
Comparisons of Isolation Methods, Structural Features, and Bioactivities of the Polysaccharides from Three Common Panax Species: A Review of Recent Progress

 ,
,
Abstract
:1. Introduction
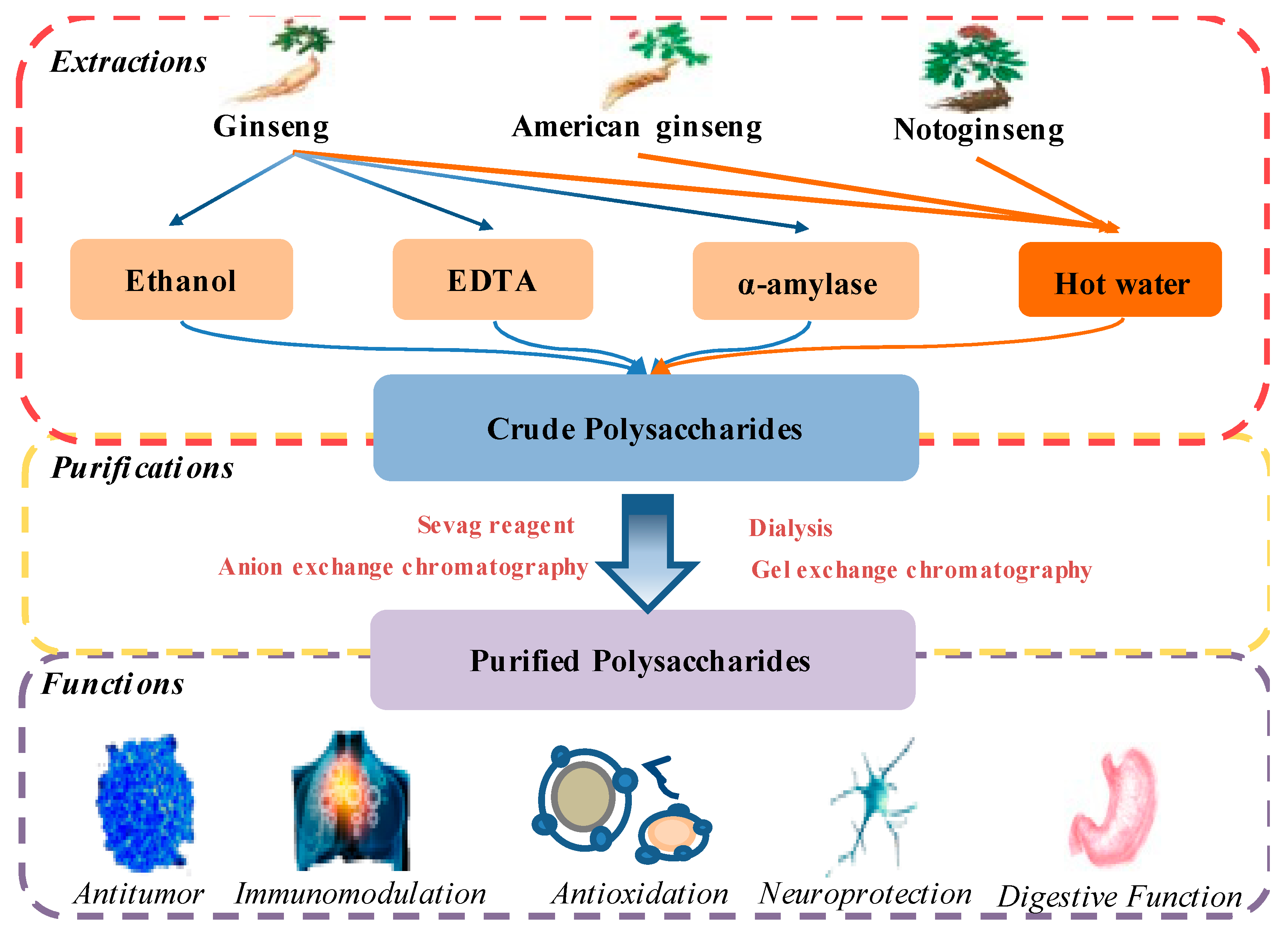
2. Extraction and Purification Methods
2.1. Extraction and Precipitation
2.2. Separation and Purification
3. Structural Characteristics
3.1. Polysaccharides from Ginseng
3.2. Polysaccharides from American Ginseng
3.3. Polysaccharides from Notoginseng
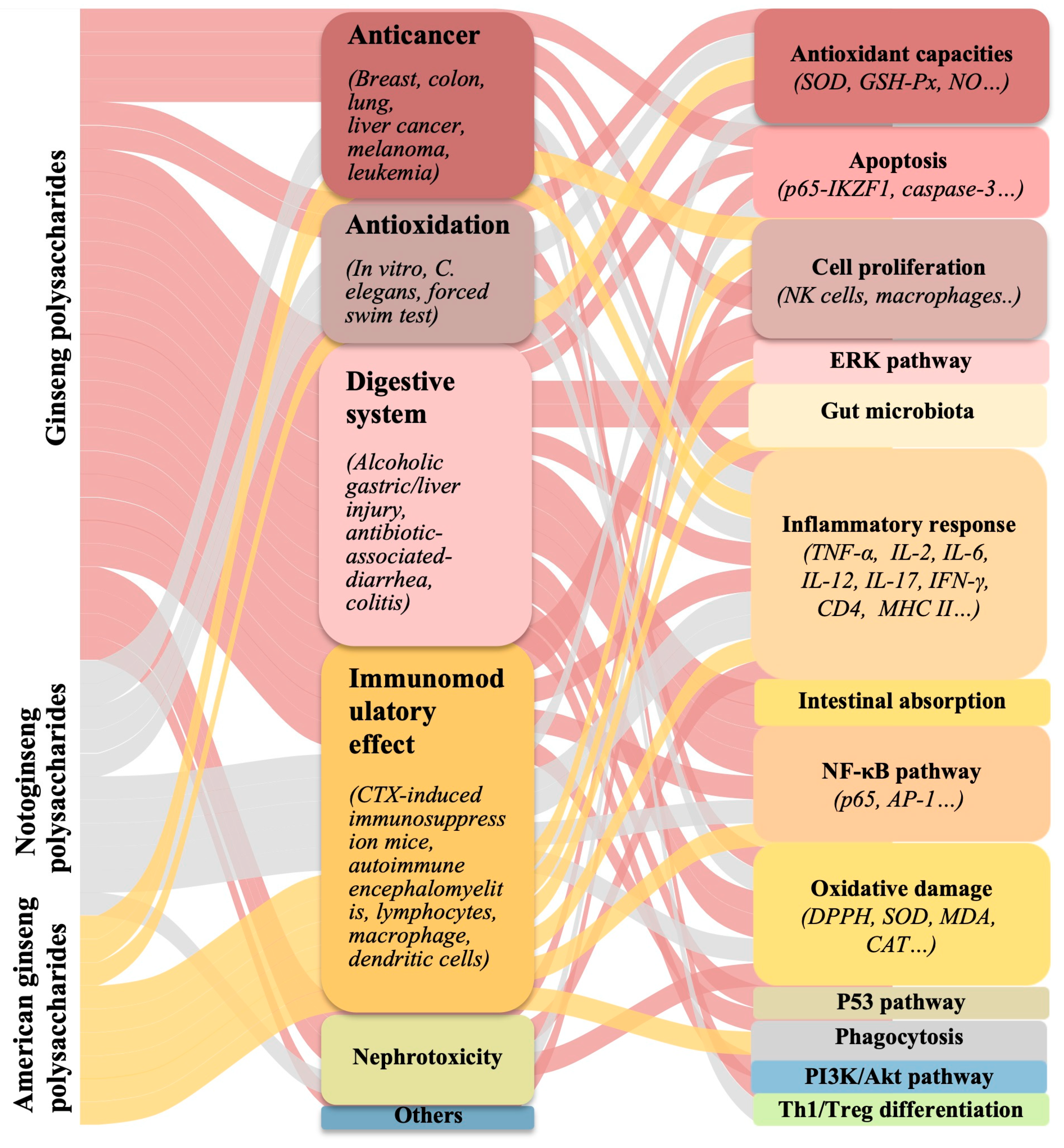
4. Biological Activities of Different Polysaccharides from the Three Species
4.1. Antitumor Activity
4.2. Immunomodulatory Activity
4.3. Anti-Oxidative Activity
4.4. Other Biological Functions
5. Conclusions and Future Perspective
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
Abbreviations
References
- Liu, L.; Xu, F.R.; Wang, Y.Z. Traditional uses, chemical diversity and biological activities of Panax, L. (Araliaceae): A review. J. Ethnopharmacol. 2020, 263, 112792. [Google Scholar] [CrossRef]
- You, S.; Shi, X.; Yu, D.; Zhao, D.; An, Q.; Wang, D.; Zhang, J.; Li, M.; Wang, C. Fermentation of Panax notoginseng root extract polysaccharides attenuates oxidative stress and promotes type I procollagen synthesis in human dermal fibroblast cells. BMC Complement. Med. 2021, 21, 34. [Google Scholar]
- Fan, S.; Zhang, Z.; Su, H.; Xu, P.; Qi, H.; Zhao, D.; Li, X. Panax ginseng clinical trials: Current status and future perspectives. Biomed. Pharm. 2020, 132, 110832. [Google Scholar] [CrossRef] [PubMed]
- Szczuka, D.; Nowak, A.; Zaklos-Szyda, M.; Kochan, E.; Szymanska, G.; Motyl, I.; Blasiak, J. American Ginseng (Panax quinquefolium L.) as a Source of Bioactive Phytochemicals with Pro-Health Properties. Nutrients 2019, 11, 5. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhang, S.; Chen, C.; Lu, W.; Wei, L. Phytochemistry, pharmacology, and clinical use of Panax notoginseng flowers buds. Phytother. Res. 2018, 32, 2155–2163. [Google Scholar] [CrossRef] [PubMed]
- Yang, W.Z.; Hu, Y.; Wu, W.Y.; Ye, M.; Guo, D.A. Saponins in the genus Panax, L. (Araliaceae): A systematic review of their chemical diversity. Phytochemistry 2014, 106, 7–24. [Google Scholar] [CrossRef]
- Yang, W.; Qiao, X.; Li, K.; Fan, J.; Bo, T.; Guo, D.A.; Ye, M. Identification and differentiation of Panax ginseng, Panax quinquefolium, and Panax notoginseng by monitoring multiple diagnostic chemical markers. Acta Pharm. Sin. B 2016, 6, 568–575. [Google Scholar] [CrossRef] [Green Version]
- Liu, W.; Leng, J.; Hou, J.G.; Jiang, S.; Wang, Z.; Liu, Z.; Gong, X.J.; Chen, C.; Wang, Y.P.; Li, W. Saponins derived from the stems and leaves of Panax ginseng attenuate scrotal heat-induced spermatogenic damage via inhibiting the MAPK mediated oxidative stress and apoptosis in mice. Phytother. Res. 2021, 35, 311–323. [Google Scholar] [CrossRef] [PubMed]
- Zhao, B.; Lv, C.; Lu, J. Natural occurring polysaccharides from Panax ginseng C. A. Meyer: A review of isolation, structures, and bioactivities. Int. J. Biol. Macromol. 2019, 133, 324–336. [Google Scholar] [CrossRef]
- Guo, M.; Shao, S.; Wang, D.; Zhao, D.; Wang, M. Recent progress in polysaccharides from Panax ginseng C. A. Meyer. Food Funct. 2021, 12, 494–518. [Google Scholar] [CrossRef]
- Jiao, L.; Zhang, X.; Li, B.; Liu, Z.; Wang, M.; Liu, S. Anti-tumour and immunomodulatory activities of oligosaccharides isolated from Panax ginseng C.A. Meyer. Int. J. Biol. Macromol. 2014, 65, 229–233. [Google Scholar] [CrossRef] [PubMed]
- Zhao, B.; Wang, X.; Liu, H.; Lv, C.; Lu, J. Structural characterization and antioxidant activity of oligosaccharides from Panax ginseng C. A. Meyer. Int. J. Biol. Macromol. 2020, 150, 737–745. [Google Scholar] [CrossRef] [PubMed]
- Yeo, C.R.; Yong, J.J.; Popovich, D.G. Isolation and characterization of bioactive polyacetylenes Panax ginseng Meyer roots. J. Pharm. Biomed. Anal. 2017, 139, 148–155. [Google Scholar] [CrossRef] [PubMed]
- Fukuyama, N.; Shibuya, M.; Orihara, Y. Antimicrobial polyacetylenes from Panax ginseng hairy root culture. Chem. Pharm. Bull. 2012, 60, 377–380. [Google Scholar] [CrossRef] [Green Version]
- Zhao, N.; Cheng, M.; Lv, W.; Wu, Y.; Liu, D.; Zhang, X. Peptides as Potential Biomarkers for Authentication of Mountain-Cultivated Ginseng and Cultivated Ginseng of Different Ages Using UPLC-HRMS. J. Agric. Food Chem. 2020, 68, 2263–2275. [Google Scholar] [CrossRef] [PubMed]
- Kwon, Y.; Byun, W.S.; Kim, B.Y.; Song, M.C.; Bae, M.; Yoon, Y.J.; Shin, J.; Lee, S.K.; Oh, D.C. Depsidomycins B and C: New Cyclic Peptides from a Ginseng Farm Soil-Derived Actinomycete. Molecules 2018, 23, 6. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chan, Y.S.; Wong, J.H.; Ng, T.B. Bioactive Proteins in Panax notoginseng Roots and Other Panax Species. Curr. Protein Pept. Sci. 2019, 20, 231–239. [Google Scholar] [CrossRef]
- Lee, D.Y.; Park, C.W.; Lee, S.J.; Park, H.R.; Kim, S.H.; Son, S.U.; Park, J.; Shin, K.S. Anti-Cancer Effects of Panax ginseng Berry Polysaccharides via Activation of Immune-Related Cells. Front. Pharm. 2019, 10, 1411. [Google Scholar] [CrossRef] [Green Version]
- Lee, D.Y.; Park, C.W.; Lee, S.J.; Park, H.R.; Seo, D.B.; Park, J.Y.; Park, J.; Shin, K.S. Immunostimulating and Antimetastatic Effects of Polysaccharides Purified from Ginseng Berry. Am. J. Chin. Med. 2019, 47, 823–839. [Google Scholar] [CrossRef]
- Li, B.; Zhang, N.; Feng, Q.; Li, H.; Wang, D.; Ma, L.; Liu, S.; Chen, C.; Wu, W.; Jiao, L. The core structure characterization and of ginseng neutral polysaccharide with the immune-enhancing activity. Int. J. Biol. Macromol. 2019, 123, 713–722. [Google Scholar] [CrossRef]
- Ghosh, R.; Bryant, D.L.; Farone, A.L. Panax quinquefolius (North American Ginseng) Polysaccharides as Immunomodulators: Current Research Status and Future Directions. Molecules 2020, 25, 24. [Google Scholar] [CrossRef]
- Lee, S.J.; In, G.; Han, S.T.; Lee, M.H.; Lee, J.W.; Shin, K.S. Structural characteristics of a red ginseng acidic polysaccharide rhamnogalacturonan I with immunostimulating activity from red ginseng. J. Ginseng Res. 2020, 44, 570–579. [Google Scholar] [CrossRef]
- Kim, M.H.; Byon, Y.Y.; Ko, E.J.; Song, J.Y.; Yun, Y.S.; Shin, T.; Joo, H.G. Immunomodulatory activity of ginsan, a polysaccharide of panax ginseng, on dendritic cells. Korean J. Physiol. Pharm. 2009, 13, 169–173. [Google Scholar] [CrossRef] [Green Version]
- Xiong, X.; Huang, G.; Huang, H. The antioxidant activities of phosphorylated polysaccharide from native ginseng. Int. J. Biol. Macromol. 2019, 126, 842–845. [Google Scholar] [CrossRef] [PubMed]
- Chen, F.; Huang, G. Antioxidant activity of polysaccharides from different sources of ginseng. Int. J. Biol. Macromol. 2019, 125, 906–908. [Google Scholar] [CrossRef] [PubMed]
- Feng, S.; Cheng, H.; Xu, Z.; Shen, S.; Yuan, M.; Liu, J.; Ding, C. Thermal stress resistance and aging effects of Panax notoginseng polysaccharides on Caenorhabditis elegans. Int. J. Biol. Macromol. 2015, 81, 188–194. [Google Scholar] [CrossRef] [PubMed]
- Liu, Y.; Sui, D.; Fu, W.; Sun, L.; Li, Y.; Yu, P.; Yu, X.; Zhou, Y.; Xu, H. Protective effects of polysaccharides from Panax ginseng on acute gastric ulcers induced by ethanol in rats. Food Funct. 2021, 12, 2741–2749. [Google Scholar] [CrossRef] [PubMed]
- Qu, Q.; Yang, F.; Zhao, C.; Liu, X.; Yang, P.; Li, Z.; Han, L.; Shi, X. Effects of fermented ginseng on the gut microbiota and immunity of rats with antibiotic-associated diarrhea. J. Ethnopharmacol. 2021, 267, 113594. [Google Scholar] [CrossRef]
- Chan, M.K.; Yu, Y.; Wulamu, S.; Wang, Y.; Wang, Q.; Zhou, Y.; Sun, L. Structural analysis of water-soluble polysaccharides isolated from Panax notoginseng. Int. J. Biol. Macromol. 2020, 155, 376–385. [Google Scholar] [CrossRef]
- Xia, Y.G.; Li, X.; Yu, L.S.; Liang, J.; Sun, H.M.; Kuang, H.X. Structural-fingerprinting of polysaccharides to discern Panax species by means of gas-liquid chromatography and mass spectrometry. Int. J. Biol. Macromol. 2020, 151, 932–943. [Google Scholar] [CrossRef]
- Kim, H.M.; Song, Y.; Hyun, G.H.; Long, N.P.; Park, J.H.; Hsieh, Y.S.Y.; Kwon, S.W. Characterization and Antioxidant Activity Determination of Neutral and Acidic Polysaccharides from Panax Ginseng, C.A. Meyer. Molecules 2020, 25, 4. [Google Scholar]
- Liu, S.; Yang, Y.; Qu, Y.; Guo, X.; Yang, X.; Cui, X.; Wang, C. Structural characterization of a novel polysaccharide from Panax notoginseng residue and its immunomodulatory activity on bone marrow dendritic cells. Int. J. Biol. Macromol. 2020, 161, 797–809. [Google Scholar] [CrossRef] [PubMed]
- Wang, M.; Guilbert, L.J.; Li, J.; Wu, Y.; Pang, P.; Basu, T.K.; Shan, J.J. A proprietary extract from North American ginseng (Panax quinquefolium) enhances IL-2 and IFN-gamma productions in murine spleen cells induced by Con-A. Int. Immunopharmacol. 2004, 4, 311–315. [Google Scholar] [CrossRef]
- Cui, L.; Wang, J.; Huang, R.; Tan, Y.; Zhang, F.; Zhou, Y.; Sun, L. Analysis of pectin from Panax ginseng flower buds and their binding activities to galectin-3. Int. J. Biol. Macromol. 2019, 128, 459–467. [Google Scholar] [CrossRef]
- Sun, C.; Chen, Y.; Li, X.; Tai, G.; Fan, Y.; Zhou, Y. Anti-hyperglycemic and anti-oxidative activities of ginseng polysaccharides in STZ-induced diabetic mice. Food Funct. 2014, 5, 845–848. [Google Scholar] [CrossRef] [PubMed]
- Cai, J.P.; Wu, Y.J.; Li, C.; Feng, M.Y.; Shi, Q.T.; Li, R.; Wang, Z.Y.; Geng, J.S. Panax ginseng polysaccharide suppresses metastasis via modulating Twist expression in gastric cancer. Int. J. Biol. Macromol. 2013, 57, 22–25. [Google Scholar] [CrossRef]
- Ji, L.; Jie, Z.; Ying, X.; Yue, Q.; Zhou, Y.; Sun, L. Structural characterization of alkali-soluble polysaccharides from Panax ginseng C. A. Meyer. R Soc. Open Sci. 2018, 5, 171644. [Google Scholar] [CrossRef] [Green Version]
- Yu, X.; Yang, X.; Cui, B.; Wang, L.; Ren, G. Antioxidant and immunoregulatory activity of alkali-extractable polysaccharides from North American ginseng. Int. J. Biol. Macromol. 2014, 65, 357–361. [Google Scholar] [CrossRef] [PubMed]
- Sun, L.; Wu, D.; Ning, X.; Yang, G.; Lin, Z.; Tian, M.; Zhou, Y. alpha-Amylase-assisted extraction of polysaccharides from Panax ginseng. Int. J. Biol. Macromol. 2015, 75, 152–157. [Google Scholar] [CrossRef]
- Song, Y.R.; Sung, S.K.; Jang, M.; Lim, T.G.; Cho, C.W.; Han, C.J.; Hong, H.D. Enzyme-assisted extraction, chemical characteristics, and immunostimulatory activity of polysaccharides from Korean ginseng (Panax ginseng Meyer). Int. J. Biol. Macromol. 2018, 116, 1089–1097. [Google Scholar] [CrossRef] [PubMed]
- Kim, H.; Kim, H.W.; Yu, K.W.; Suh, H.J. Polysaccharides fractionated from enzyme digests of Korean red ginseng water extracts enhance the immunostimulatory activity. Int. J. Biol. Macromol. 2019, 121, 913–920. [Google Scholar] [CrossRef]
- Zheng, Y.; Yang, G.; Zhao, Z.H.; Guo, T.; Shi, H.M.; Zhou, Y.F.; Sun, L. Structural analysis of ginseng polysaccharides extracted by EDTA solution. RSC Adv. 2016, 6, 2724–2730. [Google Scholar] [CrossRef]
- Wang, C.; Zheng, L.; Liu, S.; Guo, X.; Qu, Y.; Gao, M.; Cui, X.; Yang, Y. A novel acidic polysaccharide from the residue of Panax notoginseng and its hepatoprotective effect on alcoholic liver damage in mice. Int. J. Biol. Macromol. 2020, 149, 1084–1097. [Google Scholar] [CrossRef]
- Yip, K.M.; Xu, J.; Tong, W.S.; Zhou, S.S.; Yi, T.; Zhao, Z.Z.; Chen, H.B. Ultrasound-Assisted Extraction May Not Be a Better Alternative Approach than Conventional Boiling for Extracting Polysaccharides from Herbal Medicines. Molecules 2016, 21, 11. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhao, J.L.; Zhang, M.; Zhou, H.L. Microwave-Assisted Extraction, Purification, Partial Characterization, and Bioactivity of Polysaccharides from Panax ginseng. Molecules 2019, 24, 8. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Li, X.; Xiong, F.; Liu, Y.; Liu, F.; Hao, Z.; Chen, H. Total fractionation and characterization of the water-soluble polysaccharides isolated from Enteromorpha intestinalis. Int. J. Biol. Macromol. 2018, 111, 319–325. [Google Scholar] [CrossRef]
- Li, C.; Cai, J.; Geng, J.; Li, Y.; Wang, Z.; Li, R. Purification, characterization and anticancer activity of a polysaccharide from Panax ginseng. Int. J. Biol. Macromol. 2012, 51, 968–973. [Google Scholar] [CrossRef]
- Zhang, X.; Yu, L.; Bi, H.T.; Li, X.H.; Ni, W.H.; Han, H.; Li, N.; Wang, B.Q.; Zhou, Y.F.; Tai, G.H. Total fractionation and characterization of the water-soluble polysaccharides isolated from Panax ginseng C. A. Meyer. Carbohydr. Polym. 2009, 77, 544–552. [Google Scholar] [CrossRef]
- Byeon, S.E.; Lee, J.; Kim, J.H.; Yang, W.S.; Kwak, Y.S.; Kim, S.Y.; Choung, E.S.; Rhee, M.H.; Cho, J.Y. Molecular mechanism of macrophage activation by red ginseng acidic polysaccharide from Korean red ginseng. Mediat. Inflamm. 2012, 2012, 732860. [Google Scholar] [CrossRef] [PubMed]
- Zhu, Y.; Pettolino, F.; Mau, S.L.; Bacic, A. Characterization of cell wall polysaccharides from the medicinal plant Panax notoginseng. Phytochemistry 2005, 66, 1067–1076. [Google Scholar] [CrossRef]
- Wang, L.; Yao, Y.; Sang, W.; Yang, X.; Ren, G. Structural features and immunostimulating effects of three acidic polysaccharides isolated from Panax quinquefolius. Int. J. Biol. Macromol. 2015, 80, 77–86. [Google Scholar] [CrossRef]
- Jiao, L.; Zhang, X.; Wang, M.; Li, B.; Liu, Z.; Liu, S. Chemical and antihyperglycemic activity changes of ginseng pectin induced by heat processing. Carbohydr. Polym. 2014, 114, 567–573. [Google Scholar] [CrossRef]
- Ren, Y.; Bai, Y.; Zhang, Z.; Cai, W.; Del Rio Flores, A. The Preparation and Structure Analysis Methods of Natural Polysaccharides of Plants and Fungi: A Review of Recent Development. Molecules 2019, 24, 17. [Google Scholar] [CrossRef] [Green Version]
- Yin, S.Y.; Kim, H.J.; Kim, H.J. A comparative study of the effects of whole red ginseng extract and polysaccharide and saponin fractions on influenza A (H1N1) virus infection. Biol. Pharm. Bull. 2013, 36, 1002–1007. [Google Scholar] [CrossRef] [Green Version]
- Zhao, B.; Wang, X.Y.; Luo, W.; Lin, Y.; Lv, C.N.; Lu, J.C. Isolation and structural elucidation of a low-molecular-weight polysaccharide from the roots of Panax ginseng C. A. Meyer. Nat. Prod. Res. 2020, 1–8. [Google Scholar] [CrossRef]
- Gao, H.; Wang, F.; Lien, E.J.; Trousdale, M.D. Immunostimulating polysaccharides from Panax notoginseng. Pharm. Res. 1996, 13, 1196–1200. [Google Scholar] [CrossRef] [PubMed]
- Yu, X.H.; Liu, Y.; Wu, X.L.; Liu, L.Z.; Fu, W.; Song, D.D. Isolation, purification, characterization and immunostimulatory activity of polysaccharides derived from American ginseng. Carbohydr. Polym. 2017, 156, 9–18. [Google Scholar] [CrossRef]
- Zhu, Y.; Pettolino, F.; Mau, S.L.; Shen, Y.C.; Chen, C.F.; Kuo, Y.C.; Bacic, A. Immunoactive polysaccharide-rich fractions from Panax notoginseng. Planta Med. 2006, 72, 1193–1199. [Google Scholar] [CrossRef]
- Wang, J.; Sun, C.; Zheng, Y.; Pan, H.; Zhou, Y.; Fan, Y. The effective mechanism of the polysaccharides from Panax ginseng on chronic fatigue syndrome. Arch. Pharm. Res. 2014, 37, 530–538. [Google Scholar] [CrossRef] [PubMed]
- Shi, H.; Yu, L.; Shi, Y.; Lu, J.; Teng, H.; Zhou, Y.; Sun, L. Structural Characterization of a Rhamnogalacturonan I Domain from Ginseng and Its Inhibitory Effect on Galectin-3. Molecules 2017, 22, 6. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cheong, K.L.; Wu, D.T.; Deng, Y.; Leong, F.; Zhao, J.; Zhang, W.J.; Li, S.P. Qualitation and quantification of specific polysaccharides from Panax species using GC-MS, saccharide mapping and HPSEC-RID-MALLS. Carbohydr. Polym. 2016, 153, 47–54. [Google Scholar] [CrossRef]
- Yu, Y.; Shen, M.; Song, Q.; Xie, J. Biological activities and pharmaceutical applications of polysaccharide from natural resources: A review. Carbohydr. Polym. 2018, 183, 91–101. [Google Scholar] [CrossRef] [PubMed]
- Lee, J.H.; Shim, J.S.; Lee, J.S.; Kim, M.K.; Chung, M.S.; Kim, K.H. Pectin-like acidic polysaccharide from Panax ginseng with selective antiadhesive activity against pathogenic bacteria. Carbohydr. Res. 2006, 341, 1154–1163. [Google Scholar] [CrossRef] [PubMed]
- Zhang, X.; Li, S.; Sun, L.; Ji, L.; Zhu, J.; Fan, Y.; Tai, G.; Zhou, Y. Further analysis of the structure and immunological activity of an RG-I type pectin from Panax ginseng. Carbohydr. Polym. 2012, 89, 519–525. [Google Scholar] [CrossRef] [PubMed]
- Wang, J.; Flaisher-Grinberg, S.; Li, S.; Liu, H.; Sun, L.; Zhou, Y.; Einat, H. Antidepressant-like effects of the active acidic polysaccharide portion of ginseng in mice. J. Ethnopharmacol. 2010, 132, 65–69. [Google Scholar] [CrossRef]
- Fan, Y.Y.; Cheng, H.R.; Li, S.S.; Wang, J.; Liu, D.; Hao, M.A.; Gao, X.G.; Fan, E.X.; Tai, G.H.; Zhou, Y.F. Relationship of the inhibition of cell migration with the structure of ginseng pectic polysaccharides. Carbohydr. Polym. 2010, 81, 340–347. [Google Scholar] [CrossRef]
- Gong, L.H.; Lei, T.; Zhang, Z.L.; Liang, Q.C.; Zhai, F.G.; Wu, Y.Y.; Zhang, X.P.; Liu, J.Q.; Liu, J.W. Purification, compositional analysis and antioxidant properties of polysaccharides from black ginseng. Trop. J. Pharm. Res. 2018, 17, 1317–1324. [Google Scholar] [CrossRef]
- Luo, D.H.; Fang, B.S. Structural identification of ginseng polysaccharides and testing of their antioxidant activities. Carbohyd. Polym. 2008, 72, 376–381. [Google Scholar] [CrossRef]
- Li, S.; Qi, Y.; Chen, L.; Qu, D.; Li, Z.; Gao, K.; Chen, J.; Sun, Y. Effects of Panax ginseng polysaccharides on the gut microbiota in mice with antibiotic-associated diarrhea. Int. J. Biol. Macromol. 2019, 124, 931–937. [Google Scholar] [CrossRef] [PubMed]
- Baek, S.H.; Lee, J.G.; Park, S.Y.; Bae, O.N.; Kim, D.H.; Park, J.H. Pectic polysaccharides from Panax ginseng as the antirotavirus principals in ginseng. Biomacromolecules 2010, 11, 2044–2052. [Google Scholar] [CrossRef]
- Gao, X.; Zhi, Y.; Sun, L.; Peng, X.; Zhang, T.; Xue, H.; Tai, G.; Zhou, Y. The inhibitory effects of a rhamnogalacturonan I (RG-I) domain from ginseng pectin on galectin-3 and its structure-activity relationship. J. Biol. Chem. 2013, 288, 33953–33965. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Park, D.H.; Han, B.; Shin, M.S.; Hwang, G.S. Enhanced Intestinal Immune Response in Mice after Oral Administration of Korea Red Ginseng-Derived Polysaccharide. Polymers 2020, 12, 10. [Google Scholar] [CrossRef] [PubMed]
- Sun, L.; Peng, X.; Sun, P.; Shi, J.; Yuan, X.; Zhu, J.; Tai, G.; Zhou, Y. Structural characterization and immunostimulatory activity of a novel linear alpha-(1-->6)-d-glucan isolated from Panax ginseng C. A. Meyer. Glycoconj J. 2012, 29, 357–364. [Google Scholar] [CrossRef]
- Lee, J.H.; Shim, J.S.; Chung, M.S.; Lim, S.T.; Kim, K.H. Inhibition of pathogen adhesion to host cells by polysaccharides from Panax ginseng. Biosci. Biotechnol. Biochem. 2009, 73, 209–212. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sun, L.; Ropartz, D.; Cui, L.; Shi, H.; Ralet, M.C.; Zhou, Y. Structural characterization of rhamnogalacturonan domains from Panax ginseng C. A. Meyer. Carbohydr. Polym. 2019, 203, 119–127. [Google Scholar] [CrossRef] [PubMed]
- Yu, L.; Zhang, X.; Li, S.S.; Liu, X.Y.; Sun, L.; Liu, H.B.; Iteku, J.; Zhou, Y.F.; Tai, G.H. Rhamnogalacturonan I domains from ginseng pectin. Carbohydr. Polym. 2010, 79, 811–817. [Google Scholar] [CrossRef]
- Guo, Q.B.; Cui, S.W.; Kang, J.; Ding, H.H.; Wang, Q.; Wang, C. Non-starch polysaccharides from American ginseng: Physicochemical investigation and structural characterization. Food Hydrocolloids 2015, 44, 320–327. [Google Scholar] [CrossRef]
- Ghosh, R.; Smith, S.A.; Nwangwa, E.E.; Arivett, B.A.; Bryant, D.L.; Fuller, M.L.; Hayes, D.; Bowling, J.L.; Nelson, D.E.; DuBois, J.D.; et al. Panax quinquefolius (North American ginseng) cell suspension culture as a source of bioactive polysaccharides: Immunostimulatory activity and characterization of a neutral polysaccharide AGC1. Int. J. Biol. Macromol. 2019, 139, 221–232. [Google Scholar] [CrossRef]
- Ghosh, R.; Bryant, D.L.; Arivett, B.A.; Smith, S.A.; Altman, E.; Kline, P.C.; Farone, A.L. An acidic polysaccharide (AGC3) isolated from North American ginseng (Panax quinquefolius) suspension culture as a potential immunomodulatory nutraceutical. Curr. Res. Food Sci. 2020, 3, 207–216. [Google Scholar] [CrossRef]
- Wang, L.; Yu, X.; Yang, X.; Li, Y.; Yao, Y.; Lui, E.M.; Ren, G. Structural and anti-inflammatory characterization of a novel neutral polysaccharide from North American ginseng (Panax quinquefolius). Int. J. Biol. Macromol. 2015, 74, 12–17. [Google Scholar] [CrossRef]
- Zhu, W.J.; Han, B.; Sun, Y.; Wang, Z.Y.; Yang, X.H. Immunoregulatory effects of a glucogalactan from the root of Panax quinquefolium L. Carbohyd. Polym. 2012, 87, 2725–2729. [Google Scholar] [CrossRef]
- Akhter, K.F.; Mumin, M.A.; Lui, E.M.K.; Charpentier, P.A. Transdermal nanotherapeutics: Panax quinquefolium polysaccharide nanoparticles attenuate UVB-induced skin cancer. Int. J. Biol. Macromol. 2021, 181, 221–231. [Google Scholar] [CrossRef]
- Oshima, Y.; Sato, K.; Hikino, H. Isolation and hypoglycemic activity of quinquefolans A, B, and C, glycans of Panax quinquefolium roots. J. Nat. Prod. 1987, 50, 188–190. [Google Scholar] [CrossRef] [PubMed]
- Ohtani, K.; Mizutani, K.; Hatono, S.; Kasai, R.; Sumino, R.; Shiota, T.; Ushijima, M.; Zhou, J.; Fuwa, T.; Tanaka, O. Sanchinan-A, a reticuloendothelial system activating arabinogalactan from sanchi-ginseng (roots of Panax notoginseng). Planta Med. 1987, 53, 166–169. [Google Scholar] [CrossRef] [PubMed]
- Wu, Y.; Wang, D. Structural characterization and DPPH radical scavenging activity of an arabinoglucogalactan from Panax notoginseng root. J. Nat. Prod. 2008, 71, 241–245. [Google Scholar] [CrossRef] [PubMed]
- Wang, P.; Zhang, L.; Yao, J.; Shi, Y.; Li, P.; Ding, K. An arabinogalactan from flowers of Panax notoginseng inhibits angiogenesis by BMP2/Smad/Id1 signaling. Carbohydr. Polym. 2015, 121, 328–335. [Google Scholar] [CrossRef]
- Feng, S.L.; Cheng, H.R.; Xu, Z.; Feng, S.C.; Yuan, M.; Huang, Y.; Liao, J.Q.; Ding, C.B. Antioxidant and anti-aging activities and structural elucidation of polysaccharides from Panax notoginseng root. Process. Biochem. 2019, 78, 189–199. [Google Scholar] [CrossRef]
- Liu, Y.H.; Qin, H.Y.; Zhong, Y.Y.; Li, S.; Wang, H.J.; Wang, H.; Chen, L.L.; Tang, X.; Li, Y.L.; Qian, Z.Y.; et al. Neutral polysaccharide from Panax notoginseng enhanced cyclophosphamide antitumor efficacy in hepatoma H22-bearing mice. BMC Cancer 2021, 21, 37. [Google Scholar]
- Wang, J.; Zuo, G.; Li, J.; Guan, T.; Li, C.; Jiang, R.; Xie, B.; Lin, X.; Li, F.; Wang, Y.; et al. Induction of tumoricidal activity in mouse peritoneal macrophages by ginseng polysaccharide. Int. J. Biol. Macromol. 2010, 46, 389–395. [Google Scholar] [CrossRef] [PubMed]
- Zhou, H.; Yan, Y.; Zhang, X.; Zhao, T.; Xu, J.; Han, R. Ginseng polysaccharide inhibits MDA-MB-231 cell proliferation by activating the inflammatory response. Exp. Med. 2020, 20, 229. [Google Scholar] [CrossRef]
- Wang, C.Z.; Hou, L.; Wan, J.Y.; Yao, H.; Yuan, J.; Zeng, J.; Park, C.W.; Kim, S.H.; Seo, D.B.; Shin, K.S.; et al. Ginseng berry polysaccharides on inflammation-associated colon cancer: Inhibiting T-cell differentiation, promoting apoptosis, and enhancing the effects of 5-fluorouracil. J. Ginseng Res. 2020, 44, 282–290. [Google Scholar] [CrossRef]
- Lee, Y.Y.; Kim, S.W.; Youn, S.H.; Hyun, S.H.; Kyung, J.S.; In, G.; Park, C.K.; Jung, H.R.; Moon, S.J.; Kang, M.J.; et al. Biological Effects of Korean Red Ginseng Polysaccharides in Aged Rat Using Global Proteomic Approach. Molecules 2020, 25, 13. [Google Scholar] [CrossRef] [PubMed]
- Shin, M.S.; Hwang, S.H.; Yoon, T.J.; Kim, S.H.; Shin, K.S. Polysaccharides from ginseng leaves inhibit tumor metastasis via macrophage and NK cell activation. Int. J. Biol. Macromol. 2017, 103, 1327–1333. [Google Scholar] [CrossRef]
- Ni, W.; Zhang, X.; Wang, B.; Chen, Y.; Han, H.; Fan, Y.; Zhou, Y.; Tai, G. Antitumor activities and immunomodulatory effects of ginseng neutral polysaccharides in combination with 5-fluorouracil. J. Med. Food 2010, 13, 270–277. [Google Scholar] [CrossRef]
- Cheng, H.; Li, S.; Fan, Y.; Gao, X.; Hao, M.; Wang, J.; Zhang, X.; Tai, G.; Zhou, Y. Comparative studies of the antiproliferative effects of ginseng polysaccharides on HT-29 human colon cancer cells. Med. Oncol. 2011, 28, 175–181. [Google Scholar] [CrossRef] [PubMed]
- Wang, Y.; Huang, M.; Sun, R.; Pan, L. Extraction, characterization of a Ginseng fruits polysaccharide and its immune modulating activities in rats with Lewis lung carcinoma. Carbohyd. Polym. 2015, 127, 215–221. [Google Scholar] [CrossRef]
- Zhou, X.; Shi, H.; Jiang, G.; Zhou, Y.; Xu, J. Antitumor activities of ginseng polysaccharide in C57BL/6 mice with Lewis lung carcinoma. Tumour Biol. 2014, 35, 12561–12566. [Google Scholar] [CrossRef]
- Ma, J.; Liu, H.; Wang, X. Effect of ginseng polysaccharides and dendritic cells on the balance of Th1/Th2 T helper cells in patients with non-small cell lung cancer. J. Tradit. Chin. Med. 2014, 34, 641–645. [Google Scholar] [CrossRef] [Green Version]
- Huang, J.; Liu, D.; Wang, Y.; Liu, L.; Li, J.; Yuan, J.; Jiang, Z.; Jiang, Z.; Hsiao, W.W.; Liu, H.; et al. Ginseng polysaccharides alter the gut microbiota and kynurenine/tryptophan ratio, potentiating the antitumour effect of antiprogrammed cell death 1/programmed cell death ligand 1 (anti-PD-1/PD-L1) immunotherapy. Gut 2021. [Google Scholar] [CrossRef]
- Li, H.; Gu, L.; Zhong, Y.; Chen, Y.; Zhang, L.; Zhang, A.R.; Sobol, R.W.; Chen, T.; Li, J. Administration of polysaccharide from Panax notoginseng prolonged the survival of H22 tumor-bearing mice. Onco Targets Ther. 2016, 9, 3433–3441. [Google Scholar]
- Sun, Y.; Guo, M.; Feng, Y.; Zheng, H.; Lei, P.; Ma, X.; Han, X.; Guan, H.; Hou, D. Effect of ginseng polysaccharides on NK cell cytotoxicity in immunosuppressed mice. Exp. Med. 2016, 12, 3773–3777. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hwang, I.; Ahn, G.; Park, E.; Ha, D.; Song, J.Y.; Jee, Y. An acidic polysaccharide of Panax ginseng ameliorates experimental autoimmune encephalomyelitis and induces regulatory T cells. Immunol. Lett. 2011, 138, 169–178. [Google Scholar] [CrossRef]
- Bing, S.J.; Ha, D.; Hwang, I.; Park, E.; Ahn, G.; Song, J.Y.; Jee, Y. Protective Effects on Central Nervous System by Acidic Polysaccharide of Panax ginseng in Relapse-Remitting Experimental Autoimmune Encephalomyelitis-Induced SJL/J Mice. Am. J. Chin. Med. 2016, 44, 1099–1110. [Google Scholar] [CrossRef]
- Akhter, K.F.; Mumin, M.A.; Lui, E.M.K.; Charpentier, P.A. Fabrication of fluorescent labeled ginseng polysaccharide nanoparticles for bioimaging and their immunomodulatory activity on macrophage cell lines. Int. J. Biol. Macromol. 2018, 109, 254–262. [Google Scholar] [CrossRef]
- Park, K.M.; Kim, Y.S.; Jeong, T.C.; Joe, C.O.; Shin, H.J.; Lee, Y.H.; Nam, K.Y.; Park, J.D. Nitric oxide is involved in the immunomodulating activities of acidic polysaccharide from Panax ginseng. Planta Med. 2001, 67, 122–126. [Google Scholar] [CrossRef]
- Youn, S.H.; Lee, S.M.; Han, C.K.; In, G.; Park, C.K.; Hyun, S.H. Immune Activity of Polysaccharide Fractions Isolated from Korean Red Ginseng. Molecules 2020, 25, 16. [Google Scholar] [CrossRef]
- Du Xiao, F.; Jiang, C.Z.; Wu, C.F.; Won, E.K.; Choung, S.Y. Synergistic immunostimulatory effect of pidotimod and red ginseng acidic polysaccharide on humoral immunity of immunosuppressed mice. Pharmazie 2008, 63, 904–908. [Google Scholar]
- Meng, J.; Meng, Y.; Liang, Z.; Du, L.; Zhang, Z.; Hu, X.; Shan, F. Phenotypic and functional analysis of the modification of murine bone marrow dendritic cells (BMDCs) induced by neutral Ginseng polysaccharides (NGP). Hum. Vaccines Immunother. 2013, 9, 233–241. [Google Scholar] [CrossRef] [Green Version]
- Zhang, S.D.; Yin, Y.X.; Wei, Q. Immunopotentiation on murine spleen lymphocytes induced by polysaccharide fraction of Panax ginseng via upregulating calcineurin activity. APMIS 2010, 118, 288–296. [Google Scholar] [CrossRef] [PubMed]
- Lim, T.S.; Na, K.; Choi, E.M.; Chung, J.Y.; Hwang, J.K. Immunomodulating activities of polysaccharides isolated from Panax ginseng. J. Med. Food 2004, 7, 1–6. [Google Scholar] [CrossRef] [PubMed]
- Wang, K.; Zhang, H.; Han, Q.; Lan, J.; Chen, G.; Cao, G.; Yang, C. Effects of astragalus and ginseng polysaccharides on growth performance, immune function and intestinal barrier in weaned piglets challenged with lipopolysaccharide. J. Anim. Physiol. Anim. Nutr. 2020, 104, 1096–1105. [Google Scholar] [CrossRef]
- Lemmon, H.R.; Sham, J.; Chau, L.A.; Madrenas, J. High molecular weight polysaccharides are key immunomodulators in North American ginseng extracts: Characterization of the ginseng genetic signature in primary human immune cells. J. Ethnopharmacol. 2012, 142, 1–13. [Google Scholar] [CrossRef] [PubMed]
- Azike, C.G.; Charpentier, P.A.; Lui, E.M. Stimulation and suppression of innate immune function by American ginseng polysaccharides: Biological relevance and identification of bioactives. Pharm. Res. 2015, 32, 876–897. [Google Scholar] [CrossRef]
- Zhou, R.; He, D.; Xie, J.; Zhou, Q.; Zeng, H.; Li, H.; Huang, L. The Synergistic Effects of Polysaccharides and Ginsenosides From American Ginseng (Panax quinquefolius L.) Ameliorating Cyclophosphamide-Induced Intestinal Immune Disorders and Gut Barrier Dysfunctions Based on Microbiome-Metabolomics Analysis. Front. Immunol. 2021, 12, 665901. [Google Scholar] [CrossRef]
- Yang, X.S.; Wang, L.J.; Dung, C.; Lui, E.M.K.; Ren, G.X. Optimization of Ultrasonic-Assisted Extraction Process of Polysaccharides from American Ginseng and Evaluation of Its Immunostimulating Activity. J. Integr. Agric. 2014, 13, 2807–2815. [Google Scholar] [CrossRef]
- Akhter, K.F.; Mumin, M.A.; Lui, E.M.K.; Charpentier, P.A. Immunoengineering with Ginseng Polysaccharide Nanobiomaterials through Oral Administration in Mice. ACS Biomater. Sci. Eng. 2019, 5, 2916–2925. [Google Scholar] [CrossRef]
- Hua, M.; Sun, Y.; Shao, Z.; Lu, J.; Lu, Y.; Liu, Z. Functional soluble dietary fiber from ginseng residue: Polysaccharide characterization, structure, antioxidant, and enzyme inhibitory activity. J. Food Biochem. 2020, 44, e13524. [Google Scholar] [CrossRef] [PubMed]
- Wang, J.; Li, S.; Fan, Y.; Chen, Y.; Liu, D.; Cheng, H.; Gao, X.; Zhou, Y. Anti-fatigue activity of the water-soluble polysaccharides isolated from Panax ginseng C. A. Meyer. J. Ethnopharmacol. 2010, 130, 421–423. [Google Scholar] [CrossRef] [PubMed]
- Wang, J.; Li, Y.; Luo, P.; Chen, Y.; Xi, Q.; Wu, H.; Zhao, W.; Shu, G.; Wang, S.; Gao, P.; et al. Oral supplementation with ginseng polysaccharide promotes food intake in mice. Brain Behav. 2019, 9, e01340. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wei, X.M.; Jiang, S.; Li, S.S.; Sun, Y.S.; Wang, S.H.; Liu, W.C.; Wang, Z.; Wang, Y.P.; Zhang, R.; Li, W. Endoplasmic Reticulum Stress-Activated PERK-eIF2alpha-ATF4 Signaling Pathway is Involved in the Ameliorative Effects of Ginseng Polysaccharides against Cisplatin-Induced Nephrotoxicity in Mice. ACS Omega 2021, 6, 8958–8966. [Google Scholar] [CrossRef]
- Li, J.; Li, R.; Li, N.; Zheng, F.; Dai, Y.; Ge, Y.; Yue, H.; Yu, S. Mechanism of antidiabetic and synergistic effects of ginseng polysaccharide and ginsenoside Rb1 on diabetic rat model. J. Pharm. Biomed. Anal. 2018, 158, 451–460. [Google Scholar] [CrossRef]
- Shen, H.; Gao, X.J.; Li, T.; Jing, W.H.; Han, B.L.; Jia, Y.M.; Hu, N.; Yan, Z.X.; Li, S.L.; Yan, R. Ginseng polysaccharides enhanced ginsenoside Rb1 and microbial metabolites exposure through enhancing intestinal absorption and affecting gut microbial metabolism. J. Ethnopharmacol. 2018, 216, 47–56. [Google Scholar] [CrossRef] [PubMed]
- Bing, S.J.; Kim, M.J.; Ahn, G.; Im, J.; Kim, D.S.; Ha, D.; Cho, J.; Kim, A.; Jee, Y. Acidic polysaccharide of Panax ginseng regulates the mitochondria/caspase-dependent apoptotic pathway in radiation-induced damage to the jejunum in mice. Acta Histochem. 2014, 116, 514–521. [Google Scholar] [CrossRef] [PubMed]
- Yoo, D.G.; Kim, M.C.; Park, M.K.; Park, K.M.; Quan, F.S.; Song, J.M.; Wee, J.J.; Wang, B.Z.; Cho, Y.K.; Compans, R.W.; et al. Protective effect of ginseng polysaccharides on influenza viral infection. PLoS ONE 2012, 7, e33678. [Google Scholar] [CrossRef]
- Zuo, Y.H.; Han, Q.B.; Dong, G.T.; Yue, R.Q.; Ren, X.C.; Liu, J.X.; Liu, L.; Luo, P.; Zhou, H. Panax ginseng Polysaccharide Protected H9c2 Cardiomyocyte From Hypoxia/Reoxygenation Injury Through Regulating Mitochondrial Metabolism and RISK Pathway. Front. Physiol. 2018, 9, 699. [Google Scholar] [CrossRef] [PubMed]
- Kim, J.E.; Jang, S.G.; Lee, C.H.; Lee, J.Y.; Park, H.; Kim, J.H.; Lee, S.; Kim, S.H.; Park, E.Y.; Lee, K.W.; et al. Beneficial effects on skin health using polysaccharides from red ginseng by-product. J. Food Biochem. 2019, 43, e12961. [Google Scholar] [CrossRef]
- Jia, D.; Deng, Y.; Gao, J.; Liu, X.; Chu, J.; Shu, Y. Neuroprotective effect of Panax notoginseng plysaccharides against focal cerebral ischemia reperfusion injury in rats. Int. J. Biol. Macromol. 2014, 63, 177–180. [Google Scholar] [CrossRef]
- Lu, Y.L.; Liang, J.; Zhou, F.Y.; Kuang, H.X.; Xia, Y.G. Discrimination and characterization of Panax polysaccharides by 2D COS-IR spectroscopy with chemometrics. Int. J. Biol. Macromol. 2021, 183, 193–202. [Google Scholar] [CrossRef]
- Cho, Y.J.; Son, H.J.; Kim, K.S. A 14-week randomized, placebo-controlled, double-blind clinical trial to evaluate the efficacy and safety of ginseng polysaccharide (Y-75). J. Transl. Med. 2014, 12, 283. [Google Scholar] [CrossRef] [Green Version]




{kind=link}
{kind=link}
{kind=link}
| No. | Polysaccharide Name | Extraction and Purification | Composition Ratio of Sugars (Mass Percentage or Molar Ratio) | Molecular Weight (Da) | Structure Feature | Refs |
|---|---|---|---|---|---|---|
| (1) | WGNP | Hot water, purification by DEAE-Cellulose, dialysis | Gal:Glu:Ara = 1.1:97.9:1.0 | 1.61–7.04 × 104 | [31] | |
| (2) | WGAP | Hot water, purification by DEAE-Cellulose, dialysis | Gal:Glu:Ara:GalA:GluA = 24.4:24.0:18.1:32.2:1.3 | 5–8 × 104 | [31] | |
| (3) | WGPN | Hot water, purification by DEAE-Cellulose | Gal:Glc:Ara = 3.3:95.3:1.3 | Starch-like glucans | [35] | |
| (4) | WGPA | Hot water, purification by DEAE-Cellulose | Gal:Glc:Ara:GalA = 17.1:18.0:15.5:44.2 | HG and RG-I-rich pectin and arabinogalactan | [35] | |
| (5) | WGPA | Hot water, purification by DEAE-Cellulose | Gal:Glc:Ara:Rha:GalA:GlcA= 18.0:18.5:15.5:2.5:44.2:1.3 | RG-I, HG-rich pectin, and arabinogalactan | [65] | |
| (6) | WGPA-A | Hot water, purification by DEAE-Cellulose twice | Gal:Glc:Ara:Rha:Man:GalA:GlcA = 13.3:3.7:22.7:6.0:0.5:51.7:2.2 | [59] | ||
| (7) | WGPA-N | Hot water, purification by DEAE-Cellulose twice | Gal:Glc:Ara = 18:66.3:15.7 | Starch-like glucans and AG | [59] | |
| (8) | WGPA-1-RG | Hot water, purification by Sepharose CL-6B column, DEAE-Cellulose | Gal:Glc:Ara:Rha:Gla A = 56.2:3.5:34.0:0.2:1.8 | AG with trace RG-I | [66] | |
| (9) | WGPA-2-RG | Hot water, purification by Sepharose CL-6B column, DEAE-Cellulose | Gal:Glc:Ara:Rha:Gla A = 44.4:2.9:40.9:4.1:5.3 | AG with minor RG-I | [66] | |
| (10) | WGPA-3-RG | Hot water, purification by Sepharose CL-6B column, DEAE-Cellulose | Gal:Glc:Ara:Rha:Gla A = 29.0:3.2:38.0:7.3:20.2 | HG- and RG-I-domains | [66] | |
| (11) | WGPA-4-RG | Hot water, purification by Sepharose CL-6B column, DEAE-Cellulose | Gal:Glc:Ara:Rha:Gla A = 13.5:4.4:26.1:11.4:38.4 | HG- and RG-I-domains | [66] | |
| (12) | WGPA-1-HG | Hot water, purification by Sepharose CL-6B column, DEAE-Cellulose | Gal:Glc:Ara:Rha:Gla A = 15.2:7.6:7.1:3.6:62.4 | 3.5 × 103 | HG with minor RG-I | [66] |
| (13) | WGPA-2-HG | Hot water, purification by Sepharose CL-6B column, DEAE-Cellulose | Gal:Glc:Ara:Rha:Gla A = 5.1:1.9:4.6:3.0:83.6 | 6.5 × 103 | HG with minor RG-I | [66] |
| (14) | WGPA-3-HG | Hot water, purification by Sepharose CL-6B column, DEAE-Cellulose | Gal:Glc:Ara:Rha:Gla A = 3.5:1.3:2.2:1.5:90.9 | 1.6 × 104 | HG with trace RG-I | [66] |
| (15) | WGPA-4-HG | Hot water, purification by Sepharose CL-6B column, DEAE-Cellulose | Gal:Glc:Gla A = 5.9:2.0:92.1 | 4.5 × 104 | HG with little RG-I | [66] |
| (16) | WGPA-1-RG | Hot water, purification by DEAE-Cellulose, dialysis, Sepharose CL-6B column | Gal:Glc:Ara:Rha:Man:GalA:GlcA = 56.2:3.5:34.0:0.2:2.5:1.8:1.9 | 1.0 × 105 | Arabinogalactans containing RG-I domains | [48] |
| (17) | WGPA-2-RG | Hot water, purification by DEAE-Cellulose, dialysis, Sepharose CL-6B column | Gal:Glc:Ara:Rha:GalA:GlcA = 23.3:3.4:25.2:2.3:41.3:4.5 | 1.1 × 105 | Arabinogalactans containing RG-I domains | [48] |
| (18) | WGPA-1-HG | Hot water, purification by DEAE-Cellulose, dialysis, Sepharose CL-6B column | Gal:Glc:Ara:Rha:Man:GalA:GlcA = 15.2:7.6:7.1:1.6:3.6:62.4:4.5 | 3.5 × 103 | α-(1→4)-GalA backbone (HG) | [48] |
| (19) | WGPA-2-HG | Hot water, purification by DEAE-Cellulose, dialysis, Sepharose CL-6B column | Gal:Glc:Ara:Rha:Man:GalA:GlcA = 5.1:1.9:4.6:3.0:0.2:83.6:1.6 | 6.5 × 103 | α-(1→4)-GalA backbone (HG) | [48] |
| (20) | WGPA-3-HG | Hot water, purification by DEAE-Cellulose, dialysis, Sepharose CL-6B column | Gal:Glc:Ara:Rha:Man:GalA:GlcA = 3.5:1.3:2.2:1.5:90.9:0.5 | 1.6 × 104 | α-(1→4)-GalA backbone (HG) | [48] |
| (21) | WGPA-4-HG | Hot water, purification by DEAE-Cellulose, dialysis, Sepharose CL-6B column | Gal:Glc:GlcA = 5.2:2.0:92.1 | 4.5 × 104 | α-(1→4)-GalA backbone (HG) | [48] |
| (22) | GBPP | Hot water, purification by dialysis (20 kDa) | Gal:Glc:Ara:Rha:Man:GalA:Xyl = 26.6:5.4:19.5:8.4:1.5:15.2:2.2 | AG-II | [18] | |
| (23) | GBPP-I | Hot water, purification by dialysis (20 kDa), Sephadex G-75, dialysis (12–14 kDa) | Gal:Ara:GalA:Xyl:Glu = 46.9:27.5:10.4:0.4:5.4 | 7.6 × 104 | [19] | |
| (24) | GBPP-II | Hot water, purification by dialysis (20 kDa), Sephadex G-75, dialysis (12–14 kDa) | Gal:Ara:GalA:Rha:Glu:Xyl = 26.2:14.2:16.1:11.3:4.8:1.8 | 1.1 × 104 | RG-II and other polysaccharides | [19] |
| (25) | GBPP-III | Hot water, purification by dialysis (20 kDa), Sephadex G-75, dialysis (12–14 kDa) | Gal:Ara:GalA:Rha:Glu:Xyl = 23.4:6.9:13.0:5.5:8.1:7.8 | 2.2 × 103 | Phenolic compounds and sugars, eight different sugars of different composition | [19] |
| (26) | GPNE-I | Hot water, purification on a Sephadex G-100 column, DEAE-Cellulose | Gal:Glc:Ara = 3.7:37.8:1 | 8.03 × 104 | AG-I and AG-II | [20] |
| (27) | GPNE-II | Hot water, purification on a Sephadex G-100 column, DEAE-Cellulose | Gal:Glc:Ara = 1.9:7:1 | 3.15 × 104 | (1→4)-α-d- Glcp backbone with a substitution at O-6 on every two residues. (1→3)-α-d-Glcp and (1→6)-α-d-Glcp | [20] |
| (28) | WGPA-UN-N1 | Hot water, purification on DEAE-Cellulose, Sephadex G-25 column, Sephadex G-50 column | Glc = 97.5 | 1.7 × 104 | α-(1→6)-d-glucan | [73] |
| (29) | WGP | Hot water | Glu:Gal:Ara:GalA:Rha:Man = 76.7:6.5:5.1:9.2:1.4:1.1 | Neutral glucan-like polysaccharides, and contains some amount of RG-I and HG | [69] | |
| (30) | MCGP-L | Hot water, purification by Sephadex G-25 column | Gal:Glc:Man = 1:14.8:1.2 | 3 × 103 | (1→4)-linked-α-d-Glcp residues and with branch chain substituted at O-6 position of (1→4,6)-linked-α-d-Glcp. The branch chain consists of →6)-α-d- Galp-(1→, →2)-α-d-Manp-(1→ and β-d-Glcp-(1→ | [55] |
| (31) | WPPG | Hot water, purification by DEAE-Cellulose column (Microwave-assisted) | Gal:Glu:Rha:Man:GalA:Rib:Arab = 1.56:67.6:1.09:3.75:1:3.42:1 | 2.07 × 106 | [45] | |
| (32) | BGP-60 | Hot water, purification on Sephadex G-75 and DEAE-Cellulose columns | Gal:Glc = 22.3:77 | 2.86 × 104 | α-Dominating configurations | [67] |
| (33) | BGP-65 | Hot water, purification on Sephadex G-75 and DEAE-Cellulose columns | Gal:Glc:Ara = 37.05:59.23:3.72 | 2.67 × 104 | α-Dominating configurations | [67] |
| (34) | BGP-70 | Hot water, purification on Sephadex G-75 and DEAE-Cellulose columns | Gal:Glc:Ara = 43.39:51.43:5.18 | 1.14 × 104 | β-configurations | [67] |
| (35) | BGP-80 | Hot water, purification on Sephadex G-75 and DEAE-Cellulose columns | Gal:Glc:Ara = 41.73:51.61:6.66 | 3.05 × 103 | α-Dominating configurations | [67] |
| (36) | GPS | Hot water | Glu:GalA:Gal:Ara:Rha:Man = 77.9:8.7:6.8:4.6:1.1:1.0 | [27] | ||
| (37) | RG Polysaccharide | Hot water, purification by DEAE-cellulose column, Sepharose CL-6B column | [54] | |||
| (38) | GP II | Hot water, purification by DEAE-Sepharose CL-6B column | 3 × 105 | 60.06% (1→)- or (1→6)-glycosidic linkages and 39.94% (1→3)-glycosidic linkages | [68] | |
| (39) | GP III | Hot water, purification by DEAE-Sepharose CL-6B column | 4 × 105 | 16.23% (1→)- or (1→6)-glycosidic linkages, 25.98% (1→2)-glycosidic linkages, and 57.79% (1→3)-glycosidic linkages | [68] | |
| (40) | GP50-dHR | Hot water, purification by DEAE-Sepharose Fast Flow, dialysis (35 kDa), Sepharose CL-6B column | 5.6 × 104 | HG backbone with hairy regions of RG-I | [70] | |
| (41) | GP50-eHR | Hot water, purification by DEAE-Sepharose, dialysis (35 kDa), Sepharose CL-6B column | 7.7 × 104 | HG backbone with hairy regions of RG-I | [70] | |
| (42) | PGPW1 | Hot water, purification by DEAE Sepharose Fast Flow, Sepharose 6 Fast Flow column | Glc:Gal:Man:Ara = 3.3:1.2:0.5:1.1 | 3.5 × 105 | [36] | |
| (43) | WGFPA-1a | Hot water, purification by DEAE-Cellulose column, Sephacryl S-200 column | GalA:Rha:Gal:Ara:Glc:GlcA:Man = 3.9:3.1:47.2:39.4:2.1:4.0:0.3 | 1.6 × 104 | [34] | |
| (44) | WGFPA-1b | Hot water, purification by DEAE-Cellulose column, Sephacryl S-200 column | GalA:Rha:Gal:Ara:Glc:GlcA:Man:Xyl:Fuc = 56.9:6.0:11.9:17.2:1.7:2.1:1.7:1.7:0.8 | 3.2 × 103 | [34] | |
| (45) | WGFPA-2a | Hot water, purification by DEAE-Cellulose column, Sephacryl S-200 column | GalA:Rha:Gal:Ara:Glc:GlcA:Man = 14.3:9.0:35.3:39.1:0.8:1.3:0.3 | 8.6 × 104 | [34] | |
| (46) | WGFPA-2b | Hot water, purification by DEAE-Cellulose column, Sephacryl S-200 column | GalA:Rha:Gal:Ara:Glc:GlcA:Xyl = 70.3:10.2:7.4:9.4:0.3:0.9:1.6 | 8.7 × 103 | [34] | |
| (47) | WGFPA-3a | Hot water, purification by DEAE-Cellulose column, Sepharose CL-6B column | GalA:Rha:Gal:Ara:Glc:GlcA:Man:Xyl:Fuc = 19.0:13.4:27.5:35.9:0.9:0.7:0.9:1.6:0.1 | 3.0 × 105 | [34] | |
| (48) | WGFPA-3b | Hot water, purification by DEAE-Cellulose column, Sepharose CL-6B column | GalA:Rha:Gal:Ara:Glc:GlcA:Xyl = 82.2:3.6:6.3:5.3:1.2:1.8:0.8 | 2.7 × 104 | [34] | |
| (49) | GPW-1 | Hot water, purification by DEAE-cellulose, Sephadex G-100 column | Fru:Rha:GalA:Glc:Gal:Ara = 0.85:1.40:6.35:3.57:26.33:61.75 | 8.51 × 105 | RG-I-rich | [52] |
| (50) | GPR-1 | Hot water, purification by DEAE-cellulose, Sephadex G-100 column | Fru:Rha:GalA:Glc:Gal:Ara = 0.62:1.92:10.64:12.29:19.96:54.57 | 8.86 × 105 | RG-I-rich | [52] |
| (51) | GPS-1 | Hot water, purification by DEAE-cellulose, Sephadex G-100 column | Fru:Rha:GalA:Glc:Gal:Ara = 2.71:2.12:11.27:13.31:39.92:30.67 | 9.61 × 105 | RG-I-rich | [52] |
| (52) | GPW-2 | Hot water, purification by DEAE-cellulose, Sephadex G-100 column | Fru:Rha:GalA:Glc:Gal:Ara =6.48:3.64:29.12:6.19:22.33:32.24 | 2.95 × 105 | HG-rich | [52] |
| (53) | GPR-2 | Hot water, purification by DEAE-cellulose, Sephadex G-100 column | Fru:Rha:GalA:Glc:Gal:Ara = 2.77:6.85:61.55:1.52:18.76:8.55 | 2.58 × 105 | HG-rich | [52] |
| (54) | GPS-2 | Hot water, purification by DEAE-cellulose, Sephadex G-100 column | Fru:Rha:GalA:Glc:Gal:Ara = 5.57:6.71:68.09:9.36:6.62:3.65 | 3.39 × 105 | HG-rich | [52] |
| (55) | NGP-1a | 50 mM Na2CO3 solution, purification by DEAE-cellulose column, Sepharose CL-6B | GalA:Rha:Gal:Ara = 6.3:4.3:58.8:30.6 | 1.23 × 105 | RG-I | [37] |
| (56) | NGP-2a | 50 mM Na2CO3 solution, purification by DEAE-cellulose column, Sepharose CL-6B | GalA:Rha:Gal:Ara = 19.1:13.2:22.5:45.2 | 1.53 × 105 | RG-I | [37] |
| (57) | NGP-2b | 50 mM Na2CO3 solution, purification by DEAE-cellulose column, Sepharose CL-6B | GalA:Rha:Gal:Ara = 8.7:7.1:60.3:23.9 | 4.9 × 103 | RG-I | [37] |
| (58) | 1-KGP-P1 | 50 mM Na2CO3 solution, 1 M KOH solution, purification by Sepharose CL-6B | Glc:Xyl:Gal:Ara:GalA:Rha = 5.4:81.5:3.8:3.3:3.4:2.5 | 4.8 × 105 | Xylan | [37] |
| (59) | 1-KGP-P2 | 50 mM Na2CO3 solution, 1 M KOH solution, purification by Sepharose CL-6B | Glc:Xyl:Gal:Ara:GalA:Rha = 44.1:19.3:8.0:19.6:3.6:5.4 | 2.7 × 103 | Xylan-xyloglucan | [37] |
| (60) | 4-KGP-P1 | 50 mM Na2CO3 solution, 1 M KOH solution, purification by Sepharose CL-6B | Glc:Xyl:Gal:Ara:GalA:Rha = 4.7:37.6:7.9:8.7:27.9:13.7 | 4.46 × 105 | Xylan-RG-1 | [37] |
| (61) | 4-KGP-P2 | 50 mM Na2CO3 solution, 4 M KOH solution, purification by Sepharose CL-6B | Glc:Xyl:Gal:Ara:GalA = 39.1:26.9:17.7:8.3:1.9 | 1.23 × 105 | Xyloglucan | [37] |
| (62) | 4-KGP-P3 | 50 mM Na2CO3 solution, 4 M KOH solution, purification by Sepharose CL-6B | Glc:Xyl:Gal:Ara = 56.8:12.9:10.6:19.7 | 2.6 × 103 | Xyloglucan | [37] |
| (63) | RG-CW-CP | Hot water, dialysis (12–14 kDa) | Ara:Fuc:Xyl:Rha:Man:Glc:Gal = 7.6:1.84:0.53:1.66:0.37:41.15:6.23:40.62 | [41] | ||
| (64) | RG-HW-CP | Hot water, dialysis (12–14 kDa) | Ara:Fuc:Xyl:Rha:Glc:Gal = 7.26:1.15:0.14:1.64:52.84:5.49:31.48 | [41] | ||
| (65) | RG-CW-EZ-CP | Hot water, dialysis (12–14 kDa), α-amylase and amyloglucosidase | Ara:Fuc:Rha:Man:Glc:Gal = 15.94:0.16:3.94:1.13:5.06:13.97:59.81 | [41] | ||
| (66) | RG-HW-EZ-CP | Hot water, dialysis (12–14 kDa), α-amylase and amyloglucosidase | Ara:Fuc:Rha:Glc:Gal = 16.88:3.00:3.44:3.48:13.69:59.51 | [41] | ||
| (67) | PG-F2 | Hot water and RNase and DNase, purification by Q-Sepharose fast-flow column | Gal:Ara:Rha:GalA:GlcA:Xyl:Fuc = 1.6:1.8:2.3:76.6:16.9:0.1:0.6 | 1.2 × 104 | [74] | |
| (68) | WGFPN | Hot water, purification by dialysis, DEAE-Cellulose column | Gal:Ara:Glc:Man = 78:14.3:5.2:2.5 | 1.1 × 104 | (1→4)-β-d-galactan and highly branched (1→6)-β-d-galactan, (1→6)-β-d-galactan, α-l-Araf through O-3 | [73] |
| (69) | KRG-P | Hot water, purification by dialysis (20 kDa) | Glu:GalA:Gal:Ara:Rha = 60.5:19.7:11.0:6.8:1.9 | 1.06 × 105 | [72] | |
| (70) | RGP-AP-I | Hot water, α-amylase, purification on a Sephadex G-100 column, dialysis (14 kDa) | Rha:Fuc:Ara:Xyl:Man:Gal:Glu:GalA:GluA = 9.5:0.1:35.0:0.3:0.4:30.4:1.7:18.4:1.5 | 9.6 × 104 | RG-I | [22] |
| (71) | RGP-AP-II | EtOH, α-amylase, purification on a Sephadex G-100 column, dialysis (14 kDa) | Rha:Fuc:Ara:Xyl:Man:Gal:Glu:GalA:GluA = 9.3:0.3:25.4:0.3:0.5:19.9:1.2:38.6:1.2 | [22] | ||
| (72) | RGP-AP-III | EtOH, α-amylase, purification on a Sephadex G-100 column, dialysis (14 kDa) | Rha:Fuc:Ara:Xyl:Man:Gal:Glu:GalA:GluA = 6.8:0.8:19.6:0.3:0.5:24.6:2.6:38.9:1.4 | [22] | ||
| (73) | RGP-AP-IV | EtOH, α-amylase, purification on a Sephadex G-100 column, dialysis (14 kDa) | Rha:Fuc:Ara:Xyl:Man:Gal:Glu:GalA:GluA = 1.4:0.5:4.2:0.3:0.5:10.5:4.1:74.4:0.7 | [22] | ||
| (74) | FGWP | Hot water | Gal:Glc:Ara:Rha:Fuc = 9.16:60.59:10.11:1.81:0.14 | 6.5 × 106 | [40] | |
| (75) | FGEP-C | Hot water, and cellulase | Gal:Glc:Ara:Rha:Fuc = 10.39:62.24:10.54:2.13:0.14 | [40] | ||
| (76) | FGEP-A | Hot water, and α-amylase | Gal:Glc:Ara:Rha:Fuc = 12.6:50.86:10.89:1.76:0.17 | [40] | ||
| (77) | FGEP-CA | Hot water, cellulase and α-amylase co-treatment | Gal:Glc:Ara:Rha:Fuc = 16.53:45.83:11.43:3.22:0.20 | 6.4 × 106 | [40] | |
| (78) | EGP-N | Hot water, α-amylase and EDTA, purification by DEAE-Cellulose column | Gal:Ara:Glc = 3.9:4.6:88.7 | Starch-like glucan, with minor AG | [42] | |
| (79) | EGP-1a | Hot water, α-amylase and EDTA, purification by DEAE-Cellulose column, Sepharose CL-6B column | Glc = 100 | 4.5 × 105 | Starch-like glucan | [42] |
| (80) | EGP-1b | Hot water, α-amylase and EDTA, purification by DEAE-Cellulose column, Sepharose CL-6B column | GalA:Rha:Gal:Ara:Glc = 2.3:1.1:11.2:9.9:75.5 | 6.2 × 103 | More glucan with minor RG-I pectin | [42] |
| (81) | EGP-2a | Hot water, α-amylase and EDTA, purification by DEAE-Cellulose column, Sepharose CL-6B column | GalA:Rha:Gal:Ara:Glc = 32.7:8.0:27.6:27.7:3.4 | 4.2 × 105 | HG and RG-I pectin | [42] |
| (82) | EGP-2b | Hot water, α-amylase and EDTA, purification by DEAE-Cellulose column, Sepharose CL-6B column | GalA:Rha:Gal:Ara:Glc = 46.5:7.0:20.7:21.9:3.9 | 1.5 × 105 | HG and RG-I pectin | [42] |
| (83) | EGP-3a | Hot water, α-amylase and EDTA, purification by DEAE-Cellulose column, Sepharose CL-6B column | GalA:Rha:Gal:Ara:Glc = 52.8:8.7:16.9:17.7:3.9 | 4.3 × 105 | HG and RG-I pectin | [42] |
| (84) | EGP-3b | Hot water, α-amylase and EDTA, purification by DEAE-Cellulose column, Sepharose CL-6B column | GalA:Rha:Gal:Ara:Glc = 64.5:7.4:12.5:9.5:6.1 | 1.1 × 105 | HG and RG-I pectin | [42] |
| (85) | RG-I-3A-2 | Enzyme-assisted, purification by DEAE-Cellulose, dialysis, Sepharose CL-6B column | Gal:Glc:Ara:Rha:Man:GalA:GlcA = 20.3:1.7:37.2:1.7:37.8:2.4 | 2.15 × 104 | [60] | |
| (86) | RG-I-3A-4 | Enzyme-assisted, purification by DEAE-Cellulose, dialysis, Sepharose CL-6B column | Gal:Glc:Ara:Rha:Man:GalA:GlcA = 14.8:1.9:39.6:0.9:40.2:2.5 | 7.2 × 103 | [60] | |
| (87) | RG-I-3A-6 | Enzyme-assisted, purification by DEAE-Cellulose, dialysis, Sepharose CL-6B column | Gal:Glc:Ara:Rha:Man:GalA:GlcA = 13.3:2.2:40.1:0.3:41.2:2.9 | 6.3 × 103 | [60] | |
| (88) | RG-I-3A-16 | Enzyme-assisted, purification by DEAE-Cellulose, dialysis, Sepharose CL-6B column | Gal:Rha:Man:GalA:GlcA = 5.7:44.8:1.9:45.6:1.9 | 6.0 × 103 | →4)-α-GalpA-(1→ and →2) -α-Rhap-(1→ | [60] |
| (89) | RG-I-2 | Enzyme-assisted, purification on a Sephadex G-25 column, dialysis | Gal:Ara:Rha:GalA = 12.4:14.5:1.7:44.3 | 4 × 103 | RG-I backbone with 1.5 side chains | [71] |
| (90) | RG-I-3B | Enzyme-assisted, purification on a Sephadex G-25 column, dialysis | Gal:Ara:Rha:GalA = 13.4:11.9:14.1:44.6 | 6 × 103 | RG-I backbone with 4 side chains | [71] |
| (91) | RG-I-4 | Enzyme-assisted, purification on a Sephadex G-25 column, dialysis | Gal:Ara:Rha:GalA = 19.5:9.2:21.8:33.8 | 6 × 104 | RG-I backbone, Ara, and Gal side chains | [71] |
| (92) | WGPE-N | 4000 U α-amylase, dialysis (3.5 kDa), purification by DEAE-Cellulose | Gal:Glc:Ara = 2.3:94.8:2.9 | Starch-like glucans | [39] | |
| (93) | WGPE-1a | 4000 U α-amylase, dialysis (3.5 kDa), purification by DEAE-Cellulose, Sephadex G-75 column | Gal:Glc:Ara:Rha = 30.9:44.3:21.9:2.1 | 1.1 × 104 | AG chains | [39] |
| (94) | WGPE-1b | 4000 U α-amylase, dialysis (3.5 kDa), purification by DEAE-Cellulose, Sephadex G-75 column | Gal:Glc:Ara:Rha:GalA = 8.3:72.9:0.8:1.9:2.8 | 5.5 × 103 | More glucan and less AG | [39] |
| (95) | WGPE-2a | 4000 U α-amylase, dialysis (3.5 kDa), purification by DEAE-Cellulose, Sepharose CL-6B column | Gal:Glc:Ara:Rha:GalA =36.5:1.2:39.7:8.3:13.7 | 4.3 × 106 | RG-I pectin branched with α→1,5/1,3,5-arabinan and β-1,4-galactan side chains | [39] |
| (96) | WGPE-2b | 4000 U α-amylase, dialysis (3.5 kDa), purification by DEAE-Cellulose, Sepharose CL-6B column | Gal:Glc:Ara:Rha:GalA = 14.1:2.9:16.1:4.6:62.2 | 1.2 × 105 | HG pectin with 4-O-methyl-α-d- GalA | [39] |
| (97) | WGPE-3a | 4000 U α-amylase, dialysis (3.5 kDa), purification by DEAE-Cellulose, Sepharose CL-6B column | Gal:Ara:Rha:GalA =23.0:30.7:11.4:34.9 | 4.2 × 106 | RG-I pectin branched with α- 1,5/1,3,5-arabinan and β-1,4-galactan side chains | [39] |
| (98) | WGPE-3b | 4000 U α-amylase, dialysis (3.5 kDa), purification by DEAE-Cellulose, Sepharose CL-6B column | Gal:Glc:Ara:Rha:GalA =5.2:4.1:5.6:3.4:81.7 | 5.0 × 104 | HG pectin with 4-O-methyl-α-d-GalA | [39] |
| (99) | WGPA-P2A | Hot water, hydrolyzed by Endo-PG, purification by Sephadex G-25, DEAE-Sepharose Fast Flow, Sephadex G-75 | Gal:Glc:Ara:Rha:Man:GalA:GlcA = 41.1:1.7:30.1:11.5:1.3:12.2:2.1 | 7.8 × 104 | RG-I | [75] |
| (100) | WGPA-P2B | Hot water, hydrolyzed by Endo-PG, purification by Sephadex G-25, DEAE-Sepharose Fast Flow, Sephadex G-75 | Gal:Glc:Ara:Rha:Man:GalA:GlcA = 11.4:4:20.3:13.4:0.6:47.9:2.4 | 3.7 × 103 | RG-II | [75] |
| (101) | RG-I-1 | Hot water, hydrolyzed by Endo-PG, purification by Sephadex G-25 column, DEAE-Sepharose Fast Flow column, Sephadex G-75 | GalA:Rha:Gal:Ara:Man:Glc:GlcA:Fuc = 26.8:12.8:21.2:13.0:5.9:7.2:7.4:2.0 | 5 × 103 | [76] | |
| (102) | RG-I-2 | Hot water, hydrolyzed by Endo-PG, purification by Sephadex G-25 column, DEAE-Sepharose Fast Flow column, Sephadex G-75 | GalA:Rha:Gal:Ara:Man:Glc:GlcA:Fuc = 44.3:11.7:12.4:14.5:1.0:4.4:5.8:3.8 | 4 × 103 | 4-O-Me-b-d-GlcAp at the non-reducing terminals of branched chains | [76] |
| (103) | RG-I-3A | Hot water, hydrolyzed by Endo-PG, purification by Sephadex G-25 column, DEAE-Sepharose Fast Flow column, Sepharose CL-6B | GalA:Rha:Gal:Ara:Man:Glc:GlcA:Fuc = 32.2:11.1:31.6:16.3:2.1:1.9:3.0:0.7 | 4.5 × 104 | AG-I | [76] |
| (104) | RG-I-3B | Hot water, hydrolyzed by Endo-PG, purification by Sephadex G-25 column, DEAE-Sepharose Fast Flow column, Sepharose CL-6B | GalA:Rha:Gal:Ara:Man:Glc:GlcA:Fuc = 44.6:14.1:13.7:11.9:1.3:2.5:3.7:2.3 | 6 × 103 | AG-I | [76] |
| (105) | RG-I-4 | Hot water, hydrolyzed by Endo-PG, purification by Sephadex G-25 column, DEAE-Sepharose Fast Flow column, Sepharose CL-6B | GalA:Rha:Gal:Ara:Man:Glc:GlcA:Fuc = 33.8:21.8:19.5:9.2:0.4:3.0:2.2:1.5 | 6 × 104 | AG-I | [76] |
| (106) | MPPG | Microwave, purification by DEAE-Cellulose column | Gal:Glu:Rha:Man:GalA:Rib:GluA:Arab = 2.7:141.42:1.85:3.94:1.43:4.55:1:5.15 | 3.69 × 106 | [45] | |
| (107) | G1 | Microwave assisted purification (1100 W), separation by ultrafiltration (3 kDa) | Gal:Glc:Ara:Rha:Man:GalA = 4.37:3.95:5.16:0.37:0.17:3.9 | 3.5 × 103–1.1 × 105 | [61] | |
| (108) | G2 | Microwave assisted purification (1100 W), separation by ultrafiltration (3 kDa) | Gal:Glc:Ara:Rha:Man:GalA = 4.53:4.12:5.03:0.79:0.57:3.85 | [61] | ||
| (109) | G3 | Microwave assisted purification (1100 W), separation by ultrafiltration (3 kDa) | Gal:Glc:Ara:Rha:Man:GalA = 4.33:4.02:5.11:0.75:0.95:4.06 | [61] | ||
| (110) | G4 | Microwave assisted purification (1100 W), separation by ultrafiltration (3 kDa) | Gal:Glc:Ara:Rha:Man:GalA = 4.62:4.34:5.1:0.65:0.65:3.9 | [61] | ||
| (111) | G5 | Microwave assisted purification (1100 W), separation by ultrafiltration (3 kDa) | Gal:Glc:Ara:Rha:Man:GalA = 4.62:4.46:5.35:0.71:0.92:4.47 | [61] | ||
| (112) | PPG | Ultrasonication extract, dialysis (3.5 kDa) | Ara:Rha:GalA:Man:Glc:Gal = 28:3:1:4:70:44 | 1.499 × 106, 5.335 × 104 | 1→4 glycosidic linkages as linear backbone | [30] |
| No. | Polysaccharide Name | Extraction and Purification | Composition Ratio of Sugars (Mass Percentage or Molar Ratio) | Molecular Weight (Da) | Structure Feature | Refs |
|---|---|---|---|---|---|---|
| (1) | WPS-1 | Hot water, purification on DEAE-Sepharose CL-6B and Sepharose CL-6B | Gal:Glc:Ara:Rha:Man = 18.7:55.2:21.2:2.3:2.6 | 1.54 × 106 | (1→6)-α-d-Glcp, (1→5)-α-l-Araf and (1→4)- β-d-Rhap | [57] |
| (2) | WPS-2 | Hot water, purification on DEAE-Sepharose CL-6B and Sepharose CL-6B | Gal:Glc:Ara:Rha:Man = 20.7:46.8:27.9:1.7:2.9 | 1.41 × 104 | (1→6)-α-d-Glcp and (1→5)-α-l-Araf | [57] |
| (3) | SPS-1 | Hot water, purification on DEAE-Sepharose CL-6B and Sepharose CL-6B | Gal:Glc:Ara:Man:GalA:GlcA:Xyl = 28.6:15.9:22.3:9.2:13.6:3.5:6.9 | 3.62 ×105 | (1→6)-α-d-Glcp, (1→4)-α-d-Manp, (1→5)-α-l-Araf, β-d-Galp and β-d-xylose | [57] |
| (4) | SPS-2 | Hot water, purification on DEAE-Sepharose CL-6B and Sepharose CL-6B | Gal:Glc:Ara:Man:GalA:GlcA:Xyl = 22.5:25.3:14.2:7.9:16.9:7.9:5.3 | 9.70 × 106 | (1→6)-α-d-Glcp, (1→4)-α-d-Manp, (1→5)-α-l-Araf, β-d-Galp, β-d-xylose and O-acetyl group | [57] |
| (5) | SPS-3 | Hot water, purification on DEAE-Sepharose CL-6B and Sepharose CL-6B | Gal:Glc:Ara:Rha:Man:GalA:Xyl = 15.2:11.5:19.2:2.1:12:26.3:9.6 | 5.12 × 105 | (1→6)-α-d-Glcp, (1→4)-α-d-Manp, (1→5)-α-l-Araf, β-d-Galp, β-d-xylose, O-acetyl group and (1→4)-β-d-Rhap | [57] |
| (6) | GSP | Hot water, purification by dialysis (12 kDa) | Rha:Ara:Gal:Glc = 1:4:8:8 | 8.5 × 104 | The main sugar residues were 4-α-d-GalpA and 4-α-d-GalpA, other residues such as 2-α-l-Rhap, 2,4-α-l-Rhap, α-l-Araf, β-d-Galp, 4-β-d-Galp | [77] |
| (7) | quinquefolans A | Hot water, purification by DEAE-Toyopearl, Sephacryl S-200 and S-50 | Glc:Man = 2.3:1 | >2 × 103 | [83] | |
| (8) | quinquefolans B | Hot water, purification by DEAE-Toyopearl, Sephacryl S-200 and S-50 | Glc:Man = 5.5:1 | >2 × 103 | [83] | |
| (9) | quinquefolans C | Hot water, purification by DEAE-Toyopearl, Sephacryl S-200 and S-50 | Xyl = 1 | >2 × 103 | [83] | |
| (10) | PPQN | Hot water, purification by DEAE Sepharose Fast Flow column | Gal:Glc = 1.15:1 | 3.1×103 | [80] | |
| (11) | PPQA2 | Hot water, purification by DEAE Sepharose and Sephacryl S-300 | Gal:Glc:Ara:Rha:Man:GalA:GlcA = 7.2:12.5:8:4:2.9:26.6:38.8 | 2.3 ×104 | O-acetyl groups and β-arabinopyranosyl residue | [51] |
| (12) | PPQA4 | Hot water, purification by DEAE Sepharose and Sephacryl S-300 | Gal:Glc:Ara:Rha:Man:GlcA = 23.9:41.3:19.7:5.1:8.1:2 | 1.2 × 105 | β-arabinopyranosyl residue | [51] |
| (13) | PPQA5 | Hot water, purification by DEAE Sepharose and Sephacryl S-300 | Gal:Glc:Ara:Rha:Man:GalA:GlcA = 10.8:32.4:8.5:3.2:5.3:15.5:24.4 | 5.3×103 | O-acetyl groups and β-arabinopyranosyl residue | [51] |
| (14) | AGC1 | Hot water, purification by DEAE-Sepharose column | Gal:Glc:Ara:Xyl:Glu:Rha:Man = 60:6.29:19.2:11.4:1.5:0.8 | 5.2 × 103 | 3-Galp (48.5%), 3,6-Galp (10.2%), t-Galp (5.2%), 6-Galp (4.4%), 4-Glcp (5.7%), 4-Arap/5-Araf (4.0%) and t-Araf (4.5%) | [78] |
| (15) | AGC3 | Hot water, purification by DEAE-Sepharose column | Gal:Glc:Ara:Rha:Man:GlcA = 74.3:92:7.8:8.1:6.8:1 | 4.81 × 103, 3.2 ×104 | RG-I pectin, t-Galp (47.7%), 4-Galp (15.6%), 2,4-Rhap (8.1%), 6-Galp (8.1%) and 4-GalAp (6.8%) | [79] |
| (16) | PPQ | Hot water, purification by DEAE Sepharose Fast Flow column, Sephacryl S-200 High Resolution column | Glc:Gal = 2.1:1 | 5.4 × 104 | [81] | |
| (17) | GPS | Hot water | 1.092 × 106 | [82] | ||
| (18) | AEP-1 | 0.3 M NaOH, purification by DEAE Sepharose Fast Flow column, Sephacryl S-300 High Resolution column | Gal:Glc:GalA = 0.97:4.67:3.92 | d-GalpA | [38] | |
| (19) | AEP-2 | 0.3 M NaOH, purification by DEAE Sepharose Fast Flow column, Sephacryl S-300 High Resolution column | Gal:Glc:Ara:Man:GalA = 1.68:3.02:1.03:0.76:3.65 | [38] | ||
| (20) | Q1 | Microwave assisted purification (1.1 kW), separation by ultrafiltration (3 kDa) | Rha:Ara:GalA:Man:Glc:Gal = 0.39:3.06:2.16:0.14:2.95:2.36:11.1 | 8.54 × 104 | [61] | |
| (21) | Q2 | Microwave assisted purification (1.1 kW), separation by ultrafiltration (3 kDa) | Rha:Ara:GalA:Man:Glc:Gal = 0.46:3.16:2.29:0.13:3.13:2.47:11.6 | [61] | ||
| (22) | Q3 | Microwave assisted purification (1.1 kW), separation by ultrafiltration (3 kDa) | Rha:Ara:GalA:Man:Glc:Gal = 0.48:3.18:2.10:0.09:3.02:2.47:11.3 | [61] | ||
| (23) | Q4 | Microwave assisted purification (1.1 kW), separation by ultrafiltration (3 kDa) | Rha:Ara:GalA:Man:Glc:Gal = 0.45:3.23:2.19:0.09:2.97:2.40:11.3 | [61] | ||
| (24) | Q5 | Microwave assisted purification (1.1 kW), separation by ultrafiltration (3 kDa) | Rha:Ara:GalA:Man:Glc:Gal = 0.49:3.09:2.12:0.10:3.06:2.43:11.3 | [61] | ||
| (25) | PPQ | Ultrasonication extract, dialysis (3.5 kDa) | Ara:Rha:GalA:Man:Glc:Gal = 31:4:1:2:72:59 | 3.626 × 106, 2.520 × 105, 5.356 × 104 | 1→4 glycosidic linkages as linear backbone | [30] |
| No. | Polysaccharide Name | Extraction and Purification | Composition Ratio of Sugars (Mass Percentage or Molar Ratio) | Molecular Weight (Da) | Structure Feature | Refs |
|---|---|---|---|---|---|---|
| (1) | Sanchinan-A | MeOH and hot water, purification by Sephadex G-50 column | 1.5 × 106 | β-d-(1→3)-linked galactopyranosyl residues | [84] | |
| (2) | PF3111 | Hot water, purification by Sephadex G-50 column, Sephadex-DEAE A-50 column twice | Gal:Glc:Ara:Man = 3.5:10.8:1:3.5 | 6.85 × 105 | [56] | |
| (3) | PF3111 | Hot water, purification by Sephadex G-50 column, Sephadex-DEAE A-50 column twice | Gal:Glc:Ara:Man = 2.9:5.3:1:2.8 | 3.7 × 104 | [56] | |
| (4) | PBGA11 | 0.01 M NaOH, purification by Sephadex G-50 column, Sephadex-DEAE A-50 column twice | Gal:Glc:Ara:Man = 3.1:4.2:1:5.3 | 4.5 × 104 | [56] | |
| (5) | PBGA12 | 0.01 M NaOH, purification by Sephadex G-50 column, Sephadex-DEAE A-50 column twice | Gal:Glc:Ara:Man = 2.5:7.2:1:4.3 | 7.6 × 105 | [56] | |
| (6) | AIR | Sonicated with MeOH | Gal:Glc:Ara:GalA = 11:75:3:11 | 4-Glcp (72 mol%) | [50] | |
| (7) | Fr1MKOH | 1 M KOH, DMSO, dialysis (3.5 kDa) | 4-galactan, heteroxylan, starch; 2,4-Rhap, 4-Xylp, 4-Galp, terminal Glcp, 4-GalAp | [58] | ||
| (8) | 1MD3-G2 | 1 M KOH, DMSO, dialysis (3.5 kDa), DEAE-Sepharose CL-6B column | 1.14 × 106 | Glucuronoarabinoxylan, HG, RG-I, 4-galactan, arabinan | [58] | |
| (9) | Arabinoglucogalactan (1) | Hot water, purification by DEAE- cellulose column twice, Sephadex G-100 column, Sephacryl S-300 column | Ara:Glu:Gal = 1:1:8 | 6.7 × 104 | a backbone of (1→3)-linked β-d-galactofuranosyl residues, with branches of α-l-Araf-(1→4)-β-d-Glcp-(1→residues at O-6 | [85] |
| (10) | MRP | Hot water | Gal:Glc:Ara:Rha:Man = 16.9:28:3.2:4.2:47.4 | [26] | ||
| (11) | BRP | Hot water | Gal:Glc:Ara:Man = 26.9:17.9:1.7:10.4 | [26] | ||
| (12) | FRP | Hot water | Gal:Glc:Ara:Man = 10.3:49.3:1.7:22.2 | [26] | ||
| (13) | RN1 | Hot water, purification by DEAE-cellulose column | Gal:Ara = 43.7:56.3 | 2.1 × 104 | A backbone of 1,6-linked Galp branched at C3 by side 1,3-linked Galp, with branches attached at position O-3,1,5-linked, 1,3,5 linked, terminal Ara and terminal Gal | [86] |
| (14) | N1 | Microwave assisted purification (1.1 kW), separation by ultrafiltration (3 kDa) | Rha:Ara:GalA:Man:Glc:Gal = 0.14:2.83:0.63:0.17:7.28:2.67:13.7 | [61] | ||
| (15) | N2 | Microwave assisted purification (1.1 kW), separation by ultrafiltration (3 kDa) | Rha:Ara:GalA:Man:Glc:Gal = 0.11:2.83:0.56:0.13:7.64:2.64:13.9 | [61] | ||
| (16) | N3 | Microwave assisted purification (1.1 kW), separation by ultrafiltration (3 kDa) | Rha:Ara:GalA:Man:Glc:Gal = 0.14:3.03:0.47:0.23:7.72:2.69:14.3 | [61] | ||
| (17) | N4 | Microwave assisted purification (1.1 kW), separation by ultrafiltration (3 kDa) | Rha:Ara:GalA:Man:Glc:Gal = 0.12:3.03:0.40:0.17:7.54:2.64:13.9 | [61] | ||
| (18) | N5 | Microwave assisted purification (1.1 kW), separation by ultrafiltration (3 kDa) | Rha:Ara:GalA:Man:Glc:Gal = 0.14:3.02:0.57:0.13:8.25:2.84:14.9 | [61] | ||
| (19) | MRP5A | Hot water, purification by DEAE-52 Cellulosee, Sephadex G-100 column | Gal:Glu:Ara:Rha = 15.9:69.8:6.6:7.7 | 1.138 × 105 | [87] | |
| (20) | MRP5 | Hot water, purification by DEAE-52 Cellulosee, Sephadex G-100 column | Gal:Glu:Ara:Rha = 29.7:51.2:14.5:3.6 | 9.16 × 104 | →3)-β-Glcp-(1→, →3)-β-Galp-(1→, →3, 6)- β-Galp-(1→, →3)-β-Galp-(1→, →3, 6)-β-Galp-(1→, →3)-α- Rhap-(1→, →3)-α-Araf-(1→, and α-Araf-(1→ residues | [87] |
| (21) | PNPS-0.5M | Hot water, purification by DEAE-FAST-FLOW Cellulose column | Glc:Ara:Rha:Man = 22.5:4.5:3.8:0.2 | 2.6 × 106 | (1→)-linked Araf, (1→) linked Rhap, (5→1)-linked Araf, (4→1)-linked GalA, and (3,6→1)-linked Galp | [43] |
| (22) | PNPS-0.3 | Hot water, purification by DEAE-52 cellulose, Sephadex G-50 | Gal:Glc:Ara:Rha:GalA = 33.3:4.5:25.2:15.5:17.1 | 7.6655 × 104 | A backbone of →4)-α-d-GalAp-(1→4-β-l-Rhap-1→4)-β-d-Galp-(1→residues, with an α-l-Araf-1→5)-α-l-Araf-(1→branch connecting to the backbone at O-3 of →4-β-l-Rhap-1→ | [32] |
| (23) | PNPN | Hot water, purification by DEAE-Cellulose column, Sepharose CL-6B column | Gal:Glc:Ara:Rha:Man = 12.3:82.9:2.3:1.3:1.2 | [29] | ||
| (24) | PNPA-1A | Hot water, purification by DEAE-Cellulose column, Sepharose CL-6B column | Gal:Glc:Ara:Rha:Man:GalA = 63.2:2.4:27.7:0.8:0.9:5 | 8.8 × 104 | 1,4-β-d-galactan and 1,5-α-l-arabinan, small amounts of AG-II, and minor RG-I domains | [29] |
| (25) | PNPA-2A | Hot water, purification by DEAE-Cellulose column, Sepharose CL-6B column | Gal:Ara:Rha:Man:GalA:GlcA = 46:33.4:6:2:11.6:1 | 1.01 × 105 | 1,4-β-d-galactan and 1,5-α-l-arabinan, small amounts of AG-II, and minor RG-I domains | [29] |
| (26) | PNPA-3A | Hot water, purification by DEAE-Cellulose column, Sepharose CL-6B column | Gal:Glc:Ara:Rha:Man:GalA = 32.7:2.2:28.3:15.5:4:15.9 | 2.70 × 105 | Typical RG-I type pectin with 1,4-β-d-galactan and 1,5/1,3,5-α-l-arabinan side chains | [29] |
| (27) | PNPA-1B | Hot water, purification by DEAE-Cellulose column, Sepharose CL-6B column | Gal:Glc:Ara:Rha:Man:GalA:GlcA = 29.3:4.5:10.4:9.6:2.9:40.6:0.6 | 3 × 103 | HG, RG-I and RG-II | [29] |
| (28) | PNPA-2B | Hot water, purification by DEAE-Cellulose column, Sepharose CL-6B column | Gal:Ara:Rha:Man:GalA:GlcA = 8.3:8.2:7.5:0.8:74.4:0.8 | 6 × 103 | HG, RG-I and RG-II | [29] |
| (29) | PNPA-3B | Hot water, purification by DEAE-Cellulose column, Sepharose CL-6B column | Gal:Glc:Ara:Rha:Man:GalA:GlcA = 8.8:1.6:5.1:5.2:1.4:75.8:0.9 | 1.8 × 104 | HG, RG-I and RG-II | [29] |
| (30) | PPN | Ultrasonication extract, dialysis (3.5 kDa) | Ara:GalA:Man:Glc:Gal = 2:1:2:83:7 | 6.014 × 105, 5.065 × 104 | 1→4 glycosidic linkages as linear backbone | [30] |
| (31) | NPPN | Hot water, purification by DEAE Sepharose-Fast Flow column, dialysis | 1.81 × 104 | [88] | ||
| (32) | APPN I | Hot water, purification by DEAE Sepharose-Fast Flow column, dialysis | 7.047 × 104 | [88] | ||
| (33) | APPN II | Hot water, purification by DEAE Sepharose-Fast Flow column, dialysis | 1.056 × 105 | [88] | ||
| (34) | APPN III | Hot water, purification by DEAE Sepharose-Fast Flow column, dialysis | 9.233 × 104 | [88] | ||
| (35) | APPN IV | Hot water, purification by DEAE Sepharose-Fast Flow column, dialysis | 9.017 × 104 | [88] | ||
| (36) | FPNP | Hot water, purification by DEAE- Sepharose Fast Flow | [2] |
| Polysaccharides | Extraction Method | Purification | M.W. (kDa) | Monosaccharide Compositions | Structure Features |
|---|---|---|---|---|---|
| ginseng | Hot water | DEAE-Cellulose Sepharose CL-6B Sephadex G-25 Sephadex G-50 Sephadex G-75 Sephadex G-100 Sephacryl S-200 Dialysis (3.5 kDa) Ultrafiltration (3 kDa) | 0.1–2070 | Ara, Arab, Fru, Fuc, Gal, GalA, Glc, GlcA, Man, Rha, Rib, Xyl | AG-I, AG-II, HG, RG-I, RG-II, and starch-like glucans |
| Alkaline | 2.6–480 | Ara, Gal, GalA, Glc, Xyl | RG-I, amyloses, xyloglucan, and starch-like glucan | ||
| Enzyme | 4–4300 | Ara, Fuc, Gal, GalA, Glc, GlcA, Man, Rha, Xyl | AG-I, AG, HG, RG-I, and starch-like glucans | ||
| EDTA | 6.2–450 | Ara, Gal, Glc, GalA, Rha | AG, HG, RG-I, and starch-like glucan | ||
| Microwave Ultrasonication | 3.5–3690 53.35–1499 | Ara, Arab, Gal, GalA, Glc, GlcA, Man, Rha, Rib Ara, Gal, GalA, Glc, Man, Rha | 1→4 glycosidic linkages as linear backbone | ||
| American ginseng | Hot water | DEAE Sepharose Fast Flow Sephacryl S-200 Sephacryl S-300 Sepharose CL-6B Ultrafiltration (3 kDa) Dialysis (3.5 kDa) | 3.1–9700 | Ara, Gal, GalA, Glc, GlcA, Man, Rha, Xyl | RG-I, (1→6)-α-d-Glcp, (1→4)-α-d-Manp, (1→5)-α-l-Araf, β→-Galp, β-d-xylose, O-acetyl group, β-arabinopyranosyl residue |
| Alkaline | Ara, Gal, GalA, Glc, Man | d-GalpA | |||
| Microwave | 85.4 | Ara, Gal, GalA, Glc, Man, Rha | |||
| Ultrasonication | 53.56–3626 | Ara, Gal, GalA, Glc, Man, Rha | 1→4 glycosidic linkages as linear backbone | ||
| notoginseng | Hot water | Sephadex G-50 Sephadex-DEAE A-50 DEAE-Sepharose CL-6B Ultrafiltration (3 kDa) Dialysis (3.5 kDa) | 3–2600 | Ara, Gal, GalA, Glc, GlcA, Man, Rha | AG-II, HG, RG-I, RG-II, (4→1)-linked GalA, 1,4-β-d-galactan and 1,5-α-l-arabinan |
| Alkaline | 45–1140 | Ara, Gal, Glc, Man | HG, RG-I, glucuronoarabinoxylan, 4-galactan, arabinan, heteroxylan, starch; 2,4-Rhap, 4-Xylp, 4-Galp, terminal Glcp, 4-GalAp | ||
| MeOH | 45–1500 | Ara, Gal, GalA, Glc | β-d-(1→3)-linked galactopyranosyl residues, 4-Glcp | ||
| Microwave | Ara, Gal, GalA, Glc, Man, Rha, | ||||
| Ultrasonication | 50.65–601.4 | Ara, Gal, GalA, Glc, Man | 1→4 glycosidic linkages as linear backbone |
| Activities/Polysaccharides | Models | Molecular Mechanisms | Refs | |
|---|---|---|---|---|
| Anti-cancer effect Ginseng polysaccharides | ||||
| (1) | GPS | Mice macrophage, K562, HL-60, or KG1 cells | Possesses a potent antitumor activity by stimulating TNF-α, IL-1, IL-6, and NO production in macrophage against leukemia | [89] |
| (2) | GPS | MDA-MB-231 breast cancer cells | Activates p65-IKZF1 signaling and apoptosis to inhibit cell proliferation | [90] |
| (3) | GBPE, GBPP | HCT-116 and HT-29 human colon cancer cells and CD4+ IFN-γ+ cells | Inhibits IL-8 secretion and CD4+ IFN-γ+ cell (Th1) and CD4+ FoxP3+ cell (Treg) differentiation to suppress cancer cell growth | [91] |
| (4) | GBPP | B16-BL6 melanoma xenograft mice model | Increases IL-6, IL-12, TNF-α, IFN-γ, and granzyme B of NK cells to suppress tumor colonies | [18] |
| (5) | NFP | B16F10 melanoma xenograft mice model | Inhibits melanoma cell metastasis to the lung by enhancing immunity | [92] |
| (6) | GS-P | Colon 26-M3 cells and xenograft mice model | Promotes the macrophages and NK cells to exhibit anti-metastatic activity | [93] |
| (7) | WGPN | S 180 tumor-bearing mice | Stimulates lymphocytes proliferation and macrophage activity and mitigates the damage of immune system induced by 5-fluorouracil | [94] |
| (8) | WGPA, WGPA-3-HG | HT-29 colon cancer cells | Inhibits cell proliferation and cell cycle arrest in the G2/M phase | [95] |
| (9) | MWGPA-3-HG | HT-29 colon cancer cells | Increases the percentages of S and G2/M phase cells and induces apoptosis by activating caspase-3 | [95] |
| (10) | PGPW1 | HGC-27 gastric cancer cells | Decreases migration and invasion by regulation of Twist, AKR1C2 and NF1 to mediate epithelial–mesenchymal transition | [36] |
| (11) | GP | Lewis lung carcinoma xenograft mice model | Increases the ratio of CD4+/CD8+ T lymphocyte in peripheral blood and promotes NK cytolytic activity | [97] |
| (12) | GFP1 | Lewis lung carcinoma xenograft mice model | Promotes ConA or LPS-induced spleen lymphocytes proliferation and elevates NK cell activity and the ratio of CD4+/CD8+ to inhibit tumor growth and metastasis | [96] |
| (13) | GPS | 96 patients with non-small cell lung cancer | Increases Th1 cytokines (INF-γ, IL-2) and decreases Th2 cytokines (IL-4, IL-5) | [98] |
| (14) | GPs | Lewis lung carcinoma xenograft mice model | Alters gut microbiota and kynurenine/tryptophan ratio to potentiate antitumor effect of anti-PD-1/PD-L1 immunotherapy | [99] |
| American ginseng polysaccharides | ||||
| (1) | GPS NPs | UVB-induced SKH1 hairless mice | Inhibits the pro-inflammatory cytokine levels, epidermal proliferation, and skin cancer | [82] |
| Notoginseng polysaccharides | ||||
| (1) | RN1 | HMEC-1 microvascular endothelial cells, BxPC3 pancreatic cancer xenograft mice model | Inhibits BMP2/Smad1/5/8/Id1 signaling to inhibit tumor angiogenesis | [86] |
| (2) | PPN | H22 cells and tumor-bearing mice | Activates CD4+ T-cells and elevates serum IL-2 to inhibit tumor growth and prolong the survival | [100] |
| (3) | NPPN | H22 cells and tumor-bearing mice | Inhibits tumor growth via myelosuppression after combination with CTX | [88] |
| Immunomodulatory effect | ||||
| Ginseng polysaccharides | ||||
| (1) | FGWP-CA | CTX-induced immunosuppressed mice | Increases spleen and thymus indices, lymphocyte proliferation, NK cell activity, leukocyte counts, and IL-6, IL-12, and TNF-α levels | [40] |
| (2) | GPS | CTX-induced mice | Increases NK cell number and upregulates the levels of perforin and granzyme to activate NK cells | [101] |
| (3) | APG | Autoimmune encephalomyelitis mice | Promotes Treg cell generation through Foxp3 activation and inhibits the production of inflammatory cytokines, including IFN-γ, IL-1β, and IL-17 | [102] |
| (4) | APG | Autoimmune encephalomyelitis-induced SJL/J mice | Modulates the infiltration of CD4+ T cells and CD11b+ macrophages into the spinal cord and decreases the amounts of IFN-γ, IL-17, and TNF-α | [103] |
| (5) | GPNE-I | ConA or LPS-induced lymphocyte cells | Increases T and B lymphocyte proliferation | [20] |
| (6) | WGPA-2-RG | Normal mice spleen lymphocytes and LPS-induced peritoneal macrophages | Decreases lymphocyte proliferation and macrophage nitrite production and slightly increases macrophage phagocytosis | [64] |
| (7) | Ginsan | Dendritic cells | Increases the levels of IL-16 and TNF-α and the expression of CD86 | [23] |
| (8) | PS-NPs | RAW 264.7 macrophages | Increases the levels of TNF-α, IL-1β, and IL-6 and NO production | [104] |
| (9) | RGAP | RAW 264.7 macrophages | Enhances NO production and increases nuclear transcription factors by activating the ERK/JNK and NF-κB/AP-1 signaling pathways | [49] |
| (10) | RGAP | BALB/c mice spleen cells | Promotes spleen cell proliferation and NO production | [105] |
| (11) | RGAP | BALB/c mice spleen cells and intraperitoneal macrophages | Increases the numbers of IgM antibody-forming cells, T cells, B cells, and macrophages | [106] |
| (12) | RGAP | Immunosuppressed mice | Increases plaque-forming cells in the spleen in response to LPS and sheep red blood cells | [107] |
| (13) | RGP-AP-I | BALB/c mic peritoneal macrophages | Augments the production of IL-6, IL-12, and TNF-α | [22] |
| (14) | RG-CW-EZ-CP | BALB/c mice intraperitoneal macrophages and Peyer’s patch cells from small intestines of C3H/He mice | Enhances intestinal immune system and macrophage activity by upregulating the phosphorylation of ERK, JNK, and p38 | [41] |
| (15) | NGP | Murine bone marrow dendritic cells | Increases the expressions of MHC II, CD80, CD86, CD83, and CD40 and decreases TNF-α levels | [108] |
| (16) | PGP-SL | Murine spleen lymphocytes | Regulates the Ca2+/calcineurin/NFAT/IL-2 signaling pathway | [109] |
| (17) | GMP | Murine peritoneal macrophages | Increases the production of reactive oxygen intermediates | [110] |
| (18) | Gps | LPS-induced weaned piglets | Regulates the TLR4/MyD88-NF-κB pathway to reduce immunological stress | [111] |
| American ginseng polysaccharides | ||||
| (1) | High molecular weight polysaccharides | Human peripheral blood mononuclear cells | Triggers the ERK, PI3K, p38, and NF-κB signaling pathways | [112] |
| (2) | AGRPS | Rat alveolar macrophage | Increases NO production and TNF-α concentration | [113] |
| (3) | WPS-1, WPS-2, SPS-1, SPS-2, SPS-3 | Mice lymphocytes and macrophages | Increases macrophage phagocytosis, NO production, and splenic lymphocyte proliferation | [57] |
| (4) | AGP | CTX-induced mice | Enhances CD4+ T cells and IgA-secreting cells and regulates gut microbiota | [114] |
| (5) | AGP | RAW 264.7 macrophages | Up-regulates the production of NO and cytokines | [115] |
| (6) | AGC1 | RAW 264.7 macrophages and primary murine splenocytes | Enhances NO, TNF-α, IL-6, MCP-1, and GM-CSF levels to increase cell proliferation via the NF-κB signaling pathway | [78] |
| (7) | AGC3 | RAW 264.7 macrophages and primary murine splenocytes | Enhances IL-6, TNF-α, GM-CSF, and MCP-1 levels via the NF-κB (p65/RelA) and p38 signaling pathways | [79] |
| (8) | PPQA2, PPQA4, PPQA5 | RAW 264.7 macrophages | Increases NO, TNF-α, and IL-6 production | [51] |
| (9) | PPQN | RAW 264.7 macrophages | Inhibits the production of TNF-α, IL-1β, and IL-6 | [80] |
| (10) | GPS NPs | RAW 264.7 macrophages and Swiss albino mice | Increases NO, TNF-α, IL-1β, and IL-6 | [116] |
| Notoginseng polysaccharides | ||||
| (1) | PBGA12 | Mice peritoneal macrophages | Enhances IFN-γ and TNF-α to stimulate complement system | [56] |
| (2) | Fr1MKOH | Human polymorphonuclear neutrophils | Shows complement-fixing activity and mitogenic effect through regulation of ROS and IFN-γ | [58] |
| (3) | PNPS-0.3 | Bone marrow dendritic cells | Increases the amounts of TNF-α and IL-12 and induces T-cell immune response (CD4, CD8, CD69, and MHC II) by triggering the TLR4/TLR2-NF-κB signaling pathway | [32] |
| Anti-oxidative activity | ||||
| Ginseng polysaccharides | ||||
| (1) | Native ginseng polysaccharides | In vitro | Scavenges DPPH, hydroxyl, or superoxide anion radicals | [24] |
| (2) | Ginseng polysaccharides | In vitro | Scavenges DPPH free radicals | [25] |
| (3) | Ginseng-SDF | In vitro | Decreases DPPH, ABTs, and ferric ion radicals | [117] |
| (4) | WGAP, WGNP | Caenorhabditis elegans | Scavenges hydroxyl radicals and reduces ROS and lipid peroxidation | [31] |
| (5) | WGPN, WGPA | STZ-induced diabetic mice | Decreases MDA level and increases SOD activity | [35] |
| (6) | WGPA | Forced swim test/ICR mice | Inhibits MDA and LDH levels and causes the increases of SOD and GSH-Px activities | [118] |
| (7) | WGPA, WGPA-A | Forced swim test/ICR mice | Lowers MDA level and enhances GSH-Px activity | [59] |
| American ginseng polysaccharides | ||||
| (1) | AEP-2 | RAW 264.7 macrophages | Shows higher values of Trolox equivalent and oxygen radical antioxidant capacities | [38] |
| Notoginseng polysaccharides | ||||
| (1) | FPNP | H2O2-induced human dermal fibroblast | Decreases ROS and MDA and increases the activities of CAT, GSH-Px, and SOD by activating the TGF-β/Smad signaling pathway | [2] |
| Other bioactive functions | ||||
| Ginseng polysaccharides | ||||
| (1) | GPS | C57BL/6 mice model | Promotes hypothalamic neuropeptide Y expression and inhibits the levels of proopiomelanocortin and dopamine D1 receptor in the midbrain to promote food intake | [119] |
| (2) | GPS | Ethanol-induced gastric injured rats | Inhibits oxidative stress (increased SOD and CAT and decreased MDA) and inflammation (reduced TNF-α, IL-6, IL-1β, and MPO) by inhibiting the NF-κB and STAT pathways | [27] |
| (3) | WGP | Antibiotic-associated diarrhea mice | Changes gut microbiota composition and diversity, balances metabolic processes to recover mucosal structure | [69] |
| (4) | WGP | Cisplatin-induced mice | Regulates the PI3K/AKT, PERK-eIF2α-ATF4, and NF-κB p65 signaling pathways to prevent endoplasmic reticulum stress, inflammatory response, and apoptotic cell death | [120] |
| (5) | GP | Diabetic rat model | Enhances Rb1 biotransformation by gut microbiota and promotes the fecal β-d-glucosidase activity | [121] |
| (6) | GP | Dextran sulphate sodium-induced colitis rat model | Enhances microbial deglycosylation and Rb1 intestinal absorption | [122] |
| (7) | APG | γ-rays irradiation-induced C57BL/6 mice model | Inhibits the p53-dependent and mitochondrial apoptosis pathways to protect small intestine | [123] |
| (8) | GP | H1N1 influenza virus infected BALB/c mice | Lowers lung viral titers and IL-6 to improve survival | [124] |
| (9) | AP1 | Hypoxia/reoxygenation-induced H9c2 cells | Increases glucocorticoid receptor and estrogen receptor to activate the reperfusion injury salvage kinase pathway and regulates the eNOS-dependent pathways to maintain endothelial function | [125] |
| (10) | WGPA | ICR mice model | Increases social interactions and decreases aggressive behaviors | [65] |
| (11) | ELHPP-RGBPs | Dermatophagoides farinae extracts-induced NC/Nga mice | Inhibits the AP-1/MMP-1 pathway to prevent solar ultraviolet-induced skin wrinkles and atopic dermatitis | [126] |
| Notoginseng polysaccharides | ||||
| (1) | PNPS | Ischemia/reperfusion injured rat model | Regulates Bcl-2/Bax ratio and caspase-3 cascade to suppress apoptosis | [127] |
| (2) | PNPS-0.5M | Alcoholic liver damage mice model | Regulates the alcohol dehydrogenase and catalase pathways to prevent peroxide accumulation | [43] |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Qi, H.; Zhang, Z.; Liu, J.; Chen, Z.; Huang, Q.; Li, J.; Chen, J.; Wang, M.; Zhao, D.; Wang, Z.; et al. Comparisons of Isolation Methods, Structural Features, and Bioactivities of the Polysaccharides from Three Common Panax Species: A Review of Recent Progress. Molecules 2021, 26, 4997. https://doi.org/10.3390/molecules26164997
Qi H, Zhang Z, Liu J, Chen Z, Huang Q, Li J, Chen J, Wang M, Zhao D, Wang Z, et al. Comparisons of Isolation Methods, Structural Features, and Bioactivities of the Polysaccharides from Three Common Panax Species: A Review of Recent Progress. Molecules. 2021; 26(16):4997. https://doi.org/10.3390/molecules26164997
Chicago/Turabian StyleQi, Hongyu, Zepeng Zhang, Jiaqi Liu, Zhaoqiang Chen, Qingxia Huang, Jing Li, Jinjin Chen, Mingxing Wang, Daqing Zhao, Zeyu Wang, and et al. 2021. "Comparisons of Isolation Methods, Structural Features, and Bioactivities of the Polysaccharides from Three Common Panax Species: A Review of Recent Progress" Molecules 26, no. 16: 4997. https://doi.org/10.3390/molecules26164997