

Shionone-Targeted Pneumolysin to Ameliorate Acute Lung Injury Induced by Streptococcus pneumoniae In Vivo and In Vitro
 ,
, {kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Results
2.1. Shionone Inhibits PLY Hemolytic Activity
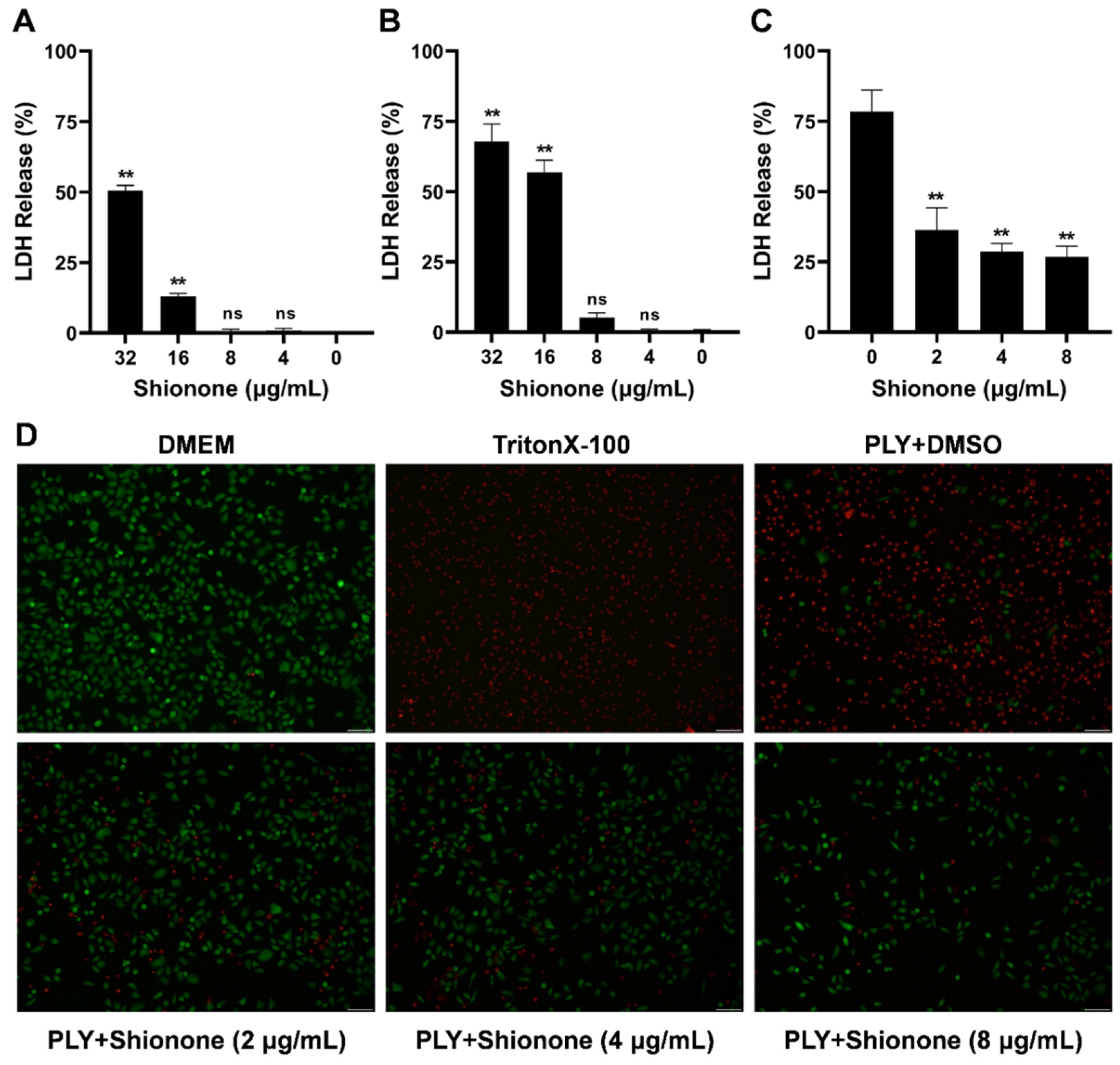
2.2. Shionone Alleviates A549 Cell Injury Induced by PLY
2.3. Shionone Neutralizes Toxicity by Inhibiting PLY Oligomerization
2.4. Shionone Alleviates S. pneumoniae Virulence in Mice
3. Discussion
4. Materials and Methods
4.1. Bacterial Strain, Cell Line, and Reagents
4.2. Hemolysis Test
4.3. Minimum Inhibitory Concentration (MIC) Determination
4.4. Growth Curve Assay
4.5. BacLight LIVE/DEAD Analysis
4.6. Cytotoxicity and Live/Dead Assays
4.7. PLY Expression Determination
4.8. Oligomerization Analysis
4.9. Molecular Docking Simulation
4.10. Mouse Pneumonia Model
4.11. Statistical Analysis
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
Sample Availability
References
- Rodrigues, C.M.C.; Groves, H. Community-Acquired Pneumonia in Children: The Challenges of Microbiological Diagnosis. J. Clin. Microbiol. 2018, 56, e01318-17. [Google Scholar] [CrossRef] [PubMed]
- Goldstein, F.W.; Vidal, B.; Kitzis, M.D. Telithromycin-resistant Streptococcus pneumoniae. Emerg. Infect. Dis. 2005, 11, 1489–1490. [Google Scholar] [CrossRef]
- Karcic, E.; Aljicevic, M.; Bektas, S.; Karcic, B. Antimicrobial Susceptibility/Resistance of Streptococcus Pneumoniae. Mater. Socio Medica 2015, 27, 180–184. [Google Scholar] [CrossRef] [PubMed]
- Casanova, C.; Küffer, M.; Leib, S.L.; Hilty, M. Re-emergence of invasive pneumococcal disease (IPD) and increase of serotype 23B after easing of COVID-19 measures, Switzerland, 2021. Emerg. Microbes Infect. 2021, 10, 2202–2204. [Google Scholar] [CrossRef] [PubMed]
- Henares, D.; Rocafort, M.; Brotons, P.; de Sevilla, M.F.; Mira, A.; Launes, C.; Cabrera-Rubio, R.; Muñoz-Almagro, C. Rapid Increase of Oral Bacteria in Nasopharyngeal Microbiota After Antibiotic Treatment in Children with Invasive Pneumococcal Disease. Front. Cell. Infect. Microbiol. 2021, 11, 744727. [Google Scholar] [CrossRef]
- Marquart, M.E. Pathogenicity and virulence of Streptococcus pneumoniae: Cutting to the chase on proteases. Virulence 2021, 12, 766–787. [Google Scholar] [CrossRef]
- Lawrence, S.L.; Feil, S.C.; Morton, C.J.; Farrand, A.J.; Mulhern, T.D.; Gorman, M.A.; Wade, K.R.; Tweten, R.K.; Parker, M.W. Crystal structure of Streptococcus pneumoniae pneumolysin provides key insights into early steps of pore formation. Sci. Rep. 2015, 5, 14352. [Google Scholar] [CrossRef]
- Li, S.F.; Lv, Q.H.; Sun, X.D.; Tang, T.Z.; Deng, X.M.; Yin, Y.H.; Li, L. Acacetin inhibits Streptococcus pneumoniae virulence by targeting pneumolysin. J. Pharm. Pharmacol. 2020, 72, 1092–1100. [Google Scholar] [CrossRef]
- Zhao, X.R.; Li, H.E.; Wang, J.F.; Guo, Y.; Liu, B.W.; Deng, X.M.; Niu, X.D. Verbascoside Alleviates Pneumococcal Pneumonia by Reducing Pneumolysin Oligomers. Mol. Pharmacol. 2016, 89, 376–387. [Google Scholar] [CrossRef]
- Li, X.; Xiang, L.; Lin, Y.; Tang, Q.; Meng, F.; Chen, W. Computational Analysis Illustrates the Mechanism of Qingfei Paidu Decoction in Blocking the Transition of COVID-19 Patients from Mild to Severe Stage. Curr. Gene Ther. 2021, 22, 277–289. [Google Scholar] [CrossRef]
- Wang, X.; Yin, H.; Fan, L.; Zhou, Y.Q.; Tang, X.L.; Fei, X.J.; Tang, H.L.; Peng, J.; Zhang, J.J.; Xue, Y.; et al. Shionone alleviates NLRP3 inflammasome mediated pyroptosis in interstitial cystitis injury. Int. Immunopharmacol. 2020, 90, 107132. [Google Scholar] [CrossRef] [PubMed]
- Xu, N.N.; Hu, J.N.; Han, K.; Ou, Y.; Ji, T.T.; Xing, J.L. Shionone suppresses the growth, migration and invasion of human breast cancer cells via induction of apoptosis and inhibition of MEK/ERK and STAT3 signalling pathways. J. BUON 2020, 25, 1821–1826. [Google Scholar] [PubMed]
- Li, H.; Zhao, X.; Wang, J.; Dong, Y.; Meng, S.; Li, R.; Niu, X.; Deng, X. β-sitosterol interacts with pneumolysin to prevent Streptococcus pneumoniae infection. Sci. Rep. 2015, 5, 17668. [Google Scholar] [CrossRef] [PubMed]
- Song, M.; Teng, Z.; Li, M.; Niu, X.; Wang, J.; Deng, X. Epigallocatechin gallate inhibits Streptococcus pneumoniae virulence by simultaneously targeting pneumolysin and sortase A. J. Cell. Mol. Med. 2017, 21, 2586–2598. [Google Scholar] [CrossRef]
- Weiser, J.N.; Ferreira, D.M.; Paton, J.C. Streptococcus pneumoniae: Transmission, colonization and invasion. Nat. Rev. Genet. 2018, 16, 355–367. [Google Scholar] [CrossRef]
- GBD 2016 Lower Respiratory Infections Collaborators. Estimates of the global, regional, and national morbidity, mortality, and aetiologies of lower respiratory infections in 195 countries, 1990–2016: A systematic analysis for the Global Burden of Disease Study 2016. Lancet Infect. Dis. 2018, 18, 1191–1210. [Google Scholar] [CrossRef]
- Wantuch, P.L.; Avci, F.Y. Invasive pneumococcal disease in relation to vaccine type serotypes. Hum. Vaccines Immunother. 2019, 15, 874–875. [Google Scholar] [CrossRef]
- Marriott, H.M.; Mitchell, T.J.; Dockrell, D.H. Pneumolysin: A Double-Edged Sword During the Host-Pathogen Interaction. Curr. Mol. Med. 2008, 8, 497–509. [Google Scholar] [CrossRef]
- Wang, T.; Zhang, P.; Lv, H.; Deng, X.; Wang, J. A Natural Dietary Flavone Myricetin as an α-Hemolysin Inhibitor for Controlling Staphylococcus aureus Infection. Front. Cell. Infect. Microbiol. 2020, 10, 330. [Google Scholar] [CrossRef]
- Rezzonico, F.; Duffy, B. Lack of genomic evidence of AI-2 receptors suggests a non-quorum sensing role for luxS in most bacteria. BMC Microbiol. 2008, 8, 154. [Google Scholar] [CrossRef] [Green Version]
- Tang, W.; Liu, J.-W.; Zhao, W.-M.; Wei, D.-Z.; Zhong, J.-J. Ganoderic acid T from Ganoderma lucidum mycelia induces mitochondria mediated apoptosis in lung cancer cells. Life Sci. 2006, 80, 205–211. [Google Scholar] [CrossRef]
- Song, Y.; Wu, Q.; Jiang, H.; Hu, A.; Xu, L.; Tan, C.; Zhang, B.; Yu, R.; Qiu, Y.; Wang, X.; et al. The Effect of Shionone on Sepsis-Induced Acute Lung Injury by the ECM1/STAT5/NF-κB Pathway. Front. Pharmacol. 2022, 12, 764247. [Google Scholar] [CrossRef] [PubMed]
- Zhang, B.; Xue, Y.; Zhao, J.; Jiang, H.; Zhu, J.; Yin, H.; Qiu, Y.; Hu, A.; Xu, L.; Song, Y.; et al. Shionone Attenuates Sepsis-Induced Acute Kidney Injury by Regulating Macrophage Polarization via the ECM1/STAT5 Pathway. Front. Med. 2022, 8, 796743. [Google Scholar] [CrossRef]
- Yin, D.-F.; Zhou, K.; Liu, J.-T.; Hu, L.; Liu, Y.; Deng, J.; Wang, S.-P.; Xiong, Y.; Zhong, W. Development and validation of an LC/MS/MS method for simultaneous determination of shionone andepi-friedelinol in rat plasma for pharmacokinetic study after oral administration of Aster tataricus extract. Biomed. Chromatogr. 2015, 30, 1112–111717. [Google Scholar] [CrossRef] [PubMed]
- Clinical and Laboratory Standards Institute. Performance Standards for Antimicrobial Susceptibility Testing, 30th ed.; CLSI supplement M100; Clinical and Laboratory Standards Institute: Wayne, PA, USA, 2020. [Google Scholar]
- Trott, O.; Olson, A.J. AutoDock Vina: Improving the speed and accuracy of docking with a new scoring function, efficient optimization, and multithreading. J. Comput. Chem. 2010, 31, 455–461. [Google Scholar] [CrossRef] [PubMed]
- O’Boyle, N.M.; Banck, M.; James, C.A.; Morley, C.; Vandermeersch, T.; Hutchison, G.R. Open babel: An open chemical toolbox. J. Cheminform. 2011, 3, 33. [Google Scholar] [CrossRef] [PubMed]
- Adasme, M.F.; Linnemann, K.L.; Bolz, S.N.; Kaiser, F.; Salentin, S.; Haupt, V.J.; Schroeder, M. PLIP 2021: Expanding the scope of the protein–ligand interaction profiler to DNA and RNA. Nucleic Acids Res. 2021, 49, W530–W534. [Google Scholar] [CrossRef]




Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Du, R.; Wang, T.; Lv, H.; Zou, Y.; Hou, X.; Hou, N.; Zhang, P.; Li, H.; Chi, G. Shionone-Targeted Pneumolysin to Ameliorate Acute Lung Injury Induced by Streptococcus pneumoniae In Vivo and In Vitro. Molecules 2022, 27, 6258. https://doi.org/10.3390/molecules27196258
Du R, Wang T, Lv H, Zou Y, Hou X, Hou N, Zhang P, Li H, Chi G. Shionone-Targeted Pneumolysin to Ameliorate Acute Lung Injury Induced by Streptococcus pneumoniae In Vivo and In Vitro. Molecules. 2022; 27(19):6258. https://doi.org/10.3390/molecules27196258
Chicago/Turabian StyleDu, Runbao, Tian Wang, Hongfa Lv, Yinuo Zou, Xiaoning Hou, Nana Hou, Peng Zhang, Hongen Li, and Gefu Chi. 2022. "Shionone-Targeted Pneumolysin to Ameliorate Acute Lung Injury Induced by Streptococcus pneumoniae In Vivo and In Vitro" Molecules 27, no. 19: 6258. https://doi.org/10.3390/molecules27196258