
The Transcriptomic Mechanism of a Novel Autolysis Induced by a Recombinant Antibacterial Peptide from Chicken Expressed in Pichia pastoris


Abstract
:1. Introduction
2. Materials and Methods
2.1. Strain and Culture Conditions
2.2. Plasmids
2.3. Construction of Plasmids Expressing the AMP and ADTZ Genes
2.4. Transformation of P. pastoris
2.5. Induction of the Recombinant AMP and ADTZ in P. pastoris at Bioreactor Scale
2.6. Sample Collection and Pretreatment
2.7. RNA Isolation, Quantification and Qualification
2.8. Library Preparation for Transcriptome Sequencing
2.9. Clustering and Sequencing
2.10. RNA-Seq Data Analysis
2.11. Quantitative Real-Time Reverse Transcription PCR (qRT-PCR)
3. Results and Discussion
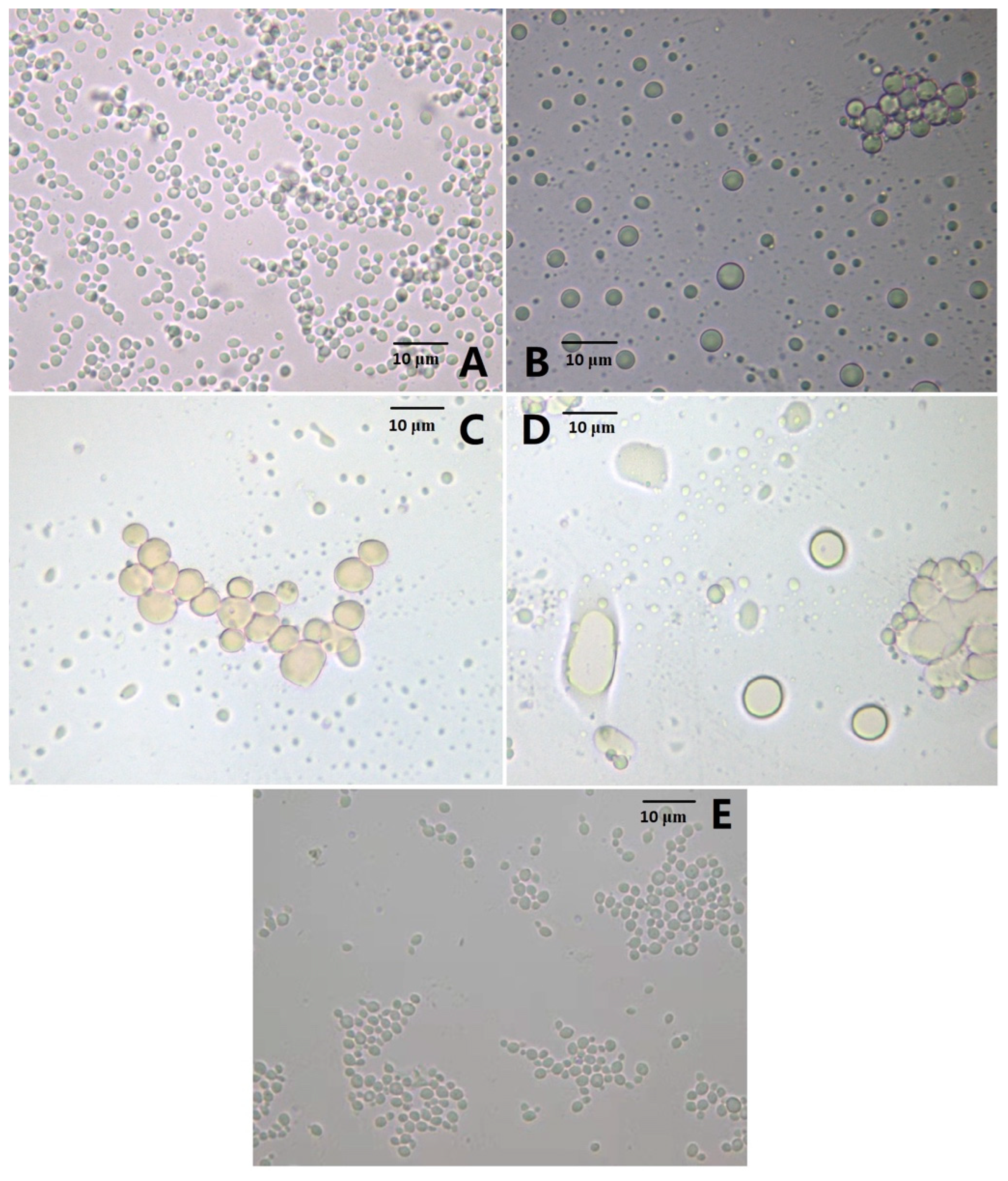
3.1. Effect of the AMP Gene Expression on Cell Morphology
3.2. Sequencing and De Novo Assembly of P. pastoris Transcriptome
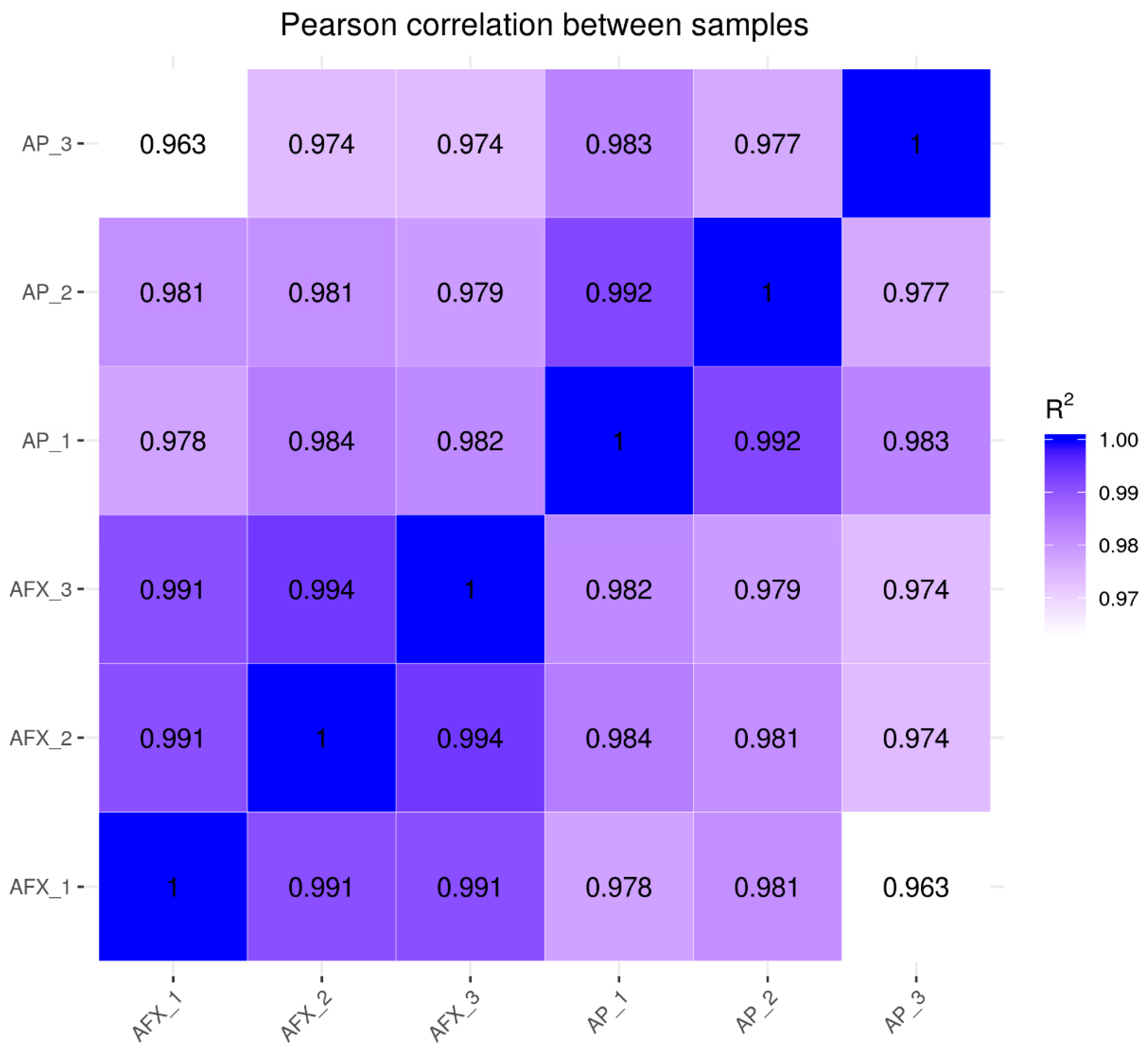
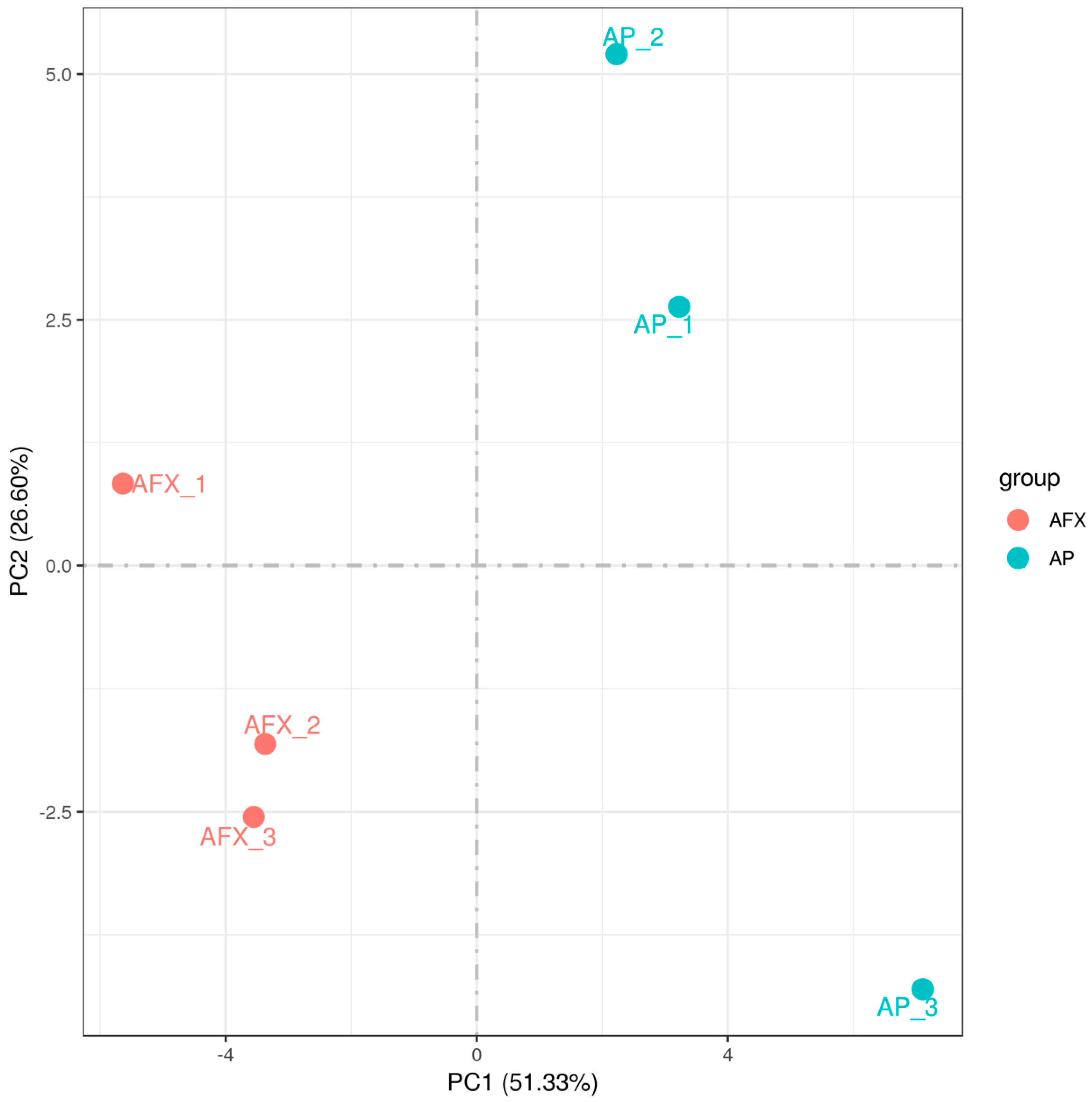
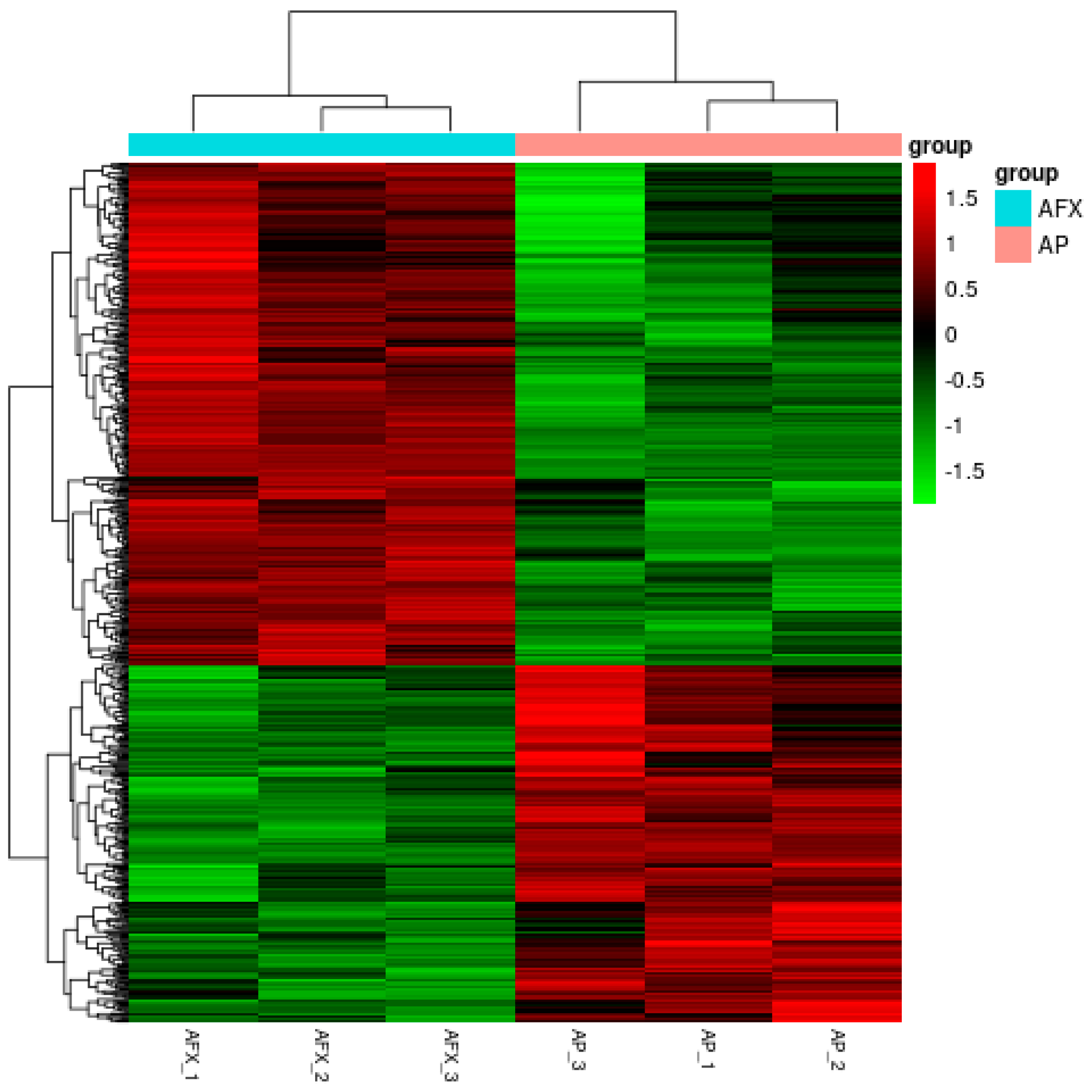
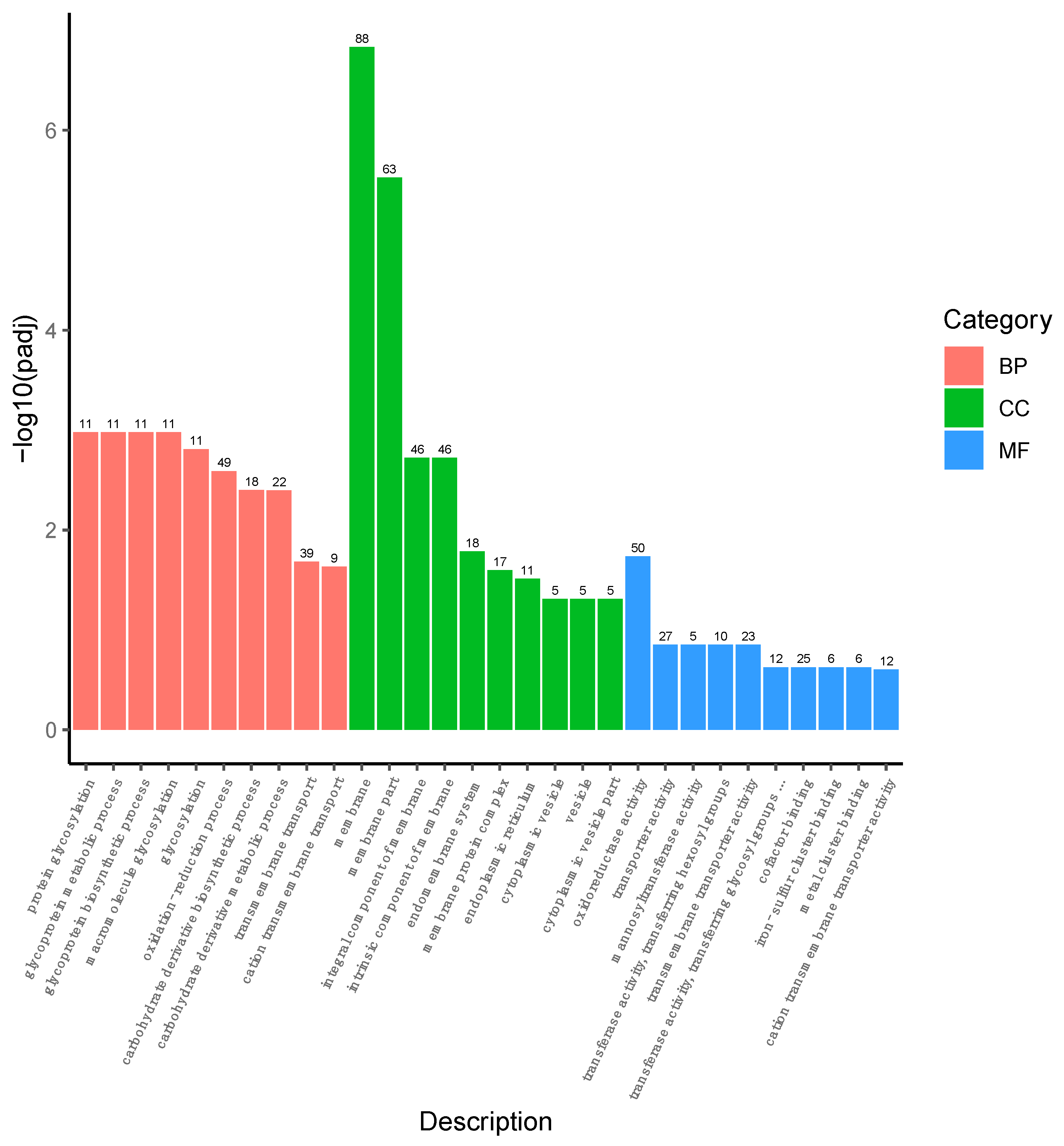
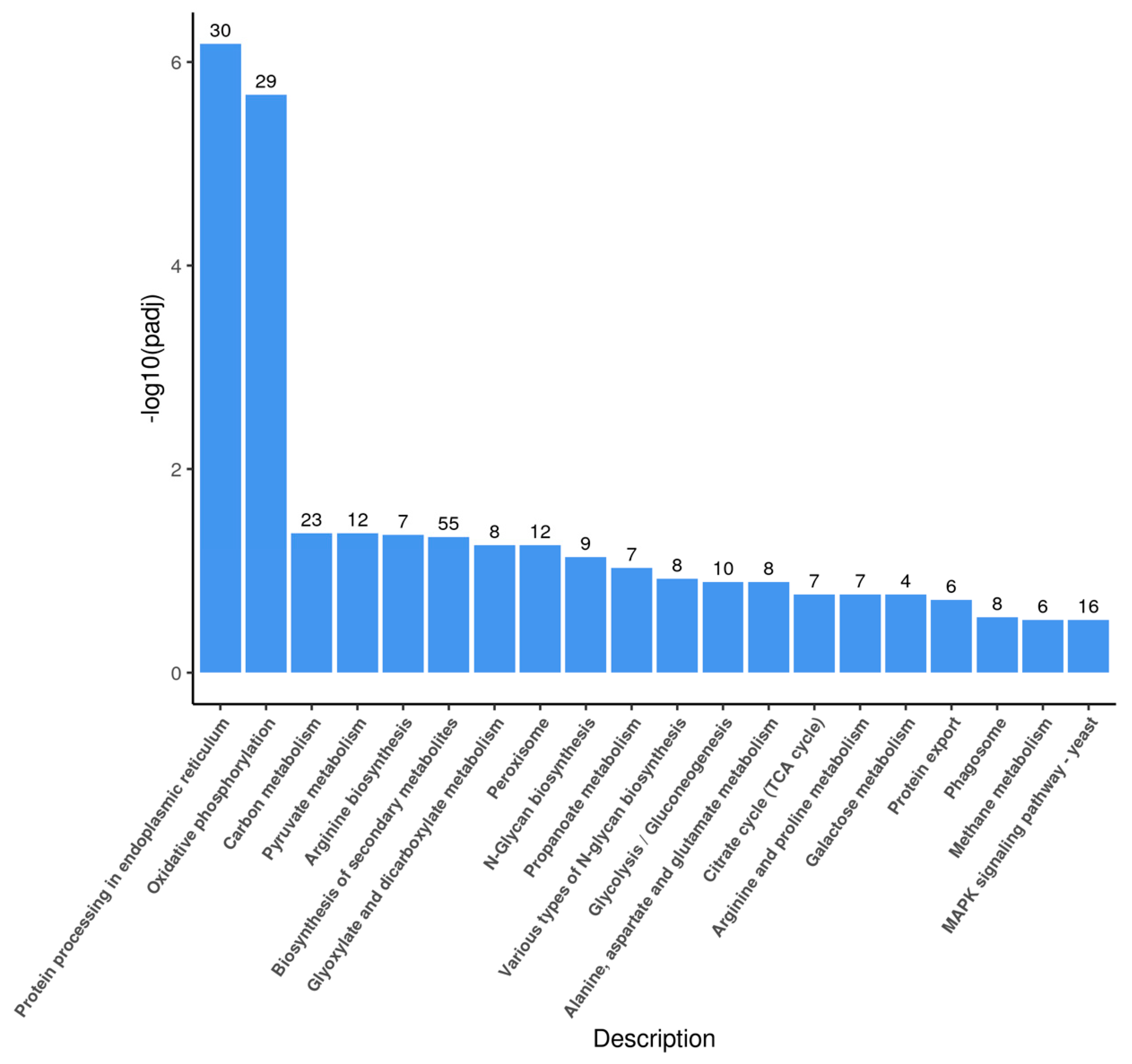
3.3. Changes in Transcriptional Profile Induced by the Recombinant AMP in P. pastoris
3.4. Overview of the DEGs
3.5. DEGs Related to the Autolysis in P. pastoris
4. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Conflicts of Interest
Sample Availability
References
- Zasloff, M. Antimicrobial peptides of multicellular organisms. Nature 2002, 415, 389–395. [Google Scholar] [CrossRef] [PubMed]
- Wan, J.; Li, Y.; Chen, D.; Yu, B.; Zheng, P.; Mao, X.; Yu, J.; He, J. Expression of a Tandemly Arrayed Plectasin Gene from Pseudoplectania nigrella in Pichia pastoris and its Antimicrobial Activity. J. Microbiol. Biotechnol. 2016, 26, 461–468. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Meng, D.-M.; Zhao, J.-F.; Ling, X.; Dai, H.-X.; Guo, Y.-J.; Gao, X.-F.; Dong, B.; Zhang, Z.-Q.; Meng, X.; Fan, Z.-C. Recombinant expression, purification and antimicrobial activity of a novel antimicrobial peptide PaDef in Pichia pastoris. Protein Expr. Purif. 2017, 130, 90–99. [Google Scholar] [CrossRef] [PubMed]
- Fujiki, Y.; Kumada, Y.; Kishimoto, M. Phase analysis in single-chain variable fragment production by recombinant Pichia pastoris based on proteomics combined with multivariate statistics. J. Biosci. Bioeng. 2015, 120, 187–192. [Google Scholar] [CrossRef]
- Garcia-Ortega, X.; Reyes, C.; Luis Montesinos, J.; Valero, F. Overall key performance indicator to optimizing operation of high-pressure homogenizers for a reliable quantification of intracellular components in Pichia pastoris. Front. Bioeng. Biotechnol. 2015, 3, 107. [Google Scholar] [CrossRef] [Green Version]
- Bartolo-Aguilar, Y.; Dendooven, L.; Chavez-Cabrera, C.; Flores-Cotera, L.B.; Hidalgo-Lara, M.E.; Villa-Tanaca, L.; Marsch, R. Autolysis of Pichia pastoris induced by cold. AMB Express 2017, 7, 95. [Google Scholar] [CrossRef] [Green Version]
- Xing, Y.; Chen, C.; Sun, W.; Zhang, B.; Sang, Y.; Xiu, Z.; Dong, Y. An environment-friendly approach to isolate and purify glucan from spent cells of recombinant Pichia pastoris and the bioactivity characterization of the purified glucan. Eng. Life Sci. 2018, 18, 227–235. [Google Scholar] [CrossRef] [Green Version]
- Lo, S.-C.; Yang, C.-Y.; Mathew, D.C.; Huang, C.-C. Growth and autolysis of the kefir yeast Kluyveromyces marxianus in lactate culture. Sci. Rep. 2021, 11, 14552. [Google Scholar] [CrossRef]
- Li, X.; Ye, H.; Xu, C.-Q.; Shen, X.-L.; Zhang, X.-L.; Huang, C.; Cheng, B.; Tan, Y.-L.; Xiao, Z.-T.; Pei, Y.-P.; et al. Transcriptomic analysis reveals MAPK signaling pathways affect the autolysis in baker’s yeast. FEMS Yeast Res. 2020, 20, foaa036. [Google Scholar] [CrossRef]
- Xu, W.; Wang, J.; Li, Q. Comparative proteome and transcriptome analysis of lager brewer’s yeast in the autolysis process. FEMS Yeast Res. 2014, 14, 1273–1285. [Google Scholar] [CrossRef] [Green Version]
- Cebollero, E.; Gonzalez, R. Induction of autophagy by second-fermentation yeasts during elaboration of sparkling wines. Appl. Environ. Microbiol. 2006, 72, 4121–4127. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Klionsky, D.J.; Cregg, J.M.; Dunn, W.A.; Emr, S.D.; Sakai, Y.; Sandoval, I.V.; Sibirny, A.; Subramani, S.; Thumm, M.; Veenhuis, M.; et al. A unified nomenclature for yeast autophagy-related genes. Dev. Cell 2003, 5, 539–545. [Google Scholar] [CrossRef] [Green Version]
- He, C.; Klionsky, D.J. Regulation Mechanisms and Signaling Pathways of Autophagy. Annu. Rev. Genet. 2009, 43, 67–93. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bernard, A.; Jin, M.; Xu, Z.; Klionsky, D.J. A large-scale analysis of autophagy-related gene expression identifies new regulators of autophagy. Autophagy 2015, 11, 2114–2122. [Google Scholar] [CrossRef]
- Cebollero, E.; Gonzalez, R. Autophagy: From basic research to its application in food biotechnology. Biotechnol. Adv. 2007, 25, 396–409. [Google Scholar] [CrossRef]
- Juturu, V.; Wu, J.C. Heterologous Protein Expression in Pichia pastoris: Latest Research Progress and Applications. Chembiochem 2018, 19, 7–21. [Google Scholar] [CrossRef]
- Zou, C.; Wang, P.; Liang, S.; Lin, Y. Deletion of Gcw13 represses autophagy in Pichia pastoris cells grown in methanol medium with sufficient amino acids. Biotechnol. Lett. 2019, 41, 1423–1431. [Google Scholar] [CrossRef]
- Vanz, A.L.; Luensdorf, H.; Adnan, A.; Nimtz, M.; Gurramkonda, C.; Khanna, N.; Rinas, U. Physiological response of Pichia pastoris GS115 to methanol-induced high level production of the Hepatitis B surface antigen: Catabolic adaptation, stress responses, and autophagic processes. Microb. Cell Fact. 2012, 11, 103. [Google Scholar] [CrossRef] [Green Version]
- Zhang, C.B.; Ma, Y.; Miao, H.B.; Tang, X.H.; Xu, B.; Wu, Q.; Mu, Y.L.; Huang, Z.X. Transcriptomic Analysis of Pichia pastoris (Komagataella phaffii) GS115 During Heterologous Protein Production Using a High-Cell-Density Fed-Batch Cultivation Strategy. Front. Microbiol. 2020, 11, 463. [Google Scholar] [CrossRef]
- Lynn, D.J.; Higgs, R.; Gaines, S.; Tierney, J.; James, T.; Lloyd, A.T.; Fares, M.A.; Mulcahy, G.; O’Farrelly, C. Bioinformatic discovery and initial characterisation of nine novel antimicrobial peptide genes in the chicken. Immunogenetics 2004, 56, 170–177. [Google Scholar] [CrossRef] [Green Version]
- Yang, H.; Li, S.; Li, F.; Yu, K.; Yang, F.; Xiang, J. Recombinant Expression of a Modified Shrimp Anti-Lipopolysaccharide Factor Gene in Pichia pastoris GS115 and Its Characteristic Analysis. Mar. Drugs 2016, 14, 152. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yang, X.; Niu, L.; Pan, Y.P.; Feng, X.H.; Liu, J.; Guo, Y.; Pan, C.L.; Geng, F.X.; Tang, X.L. LL-37-Induced Autophagy Contributed to the Elimination of Live Porphyromonas gingivalis Internalized in Keratinocytes. Front. Cell. Infect. Microbiol. 2020, 10, 561761. [Google Scholar] [CrossRef]
- Chen, K.; Yoshimura, T.; Gong, W.; Tian, C.; Huang, J.; Trinchieri, G.; Wang, J.M. Requirement of CRAMP for mouse macrophages to eliminate phagocytosed E. coli through an autophagy pathway. J. Cell Sci. 2021, 134, jcs252148. [Google Scholar] [CrossRef] [PubMed]
- Yang, R.; Yang, E.; Shen, L.; Modlin, R.L.; Shen, H.; Chen, Z.W. IL-12+IL-18 Cosignaling in Human Macrophages and Lung Epithelial Cells Activates Cathelicidin and Autophagy, Inhibiting Intracellular Mycobacterial Growth. J. Immunol. 2018, 200, 2405–2417. [Google Scholar] [CrossRef] [Green Version]
- Martinez-Culebras, P.V.; Gandia, M.; Garrigues, S.; Marcos, J.F.; Manzanares, P. Antifungal Peptides and Proteins to Control Toxigenic Fungi and Mycotoxin Biosynthesis. Int. J. Mol. Sci. 2021, 22, 13261. [Google Scholar] [CrossRef] [PubMed]
- White, S.; McIntyre, M.; Berry, D.R.; McNeil, B. The autolysis of industrial filamentous fungi. Crit. Rev. Biotechnol. 2002, 22, 1–14. [Google Scholar] [CrossRef]
- Amrane, A.; Prigent, Y. Behaviour of the yeast Kluyveromyces marxianus var marxianus during its autolysis. Antonie Van Leeuwenhoek 1996, 69, 267–272. [Google Scholar] [CrossRef]
- Farkas, V.; Takeo, K.; Macekova, D.; Ohkusu, M.; Yoshida, S.; Sipiczki, M. Secondary cell wall formation in Cryptococcus neoformans as a rescue mechanism against acid-induced autolysis. Fems Yeast Res. 2009, 9, 311–320. [Google Scholar] [CrossRef] [Green Version]
- Wibowo, D.; Zhao, C.X. Recent achievements and perspectives for large-scale recombinant production of antimicrobial peptides. Appl. Microbiol. Biotechnol. 2019, 103, 659–671. [Google Scholar] [CrossRef] [Green Version]
- Wang, X.L.; Lu, D.X.; Tian, C.M. Mitogen-activated protein kinase cascade CgSte50-Ste11-Ste7-Mk1 regulates infection-related morphogenesis in the poplar anthracnose fungus Colletotrichum gloeosporioides. Microbiol. Res. 2021, 248, 126748. [Google Scholar] [CrossRef]
- Lei, Y.; Liu, G.; Li, Z.; Gao, L.; Qin, Y.; Qu, Y. Functional characterization of protein kinase CK2 regulatory subunits regulating Penicillium oxalicum asexual development and hydrolytic enzyme production. Fungal Genet. Biol. 2014, 66, 44–53. [Google Scholar] [CrossRef] [PubMed]
- Huang, W.; Shang, Y.; Chen, P.; Cen, K.; Wang, C. Basic Leucine Zipper (bZIP) Domain Transcription Factor MBZ1 Regulates Cell Wall Integrity, Spore Adherence, and Virulence in Metarhizium robertsii. J. Biol. Chem. 2015, 290, 8218–8231. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Etxebeste, O.; Herrero-Garcia, E.; Araujo-Bazan, L.; Rodriguez-Urra, A.B.; Garzia, A.; Ugalde, U.; Espeso, E.A. The bZIP-type transcription factor FlbB regulates distinct morphogenetic stages of colony formation in Aspergillus nidulans. Mol. Microbiol. 2009, 73, 775–789. [Google Scholar] [CrossRef] [Green Version]
- Khatami, M.; Hosseini, S.N.; Hasannia, S. Co-expression of alpha-1 antitrypsin with cytoplasmic domain of v-SNARE in Pichia pastoris: Preserving biological activity of alpha-1 antitrypsin. Biotechnol. Appl. Biochem. 2018, 65, 181–187. [Google Scholar] [CrossRef]
- Wang, Y.X.; Kauffman, E.J.; Duex, J.E.; Weisman, L.S. Fusion of docked membranes requires the armadillo repeat protein Vac8p. J. Biol. Chem. 2001, 276, 35133–35140. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Marques, M.; Mojzita, D.; Amorim, M.A.; Almeida, T.; Hohmann, S.; Moradas-Ferreira, P.; Costa, V. The Pep4p vacuolar proteinase contributes to the turnover of oxidized proteins but PEP4 overexpression is not sufficient to increase chronological lifespan in Saccharomyces cerevisiae. Microbiology 2006, 152, 3595–3605. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Li, Y.; Tran, Q.; Shrestha, R.; Piao, L.; Park, S.; Park, J.; Park, J. LETM1 is required for mitochondrial homeostasis and cellular viability. Mol. Med. Rep. 2019, 19, 3367–3375. [Google Scholar] [CrossRef]
- Che, N.; Yang, Z.; Liu, X.; Li, M.; Feng, Y.; Zhang, C.; Li, C.; Cui, Y.; Xuan, Y. Suppression of LETM1 inhibits the proliferation and stemness of colorectal cancer cells through reactive oxygen species-induced autophagy. J. Cell. Mol. Med. 2021, 25, 2110–2120. [Google Scholar] [CrossRef]
- Li, T.; Ma, J.; Xu, Z.; Wang, S.; Wang, N.; Shao, S.; Yang, W.; Huang, L.; Liu, Y. Transcriptomic Analysis of the Influence of Methanol Assimilation on the Gene Expression in the Recombinant Pichia pastoris Producing Hirudin Variant 3. Genes 2019, 10, 606. [Google Scholar] [CrossRef] [Green Version]
- Shi, L.; Wang, X.; Wang, J.; Zhang, P.; Qi, F.; Cai, M.; Zhang, Y.; Zhou, X. Transcriptome analysis of Delta mig1 Delta mig2 mutant reveals their roles in methanol catabolism, peroxisome biogenesis and autophagy in methylotrophic yeast Pichia pastoris. Genes Genom. 2018, 40, 399–412. [Google Scholar] [CrossRef]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Group | Gene ID | Description | FPKM | RT-PCR log2-Fold Change | ||
|---|---|---|---|---|---|---|
| AFX | AP | log2-Fold Change | ||||
| Autophagy | PAS_chr3_1085 | Vesicle transport SNARE protein | 1044 ± 190.3 | 1362 ± 349.3 | 0.38 | 0.32 |
| PAS_chr1-4_0548 | Vacuolar proteinase | 8502 ± 100.4 | 10,225 ± 2434.9 | 0.27 | 0.34 | |
| PAS_chr2-2_0264 | Vacuolar sporting-related protein | 690 ± 49.3 | 844 ± 29.7 | 0.29 | 0.22 | |
| PAS_chr4_0998 | LETM1 homolog | 732 ± 149.2 | 605 ± 32.5 | 0.28 | 0.42 | |
| PAS_chr3_1047 | Clathrin assembly protein | 348 ± 50.4 | 449 ± 78.5 | 0.37 | 0.45 | |
| PAS_chr3_0230 | Heat shock protein 70 | 105,004 ± 10,357.2 | 124,125 ± 2985.4 | 0.2 | 0.14 | |
| MAPK signaling pathway | PAS_chr3_0449 | NAD-dependent dehydratase | 6376 ± 190.4 | 4661 ± 684.3 | 0.45 | 0.49 |
| PAS_chr4_0530 | Mitogen-activated protein kinase | 1990 ± 243.2 | 1445 ± 684.2 | 0.28 | 0.22 | |
| PAS_chr2-1_0124 | Protein kinase C | 4423 ± 146.3 | 3721 ± 283.5 | 0.25 | 0.22 | |
| PAS_chr2-1_0358 | Casein kinase | 1440 ± 424.2 | 1692 ± 168.4 | 0.23 | 0.42 | |
| Transcriptional factor | PAS_chr2-1_0142 | TBP-associated factor | 1188 ± 59.3 | 985 ± 38.5 | 0.27 | 0.46 |
| PAS_chr4_0204 | bZIP transcription factor | 4913 ± 484.2 | 5729 ± 258.6 | 0.22 | 0.25 | |
| Carbon metabolism | PAS_chr4_0470 | Aldehyde dehydrogenase | 6260 ± 243.1 | 2527 ± 864.3 | 1.3 | 2.52 |
| PAS_chr4_0152 | Methanol oxidase | 64,007 ± 734.2 | 35,007 ± 2698.5 | 0.83 | 0.92 | |
| PAS_chr1-4_0338 | Isocitrate lyase | 11,657 ± 985.2 | 6844 ± 49.3 | 0.77 | 1.03 | |
| PAS_chr4_0191 | Malate synthase | 64,415 ± 484.2 | 45,077 ± 5483.2 | 0.51 | 1.42 | |
| PAS_chr3_0403 | Acetyl-CoA synthase | 2023 ± 354.2 | 1435 ± 235.2 | 0.5 | 0.94 | |
| PAS_chr1-1_0475 | Citrate synthase | 38,285 ± 6323.2 | 28,902 ± 1984.3 | 0.41 | 0.63 | |
| Anti-stress | PAS_chr1-1_0433 | Mitochondrial thiol peroxidase | 4593 ± 354.2 | 5692 ± 254.6 | 0.29 | 0.37 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Wang, D.; Yu, X.; Sheng, P.; Zhang, G. The Transcriptomic Mechanism of a Novel Autolysis Induced by a Recombinant Antibacterial Peptide from Chicken Expressed in Pichia pastoris. Molecules 2022, 27, 2029. https://doi.org/10.3390/molecules27062029
Wang D, Yu X, Sheng P, Zhang G. The Transcriptomic Mechanism of a Novel Autolysis Induced by a Recombinant Antibacterial Peptide from Chicken Expressed in Pichia pastoris. Molecules. 2022; 27(6):2029. https://doi.org/10.3390/molecules27062029
Chicago/Turabian StyleWang, Dongsheng, Xinjun Yu, Ping Sheng, and Guohua Zhang. 2022. "The Transcriptomic Mechanism of a Novel Autolysis Induced by a Recombinant Antibacterial Peptide from Chicken Expressed in Pichia pastoris" Molecules 27, no. 6: 2029. https://doi.org/10.3390/molecules27062029