
β-Sitosterol-D-Glucopyranoside Mimics Estrogenic Properties and Stimulates Glucose Utilization in Skeletal Muscle Cells

 , ,
, ,
Abstract
:1. Introduction
2. Materials and Methods
2.1. Materials
2.2. Plant Material
2.3. Extraction, Isolation, and Characterization of β-Sitosterol-D-Glucopyranoside(BSD)
2.4. Cell Culture
2.5. Transfections and Luciferase Assays
2.6. Glucose Uptake Assay
2.7. GLUT4 Translocation Assay
2.8. Animal
2.9. Western Blot Analysis
2.10. Gene Expression Analysis
2.11. Molecular Docking Study
2.12. Statistical Analysis
3. Results
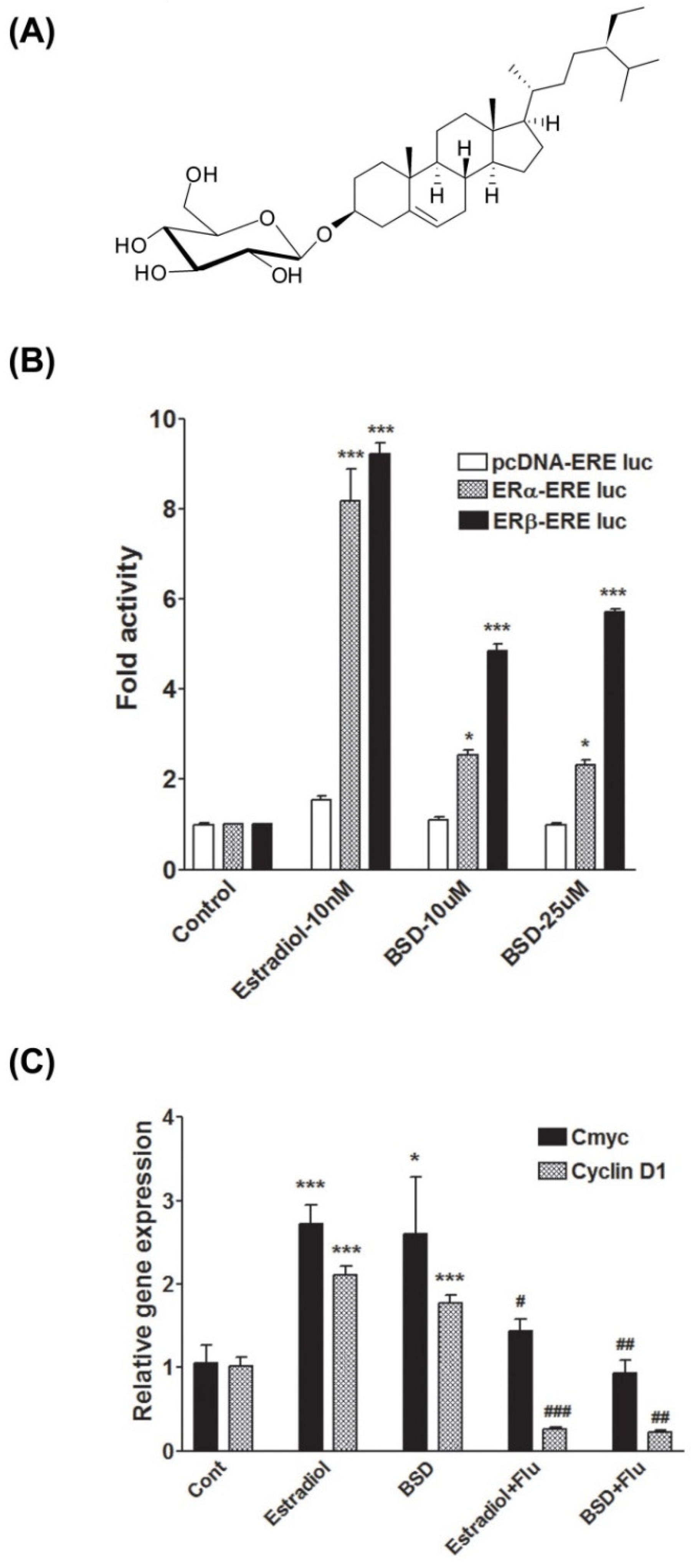
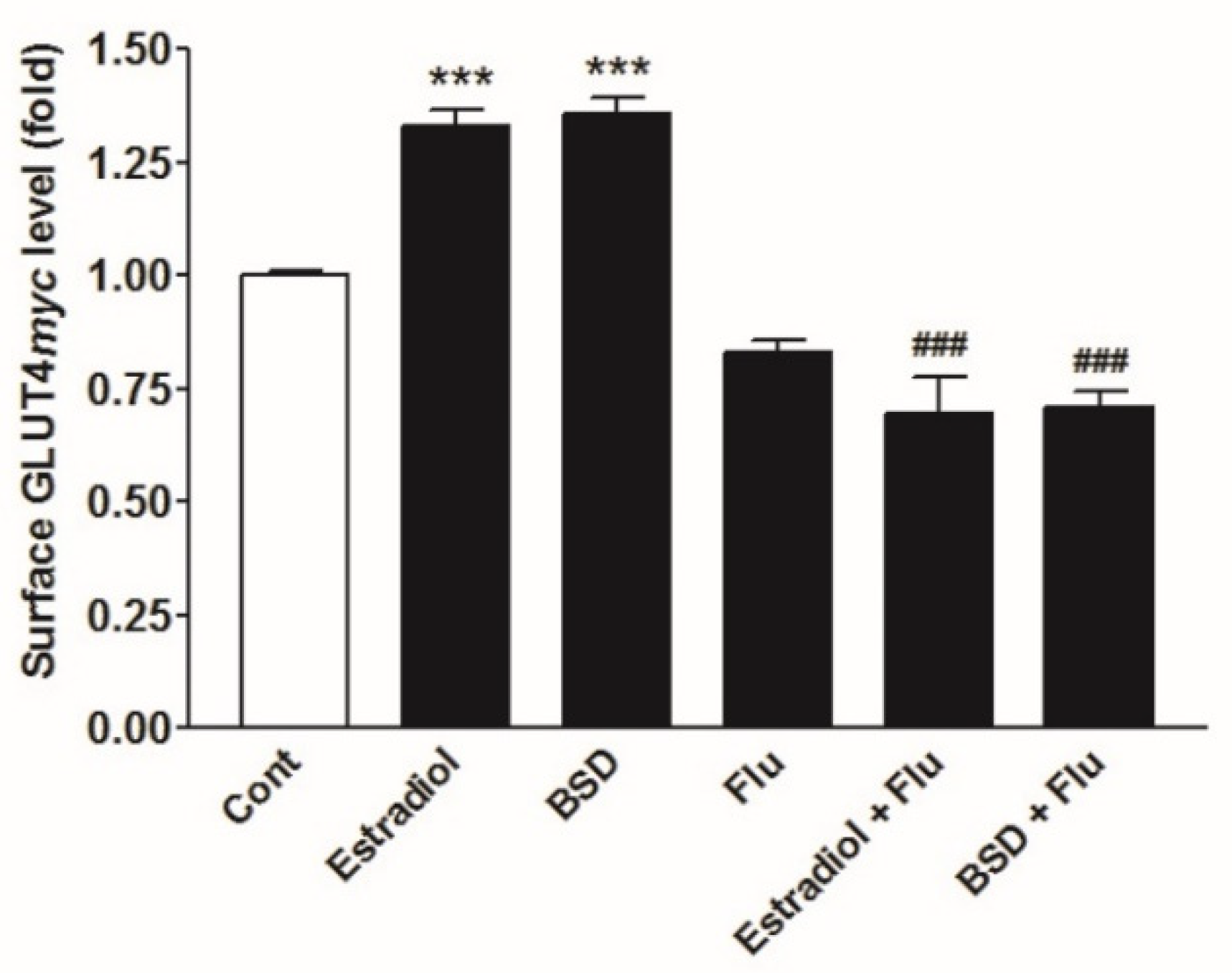
3.1. Phytoestrogenic Activity of BSD
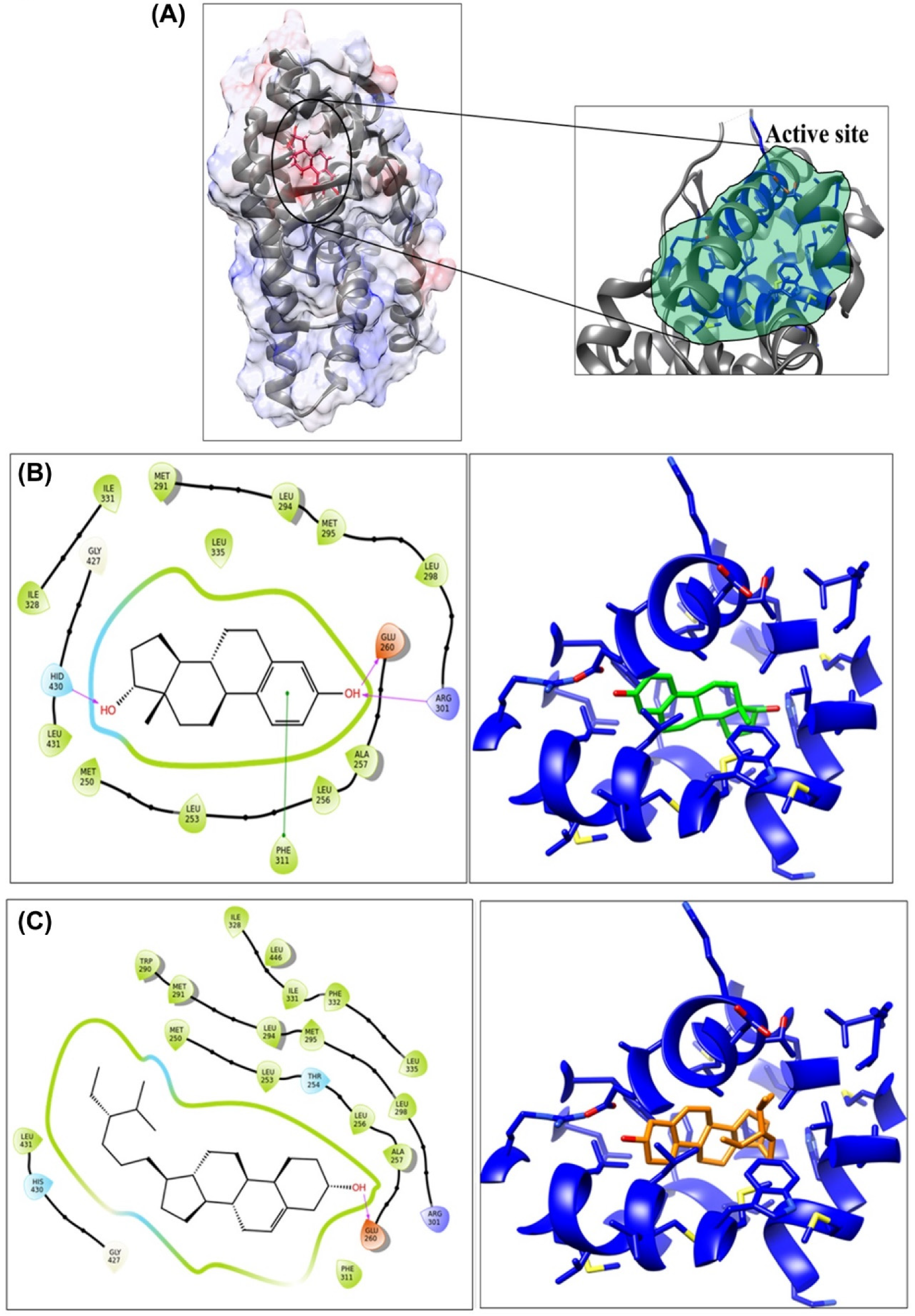
3.2. Molecular Interaction Study
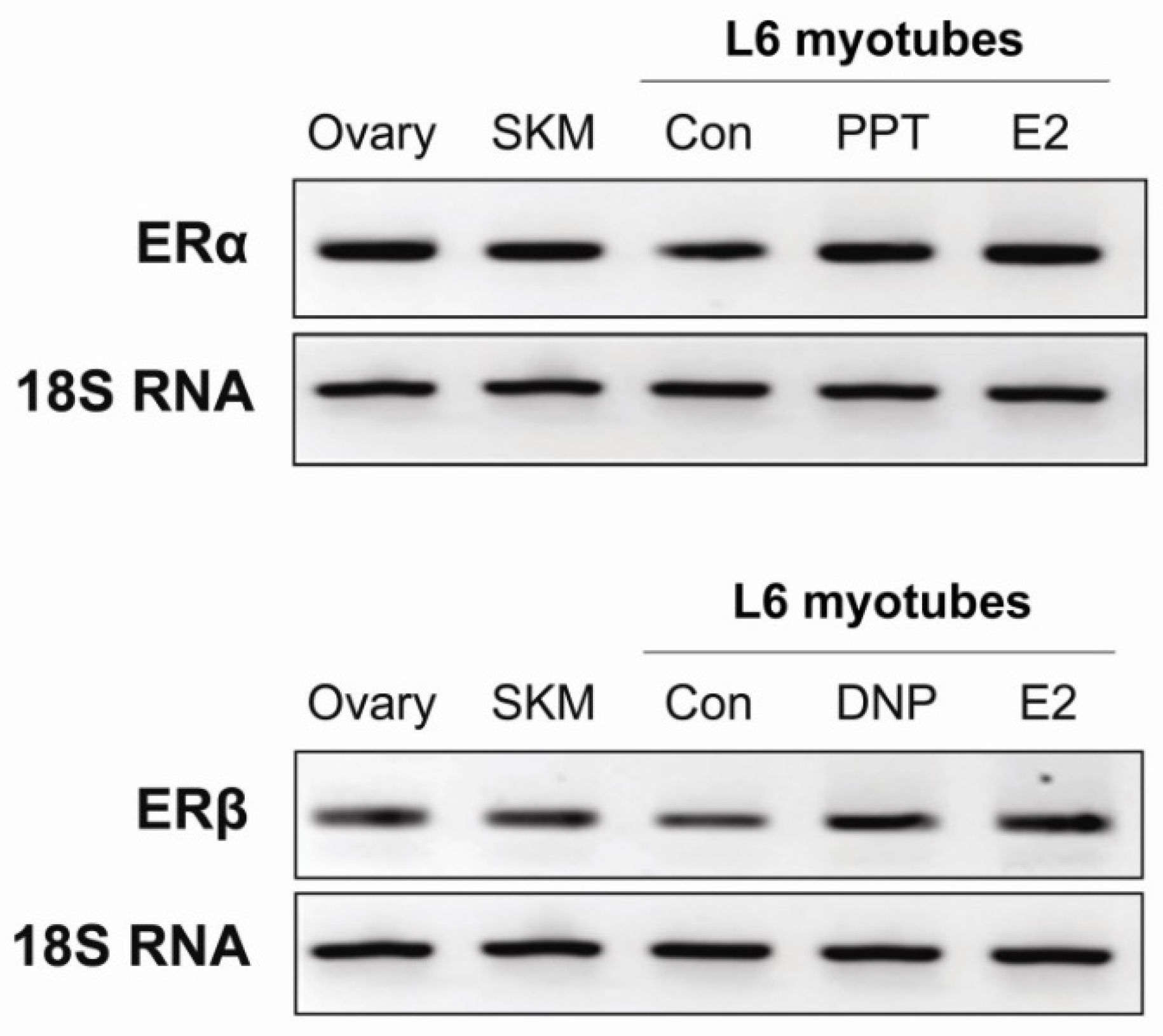
3.3. Skeletal Muscle Expresses Both Estrogen Receptor α and β
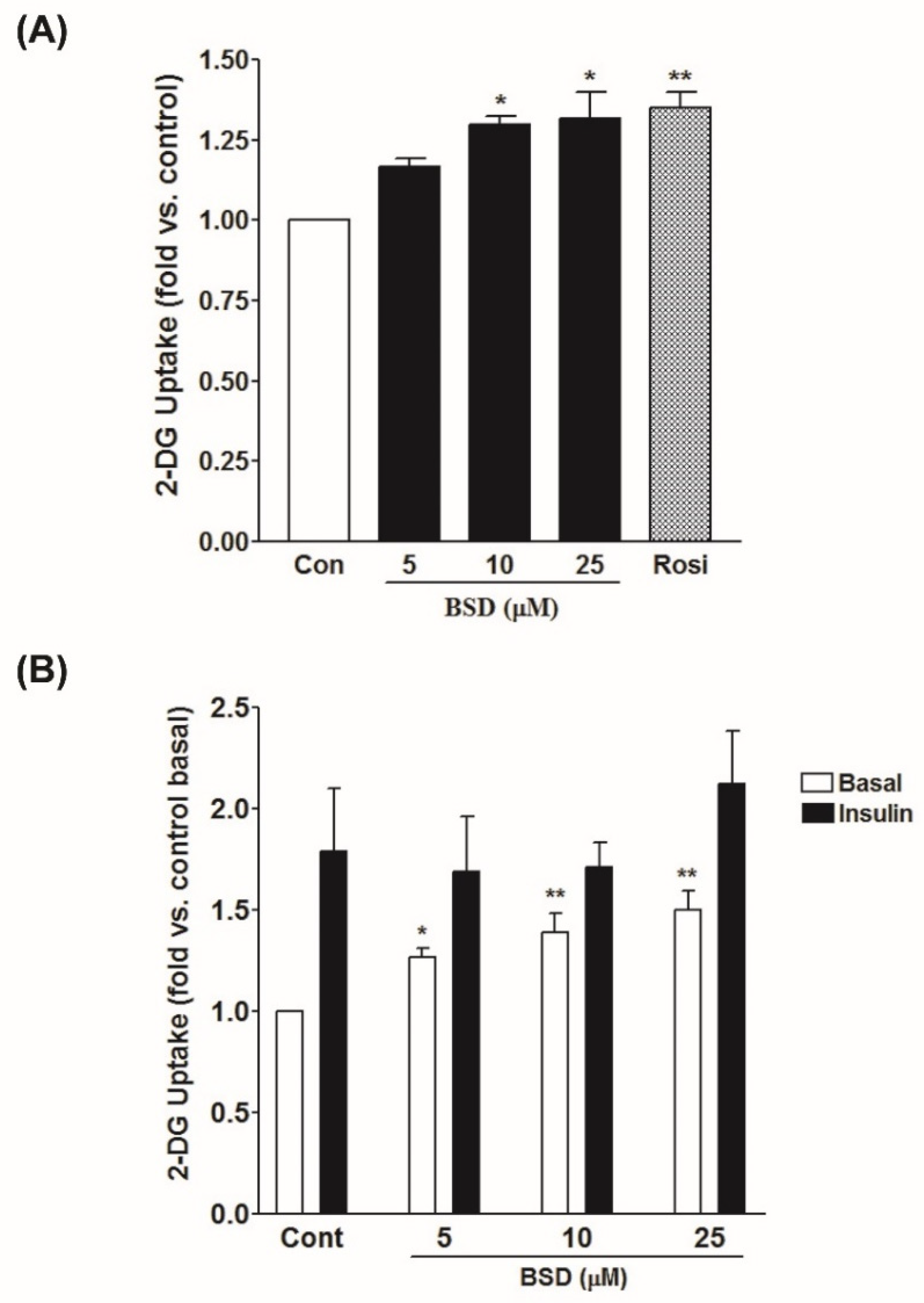
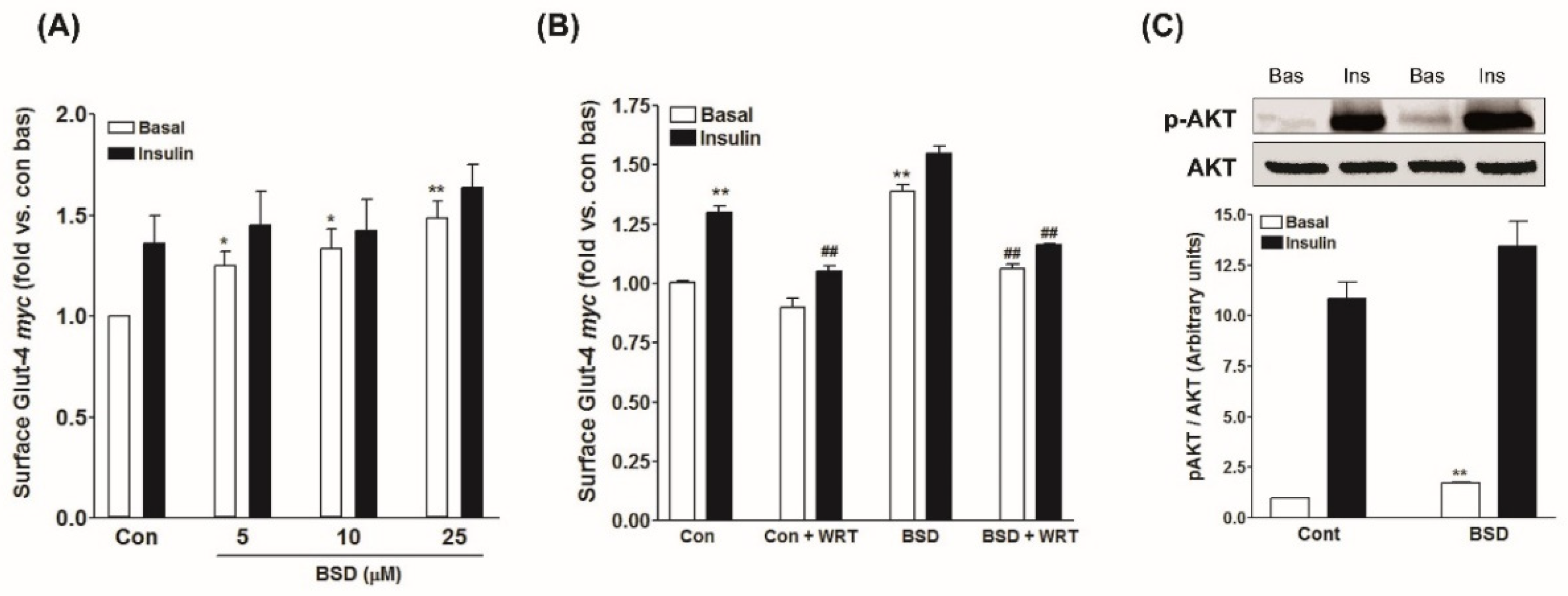
3.4. Effect of BSD on Glucose Uptake in L6-GLUT4myc Cells
3.5. Effect of BSD on GLUT4 Translocation in L6-GLUT4myc Cells
3.6. Effect of BSD on PI-3-K/Akt Mediated Signaling in Skeletal Muscle Cells
3.7. Effect of ER Inhibitor on BSD-Stimulated GLUT4myc Translocation
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
Sample Availability
References
- Meyer, M.R.; Clegg, D.J.; Prossnitz, E.R.; Barton, M. Obesity, insulin resistance and diabetes: Sex differences and role of oestrogen receptors. Acta Physiol. 2011, 203, 259–269. [Google Scholar] [CrossRef] [PubMed]
- Heine, P.A.; Taylor, J.A.; Iwamoto, G.A.; Lubahn, D.B.; Cooke, P.S. Increased adipose tissue in male and female estrogen receptor-alpha knockout mice. Proc. Natl. Acad. Sci. USA 2000, 97, 12729–12734. [Google Scholar] [CrossRef] [Green Version]
- Heldring, N.; Pike, A.; Andersson, S.; Matthews, J.; Cheng, G.; Hartman, J.; Tujague, M.; Strom, A.; Treuter, E.; Warner, M.; et al. Estrogen receptors: How do they signal and what are their targets. Physiol. Rev. 2007, 87, 905–931. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Nilsson, S.; Gustafsson, J.A. Estrogen receptors: Therapies targeted to receptor subtypes. Clin. Pharmacol. Ther. 2011, 89, 44–55. [Google Scholar] [CrossRef]
- Barros, R.P.; Machado, U.F.; Gustafsson, J.A. Estrogen receptors: New players in diabetes mellitus. Trends Mol. Med. 2006, 12, 425–431. [Google Scholar] [CrossRef] [PubMed]
- Barros, R.P.; Morani, A.; Moriscot, A.; Machado, U.F. Insulin resistance of pregnancy involves estrogen-induced repression of muscle glut4. Mol. Cell. Endocrinol. 2008, 295, 24–31. [Google Scholar] [CrossRef] [Green Version]
- Barros, R.P.; Gabbi, C.; Morani, A.; Warner, M.; Gustafsson, J.A. Participation of eralpha and erbeta in glucose homeostasis in skeletal muscle and white adipose tissue. Am. J. Physiol. Endocrinol. Metab. 2009, 297, E124–E133. [Google Scholar] [CrossRef]
- Levin, E.R. Translating extranuclear steroid receptor signaling to clinical medicine. Horm. Cancer 2014, 5, 140–145. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kang, L.; Zhang, X.; Xie, Y.; Tu, Y.; Wang, D.; Liu, Z.; Wang, Z.Y. Involvement of estrogen receptor variant er-alpha36, not gpr30, in nongenomic estrogen signaling. Mol. Endocrinol. 2010, 24, 709–721. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Prossnitz, E.R.; Barton, M. Estrogen biology: New insights into gper function and clinical opportunities. Mol. Cell. Endocrinol. 2014, 389, 71–83. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Garrido, P.; Moran, J.; Alonso, A.; Gonzalez, S.; Gonzalez, C. 17beta-estradiol activates glucose uptake via glut4 translocation and pi3k/akt signaling pathway in mcf-7 cells. Endocrinology 2013, 154, 1979–1989. [Google Scholar] [CrossRef] [Green Version]
- Lucas, T.F.; Siu, E.R.; Esteves, C.A.; Monteiro, H.P.; Oliveira, C.A.; Porto, C.S.; Lazari, M.F. 17beta-estradiol induces the translocation of the estrogen receptors esr1 and esr2 to the cell membrane, mapk3/1 phosphorylation and proliferation of cultured immature rat sertoli cells. Biol. Reprod. 2008, 78, 101–114. [Google Scholar] [CrossRef] [Green Version]
- Campello, R.S.; Fatima, L.A.; Barreto-Andrade, J.N.; Lucas, T.F.; Mori, R.C.; Porto, C.S.; Machado, U.F. Estradiol-induced regulation of glut4 in 3t3-l1 cells: Involvement of esr1 and akt activation. J. Mol. Endocrinol. 2017, 59, 257–268. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Muraki, K.; Okuya, S.; Tanizawa, Y. Estrogen receptor alpha regulates insulin sensitivity through irs-1 tyrosine phosphorylation in mature 3t3-l1 adipocytes. Endocr. J. 2006, 53, 841–851. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Velders, M.; Schleipen, B.; Fritzemeier, K.H.; Zierau, O.; Diel, P. Selective estrogen receptor-beta activation stimulates skeletal muscle growth and regeneration. FASEB J. 2012, 26, 1909–1920. [Google Scholar] [CrossRef]
- Kuiper, G.G.; Carlsson, B.; Grandien, K.; Enmark, E.; Haggblad, J.; Nilsson, S.; Gustafsson, J.A. Comparison of the ligand binding specificity and transcript tissue distribution of estrogen receptors alpha and beta. Endocrinology 1997, 138, 863–870. [Google Scholar] [CrossRef]
- Goodman-Gruen, D.; Kritz-Silverstein, D. Usual dietary isoflavone intake is associated with cardiovascular disease risk factors in postmenopausal women. J. Nutr. 2001, 131, 1202–1206. [Google Scholar] [CrossRef] [PubMed]
- Wagner, J.D.; Cefalu, W.T.; Anthony, M.S.; Litwak, K.N.; Zhang, L.; Clarkson, T.B. Dietary soy protein and estrogen replacement therapy improve cardiovascular risk factors and decrease aortic cholesteryl ester content in ovariectomizedcynomolgus monkeys. Metabolism 1997, 46, 698–705. [Google Scholar] [CrossRef]
- Cederroth, C.R.; Vinciguerra, M.; Kuhne, F.; Madani, R.; Doerge, D.R.; Visser, T.J.; Foti, M.; Rohner-Jeanrenaud, F.; Vassalli, J.D.; Nef, S. A phytoestrogen-rich diet increases energy expenditure and decreases adiposity in mice. Environ. Health Perspect. 2007, 115, 1467–1473. [Google Scholar] [CrossRef]
- Petersen, K.F.; Shulman, G.I. New insights into the pathogenesis of insulin resistance in humans using magnetic resonance spectroscopy. Obesity 2006, 14 (Suppl. S2), 34S–40S. [Google Scholar] [CrossRef] [Green Version]
- Zaid, H.; Antonescu, C.N.; Randhawa, V.K.; Klip, A. Insulin action on glucose transporters through molecular switches, tracks and tethers. Biochem. J. 2008, 413, 201–215. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Arha, D.; Ramakrishna, E.; Gupta, A.P.; Rai, A.K.; Sharma, A.; Ahmad, I.; Riyazuddin, M.; Gayen, J.R.; Maurya, R.; Tamrakar, A.K. Isoalantolactone derivative promotes glucose utilization in skeletal muscle cells and increases energy expenditure in db/db mice via activating ampk-dependent signaling. Mol. Cell. Endocrinol. 2018, 460, 134–151. [Google Scholar] [CrossRef] [PubMed]
- Moller, D.E. New drug targets for type 2 diabetes and the metabolic syndrome. Nature 2001, 414, 821–827. [Google Scholar] [CrossRef]
- Pandeti, S.; Arha, D.; Mishra, A.; Reddy, S.S.; Srivastava, A.K.; Narender, T.; Tamrakar, A.K. Glucose uptake stimulatory potential and antidiabetic activity of the arnebin-1 from arnabianobelis. Eur. J. Pharmacol. 2016, 789, 449–457. [Google Scholar] [CrossRef] [PubMed]
- Drexel, H.; Breier, C.; Lisch, H.J.; Sailer, S. Lowering plasma cholesterol with beta-sitosterol and diet. Lancet 1981, 317, 1157. [Google Scholar] [CrossRef]
- Gupta, R.; Sharma, A.K.; Dobhal, M.P.; Sharma, M.C.; Gupta, R.S. Antidiabetic and antioxidant potential of beta-sitosterol in streptozotocin-induced experimental hyperglycemia. J. Diabetes 2011, 3, 29–37. [Google Scholar] [CrossRef] [PubMed]
- Ivorra, M.D.; Paya, M.; Villar, A. Effect of beta-sitosterol-3-beta-d-glucoside on insulin secretion in vivo in diabetic rats and in vitro in isolated rat islets of langerhans. Pharmazie 1990, 45, 271–273. [Google Scholar]
- Flamini, G.; Antognoli, E.; Morelli, I. Two flavonoids and other compounds from the aerial parts of centaureabracteata from italy. Phytochemistry 2001, 57, 559–564. [Google Scholar] [CrossRef]
- Tamrakar, A.K.; Jaiswal, N.; Yadav, P.P.; Maurya, R.; Srivastava, A.K. Pongamol from pongamiapinnata stimulates glucose uptake by increasing surface glut4 level in skeletal muscle cells. Mol. Cell. Endocrinol. 2011, 339, 98–104. [Google Scholar] [CrossRef]
- Tamrakar, A.K.; Schertzer, J.D.; Chiu, T.T.; Foley, K.P.; Bilan, P.J.; Philpott, D.J.; Klip, A. Nod2 activation induces muscle cell-autonomous innate immune responses and insulin resistance. Endocrinology 2010, 151, 5624–5637. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Halgren, T.A. Identifying and characterizing binding sites and assessing druggability. J. Chem. Inf. Model. 2009, 49, 377–389. [Google Scholar] [CrossRef] [PubMed]
- Friesner, R.A.; Banks, J.L.; Murphy, R.B.; Halgren, T.A.; Klicic, J.J.; Mainz, D.T.; Repasky, M.P.; Knoll, E.H.; Shelley, M.; Perry, J.K.; et al. Glide: A new approach for rapid, accurate docking and scoring. 1. Method and assessment of docking accuracy. J. Med. Chem. 2004, 47, 1739–1749. [Google Scholar] [CrossRef] [PubMed]
- Sastry, G.M.; Adzhigirey, M.; Day, T.; Annabhimoju, R.; Sherman, W. Protein and ligand preparation: Parameters, protocols, and influence on virtual screening enrichments. J. Comput.-Aided Mol. Des. 2013, 27, 221–234. [Google Scholar] [CrossRef]
- Friesner, R.A.; Murphy, R.B.; Repasky, M.P.; Frye, L.L.; Greenwood, J.R.; Halgren, T.A.; Sanschagrin, P.C.; Mainz, D.T. Extra precision glide: Docking and scoring incorporating a model of hydrophobic enclosure for protein-ligand complexes. J. Med. Chem. 2006, 49, 6177–6196. [Google Scholar] [CrossRef] [Green Version]
- Pettersen, E.F.; Goddard, T.D.; Huang, C.C.; Couch, G.S.; Greenblatt, D.M.; Meng, E.C.; Ferrin, T.E. Ucsf chimera—A visualization system for exploratory research and analysis. J. Comput. Chem. 2004, 25, 1605–1612. [Google Scholar] [CrossRef] [Green Version]
- DeFronzo, R.A.; Tripathy, D. Skeletal muscle insulin resistance is the primary defect in type 2 diabetes. Diabetes Care 2009, 32 (Suppl. 2), S157–S163. [Google Scholar] [CrossRef] [Green Version]
- Huang, S.; Czech, M.P. The glut4 glucose transporter. Cell Metab. 2007, 5, 237–252. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bjornholm, M.; Zierath, J.R. Insulin signal transduction in human skeletal muscle: Identifying the defects in type ii diabetes. Biochem. Soc. Trans. 2005, 33, 354–357. [Google Scholar] [CrossRef] [Green Version]
- Louet, J.F.; LeMay, C.; Mauvais-Jarvis, F. Antidiabetic actions of estrogen: Insight from human and genetic mouse models. Curr. Atheroscler. Rep. 2004, 6, 180–185. [Google Scholar] [CrossRef] [PubMed]
- Kuiper, G.G.; Lemmen, J.G.; Carlsson, B.; Corton, J.C.; Safe, S.H.; van der Saag, P.T.; van der Burg, B.; Gustafsson, J.A. Interaction of estrogenic chemicals and phytoestrogens with estrogen receptor beta. Endocrinology 1998, 139, 4252–4263. [Google Scholar] [CrossRef]
- Weihua, Z.; Saji, S.; Makinen, S.; Cheng, G.; Jensen, E.V.; Warner, M.; Gustafsson, J.A. Estrogen receptor (er) beta, a modulator of eralpha in the uterus. Proc. Natl. Acad. Sci. USA 2000, 97, 5936–5941. [Google Scholar] [CrossRef] [Green Version]
- Jorgensen, M.; Vendelbo, B.; Skakkebaek, N.E.; Leffers, H. Assaying estrogenicity by quantitating the expression levels of endogenous estrogen-regulated genes. Environ. Health Perspect. 2000, 108, 403–412. [Google Scholar] [PubMed] [Green Version]
- Dubik, D.; Dembinski, T.C.; Shiu, R.P. Stimulation of c-myc oncogene expression associated with estrogen-induced proliferation of human breast cancer cells. Cancer Res. 1987, 47, 6517–6521. [Google Scholar] [PubMed]
- Sabbah, M.; Courilleau, D.; Mester, J.; Redeuilh, G. Estrogen induction of the cyclin d1 promoter: Involvement of a camp response-like element. Proc. Natl. Acad. Sci. USA 1999, 96, 11217–11222. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Taniguchi, C.M.; Emanuelli, B.; Kahn, C.R. Critical nodes in signalling pathways: Insights into insulin action. Nat. Rev. Mol. Cell Biol. 2006, 7, 85–96. [Google Scholar] [CrossRef]
- Alessi, D.R.; Andjelkovic, M.; Caudwell, B.; Cron, P.; Morrice, N.; Cohen, P.; Hemmings, B.A. Mechanism of activation of protein kinase b by insulin and igf-1. EMBO J. 1996, 15, 6541–6551. [Google Scholar] [CrossRef] [Green Version]
- Guo, R.X.; Wei, L.H.; Tu, Z.; Sun, P.M.; Wang, J.L.; Zhao, D.; Li, X.P.; Tang, J.M. 17 beta-estradiol activates pi3k/akt signaling pathway by estrogen receptor (er)-dependent and er-independent mechanisms in endometrial cancer cells. J. Steroid Biochem. Mol. Biol. 2006, 99, 9–18. [Google Scholar] [CrossRef]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Gene | Forward Primer | Reverse Primer |
|---|---|---|
| H-c-Myc | TTCGGGTAGTGGAAAACCAG | CAGCAGCTCGAATTTCTTCC |
| H-cyclin D1 | AACTACCTGGACCGCTTCCT | CCACTTGAGCTTGTTCACCA |
| H-18S RNA | TAGTTGGATCTTGGGAGCGG | TAGAACCGCGGTCCTATTCC |
| R-ERα | CAGCAGCGAGAAGGGAAACA | GGGCGGGGCTATTCTTCTTA |
| R-ERβ | GGACCCCAATGAACCAACG | CCTTCCTCTTCCCTATGCCC |
| R-18S RNA | AAACGGCTACCACATCCAAG | CCCTCTTAATCATGGCCTCA |
| Sr. No. | Name of Compound | Glide Score-XP (Kcal/mol) |
|---|---|---|
| 1. | Estradiol | −8.95 |
| 2. | BSD | −9.42 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Pandey, J.; Dev, K.; Chattopadhyay, S.; Kadan, S.; Sharma, T.; Maurya, R.; Sanyal, S.; Siddiqi, M.I.; Zaid, H.; Tamrakar, A.K. β-Sitosterol-D-Glucopyranoside Mimics Estrogenic Properties and Stimulates Glucose Utilization in Skeletal Muscle Cells. Molecules 2021, 26, 3129. https://doi.org/10.3390/molecules26113129
Pandey J, Dev K, Chattopadhyay S, Kadan S, Sharma T, Maurya R, Sanyal S, Siddiqi MI, Zaid H, Tamrakar AK. β-Sitosterol-D-Glucopyranoside Mimics Estrogenic Properties and Stimulates Glucose Utilization in Skeletal Muscle Cells. Molecules. 2021; 26(11):3129. https://doi.org/10.3390/molecules26113129
Chicago/Turabian StylePandey, Jyotsana, Kapil Dev, Sourav Chattopadhyay, Sleman Kadan, Tanuj Sharma, Rakesh Maurya, Sabyasachi Sanyal, Mohammad Imran Siddiqi, Hilal Zaid, and Akhilesh Kumar Tamrakar. 2021. "β-Sitosterol-D-Glucopyranoside Mimics Estrogenic Properties and Stimulates Glucose Utilization in Skeletal Muscle Cells" Molecules 26, no. 11: 3129. https://doi.org/10.3390/molecules26113129