
Baicalin Alleviate Apoptosis via PKC-MAPK Pathway in Porcine Peritoneal Mesothelial Cells Induced by Glaesserella parasuis

Abstract
:1. Introduction
2. Results
2.1. Baicalin Inhibits GPS Induced Apoptosis
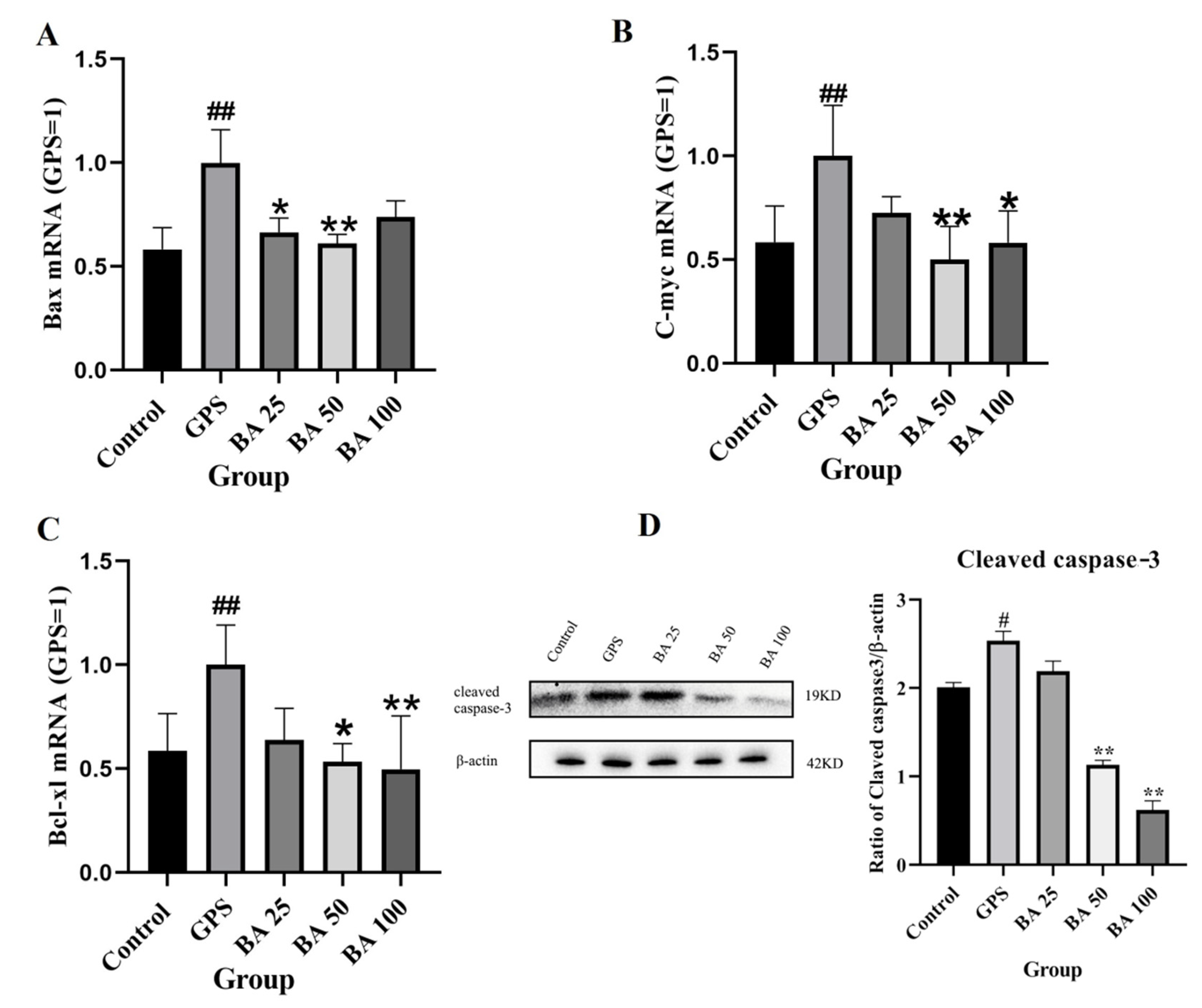
2.2. Baicalin Inhibits the Expression of Apoptosis-Related Genes Induced by GPS
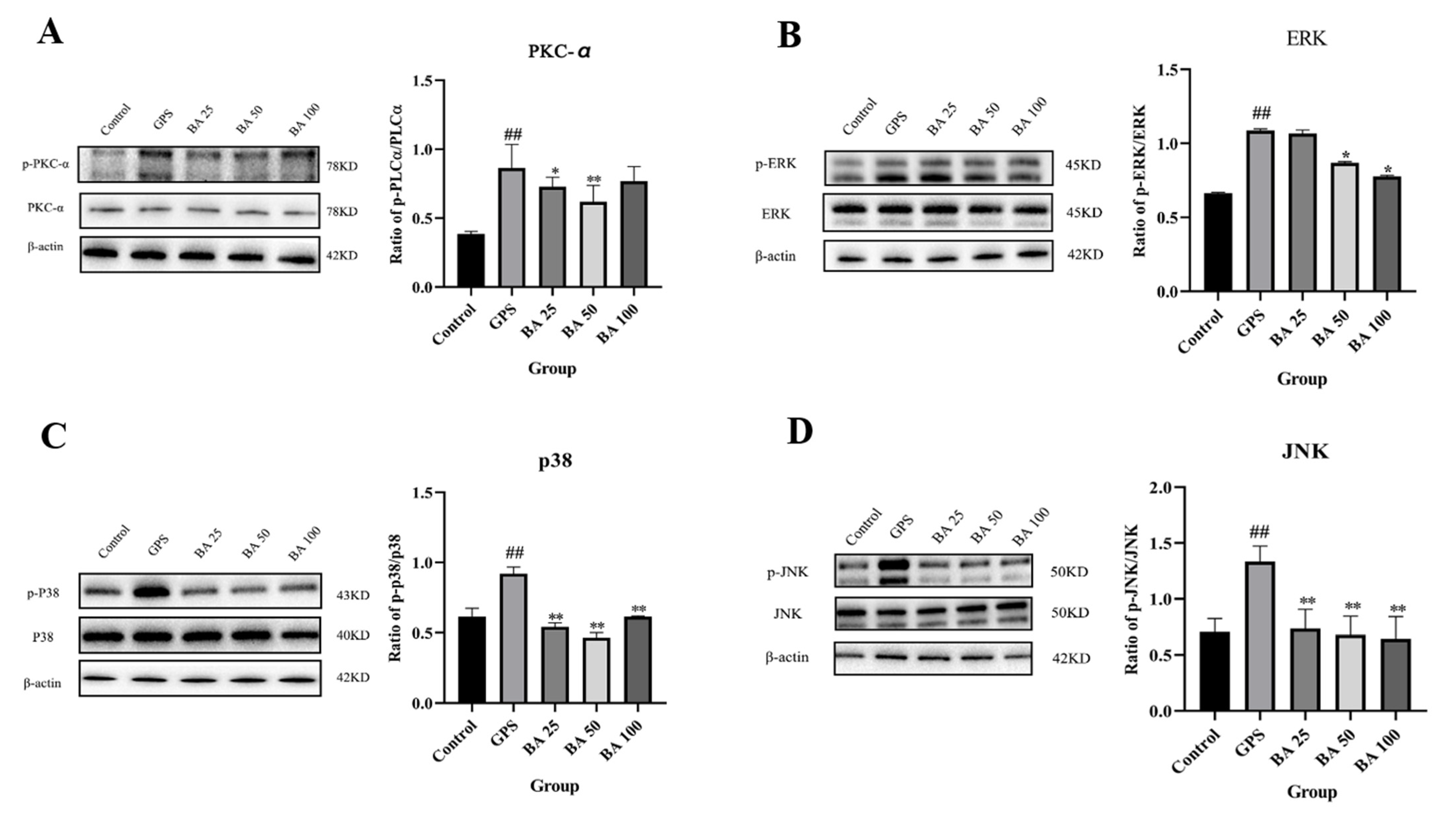
2.3. Baicalin Inhibits GPS Induced Apoptosis via PKC-MAPK Pathway
3. Discussion
4. Materials and Methods
4.1. Drugs
4.2. Bacterial Strains
4.3. Cell Culture
4.4. Cell Apoptosis
4.5. RNA Isolation and Reverse Transcription Quantitative Polymerase Chain Reaction (RT-qPCR)
4.6. Western Blot
4.7. Statistical Analysis
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Ni, H.B.; Gong, Q.L.; Zhao, Q.; Li, X.Y.; Zhang, X.X. Prevalence of Haemophilus parasuis “Glaesserella parasuis” in pigs in China: A systematic review and meta-analysis. Prev. Vet. Med. 2020, 182, 105083. [Google Scholar] [CrossRef]
- Luo, X.; Chang, X.; Zhou, H.; Lin, H.; Fan, H. Glaesserella parasuis induces inflammatory response in 3D4/21 cells through activation of NLRP3 inflammasome signaling pathway via ROS. Vet. Microbiol. 2021, 256, 109057. [Google Scholar] [CrossRef] [PubMed]
- Wang, Y.; Liu, C.; Fang, Y.; Liu, X.; Li, W.; Liu, S.; Liu, Y.; Liu, Y.; Charreyre, C.; Audonnet, J.C.; et al. Transcription analysis on response of porcine alveolar macrophages to Haemophilus parasuis. BMC Genom. 2012, 13, 68. [Google Scholar] [CrossRef] [Green Version]
- Zhang, J.; Zhang, Z.; Xu, J.; Ye, C.; Fu, S.; Hu, C.A.; Qiu, Y.; Liu, Y. Protective Effects of Baicalin on Peritoneal Tight Junctions in Piglets Challenged with Glaesserella parasuis. Molecules 2021, 26, 1268. [Google Scholar] [CrossRef] [PubMed]
- Costa-Hurtado, M.; Barba-Vidal, E.; Maldonado, J.; Aragon, V. Update on Glässer’s disease: How to control the disease under restrictive use of antimicrobials. Vet. Microbiol. 2020, 242, 108595. [Google Scholar] [CrossRef]
- An, J.; Guo, G.; Yu, D.; Zhu, K.; Zhang, C.; Li, Y. ICEHpsaHPS7, a Novel Multiple Drug Resistance Integrative Conjugative Element in Glaesserella parasuis. Antimicrob. Agents Chemother. 2021, 65, e01716-20. [Google Scholar] [CrossRef] [PubMed]
- Aarestrup, F.M.; Seyfarth, A.M.; Angen, Ø. Antimicrobial susceptibility of Haemophilus parasuis and Histophilus somni from pigs and cattle in Denmark. Vet. Microbiol. 2004, 101, 143–146. [Google Scholar] [CrossRef] [PubMed]
- McEwen, S.A.; Collignon, P.J. Antimicrobial Resistance: A One Health Perspective. Microbiol. Spectr. 2018, 6, 10. [Google Scholar] [CrossRef] [Green Version]
- Lee, C.Y.; Ooi, P.T.; Zunita, Z.; Noordin, M.M. Molecular detection of Haemophilus parasuis serotypes 4, 5 or 12 and 13 in Peninsular Malaysia. Trop. Biomed. 2019, 36, 482–487. [Google Scholar] [PubMed]
- Sanchez-Vazquez, M.J.; Nielen, M.; Edwards, S.A.; Gunn, G.J.; Lewis, F.I. Identifying associations between pig pathologies using a multi-dimensional machine learning methodology. BMC Vet. Res. 2012, 8, 151. [Google Scholar] [CrossRef] [Green Version]
- Santamaría, B.; Ucero, A.C.; Benito-Martin, A.; Vicent, M.J.; Orzáez, M.; Celdrán, A.; Selgas, R.; Ruíz-Ortega, M.; Ortiz, A. Biocompatibility reduces inflammation-induced apoptosis in mesothelial cells exposed to peritoneal dialysis fluid. Blood Purif. 2015, 39, 200–209. [Google Scholar] [CrossRef] [PubMed]
- van Till, J.W.; van Veen, S.Q.; den Broeder, V.; Bresser, P.; Lutter, R.; Out, T.A.; Schultz, M.J.; Gouma, D.J.; Boermeester, M.A. Compartmental apoptosis and neutrophil accumulation in severe peritonitis. J. Surg. Res. 2010, 164, 321–328. [Google Scholar] [CrossRef] [PubMed]
- Lim, J.H.; Park, S.M.; Yook, J.M.; Ahn, J.S.; Choi, S.Y.; Oh, S.H.; Jung, H.Y.; Choi, J.Y.; Cho, J.H.; Park, S.H.; et al. Alpha-1 antitrypsin inhibits formaldehyde-induced apoptosis of human peritoneal mesothelial cells. Perit. Dial. Int. 2020, 40, 124–131. [Google Scholar] [CrossRef] [PubMed]
- Tomita, T.; Arai, S.; Kitada, K.; Mizuno, M.; Suzuki, Y.; Sakata, F.; Nakano, D.; Hiramoto, E.; Takei, Y.; Maruyama, S.; et al. Apoptosis inhibitor of macrophage ameliorates fungus-induced peritoneal injury model in mice. Sci. Rep. 2017, 7, 6450. [Google Scholar] [CrossRef] [Green Version]
- Santamaria, B.; Ucero, A.C.; Benito-Martin, A.; Selgas, R.; Ruiz-Ortega, M.; Sanz, A.B.; Egido, J.; Ortiz, A. Taming apoptosis in peritoneal dialysis. Perit. Dial. Int. 2009, 29 (Suppl. 2), S45–S48. [Google Scholar] [CrossRef]
- Cao, S.; Li, S.; Wang, Y.; Shen, J.; Zhou, Y.; Li, H.; Yu, X.; Mao, H. Acetylation of HMGB1 by JNK1 Signaling Promotes LPS-Induced Peritoneal Mesothelial Cells Apoptosis. Biomed. Res. Int. 2018, 2018, 2649585. [Google Scholar] [CrossRef]
- Huang, T.; Liu, Y.; Zhang, C. Pharmacokinetics and Bioavailability Enhancement of Baicalin: A Review. Eur. J. Drug. Metab. Pharmacokinet. 2019, 44, 159–168. [Google Scholar] [CrossRef]
- Yu, H.; Chen, B.; Ren, Q. Baicalin relieves hypoxia-aroused H9c2 cell apoptosis by activating Nrf2/HO-1-mediated HIF1α/BNIP3 pathway. Artif. Cells. Nanomed. Biotechnol. 2019, 47, 3657–3663. [Google Scholar] [CrossRef] [Green Version]
- Guo, L.T.; Wang, S.Q.; Su, J.; Xu, L.X.; Ji, Z.Y.; Zhang, R.Y.; Zhao, Q.W.; Ma, Z.Q.; Deng, X.Y.; Ma, S.P. Baicalin ameliorates neuroinflammation-induced depressive-like behavior through inhibition of toll-like receptor 4 expression via the PI3K/AKT/FoxO1 pathway. J. Neuroinflamm. 2019, 16, 95. [Google Scholar] [CrossRef] [Green Version]
- Liu, Y.; Zhao, W.; Xu, J.; Yu, X.; Ye, C.; Fu, S.; Qiu, Y. Pharmacokinetics of sodium baicalin following intravenous and intramuscular administration to piglets. J. Vet. Pharmacol. Ther. 2019, 42, 580–584. [Google Scholar] [CrossRef]
- Ye, C.; Li, R.; Xu, L.; Qiu, Y.; Fu, S.; Liu, Y.; Wu, Z.; Hou, Y.; Hu, C.-A.A. Effects of Baicalin on piglet monocytes involving PKC–MAPK signaling pathways induced by Haemophilus parasuis. BMC Vet. Res. 2019, 15, 98. [Google Scholar] [CrossRef] [PubMed]
- Fu, S.; Yin, R.; Zuo, S.; Liu, J.; Zhang, Y.; Guo, L.; Qiu, Y.; Ye, C.; Liu, Y.; Wu, Z.; et al. The effects of baicalin on piglets challenged with Glaesserella parasuis. Vet. Res. 2020, 51, 102. [Google Scholar] [CrossRef] [PubMed]
- Fu, S.; Zhao, W.; Xiong, C.; Guo, L.; Guo, J.; Qiu, Y.; Hu, C.A.; Ye, C.; Liu, Y.; Wu, Z.; et al. Baicalin modulates apoptosis via RAGE, MAPK, and AP-1 in vascular endothelial cells during Haemophilus parasuis invasion. Innate Immun. 2019, 25, 420–432. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kaddour, H.; Hamdi, Y.; Vaudry, D.; Basille, M.; Desrues, L.; Leprince, J.; Castel, H.; Vaudry, H.; Tonon, M.C.; Amri, M.; et al. The octadecaneuropeptide ODN prevents 6-hydroxydopamine-induced apoptosis of cerebellar granule neurons through a PKC-MAPK-dependent pathway. J. Neurochem. 2013, 125, 620–633. [Google Scholar] [CrossRef] [PubMed]
- Nedbalcova, K.; Satran, P.; Jaglic, Z.; Ondriasova, R.; Kucerova, Z. Haemophilus parasuis and Glässer’s disease in pigs: A review. Vet. Med. 2018, 51, 168–179. [Google Scholar] [CrossRef] [Green Version]
- Manrique Ramírez, P.C. Identification and Partial Characterization of Acid Phosphatases from Haemophilus parasuis; Universitat Autònoma de Barcelona: Barcelona, Spain, 2014. [Google Scholar]
- Takano, T.; Hohdatsu, T.; Hashida, Y.; Kaneko, Y.; Tanabe, M.; Koyama, H. A “possible” involvement of TNF-alpha in apoptosis induction in peripheral blood lymphocytes of cats with feline infectious peritonitis. Vet. Microbiol. 2007, 119, 121–131. [Google Scholar] [CrossRef] [PubMed]
- Kolaczkowska, E.; Koziol, A.; Plytycz, B.; Arnold, B. Inflammatory macrophages, and not only neutrophils, die by apoptosis during acute peritonitis. Immunobiology 2010, 215, 492–504. [Google Scholar] [CrossRef]
- Beyer, K.; Menges, P.; Keßler, W.; Heidecke, C.D. Pathophysiology of peritonitis. Chirurgie 2016, 87, 5–12. [Google Scholar] [CrossRef]
- Feterowski, C.; Weighardt, H.; Emmanuilidis, K.; Hartung, T.; Holzmann, B. Immune protection against septic peritonitis in endotoxin-primed mice is related to reduced neutrophil apoptosis. Eur. J. Immunol. 2001, 31, 1268–1277. [Google Scholar] [CrossRef] [Green Version]
- Ameyar, M.; Wisniewska, M.; Weitzman, J.B. A role for AP-1 in apoptosis: The case for and against. Biochimie 2003, 85, 747–752. [Google Scholar] [CrossRef]
- Wang, L.; Zhang, L.; Chow, B.K.C. Secretin prevents apoptosis in the developing cerebellum through Bcl-2 and Bcl-xL. J. Mol. Neurosci. 2019, 68, 494–503. [Google Scholar] [CrossRef] [PubMed]
- Vrana, J.A.; Grant, S. Synergistic induction of apoptosis in human leukemia cells (U937) exposed to bryostatin 1 and the proteasome inhibitor lactacystin involves dysregulation of the PKC/MAPK cascade. Blood J. Am. Soc. Hematol. 2001, 97, 2105–2114. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wu, G.; Wang, Z.; Shan, P.; Huang, S.; Lin, S.; Huang, W.; Huang, Z. Suppression of Netrin-1 attenuates angiotension II-induced cardiac remodeling through the PKC/MAPK signaling pathway. Biomed. Pharmacother. 2020, 130, 110495. [Google Scholar] [CrossRef]
- Chen, Y.; Wu, Q.; Song, S.Y.; Su, W.J. Activation of JNK by TPA promotes apoptosis via PKC pathway in gastric cancer cells. World J. Gastroenterol. 2002, 8, 1014–1018. [Google Scholar] [CrossRef] [PubMed]
- An, W.W.; Wang, M.W.; Tashiro, S.; Onodera, S.; Ikejima, T. Mitogen-activated protein kinase-dependent apoptosis in norcan-tharidin-treated A375-S2 cells is proceeded by the activation of protein kinase C. Chin. Med. J. 2005, 118, 198–203. [Google Scholar] [PubMed]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Gene | Nucleotide Sequences (5’–3’) | Tm (°C) | Length (bp) | |
|---|---|---|---|---|
| β-actin | Forward | TGCGGGACATCAAGGAGAAG | 57.4 | 216 |
| Reverse | AGTTGAAGGTGGTCTCGTGG | 57.4 | ||
| Bax | Forward | GCCGAAATGTTTGCTGACG | 55.2 | 156 |
| Reverse | GAGCCGATCTCGAAGGAAGT | 57.4 | ||
| C-myc | Forward | GGTCTTCCCCTACCCACT | 57.2 | 200 |
| Reverse | CCTCATCCTCTTGTTCTTCC | 55.4 | ||
| Bcl-xl | Forward | GCAACCCATCCTGGCACCT | 59.5 | 136 |
| Reverse | TCAAACTCATCGCCCGCCT | 57.3 | ||
| c-fos | Forward | GCTGACAGATACACTCCAAGCGG | 61.3 | 542 |
| Reverse | AGGAAGACGTGTAAGTAGTGCAG | 57.8 | ||
| c-jun | Forward | CGCCAGTCTACGCTAATC | 55.1 | 288 |
| Reverse | GGTTCCTCATACGCTTCC | 54.8 | ||
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Lu, Q.; Zhou, L.; Wang, Z.; Li, X.; Ding, L.; Qiu, Y.; Guo, P.; Ye, C.; Fu, S.; Wu, Z.; et al. Baicalin Alleviate Apoptosis via PKC-MAPK Pathway in Porcine Peritoneal Mesothelial Cells Induced by Glaesserella parasuis. Molecules 2022, 27, 5083. https://doi.org/10.3390/molecules27165083
Lu Q, Zhou L, Wang Z, Li X, Ding L, Qiu Y, Guo P, Ye C, Fu S, Wu Z, et al. Baicalin Alleviate Apoptosis via PKC-MAPK Pathway in Porcine Peritoneal Mesothelial Cells Induced by Glaesserella parasuis. Molecules. 2022; 27(16):5083. https://doi.org/10.3390/molecules27165083
Chicago/Turabian StyleLu, Qirong, Lang Zhou, Ziyue Wang, Xiaomin Li, Li Ding, Yinsheng Qiu, Pu Guo, Chun Ye, Shulin Fu, Zhongyuan Wu, and et al. 2022. "Baicalin Alleviate Apoptosis via PKC-MAPK Pathway in Porcine Peritoneal Mesothelial Cells Induced by Glaesserella parasuis" Molecules 27, no. 16: 5083. https://doi.org/10.3390/molecules27165083