
Effects of High Pressure Processing and Thermal Treatment on the Interaction between α-Lactalbumin and Pelargonium-3-Glucoside


Abstract
:1. Introduction
2. Materials and Methods
2.1. Materials
2.2. Preparation of Samples
2.3. Fluorescence Measurement and Quenching Analysis
2.4. Circular Dichroism Spectrometry
2.5. Dynamic Light Scattering
2.6. Molecular Dynamic Simulation
2.7. Molecular Docking
2.8. Residue pKa Analysis
2.9. Statistical Analysis
3. Results and Discussion
3.1. Fluorescence Quenching Analysis
3.2. Protein Secondary Structure Analysis
3.3. Size Distribution Analysis
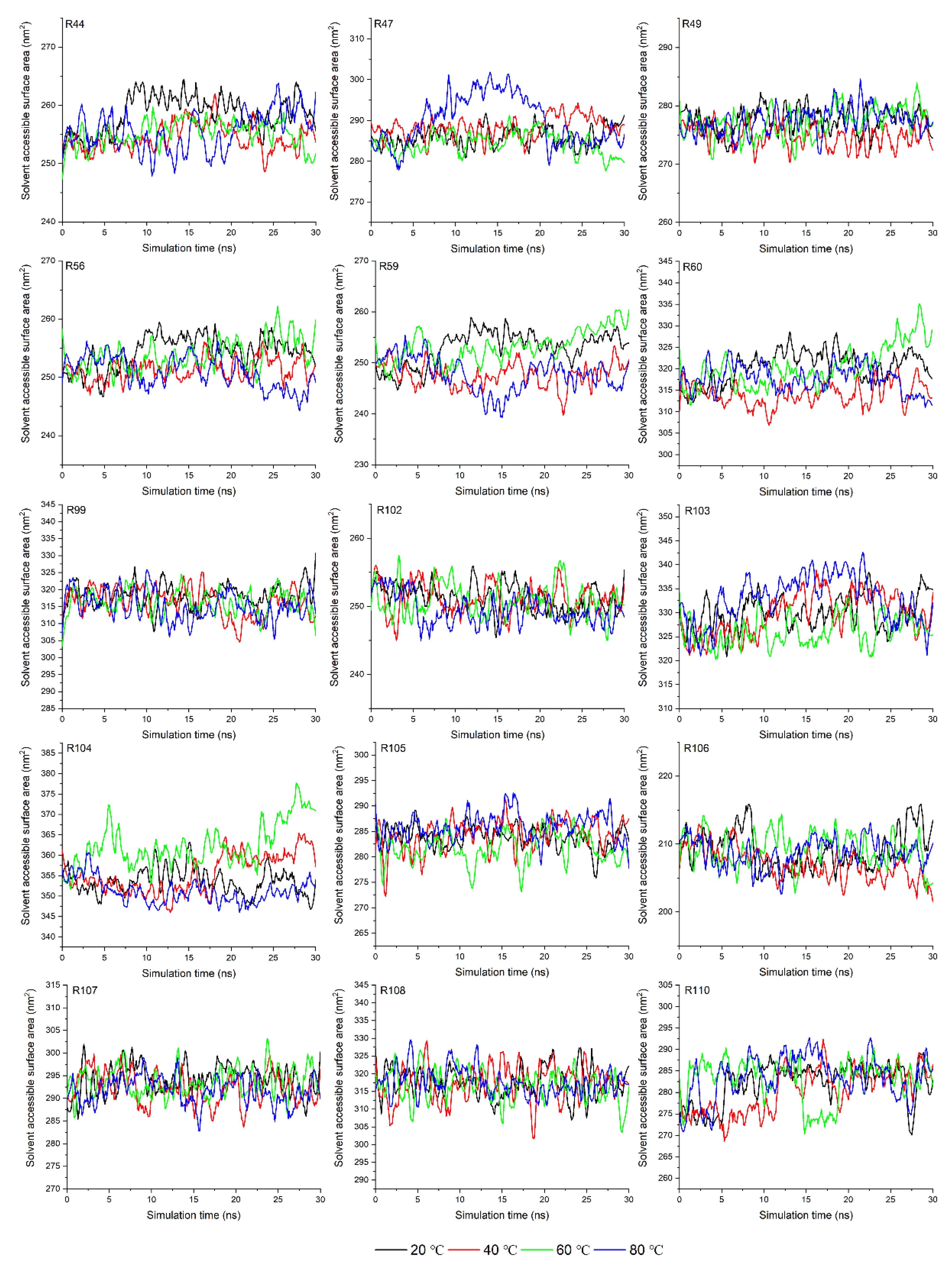
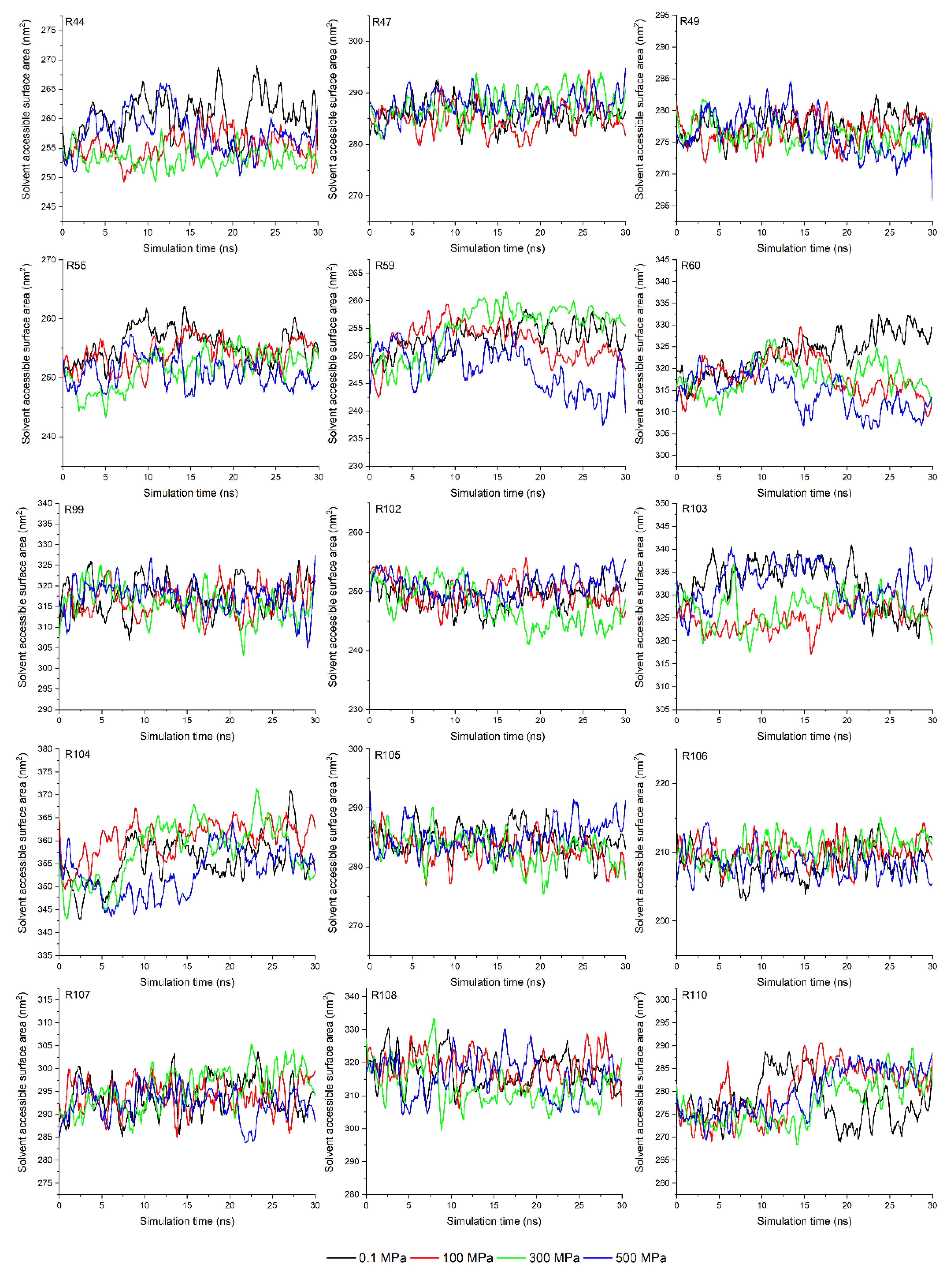
3.4. Analysis of Solvent Accessible Surface Area in Molecular Dynamic Simulation
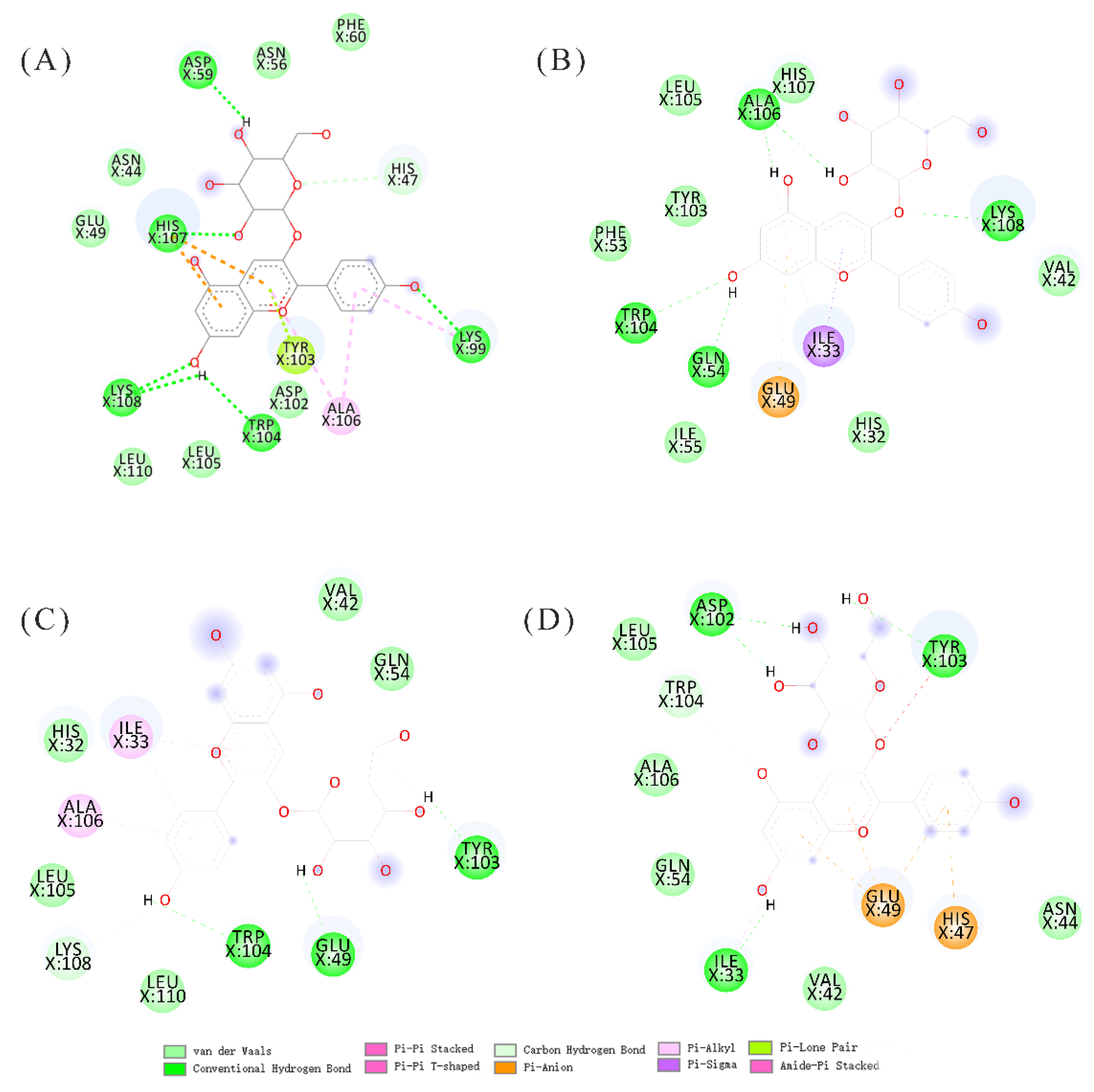
3.5. Molecular Docking Analysis
3.6. Residues pKa Analysis
4. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Mateus, N.; Freitas, V.D. Anthocyanins as Food Colorants. In Anthocyanins; Springer: Berlin, Germany, 2008. [Google Scholar]
- Alu’datt, M.H.; Rababah, T.; Alhamad, M.N.; Gammoh, S.; Ereifej, K.; Alodat, M.; Hussein, N.M.; Kubow, S.; Torley, P.J. Antioxidant and antihypertensive properties of phenolic-protein complexes in extracted protein fractions from Nigella damascena and Nigella arvensis. Food Hydrocoll. 2016, 56, 84–92. [Google Scholar] [CrossRef]
- Boye, J.I. Protein-polyphenol interactions in fruit juices. Recent Res Dev Agricult Food Chem. 1999, 3, 85–107. [Google Scholar]
- Castaneda-Ovando, A.; Pacheco-Hernandez, M.D.; Paez-Hernandez, M.E.; Rodriguez, J.A.; Galan-Vidal, C.A. Chemical studies of anthocyanins: A review. Food Chem. 2009, 113, 859–871. [Google Scholar] [CrossRef]
- Rein, M. Copigmentation Reactions and Color Stability of Berry Anthocyanins. EKT Series 1331. University of Helsinki, Department of Applied Chemistry and Microbiology, 2005, Volume 84, p. 34. Available online: https://helda.helsinki.fi/bitstream/handle/10138/20822/copigmen.pdf (accessed on 24 June 2022).
- Giusti, M.M.; Wrolstad, R.E. Acylated anthocyanins from edible sources and their applications in food systems. Biochem. Eng. J. 2003, 14, 217–225. [Google Scholar] [CrossRef]
- Yawadio, R.; Morita, N. Color enhancing effect of carboxylic acids on anthocyanins. Food Chem. 2007, 105, 421–427. [Google Scholar] [CrossRef]
- Teixeira, N.; Cruz, L.; Bras, N.F.; Mateus, N.; Ramos, M.J.; de Freitas, V. Structural Features of Copigmentation of Oenin with Different Polyphenol Copigments. J. Agric. Food Chem. 2013, 61, 6942–6948. [Google Scholar] [CrossRef]
- Li, Q.; Li, Y.W.; Rao, L.; Zhao, L.; Wang, Y.T.; Liao, X.J. Insights into pH-modulated interactions between native potato starch and cyanidin-3-O-glucoside: Electrostatic interaction-dependent binding. Food Res. Int. 2022, 156, 111129. [Google Scholar] [CrossRef] [PubMed]
- Croitoru, C.; Mureșan, C.; Turturică, M.; Stănciuc, N.; Andronoiu, D.G.; Dumitrașcu, L.; Barbu, V.; Enachi, E.; Horincar, G.; Râpeanu, G. Improvement of Quality Properties and Shelf Life Stability of New Formulated Muffins Based on Black Rice. Molecules 2018, 23, 3047. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Erna, K.H.; Felicia, W.X.L.; Vonnie, J.M.; Rovina, K.; Yin, K.W.; Nur’Aqilah, M.N. Synthesis and Physicochemical Characterization of Polymer Film-Based Anthocyanin and Starch. Biosensors 2022, 12, 211. [Google Scholar] [CrossRef] [PubMed]
- Lin, Z.; Fischer, J.; Wicker, L. Intermolecular binding of blueberry pectin-rich fractions and anthocyanin. Food Chem. 2016, 194, 986–993. [Google Scholar] [CrossRef] [Green Version]
- Zhao, L.; Pan, F.; Mehmood, A.; Zhang, H.M.; Rehman, A.U.; Li, J.Y.; Hao, S.; Wang, C.T. Improved color stability of anthocyanins in the presence of ascorbic acid with the combination of rosmarinic acid and xanthan gum. Food Chem. 2021, 351, 129317. [Google Scholar] [CrossRef]
- Liao, M.; Chen, F.; Hu, X.; Liao, X.; Miao, S.; Ma, L.; Ji, J. Controlled gastrointestinal digestion of micellar casein loaded anthocyanins: The chelating and complexing effect of dextran sulfate. Food Hydrocoll. 2022, 132, 107863. [Google Scholar] [CrossRef]
- Zang, Z.H.; Chou, S.R.; Geng, L.J.; Si, X.; Ding, Y.M.; Lang, Y.X.; Cui, H.J.; Gao, N.X.; Chen, Y.; Wang, M.S.; et al. Interactions of blueberry anthocyanins with whey protein isolate and bovine serum protein: Color stability, antioxidant activity, in vitro simulation, and protein functionality. LWT 2021, 152, 112269. [Google Scholar] [CrossRef]
- Wu, Y.; Yin, Z.; Qie, X.; Chen, Y.; Zeng, M.; Wang, Z.; Qin, F.; Chen, J.; He, Z. Interaction of Soy Protein Isolate Hydrolysates with Cyanidin-3-O-Glucoside and Its Effect on the In Vitro Antioxidant Capacity of the Complexes under Neutral Condition. Molecules 2021, 26, 1721. [Google Scholar] [CrossRef] [PubMed]
- Lowe, P.N.; Vaughan, C.K.; Daviter, T. Measurement of Protein–Ligand Complex Formation. In Protein-Ligand Interactions: Methods and Applications; Williams, M.A., Daviter, T., Eds.; Humana Press: Totowa, NJ, USA, 2013; pp. 63–99. [Google Scholar]
- Salleh, N.; Goh, K.K.T.; Waterland, M.R.; Huffman, L.M.; Weeks, M.; Matia-Merino, L. Complexation of Anthocyanin-Bound Blackcurrant Pectin and Whey Protein: Effect of pH and Heat Treatment. Molecules 2022, 27, 4202. [Google Scholar] [CrossRef] [PubMed]
- Oey, I.; Lille, M.; Van Loey, A.; Hendrickx, M. Effect of high-pressure processing on colour, texture and flavour of fruit- and vegetable-based food products: A review. Trends Food Sci. Technol. 2008, 19, 320–328. [Google Scholar] [CrossRef]
- St John, R.J.; Carpenter, J.F.; Randolph, T.W. High pressure fosters protein refolding from aggregates at high concentrations. Proc. Natl. Acad. Sci. USA 1999, 96, 13029–13033. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Katouzian, I.; Jafari, S.M.; Maghsoudlou, Y.; Karami, L.; Eikani, M.H. Experimental and molecular docking study of the binding interactions between bovine alpha-lactalbumin and oleuropein. Food Hydrocoll. 2020, 105, 105859. [Google Scholar] [CrossRef]
- Zou, H.; Xu, Z.Z.; Zhao, L.; Wang, Y.T.; Liao, X.J. Effects of high pressure processing on the interaction of alpha-lactalbumin and pelargonidin-3-glucoside. Food Chem. 2019, 285, 22–30. [Google Scholar] [CrossRef]
- Chang, R.; Liu, B.; Wang, Q.; Zhang, J.; Yuan, F.; Zhang, H.; Chen, S.; Liang, S.; Li, Y. The encapsulation of lycopene with α-lactalbumin nanotubes to enhance their anti-oxidant activity, viscosity and colloidal stability in dairy drink. Food Hydrocoll. 2022, 131, 107792. [Google Scholar] [CrossRef]
- Zou, H.; Ma, Y.; Xu, Z.Z.; Liao, X.J.; Chen, A.L.; Yang, S.M. Isolation of strawberry anthocyanins using high-speed counter-current chromatography and the copigmentation with catechin or epicatechin by high pressure processing. Food Chem. 2018, 247, 81–88. [Google Scholar] [CrossRef]
- Cao, Q.; Huang, Y.; Zhu, Q.F.; Song, M.; Xiong, S.; Manyande, A.; Du, H. The mechanism of chlorogenic acid inhibits lipid oxidation: An investigation using multi-spectroscopic methods and molecular docking. Food Chem. 2020, 333, 127528. [Google Scholar] [CrossRef] [PubMed]
- Zhou, H.L.; Wang, F.H.; Niu, H.H.; Yuan, L.; Tian, J.; Cai, S.B.; Bi, X.F.; Zhou, L.Y. Structural studies and molecular dynamic simulations of polyphenol oxidase treated by high pressure processing. Food Chem. 2022, 372, 131243. [Google Scholar] [CrossRef] [PubMed]
- Lupo, C.; Boulos, S.; Gramm, F.; Wu, X.W.; Nystrom, L. A microcalorimetric and microscopic strategy to assess the interaction between dietary fibers and small molecules. Carbohydr. Polym. 2022, 287, 119229. [Google Scholar] [CrossRef] [PubMed]
- Tian, X.; Rao, L.; Zhao, L.; Wang, Y.; Liao, X. Multispectroscopic and computational simulation insights into the inhibition mechanism of epigallocatechin-3-gallate on polyphenol oxidase. Food Chem. 2022, 393, 133415. [Google Scholar] [CrossRef] [PubMed]
- Morris, G.M.; Huey, R.; Lindstrom, W.; Sanner, M.F.; Belew, R.K.; Goodsell, D.S.; Olson, A.J. AutoDock4 and AutoDockTools4: Automated Docking with Selective Receptor Flexibility. J. Comput. Chem. 2009, 30, 2785–2791. [Google Scholar] [CrossRef] [Green Version]
- Sarkar, T.; Bharadwaj, K.K.; Salauddin, M.; Pati, S.; Chakraborty, R. Phytochemical Characterization, Antioxidant, Anti-inflammatory, Anti-diabetic properties, Molecular Docking, Pharmacokinetic Profiling, and Network Pharmacology Analysis of the Major Phytoconstituents of Raw and Differently Dried Mangifera indica (Himsagar cultivar): An In Vitro and In Silico Investigations. Appl. Biochem. Biotechnol. 2022, 194, 950–987. [Google Scholar] [CrossRef]
- Olsson, M.H.M.; Søndergaard, C.R.; Rostkowski, M.; Jensen, J.H. PROPKA3: Consistent Treatment of Internal and Surface Residues in Empirical pKa Predictions. J. Chem. Theory Comput. 2011, 7, 525–537. [Google Scholar] [CrossRef] [PubMed]
- Lakowicz, J.R. Principles of Fluorescence Spectroscopy; Springer: Berlin, Germany, 2006. [Google Scholar]
- Nishizawa, M.; Walinda, E.; Morimoto, D.; Kohn, B.; Scheler, U.; Shirakawa, M.; Sugase, K. Effects of Weak Nonspecific Interactions with ATP on Proteins. J. Am. Chem. Soc. 2021, 143, 11982–11993. [Google Scholar] [CrossRef] [PubMed]
- Ma, J.; Yao, Q.; Chen, X.; Lv, C.; Zang, J.; Zhao, G. Weak Binding of Epigallocatechin to α-Lactalbumin Greatly Improves Its Stability and Uptake by Caco-2 Cells. J. Agric. Food Chem. 2021, 69, 8482–8491. [Google Scholar] [CrossRef]
- Li, Y.; Wan, Y.; Mamu, Y.; Liu, X.; Guo, S. Protein aggregation and Ca2+-induced gelation of soymilk after heat treatment under slightly alkaline conditions. Food Hydrocoll. 2022, 124, 107274. [Google Scholar] [CrossRef]
- Singh, A.; Vanga, S.K.; Orsat, V.; Raghavan, V. Application of molecular dynamic simulation to study food proteins: A review. Crit. Rev. Food Sci. 2018, 58, 2779–2789. [Google Scholar] [CrossRef]
- Durham, E.; Dorr, B.; Woetzel, N.; Staritzbichler, R.; Meiler, J. Solvent accessible surface area approximations for rapid and accurate protein structure prediction. J. Mol. Model. 2009, 15, 1093–1108. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tian, X.; Liu, Y.; Zhao, L.; Rao, L.; Wang, Y.; Liao, X. Inhibition effect of high hydrostatic pressure combined with epigallocatechin gallate treatments on pectin methylesterase in orange juice and model system. Food Chem. 2022, 390, 133147. [Google Scholar] [CrossRef]
- Chen, G.; Huang, K.; Miao, M.; Feng, B.; Campanella, O.H. Molecular Dynamics Simulation for Mechanism Elucidation of Food Processing and Safety: State of the Art. Compr. Rev. Food Sci. Food 2019, 18, 243–263. [Google Scholar] [CrossRef] [Green Version]
- Le Bourvellec, C.; Renard, C.M.G.C. Interactions between Polyphenols and Macromolecules: Quantification Methods and Mechanisms. Crit. Rev. Food Sci. 2012, 52, 213–248. [Google Scholar] [CrossRef]
- Ulrih, N.P. Analytical techniques for the study of polyphenol-protein interactions. Crit. Rev. Food Sci. 2017, 57, 2144–2161. [Google Scholar] [CrossRef] [PubMed]
- Ozdal, T.; Capanoglu, E.; Altay, F. A review on protein-phenolic interactions and associated changes. Food Res. Int. 2013, 51, 954–970. [Google Scholar] [CrossRef]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| pH | Control | 40 °C | 60 °C | 80 °C | |
|---|---|---|---|---|---|
| Ka | 6.0 | 0.498 ± 0.030 a | 0.388 ± 0.078 a | 0.495 ± 0.084 a | 0.542 ± 0.177 a |
| 7.4 | 0.463 ± 0.042 a | 0.448 ± 0.092 ab | 0.304 ± 0.084 ab | 0.442 ± 0.075 b | |
| 8.0 | 0.392 ± 0.084 a | 0.462 ± 0.092 ab | 0.526 ± 0.015 bc | 0.633 ± 0.001 c | |
| fa | 6.0 | 0.386 ± 0.194 a | 0.565 ± 0.237 a | 0.331 ± 0.054 a | 0.714 ± 0.371 a |
| 7.4 | 0.417 ± 0.048 a | 0.515 ± 0.125 a | 0.464 ± 0.245 a | 0.473 ± 0.293 a | |
| 8.0 | 0.428 ± 0.244 a | 0.732 ± 0.272 ab | 0.563 ± 0.232 ab | 0.768 ± 0.196 b |
| pH | Content (%) | Control | 100 MPa | 300 MPa | 500 MPa | 40 °C | 60 °C | 80 °C |
|---|---|---|---|---|---|---|---|---|
| 6.0 | α-helix | 37.60 | 37.80 | 38.70 | 44.20 | 45.60 | 30.10 | 44.30 |
| β-turn | 17.00 | 17.10 | 17.00 | 18.00 | 16.50 | 19.20 | 16.40 | |
| random coil | 22.50 | 22.30 | 21.20 | 10.80 | 14.40 | 23.40 | 16.20 | |
| 7.4 | α-helix | 38.20 | 40.70 | 40.70 | 40.50 | 43.90 | 32.30 | 47.90 |
| β-turn | 18.50 | 17.90 | 17.90 | 17.90 | 16.10 | 18.60 | 15.80 | |
| random coil | 14.40 | 14.30 | 14.50 | 14.80 | 18.10 | 22.80 | 14.80 | |
| 8.0 | α-helix | 35.00 | 38.60 | 39.60 | 41.30 | 43.40 | 42.80 | 41.30 |
| β-turn | 17.90 | 17.30 | 17.70 | 17.80 | 16.10 | 16.60 | 17.10 | |
| random coil | 21.60 | 19.50 | 16.40 | 14.10 | 18.70 | 17.10 | 16.90 |
| Treatments | Peak Intensity (nm) | Peak Intensity Percentage (%) | |||||
|---|---|---|---|---|---|---|---|
| Peak I | Peak II | Peak III | Peak I | Peak II | Peak III | ||
| High pressure processing | Control/pH 6.0 | 229.40 | 3.95 | 0 | 56.53 | 43.46 | 0 |
| Control/pH 7.4 | 220.23 | 3.77 | 0 | 55.50 | 44.50 | 0 | |
| Control/pH 8.0 | 191.54 | 3.83 | 0 | 57.06 | 42.94 | 0 | |
| 100 MPa/pH 6.0 | 189.56 | 4.06 | 39.78 | 51.43 | 30.93 | 17.66 | |
| 100 MPa/pH 7.4 | 178.32 | 3.80 | 23.40 | 55.90 | 29.38 | 14.74 | |
| 100 MPa/pH 8.0 | 136.03 | 3.55 | 13.73 | 61.13 | 25.20 | 13.67 | |
| 300 MPa/pH 6.0 | 247.53 | 4.09 | 49.99 | 49.63 | 31.16 | 19.20 | |
| 300 MPa/pH 7.4 | 297.65 | 4.00 | 46.87 | 47.70 | 31.83 | 19.32 | |
| 300 MPa/pH 8.0 | 126.25 | 3.85 | 17.17 | 54.80 | 34.95 | 10.20 | |
| 500 MPa/pH 6.0 | 187.32 | 3.97 | 33.37 | 54.14 | 35.98 | 9.92 | |
| 500 MPa/pH 7.4 | 225.13 | 10.30 | 28.84 | 54.65 | 26.23 | 19.10 | |
| 500 MPa/pH 8.0 | 261.16 | 3.89 | 30.44 | 59.80 | 25.50 | 14.70 | |
| Thermal treatment | 40 °C/pH 6.0 | 197.26 | 3.76 | 0 | 45.63 | 54.36 | 0 |
| 40 °C/pH 7.4 | 146.85 | 4.17 | 0 | 58.90 | 41.10 | 0 | |
| 40 °C/pH 8.0 | 140.10 | 3.67 | 0 | 47.05 | 52.95 | 0 | |
| 60 °C/pH 6.0 | 323.76 | 4.01 | 0 | 56.22 | 43.78 | 0 | |
| 60 °C/pH 7.4 | 320.95 | 3.97 | 0 | 59.32 | 40.68 | 0 | |
| 60 °C/pH 8.0 | 265.17 | 3.70 | 0 | 63.05 | 36.95 | 0 | |
| 80 °C/pH 6.0 | 188.40 | 3.73 | 0 | 61.00 | 39.00 | 0 | |
| 80 °C/pH 7.4 | 262.01 | 3.98 | 0 | 45.83 | 54.18 | 0 | |
| 80 °C/pH 8.0 | 870.6 | 0 | 0 | 100.00 | 0 | 0 | |
| Histidine Number | Control | 40 °C | 60 °C | 80 °C |
|---|---|---|---|---|
| 10 | 6.51 | 6.42 | 6.45 | 6.53 |
| 32 | 6.50 | 6.66 | 6.78 | 6.86 |
| 47 | 7.24 | 6.32 | 6.80 | 7.47 |
| 107 | 6.60 | 6.39 | 6.55 | 6.34 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Tian, X.; Zou, H.; Yang, P.; Ma, Y.; Li, Y.; Zhao, L.; Wang, Y.; Liao, X. Effects of High Pressure Processing and Thermal Treatment on the Interaction between α-Lactalbumin and Pelargonium-3-Glucoside. Molecules 2022, 27, 4944. https://doi.org/10.3390/molecules27154944
Tian X, Zou H, Yang P, Ma Y, Li Y, Zhao L, Wang Y, Liao X. Effects of High Pressure Processing and Thermal Treatment on the Interaction between α-Lactalbumin and Pelargonium-3-Glucoside. Molecules. 2022; 27(15):4944. https://doi.org/10.3390/molecules27154944
Chicago/Turabian StyleTian, Xuezhi, Hui Zou, Peiqing Yang, Yan Ma, Yuwan Li, Liang Zhao, Yongtao Wang, and Xiaojun Liao. 2022. "Effects of High Pressure Processing and Thermal Treatment on the Interaction between α-Lactalbumin and Pelargonium-3-Glucoside" Molecules 27, no. 15: 4944. https://doi.org/10.3390/molecules27154944