
Optimization of Fermentation Process of Pomegranate Peel and Schisandra Chinensis and the Biological Activities of Fermentation Broth: Antioxidant Activity and Protective Effect Against H2O2-induced Oxidative Damage in HaCaT Cells

Abstract
:1. Introduction
2. Results and Discussion
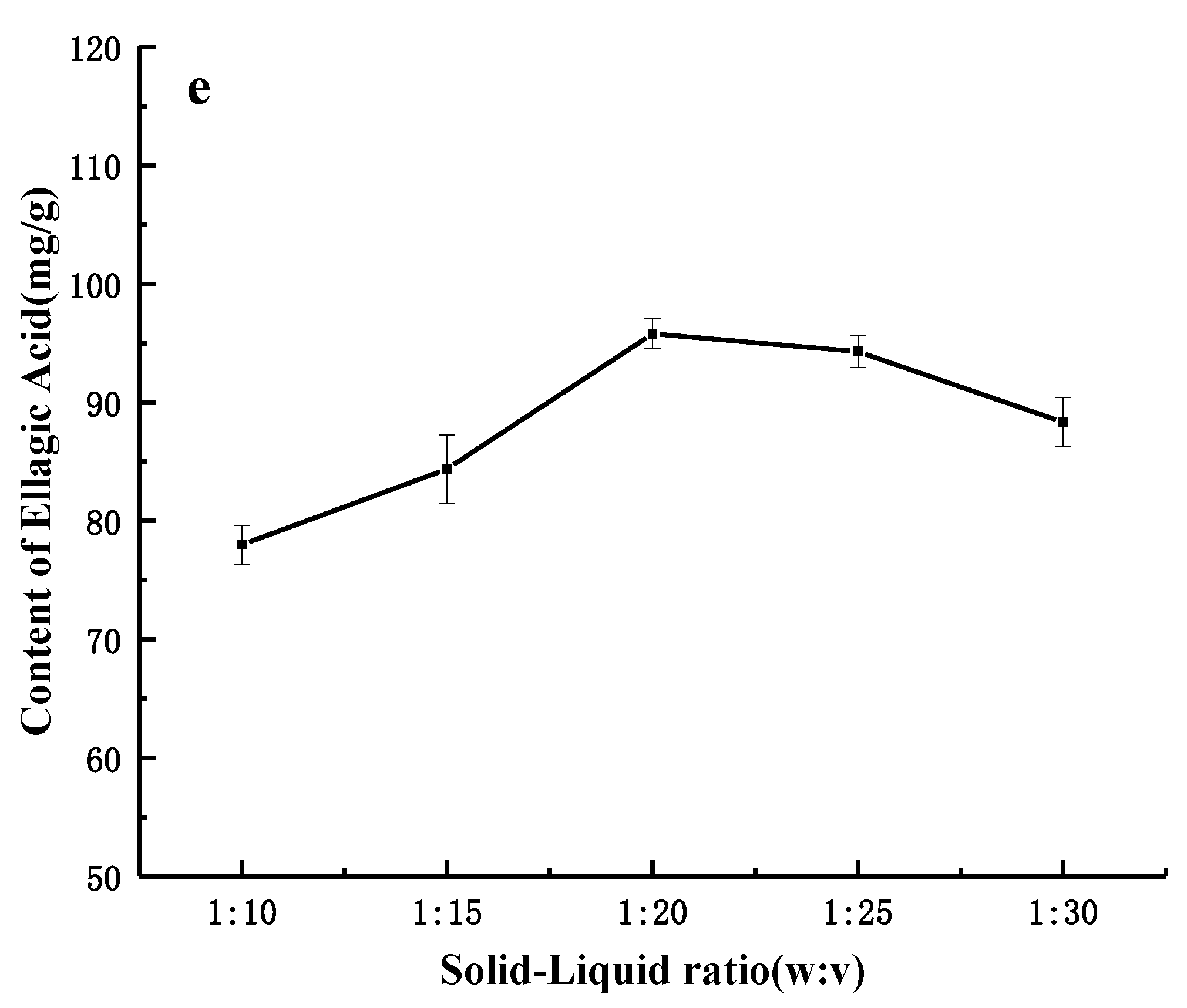
2.1. Single-Factor Experiments
2.2. Building Models and Analyzing Statistics
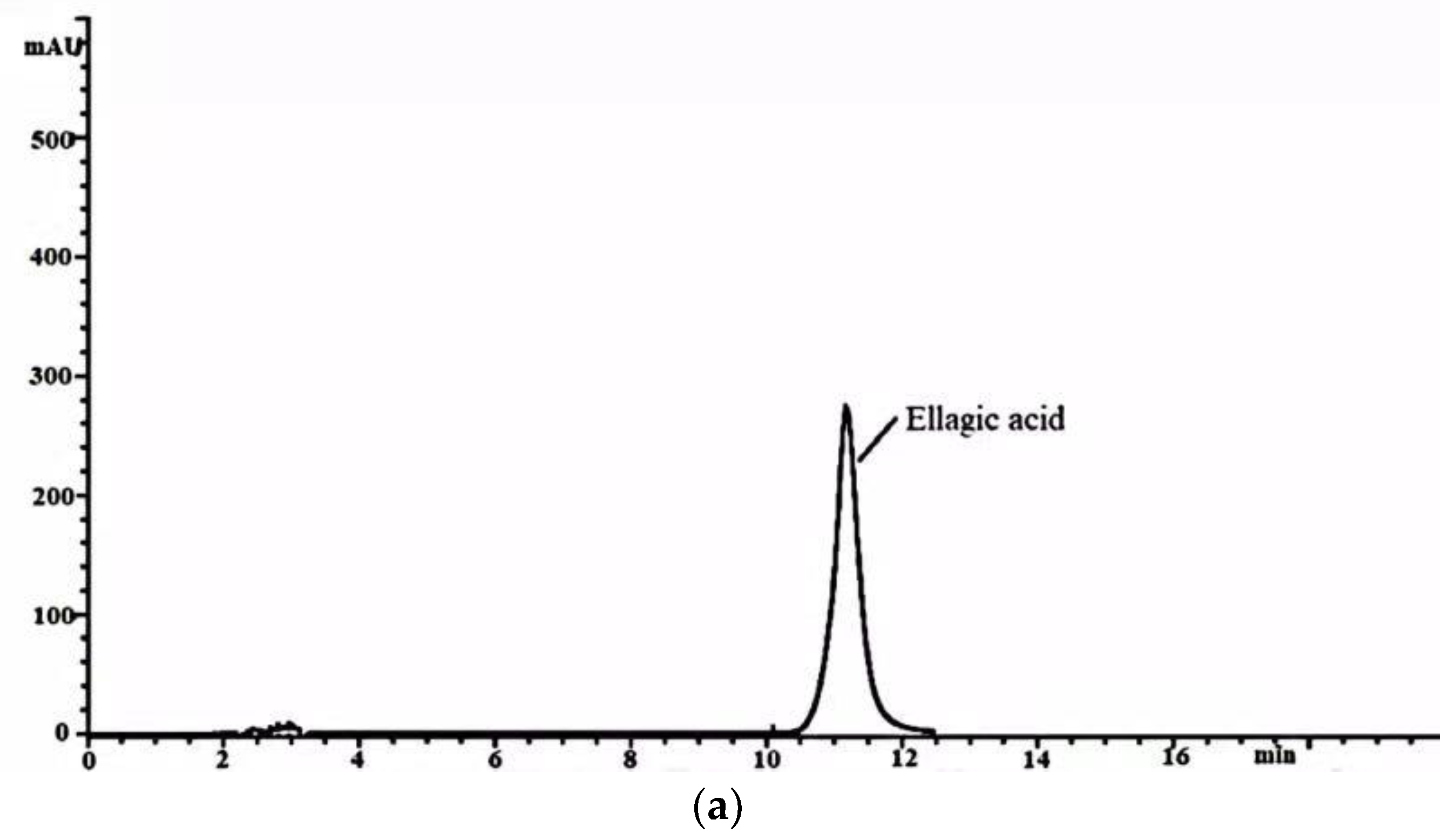
2.3. HPLC Analysis of Ellagic Acid in the FB and WE
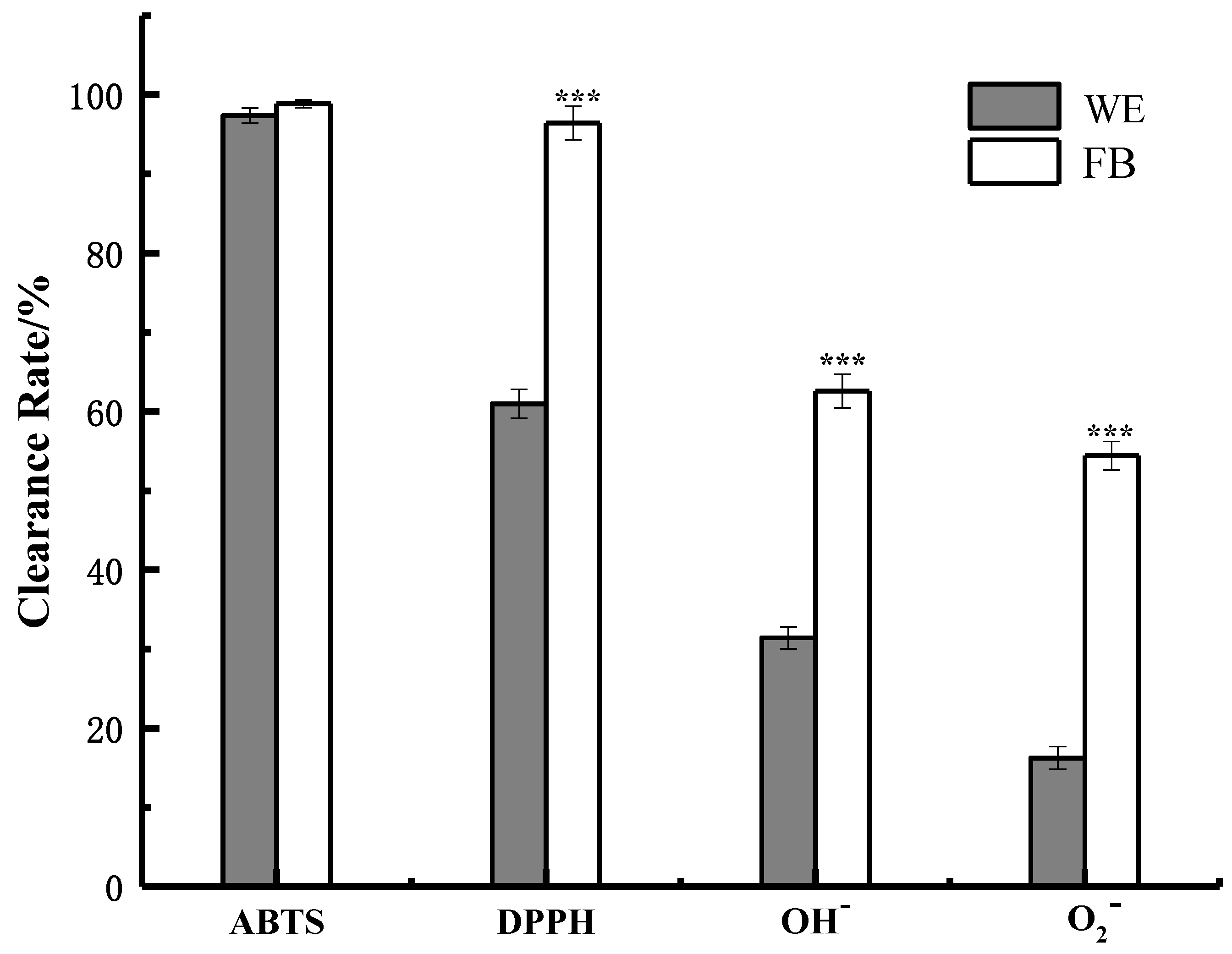
2.4. Antioxidant Ability of WE and FB
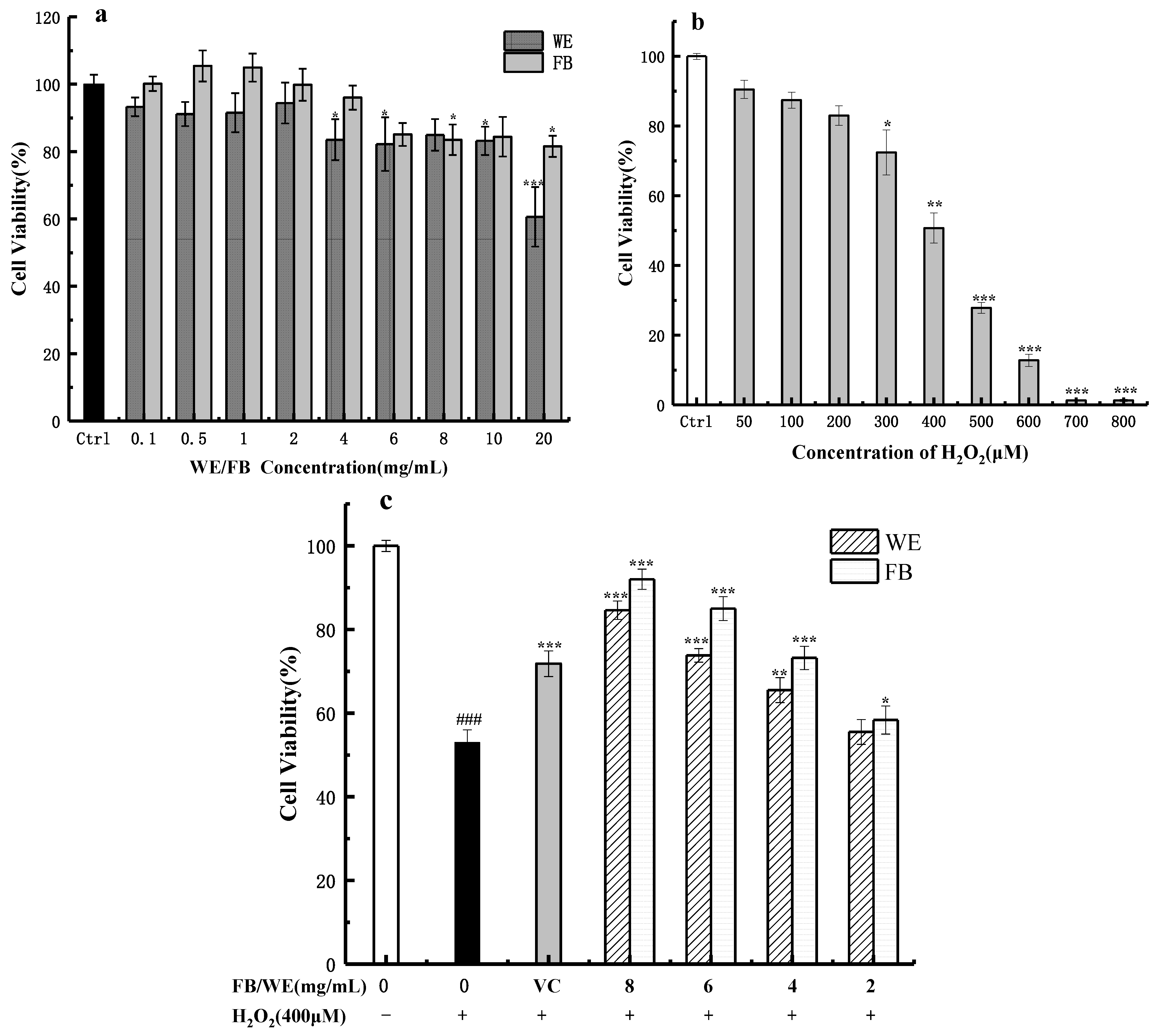
2.5. Effects of FB, WE and H2O2 on the Activity of HaCaT Cells
2.6. Effect of FB and WE on ROS Level in HaCaT Cells Treated with H2O2
3. Materials and Methods
3.1. Experimental Materials and Chemical Reagents
3.2. Preparation of FB
3.3. Determination of Ellagic Acid in the FB
3.4. Determination of Antioxidant Activity
3.5. Characterization of Ellagic Acid by HPLC
3.6. Effects of FB, WE and H2O2 on the Activity of HaCaT Cells
3.7. Effects of FB and WE on the ROS Level in HaCaT Cells Treated with H2O2
3.8. Experimental Design and Statistical Analysis
4. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
Sample Availability
References
- Castro-Giráldez, M.; Fito, P.; Ortolá, M.D.O.; Balaguer, N. Study of pomegranate ripening by dielectric spectroscopy. Postharvest Biol. Technol. 2013, 86, 346–353. [Google Scholar] [CrossRef]
- Li, X.; Liu, L.; Pischetsrieder, M. Pomegranate (Punica granatum L.) wine polyphenols affect Nrf2 activation and antioxidant enzyme expression in human neuroblastoma cells (SH-SY5Y). J. Funct. Foods 2017, 38, 140–150. [Google Scholar] [CrossRef]
- Yoshime, L.T.; De Melo, I.L.P.; Sattler, J.A.G.; Torres, R.P.; Mancini-Filho, J. Bioactive compounds and the antioxidant capacities of seed oils from pomegranate (Punica granatum L.) and bitter gourd (Momordica charantia L.). Food Sci. Technol. 2019, 39, 571–580. [Google Scholar] [CrossRef] [Green Version]
- Ambigaipalan, P.; de Camargo, A.C.; Shahidi, F. Phenolic Compounds of Pomegranate Byproducts (Outer Skin, Mesocarp, Divider Membrane) and Their Antioxidant Activities. J. Agric. Food Chem. 2016, 64, 6584–6604. [Google Scholar] [CrossRef]
- Kang, S.; Choi, B.; Kim, S.; Yi, H.; Park, H.; Park, S.; Song, C.; Park, J.; Lee, Y.; Ku, S. Inhibitory effects of Pomegranate concentrated solution on the activities of Hyaluronidase, Tyrosinase, and Metalloproteinase 1. Clin. Ther. 2015, 37, e118. [Google Scholar] [CrossRef]
- Ordoudi, S.A.; Mantzouridou, F.; Daftsiou, E.; Malo, C.; Hatzidimitriou, E.; Nenadis, N.; Tsimidou, M. Pomegranate juice functional constituents after alcoholic and acetic acid fermentation. J. Funct. Foods 2014, 8, 161–168. [Google Scholar] [CrossRef]
- Natalello, A.; Hervás, G.; Toral, P.G.; Luciano, G.; Valenti, B.; Mendoza, A.G.; Pauselli, M.; Priolo, A.; Frutos, P. Bioactive compounds from pomegranate by-products increase the in vitro ruminal accumulation of potentially health promoting fatty acids. Anim. Feed. Sci. Technol. 2020, 259, 114355. [Google Scholar] [CrossRef]
- Lekshmi, R.; Nisha, S.A.; Kaleeswaran, B.; Alfarhan, A. Pomegranate peel is a low-cost substrate for the production of tannase by Bacillus velezensis TA3 under solid state fermentation. J. King Saud Univ. Sci. 2020, 32, 1831–1837. [Google Scholar] [CrossRef]
- Giamogante, F.; Marrocco, I.; Cervoni, L.; Eufemi, M.; Chichiarelli, S.; Altieri, F. Punicalagin, an active pomegranate component, is a new inhibitor of PDIA3 reductase activity. Biochimie 2018, 147, 122–129. [Google Scholar] [CrossRef]
- Wang, P.; Zhang, Q.; Hou, H.; Liu, Z.; Wang, L.; Rasekhmagham, R.; Kord-Varkaneh, H.; Santos, H.O.; Yao, G. The effects of pomegranate supplementation on biomarkers of inflammation and endothelial dysfunction: A meta-analysis and systematic review. Complement. Ther. Med. 2020, 49, 102358. [Google Scholar] [CrossRef]
- Amor, A.J.; Gómez-Guerrero, C.; Ortega, E.; Sala-Vila, A.; Lázaro, I. Ellagic Acid as a Tool to Limit the Diabetes Burden: Updated Evidence. Antioxidants 2020, 9, 1226. [Google Scholar] [CrossRef] [PubMed]
- Szopa, A.; Klimek-Szczykutowicz, M.; Kokotkiewicz, A.; Dziurka, M.; Luczkiewicz, M.; Ekiert, H. Phenolic acid and flavonoid production in agar, agitated and bioreactor-grown microshoot cultures of Schisandra chinensis cv. Sadova No. 1—A valuable medicinal plant. J. Biotechnol. 2019, 305, 61–70. [Google Scholar] [CrossRef]
- Sobstyl, E.; Szopa, A.; Ekiert, H.; Gnat, S.; Typek, R.; Choma, I.M. Effect directed analysis and TLC screening of Schisandra chinensis fruits. J. Chromatogr. A 2020, 1618, 460942. [Google Scholar] [CrossRef] [PubMed]
- Zhai, J.; Zhang, F.; Gao, S.; Chen, L.; Feng, G.; Yin, J.; Chen, W. Schisandra chinensis extract decreases chloroacetaldehyde production in rats and attenuates cyclophosphamide toxicity in liver, kidney and brain. J. Ethnopharmacol. 2018, 210, 223–231. [Google Scholar] [CrossRef] [PubMed]
- Song, Y.; Shan, B.; Zeng, S.; Zhang, J.; Jin, C.; Liao, Z.; Wang, T.; Zeng, Q.; He, H.; Wei, F.; et al. Raw and wine processed Schisandra chinensis attenuate anxiety like behavior via modulating gut microbiota and lipid metabolism pathway. J. Ethnopharmacol. 2021, 266, 113426. [Google Scholar] [CrossRef]
- Bae, S.J.; Kim, J.E.; Choi, Y.J.; Lee, S.J.; Gong, J.E.; Choi, Y.-W.; Hwang, D.Y. Novel Function of α-Cubebenoate Derived from Schisandra chinensis as Lipogenesis Inhibitor, Lipolysis Stimulator and Inflammasome Suppressor. Molecules 2020, 25, 4995. [Google Scholar] [CrossRef]
- Lee, S.; Chun, J.; Lee, H.-J.; Park, H.; So, I.; Jeon, J.-H.; Park, E.-J. Transcriptome Analysis of the Anti-TGFβ Effect of Schisandra chinensis Fruit Extract and Schisandrin B in A7r5 Vascular Smooth Muscle Cells. Life 2021, 11, 163. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Y.-H.; Zhao, Z.-Y.; Wang, B.-J.; Zhang, Y.-Q.; Zhang, M.; Gao, Y.-Y. Protective effect of Schisandra chinensis lignans on hypoxia-induced PC12 cells and signal transduction. J. Toxicol. Environ. Health Part A 2018, 81, 1224–1230. [Google Scholar] [CrossRef]
- Mapelli-Brahm, P.; Barba, F.J.; Remize, F.; Garcia, C.; Fessard, A.; Khaneghah, A.M.; Sant’Ana, A.S.; Lorenzo, J.M.; Montesano, D.; Meléndez-Martínez, A.J. The impact of fermentation processes on the production, retention and bioavailability of carotenoids: An overview. Trends Food Sci. Technol. 2020, 99, 389–401. [Google Scholar] [CrossRef]
- Palmieri, M.G.S.; Cruz, L.T.; Bertges, F.S.; Húngaro, H.M.; Batista, L.R.; da Silva, S.S.; Fonseca, M.J.V.; Rodarte, M.P.; Vilela, F.M.P.; Amaral, M.D.P.H.D. Enhancement of antioxidant properties from green coffee as promising ingredient for food and cosmetic industries. Biocatal. Agric. Biotechnol. 2018, 16, 43–48. [Google Scholar] [CrossRef]
- Xu, N.; Marchionni, K.; Hu, Y.; Zhang, W.; Sosic, Z. Quantitative analysis of a biopharmaceutical protein in cell culture samples using automated capillary electrophoresis (CE) western blot. J. Pharm. Biomed. Anal. 2017, 145, 10–15. [Google Scholar] [CrossRef]
- Aslani, A.; Mazzuca-Sobczuk, T.; Eivazi, S.; Bekhrad, K. Analysis of bioenergy technologies development based on life cycle and adaptation trends. Renew. Energy 2018, 127, 1076–1086. [Google Scholar] [CrossRef]
- Balwani, I.; Chakravarty, K.; Gaur, S. Role of phytase producing microorganisms towards agricultural sustainability. Biocatal. Agric. Biotechnol. 2017, 12, 23–29. [Google Scholar] [CrossRef]
- Viji, R.; Shrinithivihahshini, N.D.; Santhanam, P.; Balakrishnan, S.; Yi, Y.; Rajivgandhi, G. Biomonitoring of the environmental indicator and pathogenic microorganisms assortment in foremost pilgrimage beaches of the Bay of Bengal, Southeast coast, India. Mar. Pollut. Bull. 2019, 149, 110548. [Google Scholar] [CrossRef]
- Szutowska, J. Functional properties of lactic acid bacteria in fermented fruit and vegetable juices: A systematic literature review. Eur. Food Res. Technol. 2020, 246, 357–372. [Google Scholar] [CrossRef]
- Verotta, L.; Panzella, L.; Antenucci, S.; Calvenzani, V.; Tomay, F.; Petroni, K.; Caneva, E.; Napolitano, A. Fermented pomegranate wastes as sustainable source of ellagic acid: Antioxidant properties, anti-inflammatory action, and controlled release under simulated digestion conditions. Food Chem. 2018, 246, 129–136. [Google Scholar] [CrossRef]
- Wang, C.; Zhang, J.; Hu, F.; Zhang, S.; Lu, J.; Liu, S. Bio-pretreatment promote hydrolysis and acidification of oilseed rape straw: Roles of fermentation broth and micro-oxygen. Bioresour. Technol. 2020, 308, 123272. [Google Scholar] [CrossRef] [PubMed]
- Walters, N.A.; de Villiers, A.; Joubert, E.; de Beer, D. Improved HPLC method for rooibos phenolics targeting changes due to fermentation. J. Food Compos. Anal. 2017, 55, 20–29. [Google Scholar] [CrossRef]
- Medjekal, S.; Bodas, R.; Bousseboua, H.; López, S. Evaluation of three medicinal plants for methane production potential, fiber digestion and rumen fermentation in vitro. Energy Procedia 2017, 119, 632–641. [Google Scholar] [CrossRef]
- Morone, J.; Alfeus, A.; Vasconcelos, V.; Martins, R. Revealing the potential of cyanobacteria in cosmetics and cosmeceuticals—A new bioactive approach. Algal Res. 2019, 41, 101541. [Google Scholar] [CrossRef]
- Rashki, M.; Azarkish, H.; Rostamian, M.; Bahrpeyma, A. Classification correction of polynomial response surface methods for accurate reliability estimation. Struct. Saf. 2019, 81, 101869. [Google Scholar] [CrossRef]
- Yu, M.; Wang, B.; Qi, Z.; Xin, G.; Li, W. Response surface method was used to optimize the ultrasonic assisted extraction of flavonoids from Crinum asiaticum. Saudi J. Biol. Sci. 2019, 26, 2079–2084. [Google Scholar] [CrossRef] [PubMed]
- Logarušić, M.; Slivac, I.; Radošević, K.; Bagović, M.; Redovnikovic, I.R.; Srček, V.G. Hempseed protein hydrolysates’ effects on the proliferation and induced oxidative stress in normal and cancer cell lines. Mol. Biol. Rep. 2019, 46, 6079–6085. [Google Scholar] [CrossRef] [PubMed]
- Shin, D.; Lee, Y.; Huang, Y.-H.; Lim, H.-W.; Jang, K.; Kim, D.-D.; Lim, C.-J. Probiotic fermentation augments the skin anti-photoaging properties of Agastache rugosa through up-regulating antioxidant components in UV-B-irradiated HaCaT keratinocytes. BMC Complement. Altern. Med. 2018, 18, 196. [Google Scholar] [CrossRef]
- Xiao, X.; He, Y.; Li, C.; Zhang, X.; Xu, H.; Wang, B. Nicastrin mutations in familial acne inversa impact keratinocyte proliferation and differentiation through the Notch and phosphoinositide 3-kinase/AKT signalling pathways. Br. J. Dermatol. 2016, 174, 522–532. [Google Scholar] [CrossRef]
- Choi, M.; Park, M.; Lee, S.; Lee, J.W.; Choi, W.J.; Lee, C. Establishment of Nrf2-deficient HaCaT and immortalized primary human foreskin keratinocytes and characterization of their responses to ROS-induced cytotoxicity. Toxicol. Vitr. 2019, 61, 104602. [Google Scholar] [CrossRef] [PubMed]
- Gao, J.; Guo, J.; Nong, Y.; Mo, W.; Fang, H.; Mi, J.; Qi, Q.; Yang, M. 18β-Glycyrrhetinic acid induces human HaCaT keratinocytes apoptosis through ROS-mediated PI3K-Akt signaling pathway and ameliorates IMQ-induced psoriasis-like skin lesions in mice. BMC Pharmacol. Toxicol. 2020, 21, 1–11. [Google Scholar] [CrossRef]
- Masci, A.; Coccia, A.; Lendaro, E.; Mosca, L.; Paolicelli, P.; Cesa, S. Evaluation of different extraction methods from pomegranate whole fruit or peels and the antioxidant and antiproliferative activity of the polyphenolic fraction. Food Chem. 2016, 202, 59–69. [Google Scholar] [CrossRef]
- Tabaraki, R.; Heidarizadi, E. Simultaneous biosorption of Arsenic (III) and Arsenic (V): Application of multiple response optimizations. Ecotoxicol. Environ. Saf. 2018, 166, 35–41. [Google Scholar] [CrossRef]
- Milessi, T.S.; Aquino, P.M.; Silva, C.R.; Moraes, G.S.; Zangirolami, T.C.; Giordano, R.C.; Giordano, R.L. Influence of key variables on the simultaneous isomerization and fermentation (SIF) of xylose by a native Saccharomyces cerevisiae strain co-encapsulated with xylose isomerase for 2G ethanol production. Biomass. Bioenergy 2018, 119, 277–283. [Google Scholar] [CrossRef]
- Lu, H.; Guo, L.; Zhang, L.; Xie, C.; Li, W.; Gu, B.; Li, K. Study on quality characteristics of cassava flour and cassava flour short biscuits. Food Sci. Nutr. 2020, 8, 521–533. [Google Scholar] [CrossRef] [Green Version]
- Stavropoulou, D.A.; Filippou, P.; De Smet, S.; De Vuyst, L.; Leroy, F. Effect of temperature and pH on the community dynamics of coagulase-negative staphylococci during spontaneous meat fermentation in a model system. Food Microbiol. 2018, 76, 180–188. [Google Scholar] [CrossRef]
- Kumari, A.; Bhawal, S.; Kapila, S.; Yadav, H.; Kapila, R. Health-promoting role of dietary bioactive compounds through epigenetic modulations: A novel prophylactic and therapeutic approach. Crit. Rev. Food Sci. Nutr. 2020, 1–21. [Google Scholar] [CrossRef]
- Santini, T.C.; Peng, Y.G. Microbial Fermentation of Organic Carbon Substrates Drives Rapid pH Neutralization and Element Removal in Bauxite Residue Leachate. Environ. Sci. Technol. 2017, 51, 12592–12601. [Google Scholar] [CrossRef] [PubMed]
- Rahmani, R.; Beaufort, S.; Villarreal-Soto, S.A.; Taillandier, P.; Bouajila, J.; Debouba, M. Kombucha fermentation of African mustard (Brassica tournefortii) leaves: Chemical composition and bioactivity. Food Biosci. 2019, 30, 100414. [Google Scholar] [CrossRef] [Green Version]
- Wei, C.; Yu, L.; Qiao, N.; Wang, S.; Tian, F.; Zhao, J.; Zhang, H.; Zhai, Q.; Chen, W. The characteristics of patulin detoxification by Lactobacillus plantarum 13M5. Food Chem. Toxicol. 2020, 146, 111787. [Google Scholar] [CrossRef]
- Vashi, H.; Iorhemen, O.; Tay, J. Degradation of industrial tannin and lignin from pulp mill effluent by aerobic granular sludge technology. J. Water Process. Eng. 2018, 26, 38–45. [Google Scholar] [CrossRef]
- Suraiya, S.; Lee, J.M.; Cho, H.J.; Jang, W.J.; Kim, D.-G.; Kim, Y.-O.; Kong, I.-S. Monascus spp. fermented brown seaweeds extracts enhance bio-functional activities. Food Biosci. 2018, 21, 90–99. [Google Scholar] [CrossRef]
- Mohammadipanah, F.; Momenilandi, M. Potential of rare actinomycetes in the production of metabolites against multiple oxidant agents. Pharm. Biol. 2017, 56, 51–59. [Google Scholar] [CrossRef] [PubMed]
- Zhang, J.; Wang, W.; Mao, X. Chitopentaose protects HaCaT cells against H2O2-induced oxidative damage through modulating MAPKs and Nrf2/ARE signaling pathways. J. Funct. Foods 2020, 72, 104086. [Google Scholar] [CrossRef]
- Hseu, Y.-C.; Chou, C.-W.; Kumar, K.S.; Fu, K.-T.; Wang, H.-M.; Hsu, L.-S.; Kuo, Y.-H.; Wu, C.-R.; Chen, S.-C.; Yang, H.-L. Ellagic acid protects human keratinocyte (HaCaT) cells against UVA-induced oxidative stress and apoptosis through the upregulation of the HO-1 and Nrf-2 antioxidant genes. Food Chem. Toxicol. 2012, 50, 1245–1255. [Google Scholar] [CrossRef]
- Ahangari, R.; Khezri, S.; Jahedsani, A.; Bakhshii, S.; Salimi, A. Ellagic acid alleviates clozapine-induced oxidative stress and mitochondrial dysfunction in cardiomyocytes. Drug Chem. Toxicol. 2020, 1–9. [Google Scholar] [CrossRef] [PubMed]
- Hou, W.; Gao, W.; Wang, D.; Liu, Q.; Zheng, S.; Wang, Y. The Protecting Effect of Deoxyschisandrin and Schisandrin B on HaCaT Cells against UVB-Induced Damage. PLoS ONE 2015, 10, e0127177. [Google Scholar] [CrossRef] [Green Version]
- Li, Y.; Chen, X.; Niu, S.; Zhou, H.; Li, Q. Protective Antioxidant Effects of Amentoflavone and Total Flavonoids from Hedyotis diffusa on H2O2 -Induced HL-O2 Cells through ASK1/p38 MAPK Pathway. Chem. Biodivers. 2020, 17. [Google Scholar] [CrossRef]
- Chang, T.-M.; Tsen, J.-H.; Yen, H.; Yang, T.-Y.; Huang, H.-C. Extract fromPeriostracum cicadaeInhibits Oxidative Stress and Inflammation Induced by Ultraviolet B Irradiation on HaCaT Keratinocytes. Evid. Based Complement. Altern. Med. 2017, 2017, 1–12. [Google Scholar] [CrossRef] [Green Version]
- Shendge, A.K.; Basu, T.; Panja, S.; Chaudhuri, D.; Mandal, N. An ellagic acid isolated from Clerodendrum viscosum leaves ameliorates iron-overload induced hepatotoxicity in Swiss albino mice through inhibition of oxidative stress and the apoptotic pathway. Biomed. Pharmacother. 2018, 106, 454–465. [Google Scholar] [CrossRef]
- Sepand, M.R.; Ghahremani, M.H.; Razavi-Azarkhiavi, K.; Aghsami, M.; Rajabi, J.; Keshavarz-Bahaghighat, H.; Soodi, M. Ellagic acid confers protection against gentamicin-induced oxidative damage, mitochondrial dysfunction and apoptosis-related nephrotoxicity. J. Pharm. Pharmacol. 2016, 68, 1222–1232. [Google Scholar] [CrossRef]
- Liu, Y.; Liu, Y.; Liu, Y.; Liu, H.; Shang, Y. Evaluating effects of ellagic acid on the quality of kumquat fruits during storage. Sci. Hortic. 2018, 227, 244–254. [Google Scholar] [CrossRef]
- Nagamallu, R.; Srinivasan, B.; Ningappa, M.B.; Kariyappa, A.K. Synthesis of novel coumarin appended bis(formylpyrazole) derivatives: Studies on their antimicrobial and antioxidant activities. Bioorg. Med. Chem. Lett. 2016, 26, 690–694. [Google Scholar] [CrossRef] [PubMed]
- Hernández-Ruiz, K.L.; Ruiz-Cruz, S.; Cira-Chávez, L.A.; Gassos-Ortega, L.E.; Ornelas-Paz, J.D.J.; Del-Toro-Sánchez, C.L.; Márquez-Ríos, E.; López-Mata, M.A.; Rodríguez-Félix, F. Evaluation of Antioxidant Capacity, Protective Effect on Human Erythrocytes and Phenolic Compound Identification in Two Varieties of Plum Fruit (Spondias spp.) by UPLC-MS. Molecules 2018, 23, 3200. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ruan, X.; Liu, Y.; Wang, G.; Frost, R.L.; Qian, G.; Tsang, D.C. Transformation of functional groups and environmentally persistent free radicals in hydrothermal carbonisation of lignin. Bioresour. Technol. 2018, 270, 223–229. [Google Scholar] [CrossRef] [PubMed]
- Dai, C.-Y.; Liao, P.-R.; Zhao, M.-Z.; Gong, C.; Dang, Y.; Qu, Y.; Qiu, L.-S. Optimization of Ultrasonic Flavonoid Extraction from Saussurea involucrate, and the Ability of Flavonoids to Block Melanin Deposition in Human Melanocytes. Molecules 2020, 25, 313. [Google Scholar] [CrossRef] [Green Version]
- Mackei, M.; Vörösházi, J.; Sebők, C.; Neogrády, Z.; Mátis, G.; Jerzsele, Á. Fermented Wheat Germ Extract as a Redox Modulator: Alleviating Endotoxin-Triggered Oxidative Stress in Primary Cultured Rat Hepatocytes. Oxidative Med. Cell. Longev. 2020, 2020, 1–9. [Google Scholar] [CrossRef]
- Xie, X.; Dai, H.; Zhuang, B.; Chai, L.; Xie, Y.; Li, Y. Exogenous hydrogen sulfide promotes cell proliferation and differentiation by modulating autophagy in human keratinocytes. Biochem. Biophys. Res. Commun. 2016, 472, 437–443. [Google Scholar] [CrossRef] [PubMed]
- Xie, Y.; Guo, Y.; Cao, S.; Xue, M.; Fan, Z.; Gao, C.; Jin, B. Hydroxysafflor Yellow A Attenuates Hydrogen Peroxide-Induced Oxidative Damage on Human Umbilical Vein Endothelial Cells. Evid. Based Complement. Altern. Med. 2020, 2020, 1–8. [Google Scholar] [CrossRef]
- Acito, M.; Bartolini, D.; Ceccarini, M.R.; Russo, C.; Vannini, S.; Dominici, L.; Codini, M.; Villarini, M.; Galli, F.; Beccari, T.; et al. Imbalance in the antioxidant defence system and pro-genotoxic status induced by high glucose concentrations: In vitro testing in human liver cells. Toxicol. In Vitro 2020, 69, 105001. [Google Scholar] [CrossRef] [PubMed]
- Jaisin, Y.; Ratanachamnong, P.; Wongsawattkul, O.; Watthammawut, A.; Malaniyom, K.; Natewong, S. Antioxidant and anti-inflammatory effects of piperine on UV-B-irradiated human HaCaT keratinocyte cells. Life Sci. 2020, 263, 118607. [Google Scholar] [CrossRef] [PubMed]
- Trajčíková, E.; Kurin, E.; Slobodníková, L.; Straka, M.; Lichváriková, A.; Dokupilová, S.; Čičová, I.; Nagy, M.; Mučaji, P.; Fialová, S.B. Antimicrobial and Antioxidant Properties of Four Lycopus Taxa and an Interaction Study of Their Major Compounds. Molecules 2020, 25, 1422. [Google Scholar] [CrossRef] [Green Version]
 Content of ellagic acid. Data shown as the mean ± S.D. (n = 5).
Content of ellagic acid. Data shown as the mean ± S.D. (n = 5).
Content of ellagic acid. Data shown as the mean ± S.D. (n = 5).
Content of ellagic acid. Data shown as the mean ± S.D. (n = 5).









{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Serial Number | A/g | B/h | C/°C | Ellagic Acid Content (mg/g) |
|---|---|---|---|---|
| 1 | 3 | 36 | 37 | 95.25 |
| 2 | 6 | 48 | 37 | 84.29 |
| 3 | 3 | 24 | 35 | 83.04 |
| 4 | 3 | 48 | 35 | 79.86 |
| 5 | 3 | 36 | 37 | 96.41 |
| 6 | 0 | 36 | 39 | 82.29 |
| 7 | 0 | 48 | 37 | 83.94 |
| 8 | 3 | 36 | 37 | 95.48 |
| 9 | 6 | 36 | 39 | 83.53 |
| 10 | 0 | 36 | 35 | 80.71 |
| 11 | 0 | 24 | 37 | 86.31 |
| 12 | 6 | 36 | 35 | 84.73 |
| 13 | 3 | 25 | 39 | 86.75 |
| 14 | 3 | 48 | 39 | 82.62 |
| 15 | 3 | 36 | 37 | 97.80 |
| 16 | 3 | 36 | 37 | 97.22 |
| 17 | 6 | 24 | 37 | 90.33 |
| Parameter | Sum of Squares | df | Mean Squares | F-Value | p-Value |
|---|---|---|---|---|---|
| Model | 625.19 | 9 | 69.47 | 50.22 | <0.0001 *** |
| A | 11.59 | 1 | 11.59 | 8.38 | 0.0232 * |
| B | 30.89 | 1 | 30.89 | 22.33 | 0.0021 ** |
| C | 5.87 | 1 | 5.87 | 4.24 | 0.0483 * |
| AB | 3.37 | 1 | 3.37 | 2.43 | 0.0027 ** |
| AC | 1.93 | 1 | 1,93 | 1.40 | 0.0258 * |
| BC | 0.23 | 1 | 0.23 | 0.16 | 0.0184 * |
| A2 | 115.32 | 1 | 115.32 | 83.37 | <0.0001 *** |
| B2 | 104.46 | 1 | 104.46 | 75.52 | <0.0001 *** |
| C2 | 295.93 | 1 | 295.93 | 213.94 | <0.0001 *** |
| Residual | 9.68 | 7 | 1.38 | ||
| Lack of Fit | 4.89 | 3 | 1.63 | 1.36 | 0.3795 |
| Pure Error | 4.80 | 4 | 1.20 | ||
| Cor total | 634.87 | 16 | |||
| R2 | 0.9820 | ||||
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Liu, H.-M.; Xu, P.-F.; Cheng, M.-Y.; Lei, S.-N.; Liu, Q.-L.; Wang, W. Optimization of Fermentation Process of Pomegranate Peel and Schisandra Chinensis and the Biological Activities of Fermentation Broth: Antioxidant Activity and Protective Effect Against H2O2-induced Oxidative Damage in HaCaT Cells. Molecules 2021, 26, 3432. https://doi.org/10.3390/molecules26113432
Liu H-M, Xu P-F, Cheng M-Y, Lei S-N, Liu Q-L, Wang W. Optimization of Fermentation Process of Pomegranate Peel and Schisandra Chinensis and the Biological Activities of Fermentation Broth: Antioxidant Activity and Protective Effect Against H2O2-induced Oxidative Damage in HaCaT Cells. Molecules. 2021; 26(11):3432. https://doi.org/10.3390/molecules26113432
Chicago/Turabian StyleLiu, Hui-Min, Peng-Fei Xu, Ming-Yan Cheng, Sheng-Nan Lei, Qing-Lei Liu, and Wei Wang. 2021. "Optimization of Fermentation Process of Pomegranate Peel and Schisandra Chinensis and the Biological Activities of Fermentation Broth: Antioxidant Activity and Protective Effect Against H2O2-induced Oxidative Damage in HaCaT Cells" Molecules 26, no. 11: 3432. https://doi.org/10.3390/molecules26113432