
Alcalase Potato Protein Hydrolysate-PPH902 Enhances Myogenic Differentiation and Enhances Skeletal Muscle Protein Synthesis under High Glucose Condition in C2C12 Cells


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Results
2.1. Effect of PPH902 on Cell Viability
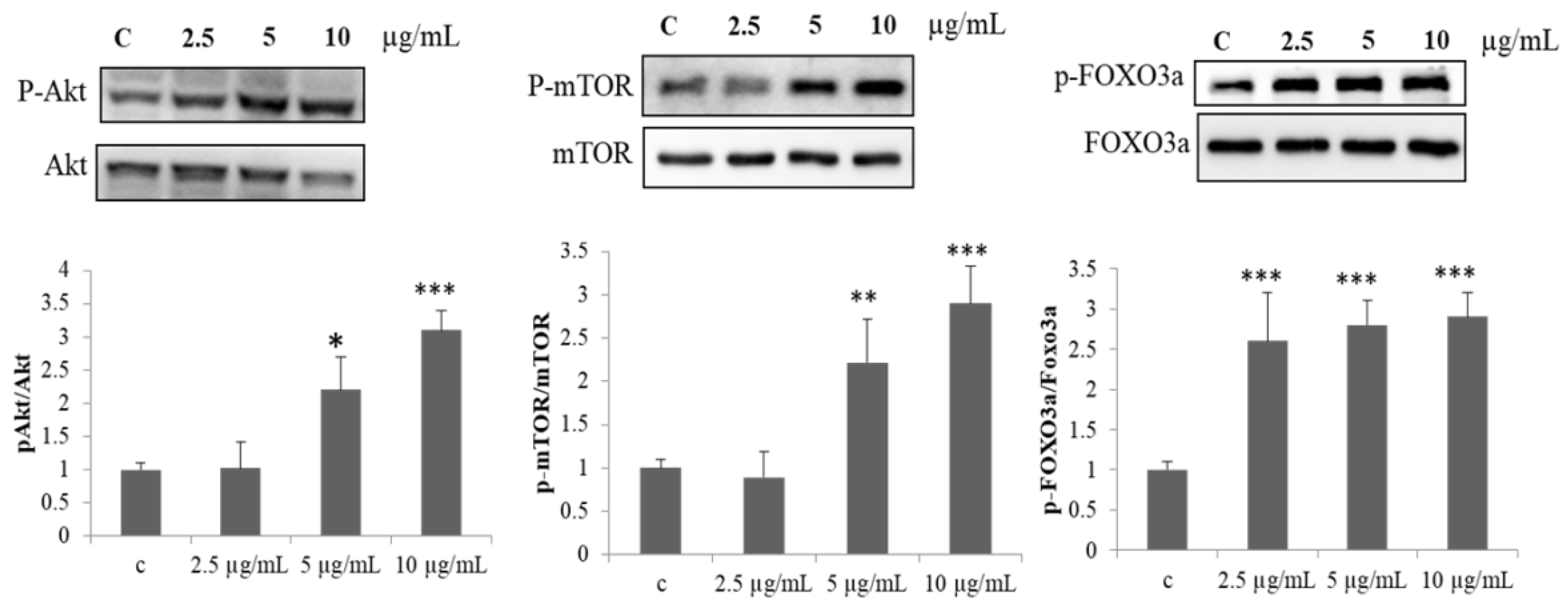
2.2. Effect of PPH902 on Protein Synthesis Mechanism
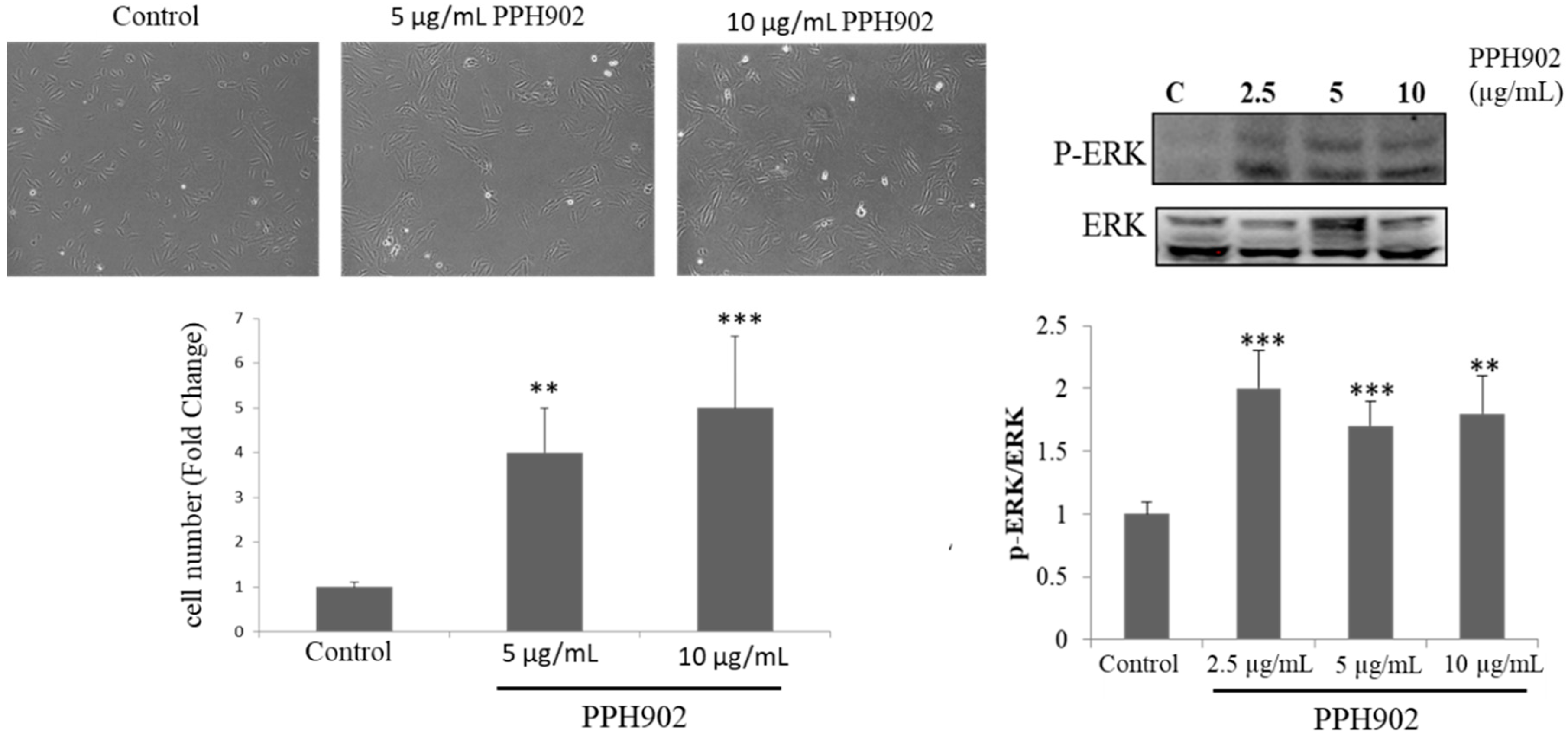
2.3. Effect of PPH902 on C2C12 Cell Proliferation
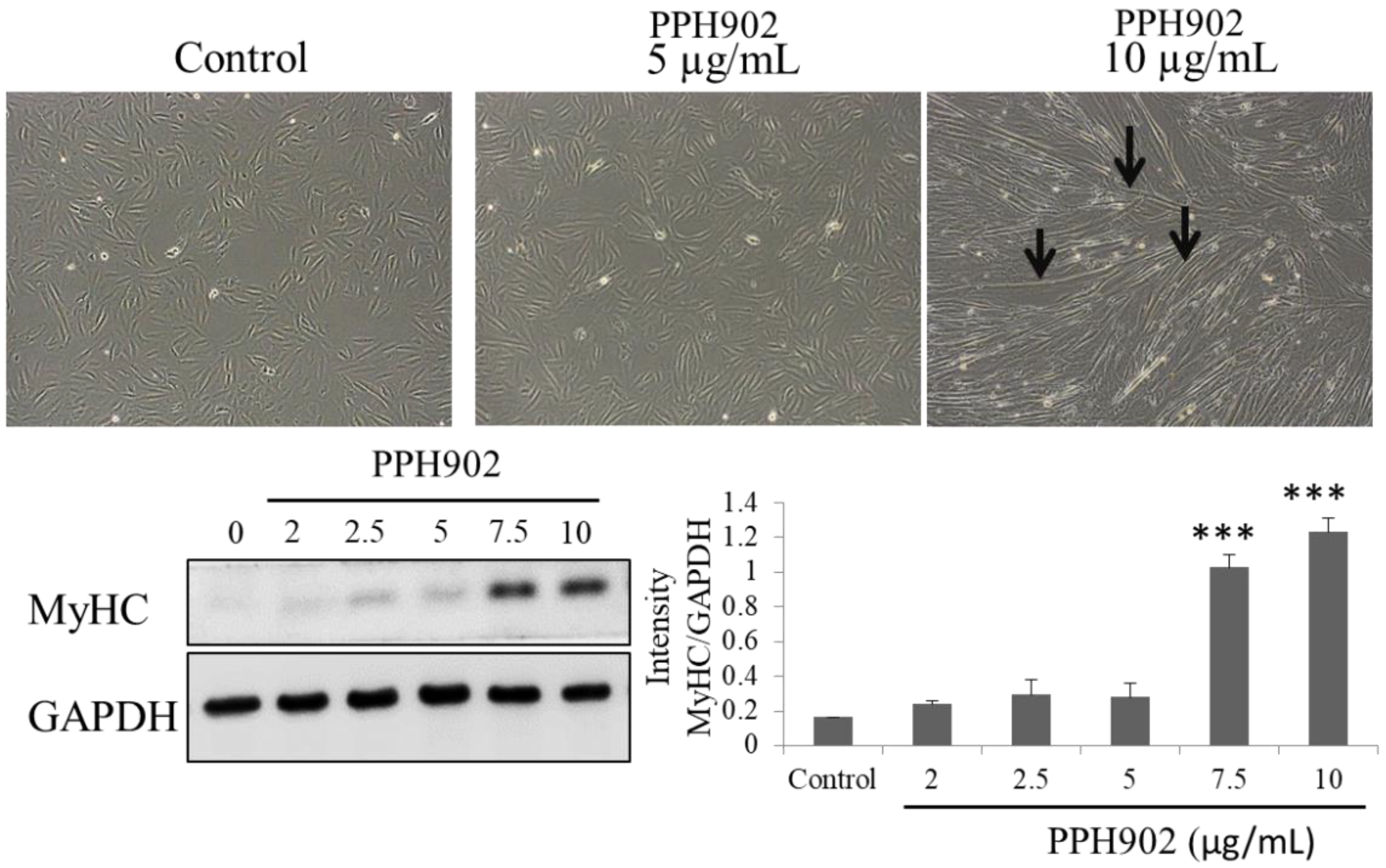
2.4. Effect of PPH902 on Myogenic Differentiation
2.5. Effect of PPH902 on High Glucose Induced C2C12 Cytotoxicity
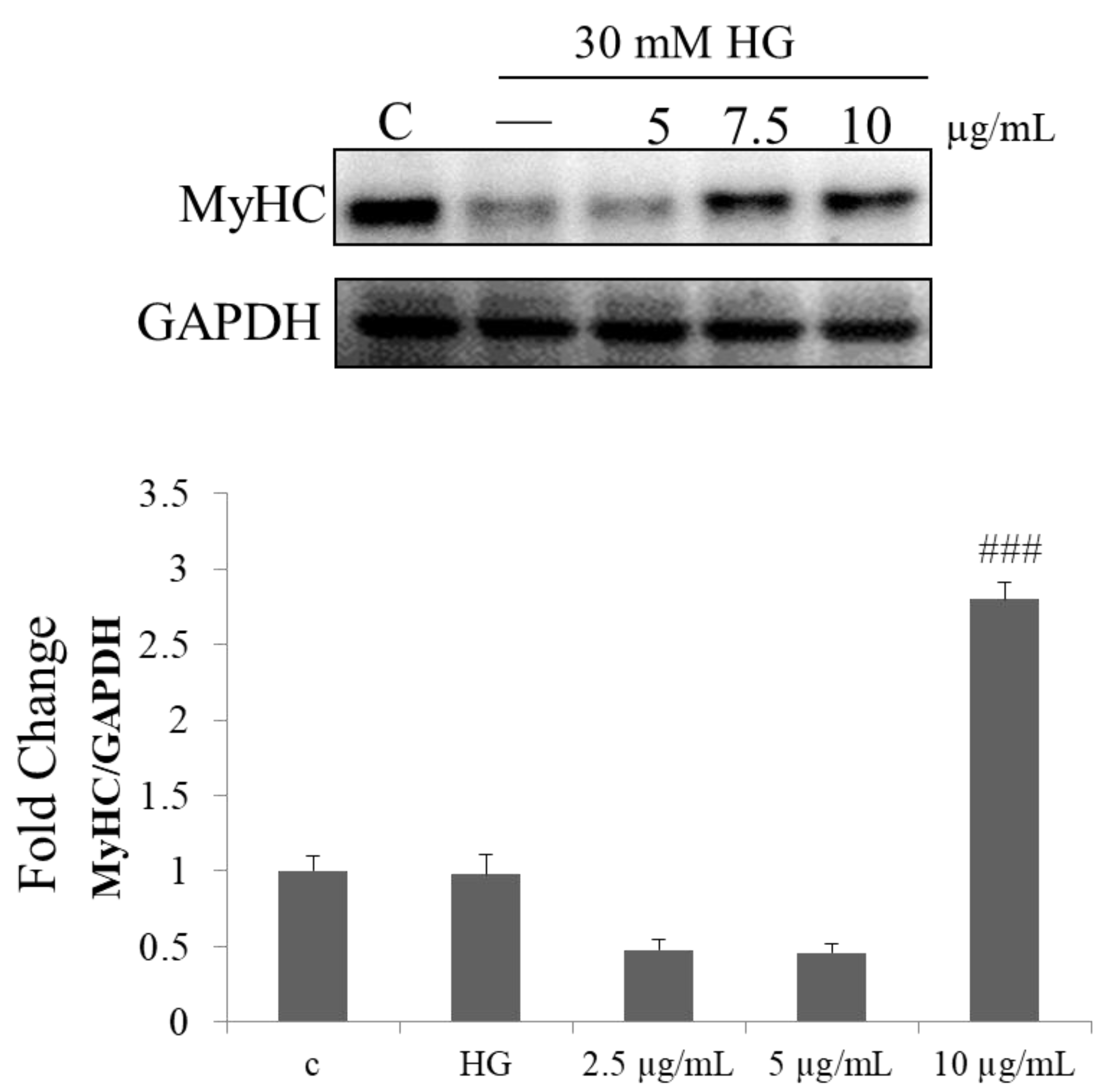
2.6. Effect of PPH902 on High Glucose Induced Myogenic Differentiation
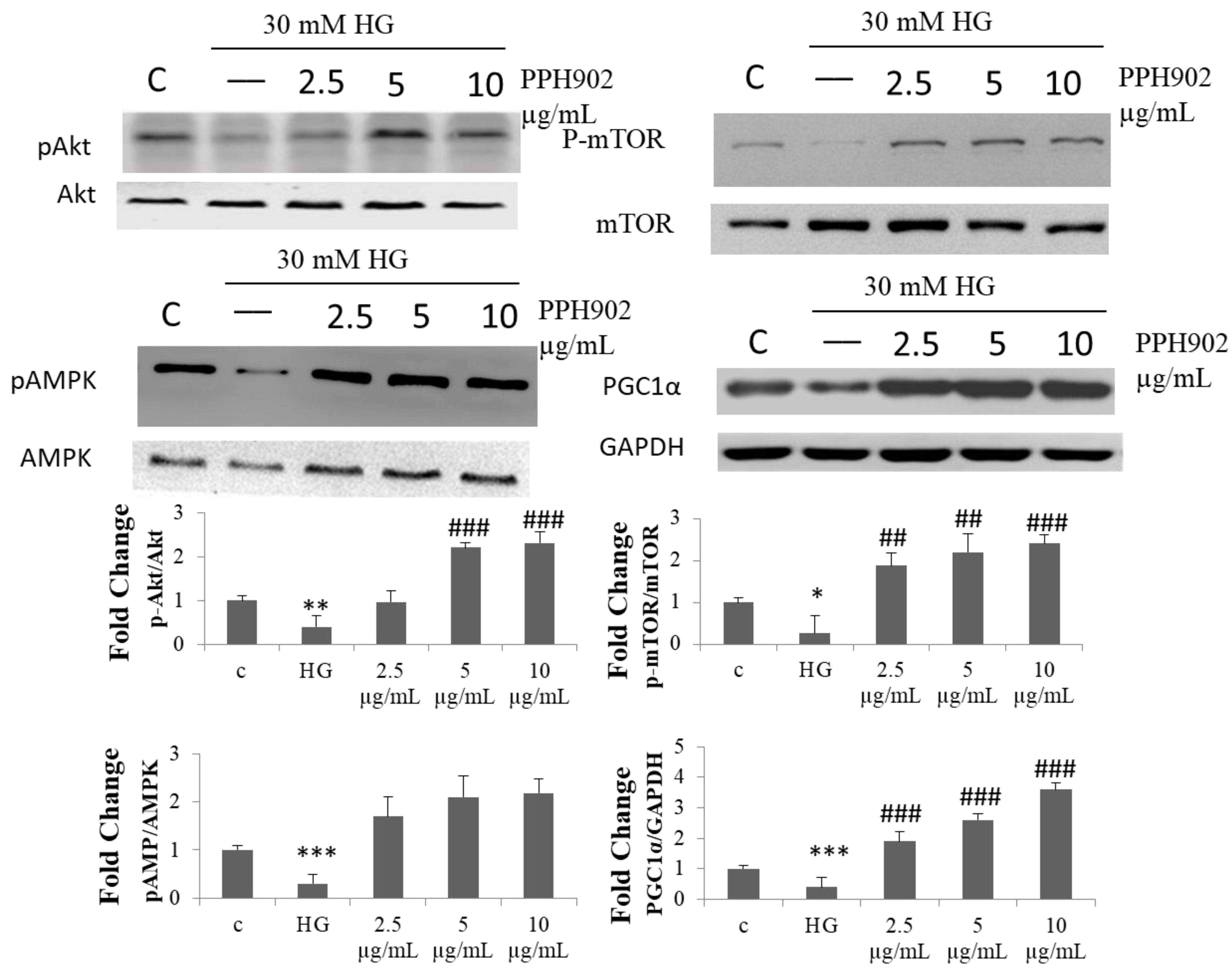
2.7. Effect of PPH902 on High Glucose Associated Modulations in Protein Synthesis Machinery in C2C12
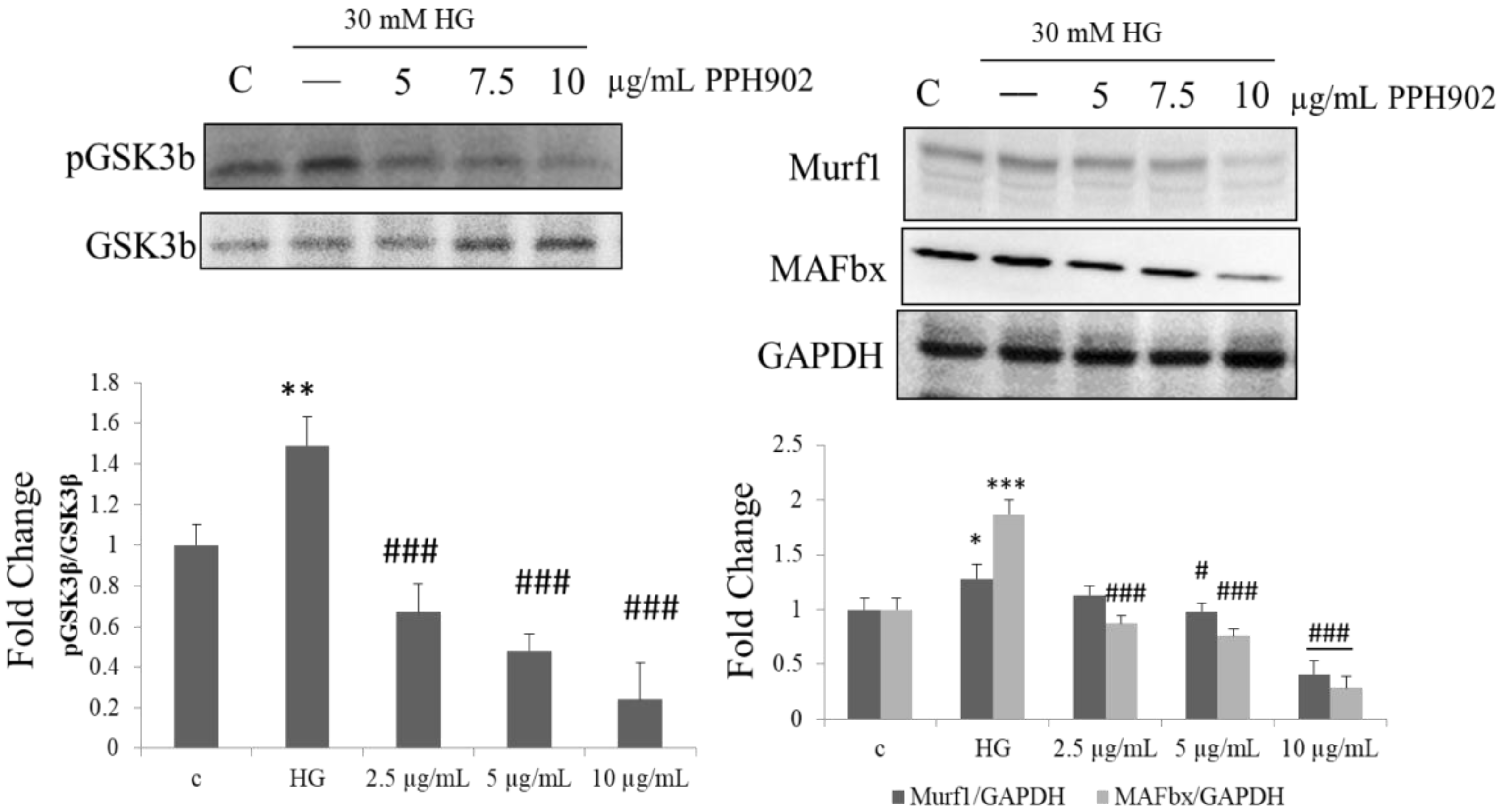
2.8. PPH902 Inhibits High Glucose Induced Muscle Atrophy by Downregulating Atrogin1/MAFbx and MuRF1
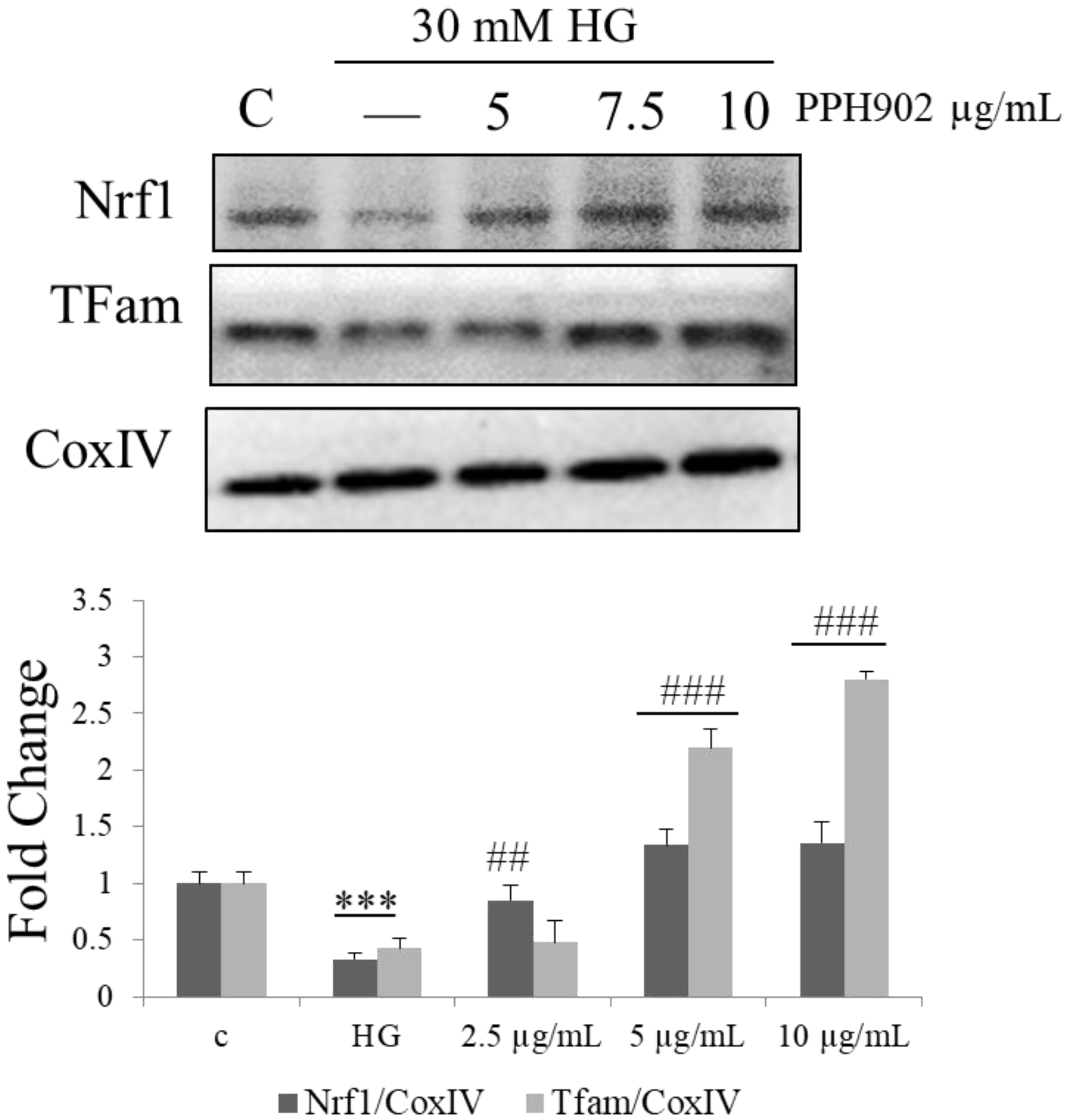
2.9. PPH902 Activates Mitochondrial Biogenesis via NRF-1 and TFAM in C2C12 Cells
3. Discussion
4. Materials and Methods
4.1. Cell Culture
4.2. MTT Assay
4.3. Differentiation of C2C12 Cells
4.4. Western Blotting Analysis
4.5. Statistical Analysis
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
Sample Availability
References
- Janssen, I.; Shepard, D.S.; Katzmarzyk, P.T.; Roubenoff, R. The healthcare costs of sarcopenia in the United States. J. Am. Geriatr. Soc. 2004, 52, 80–85. [Google Scholar] [CrossRef] [PubMed]
- Beaudart, C.; Rizzoli, R.; Bruyère, O.; Reginster, J.-Y.; Biver, E. Sarcopenia: Burden and challenges for public health. Arch. Public Health 2014, 72, 45. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Prokopidis, K.; Cervo, M.M.; Gandham, A.; Scott, D. Impact of Protein Intake in Older Adults with Sarcopenia and Obesity: A Gut Microbiota Perspective. Nutrients 2020, 12, 2285. [Google Scholar] [CrossRef]
- Paddon-Jones, D.; Rasmussen, B.B. Dietary protein recommendations and the prevention of sarcopenia. Curr Opin Clin. Nutr. Metab Care 2009, 12, 86–90. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sandri, M.; Sandri, C.; Gilbert, A.; Skurk, C.; Calabria, E.; Picard, A.; Walsh, K.; Schiaffino, S.; Lecker, S.H.; Goldberg, A.L. Foxo transcription factors induce the atrophy-related ubiquitin ligase atrogin-1 and cause skeletal muscle atrophy. Cell 2004, 117, 399–412. [Google Scholar] [CrossRef] [Green Version]
- Ruiz Ruiz, J.C.; Betancur Ancona, D.A.; Segura Campos, M.R. Bioactive vegetable proteins and peptides in lipid-lowering; nutraceutical potential. Nutr. Hosp. 2014, 29, 776–784. [Google Scholar] [CrossRef] [PubMed]
- Singh, B.P.; Vij, S.; Hati, S. Functional significance of bioactive peptides derived from soybean. Peptides 2014, 54, 171–179. [Google Scholar] [CrossRef]
- Erdmann, K.; Cheung, B.W.; Schröder, H. The possible roles of food-derived bioactive peptides in reducing the risk of cardiovascular disease. J. Nutr. Biochem. 2008, 19, 643–654. [Google Scholar] [CrossRef]
- Hartmann, R.; Meisel, H. Food-derived peptides with biological activity: From research to food applications. Curr. Opin. Biotechnol. 2007, 18, 163–169. [Google Scholar] [CrossRef]
- Zhao, J.; Xiong, Y.L.; McNear, D.H. Changes in structural characteristics of antioxidative soy protein hydrolysates resulting from scavenging of hydroxyl radicals. J. Food Sci. 2013, 78, C152–C159. [Google Scholar] [CrossRef] [PubMed]
- Kudoh, K.; Matsumoto, M.; Onodera, S.; Takeda, Y.; Ando, K.; Shiomi, N. Antioxidative activity and protective effect against ethanol-induced gastric mucosal damage of a potato protein hydrolysate. J. Nutr. Sci. Vitaminol. 2003, 49, 451–455. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Valiñas, M.A.; Lanteri, M.L.; Ten Have, A.; Andreu, A.B. Chlorogenic Acid Biosynthesis Appears Linked with Suberin Production in Potato Tuber (Solanum tuberosum). J. Agric. Food Chem. 2015, 63, 4902–4913. [Google Scholar] [CrossRef] [PubMed]
- Huang, C.Y.; Chiang, W.-D.; Pai, P.; Lin, W.-T. Potato protein hydrolysate attenuates high fat diet-induced cardiac apoptosis through SIRT1/ PGC-1á/Akt signalling. J. Funct. Foods 2015, 12, 389–398. [Google Scholar] [CrossRef]
- Chiang, W.-D.; Huang, C.Y.; Paul, C.R.; Lee, Z.-Y.; Lin, W.-T. Lipolysis stimulating peptides of potato protein hydrolysate effectively suppresses high-fat-diet-induced hepatocyte apoptosis and fibrosis in aging rats. Food Nutr. Res. 2016, 60, 31417. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sarti, S.; Ruggiero, E.; Coin, A.; Toffanello, E.D.; Perissinotto, E.; Miotto, F.; Pintore, G.; Inelmen, E.M.; Manzato, E.; Sergi, G. Dietary intake and physical performance in healthy elderly women: A 3-year follow-up. Exp. Gerontol. 2013, 48, 250–254. [Google Scholar] [CrossRef]
- Marthandam Asokan, S.; Wang, T.; Su, W.-T.; Lin, W.-T. Antidiabetic Effects of a Short Peptide of Potato Protein Hydrolysate in STZ-Induced Diabetic Mice. Nutrients 2019, 11, 779. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hu, W.-S.; Ting, W.-J.; Chiang, W.-D.; Pai, P.; Yeh, Y.-L.; Chang, C.-H.; Lin, W.-T.; Huang, C.-Y. The Heart Protection Effect of Alcalase Potato Protein Hydrolysate Is through IGF1R-PI3K-Akt Compensatory Reactivation in Aging Rats on High Fat Diets. Int J. Mol. Sci. 2015, 16, 10158–10172. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Candow, D.G.; Forbes, S.C.; Little, J.P.; Cornish, S.M.; Pinkoski, C.; Chilibeck, P.D. Effect of nutritional interventions and resistance exercise on aging muscle mass and strength. Biogerontology 2012, 13, 345–358. [Google Scholar] [CrossRef]
- Gutierrez-Salmean, G.; Ciaraldi, T.P.; Nogueira, L.; Barboza, J.; Taub, P.R.; Hogan, M.C.; Henry, R.R.; Meaney, E.; Villarreal, F.; Ceballos, G.; et al. Effects of (-)-epicatechin on molecular modulators of skeletal muscle growth and differentiation. J. Nutr. Biochem. 2014, 25, 91–94. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wang, S.H.; Wu, H.C.; Badrealam, K.F.; Kuo, Y.H.; Chao, Y.P.; Hsu, H.H.; Bau, D.T.; Viswanadha, V.P.; Chen, Y.H.; Lio, P.J.; et al. Taiwanin E Induces Cell Cycle Arrest and Apoptosis in Arecoline/4-NQO-Induced Oral Cancer Cells Through Modulation of the ERK Signaling Pathway. Front. Oncol. 2019, 9, 1309. [Google Scholar] [CrossRef]
- Asokan, S.M.; Wang, T.; Wang, M.-F.; Lin, W.-T. A novel dipeptide from potato protein hydrolysate augments the effects of exercise training against high-fat diet-induced damages in senescence-accelerated mouse-prone 8 by boosting pAMPK / SIRT1/ PGC-1α/ pFOXO3 pathway. Aging 2020, 12, 7334–7349. [Google Scholar] [CrossRef]
- Marthandam Asokan, S.; Yang, J.Y.; Lin, W.T. Anti-hypertrophic and anti-apoptotic effects of short peptides of potato protein hydrolysate against hyperglycemic condition in cardiomyoblast cells. Biomed. Pharmacother. Biomed. Pharmacother. 2018, 107, 1667–1673. [Google Scholar] [CrossRef] [PubMed]
- Chen, C.H.; Lin, J.W.; Huang, C.Y.; Yeh, Y.L.; Shen, C.Y.; Badrealam, K.F.; Ho, T.J.; Padma, V.V.; Kuo, W.W.; Huang, C.Y. The combined inhibition of the CaMKIIdelta and calcineurin signaling cascade attenuates IGF-IIR-induced cardiac hypertrophy. J. Cell. Physiol. 2020, 235, 3539–3547. [Google Scholar] [CrossRef] [PubMed]
- Liu, S.P.; Shibu, M.A.; Tsai, F.J.; Hsu, Y.M.; Tsai, C.H.; Chung, J.G.; Yang, J.S.; Tang, C.H.; Wang, S.; Li, Q.; et al. Tetramethylpyrazine reverses high-glucose induced hypoxic effects by negatively regulating HIF-1alpha induced BNIP3 expression to ameliorate H9c2 cardiomyoblast apoptosis. Nutr. Metab. 2020, 17, 12. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lin, C.C.; Chen, K.B.; Tsai, C.H.; Tsai, F.J.; Huang, C.Y.; Tang, C.H.; Yang, J.S.; Hsu, Y.M.; Peng, S.F.; Chung, J.G. Casticin inhibits human prostate cancer DU 145 cell migration and invasion via Ras/Akt/NF-kappaB signaling pathways. J. Food Biochem. 2019, 43, e12902. [Google Scholar] [CrossRef] [PubMed]









Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Chen, Y.-J.; Chang, C.-F.; Angayarkanni, J.; Lin, W.-T. Alcalase Potato Protein Hydrolysate-PPH902 Enhances Myogenic Differentiation and Enhances Skeletal Muscle Protein Synthesis under High Glucose Condition in C2C12 Cells. Molecules 2021, 26, 6577. https://doi.org/10.3390/molecules26216577
Chen Y-J, Chang C-F, Angayarkanni J, Lin W-T. Alcalase Potato Protein Hydrolysate-PPH902 Enhances Myogenic Differentiation and Enhances Skeletal Muscle Protein Synthesis under High Glucose Condition in C2C12 Cells. Molecules. 2021; 26(21):6577. https://doi.org/10.3390/molecules26216577
Chicago/Turabian StyleChen, Yi-Ju, Ching-Fang Chang, Jayaraman Angayarkanni, and Wan-Teng Lin. 2021. "Alcalase Potato Protein Hydrolysate-PPH902 Enhances Myogenic Differentiation and Enhances Skeletal Muscle Protein Synthesis under High Glucose Condition in C2C12 Cells" Molecules 26, no. 21: 6577. https://doi.org/10.3390/molecules26216577