
Red Blood Cell Stiffness and Adhesion Are Species-Specific Properties Strongly Affected by Temperature and Medium Changes in Single Cell Force Spectroscopy


Abstract
:1. Introduction
2. Results
2.1. AFM Tests: Apparent Young’s Modulus (Ea) in 0.9% Aqueous NaCl Solution
2.2. AFM Tests: Apparent Young’s Modulus (Ea) in Autologous Plasma
2.3. AFM Tests: Adhesion in 0.9% Aqueous NaCl Solution
2.4. AFM Tests: Adhesion in Autologous Plasma
3. Discussion
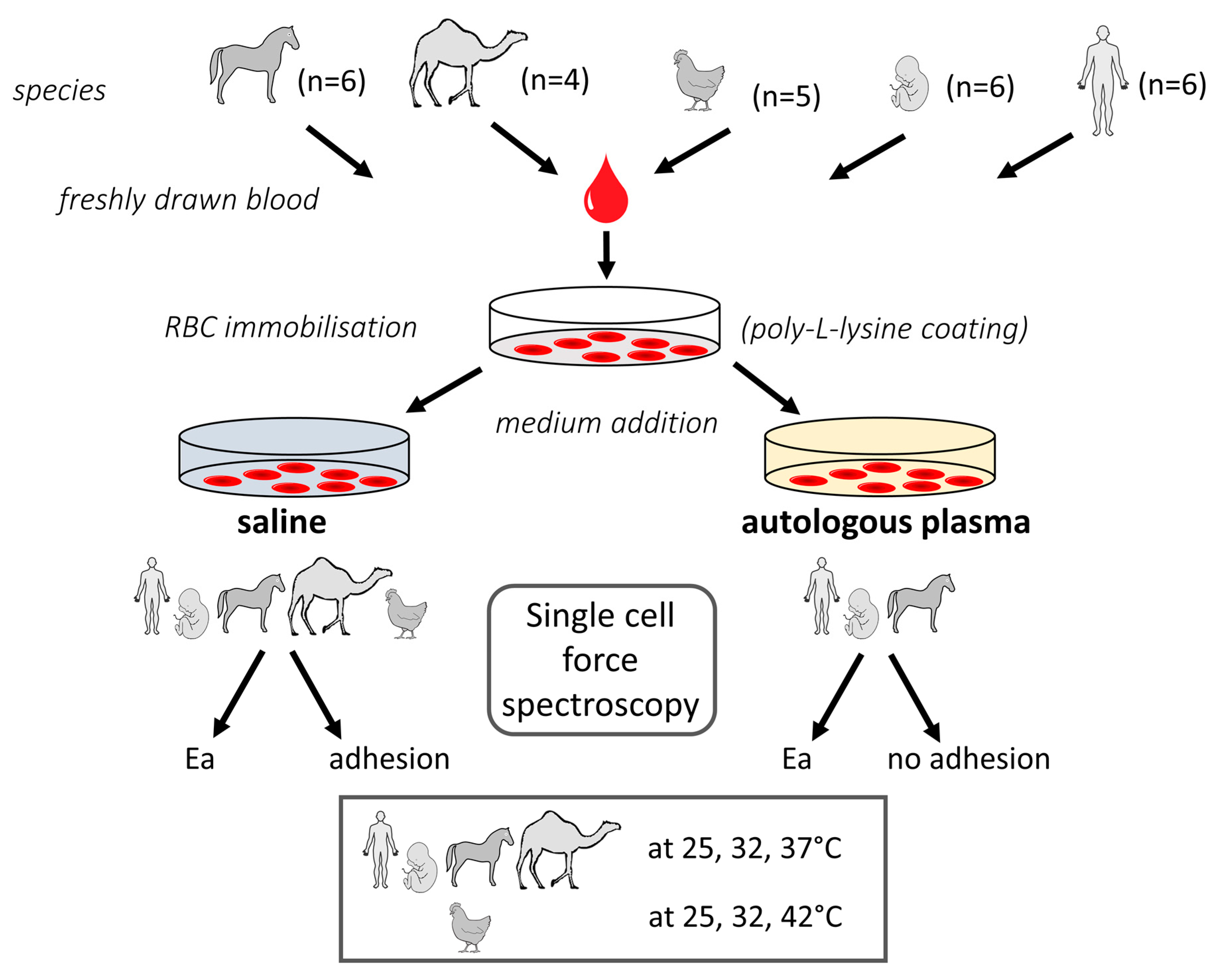
4. Materials and Methods
4.1. Sample and Medium Preparation
4.1.1. Blood Samples
4.1.2. Sample Preparation
4.2. AFM Tests
4.2.1. AFM Setup
4.2.2. Force Measurements
4.2.3. Theoretical Model and Data Processing
4.3. Statistics
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
Sample Availability
References
- De Oliveira, S.; Saldanha, C. An Overview About Erythrocyte Membrane. Clin. Hemorheol. Microcirc. 2010, 44, 63–74. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mohandas, N.; Gallagher, P.G. Red Cell Membrane: Past, Present, and Future. Blood 2008, 112, 3939–3948. [Google Scholar] [CrossRef] [Green Version]
- Mukherjee, R.; Saha, M.; Routray, A.; Chakraborty, C. Nanoscale Surface Characterization of Human Erythrocytes by Atomic Force Microscopy: A Critical Review. IEEE Trans. Nanobioscience 2015, 14, 625–633. [Google Scholar] [CrossRef]
- Choi, D. Medical Applications of Atomic Force Microscopy and Raman Spectroscopy. J. Nanosci. Nanotechnol. 2014, 14, 71–97. [Google Scholar] [CrossRef] [PubMed]
- Li, M.; Dang, D.; Liu, L.; Xi, N.; Wang, Y. Atomic Force Microscopy in Characterizing Cell Mechanics for Biomedical Applications: A Review. IEEE Trans. Nanobiosci. 2017, 16, 523–540. [Google Scholar] [CrossRef] [PubMed]
- Jembrek, M.J. Atomic Force Microscopy as an Advanced Tool in Neuroscience. Transl. Neurosci. 2015, 6, 117–130. [Google Scholar] [CrossRef] [Green Version]
- Musielak, M. Red Blood Cell-Deformability Measurement: Review of Techniques. Clin. Hemorheol. Microcirc. 2008, 42, 47–64. [Google Scholar] [CrossRef]
- Abkarian, M.; Viallat, A. On the Importance of the Deformability of Red Blood Cells in Blood Flow. In RSC Soft Matter; Royal Society of Chemistry: London, UK, 2016; pp. 347–362. [Google Scholar]
- Berdyyeva, T.; Woodworth, C.D.; Sokolov, I. Visualization of Cytoskeletal Elements by the Atomic Force Microscope. Ultramicroscopy 2005, 102, 189–198. [Google Scholar] [CrossRef] [Green Version]
- Bryk, A.H.; Wiśniewski, J.R. Quantitative Analysis of Human Red Blood Cell Proteome. J. Proteome Res. 2017, 16, 2752–2761. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Oberleithner, H.; Wälte, M.; Kusche-Vihrog, K. Sodium Renders Endothelial Cells Sticky for Red Blood Cells. Front. Physiol. 2015, 6, 2011–2016. [Google Scholar] [CrossRef] [Green Version]
- Kidoaki, S.; Matsuda, T. Adhesion Forces of the Blood Plasma Proteins on Self-Assembled Monolayer Surfaces of Alkanethiolates with Different Functional Groups Measured by an Atomic Force Microscope. Langmuir 1999, 15, 7639–7646. [Google Scholar] [CrossRef]
- Wang, M.S.; Palmer, L.; Schwartz, J.; Razatos, A. Evaluating Protein Attraction and Adhesion to Biomaterials with the Atomic Force Microscope. Langmuir 2004, 20, 7753–7759. [Google Scholar] [CrossRef] [PubMed]
- Scheffer, L.; Bitler, A.; Ben-Jacob, E.; Korenstein, R. Atomic Force Pulling: Probing the Local Elasticity of the Cell Membrane. Eur. Biophys. J. Biophys. Lett. 2001, 30, 83–90. [Google Scholar] [CrossRef] [PubMed]
- Morkvenaite-Vilkončiene, I.; Ramanavičiene, A.; Ramanavičius, A. Atomic Force Microscopy as a Tool for the Investigation of Living Cells. Medicina 2013, 49, 25. [Google Scholar] [CrossRef]
- Barns, S.; Balanant, M.A.; Sauret, E.; Flower, R.; Saha, S.; Gu, Y.T. Investigation of Red Blood Cell Mechanical Properties Using AFM Indentation And Coarse-Grained Particle Method. Biomed. Eng. Online 2017, 16, 1–21. [Google Scholar] [CrossRef] [Green Version]
- Ciasca, G.; Papi, M.; Di Claudio, S.; Chiarpotto, M.; Palmieri, V.; Maulucci, G.; Nocca, G.; Rossi, C.; De Spirito, M. Mapping Viscoelastic Properties of Healthy and Pathological Red Blood Cells at the Nanoscale Level. Nanoscale 2015, 7, 17030–17037. [Google Scholar] [CrossRef]
- Diez-Silva, M.; Dao, M.; Han, J.; Lim, C.T.; Suresh, S. Shape and Biomechanics Characteristics of Human Red Blood Cells in Health and Disease. MRS Bull. 2010, 35, 382–388. [Google Scholar] [CrossRef]
- Windberger, U. Blood suspensions in animals. In Dynamics of Blood Cell Suspensions in Microflows; CRC Press: Boca Raton, FL, USA, 2019; pp. 371–419. [Google Scholar]
- Linderkamp, O.; Wu, P.Y.; Meiselman, H. Geometry of neonatal and Adult Red Blood Cells. Pedriatr. Res. 1983, 17, 250–253. [Google Scholar] [CrossRef] [Green Version]
- El Bouhmadi, A.; Boulot, P.; Laffargue, F.; Brun, J.F. Rheological Properties of Fetal red cells with Special Reference to Aggregability and Disaggregability Analyzed by Light Transmission and Laser Backscattering Techniques. Clin. Hemorheol. Microcirc. 2000, 22, 79–90. [Google Scholar]
- Linderkamp, O.; Nash, G.B.; Wu, P.Y.K.; Meiselman, H.J. Deformability and Intrinsic Material Properties of Neonatal Red Blood Cells. Blood 1986, 67, 1244–1250. [Google Scholar] [CrossRef] [Green Version]
- Shapiro, D.L.; Pasqualini, P. Erythrocyte Membrane Proteins of Premature and Full-Term Newborn Infants. Pediatr. Res. 1978, 12, 176–178. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Calatroni, A.; Cordaro, V.; Salpietro, C.; Barberi, I. Erythrocyte Membrane Sialic Acid in New-Born Infants. Acta Haematol. 1984, 71, 198–203. [Google Scholar] [CrossRef] [PubMed]
- Inoue, S.; Lin, S.L.; Chang, T.; Wu, S.H.; Yao, C.W.; Chu, T.Y.; Troy, F.A.; Inoue, Y. Identification of Free Deaminated Sialic Acid (2-keto-3-deoxy-D-glycero- D-galacto-nononic acid) in Human Red Blood Cells And Its Elevated Expression in Fetal Cord Red Blood Cells and Ovarian Cancer Cells. J. Biol. Chem. 1998, 273, 27199–27204. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Matei, H.; Frentescu, L.; Benga, G. Comparative Studies of the Protein Composition of Red Blood Cell Membranes From Eight Mammalian Species. J. Cell. Mol. Med. 2000, 4, 270–276. [Google Scholar] [CrossRef] [PubMed]
- Baskurt, O.K.; Farley, R.A.; Meiselman, H.J. Erythrocyte Aggregation Tendency and Cellular Properties in Horse, Human, and Rat: A Comparative Study. Am. J. Physiol. Hear. Circ. Physiol. 1997, 273. [Google Scholar] [CrossRef] [PubMed]
- Bäumler, H.; Neu, B.; Mitlöhner, R.; Georgieva, R.; Meiselman, H.J.; Kiesewetter, H. Electrophoretic and Aggregation Behavior of Bovine, Horse and Human Red Blood Cells in Plasma and in Polymer Solutions. Biorheology 2001, 38, 39–51. [Google Scholar]
- Eylar, E.H.; Madoff, M.A.; Brody, O.V.; Oncley, J.L. The Contribution of Sialic Acid to the Surface Charge of the Erythrocyte. J. Biol. Chem. 1962, 237, 1992–2000. [Google Scholar] [CrossRef]
- Gulliver, G. On the Size of the Red Corpuscules of the Blood in the Vertebrates, with Copious Tables of Measurements. In Proceedings of the Zoological Society of London, Part XIII; Taylor, R., Taylor, J.E., Eds.; Red Lion Court; Trieste Publishing: Los Angeles, CA, USA, 1885; pp. 93–102. [Google Scholar]
- Windberger, U.; Auer, R.; Seltenhammer, M.; MacH, G.; Skidmore, J.A. Near-Newtonian Blood Behavior -Is It Good to Be a Camel? Front. Physiol. 2019. [Google Scholar] [CrossRef] [Green Version]
- Khodadad, J.K.; Weinstein, R.S. The Band 3-Rich Membrane of Llama Erythrocytes: Studies on Cell Shape and the Organization of Membrane Proteins. J. Membr. Biol. 1983, 72, 161–171. [Google Scholar] [CrossRef]
- McPherson, R.A.; Tilley, L.; Sawyer, W.H. Band 3 Mobility in Camelid Elliptocytes: Implications for Erythrocyte Shape. Biochemistry 1993, 32, 6696–6702. [Google Scholar] [CrossRef]
- Jay, D.G. Characterization of the Chicken Erythrocyte Anion Exchange Protein. J. Biol. Chem. 1983, 258, 9431–9436. [Google Scholar] [CrossRef]
- Blanchet, J.P. Chicken erythrocyte membranes: Comparison of Nuclear and Plasma Membranes from Adults And Embryos. Exp. Cell Res. 1974, 84, 159–166. [Google Scholar] [CrossRef]
- Jackson, R.C. The Exterior Surface of the Chicken erythrocyte. J. Biol. Chem. 1975, 250, 617–622. [Google Scholar] [CrossRef]
- Stier, A.; Bize, P.; Schull, Q.; Zoll, J.; Singh, F.; Geny, B.; Gros, F.; Royer, C.; Massemin, S.; Criscuolo, F. Avian Erythrocytes Have Functional Mitochondria, Opening Novel Perspectives for Birds as Animal Models in the Study Of Ageing. Front. Zool. 2013, 10, 1–9. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Woodcock, C.L.F. Nucleus-Associated Intermediate Filaments From Chicken Erythrocytes. J. Cell Biol. 1980, 85, 881–889. [Google Scholar] [CrossRef] [Green Version]
- Kitchen, H. Comparative biology: Animal Models of Human Hematologic Disease. Pediatr. Res. 1968, 2, 215. [Google Scholar] [CrossRef] [Green Version]
- Virtanen, I.; Kurkinen, M.; Lehto, V.P. Nucleus-Anchoring Cytoskeleton in Chicken Red Blood-Cells. Cell Biol. Int. Rep. 1979, 3, 157–162. [Google Scholar] [CrossRef]
- Steen, I.; Steen, J.B. The Importance of the Legs in the Thermoregulation of Birds. Acta Physiol Scand. 1965, 63, 285–291. [Google Scholar] [CrossRef]
- Granger, B.L.; Lazarides, E. Membrane Skeletal Protein 4.1 Of Avian Erythrocytes Is Composed Of Multiple Variants That Exhibit Tissue-Specific Expression. Cell 1984, 37, 595–607. [Google Scholar] [CrossRef]
- Cappella, B.; Stark, W. Adhesion of Amorphous Polymers As a Function of Temperature Probed with AFM Force-Distance Curves. J. Colloid Interface Sci. 2006, 296, 507–514. [Google Scholar] [CrossRef]
- Bassagañas, S.; Pérez-Garay, M.; Peracaula, R. Cell Surface Sialic Acid Modulates Extracellular Matrix Adhesion And Migration In Pancreatic Adenocarcinoma Cells. Pancreas 2014, 43, 109–117. [Google Scholar] [CrossRef] [PubMed]
- Seiradake, E.; Henaff, D.; Wodrich, H.; Billet, O.; Perreau, M.; Hippert, C.; Mennechet, F.; Schoehn, G.; Lortat-Jacob, H.; Dreja, H.; et al. The Cell Adhesion Molecule “CAR” and Sialic Acid On Human Erythrocytes Influence Adenovirus In Vivo Biodistribution. PLoS Pathog. 2009, 5. [Google Scholar] [CrossRef] [Green Version]
- Silker, A.; Engelhardt, H.; Sackmann, E. Dynamic Reflection Interference Contrast (Ric-) Microscopy: A New Method To Study Surface Excitations of Cells and To Measure Membrane Bending Elastic Moduli. J. Phys. Paris 1987, 48, 2139–2151. [Google Scholar] [CrossRef]
- Khodadad, J.K.; Weinstein, R.S. Band-3 Protein of the Red-Cell Membrane of the Llama—Crosslinking and Cleavage of the Cytoplasmic Domain. Biochem. Biophys. Res. Commun. 1985, 130, 493–499. [Google Scholar] [CrossRef]
- Steffen, P.; Verdier, C.; Wagner, C. Quantification of Depletion-Induced Adhesion Of Red Blood Cells. Phys. Rev. Lett. 2013, 110, 1–5. [Google Scholar] [CrossRef] [Green Version]
- Curry, F.E.; Michel, C.C. A Fiber Matrix Model Of Capillary Permeability. Microvasc. Res. 1980, 20, 96–99. [Google Scholar] [CrossRef]
- Siren, E.M.J.; Chapanian, R.; Constantinescu, I.; Brooks, D.E.; Kizhakkedathu, J.N. Oncotically Driven Control over Glycocalyx Dimension for Cell Surface Engineering and Protein Binding in the Longitudinal Direction. Sci. Rep. 2018, 8, 1–11. [Google Scholar] [CrossRef] [Green Version]
- Schnitzer, J.E.; Oh, P. Albondin-mediated capillary permeability to albumin. Differential Role Of Receptors In Endothelial Transcytosis And Endocytosis Of Native And Modified Albumins. J. Biol. Chem. 1994, 269, 6072–6082. [Google Scholar] [CrossRef]
- Wall, J.; Ayoub, F.; O’Shea, P. Interactions Of Macromolecules With The Mammalian Cell Surface. J. Cell Sci. 1995, 108, 2673–2682. [Google Scholar] [CrossRef]
- Jensen, T.B.; Cheema, M.U.; Szymiczek, A.; Damkier, H.H.; Praetorius, J. Renal Type A Intercalated Cells Contain Albumin In Organelles With Aldosterone- Regulated Abundance. PLoS ONE 2015, 10, 1–17. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Xu, Z.; Dou, W.; Wang, C.; Sun, Y. Stiffness and ATP Recovery Of Stored Red Blood Cells In Serum. Microsyst. Nanoeng. 2019, 5. [Google Scholar] [CrossRef] [PubMed]
- Manno, S.; Takakuwa, Y.; Mohandas, N. Modulation of Erythrocyte Membrane Mechanical Function by Protein 4.1 Phosphorylation. J. Biol. Chem. 2005, 280, 7581–7587. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bai, K.; Wang, W. Spatio-Temporal Development of the Endothelial Glycocalyx Layer and Its Mechanical Property In Vitro. J. R. Soc. Interface 2012, 9, 2290–2298. [Google Scholar] [CrossRef] [Green Version]
- Alcoutlabi, M.; McKenna, G.B. Effects of Confinement On Material Behaviour at the Nanometre Size Scale. J. Phys. Condens. Matter 2005, 17. [Google Scholar] [CrossRef]
- Li, T.; Riedo, E. Nonlinear Viscoelastic Dynamics of Nanoconfined Wetting Liquids. Phys. Rev. Lett. 2008, 100, 106102. [Google Scholar] [CrossRef] [Green Version]
- Pries, A.R.; Secomb, T.W.; Gaehtgens, P. The Endothelial Surface Layer. Pflugers Arch. Eur. J. Physiol. 2000, 440, 653–666. [Google Scholar] [CrossRef] [PubMed]
- Jeffrey, L.; Hutter, J.B. Calibration Of Atomic-Force Microscope Tips. Rev. Sci. Instrum. 1993, 64, 1868. [Google Scholar] [CrossRef] [Green Version]
- Rosenbluth, M.J. Probing the Role of Single Cell Mechanics in Disease with Atomic Force Microscopy and Microfluidics. Ph.D. Thesis, University of California, Berkeley, CA, USA, 2008. [Google Scholar]
- Chopinet, L.; Formosa, C.; Rols, M.P.; Duval, R.E.; Dague, E. Imaging Living Cells Surface and Quantifying Its Properties at High Resolution Using AFM in QITM mode. Micron 2013, 48, 26–33. [Google Scholar] [CrossRef]
- Friedrichs, J.; Helenius, J.; Muller, D.J. Quantifying Cellular Adhesion to Extracellular Matrix Components by Single-Cell Force Spectroscopy. Nat. Protoc. 2010, 5, 1353–1361. [Google Scholar] [CrossRef]
- Hertz, H. Ueber die Beruehrung Fester Elastischer Koerper. J. für die Reine und Angew. Math. 1881, 92, 156–171. [Google Scholar]
- Johnson, A.D.; Kendall, K.; Roberts, A.D. Surface Energy and the Contact of Elastic Solids. Proc. R. Soc. Lond. A Math. Phys. Sci. 1971, 234, 1558. [Google Scholar]
- Derjaguin, B.V.; Muller, V.M.; Toporov, Y.P. Effect of Contact Deformations on the Adhesion of Particles. J. Colloid Interface Sci. 1975, 53, 314–326. [Google Scholar] [CrossRef]
- Muller, V.M.; Derjaguin, B.V.; Toporov, Y.P. On Two Methods of Calculation of the Force of Sticking of An Elastic Sphere to a Rigid Plane. Colloids and Surfaces 1983, 7, 251–259. [Google Scholar] [CrossRef]
- Bilodeau, G.G. Regular Pyramid Punch Problem. J. Appl. Mech. 1992, 59, 518–523. [Google Scholar] [CrossRef]


{kind=link}
| RBC Type | RBC Shape and Size | Aggregation Index M1 | Elongation Index EImax | Membrane Proteins (Differences to Human) | Sialic Acid Content (Differences to Human) | Intracellular Structures |
|---|---|---|---|---|---|---|
Human adult  | biconcave diameter 7.5–8.7 μm [18] | 28.5 ± 6.6 [19] | 0.618 (0.612/0.623) [19] | - | - | none |
Human fetus  | 11% wider diameter than adult [20] | 4.84 ± 0.89 [21] | higher rigidity index [21], lower shear elastic modulus [22] | comparable [23] | increased [24,25] | none, 21% bigger cell volume [20] |
Horse  | biconcave diameter 5.7 µm | 55.9 ± 13.4 [19] | 0.679 (0.590/0.878) [19] | deficient in band 4.2 and band 6 [26,27] | increased [28,29] | none |
Camel  | elliptic dimension 7 × 4 µm [30] | none [31] | n.a. | higher amount of band 3, less mobile [32,33] | increased [32] | none |
Chicken  | elliptic dimension 12.1 × 7.3 µm nucleus: 4.2 × 2.8 µm [30] | none | n.a. | less band 3 copies, no adherence to GAPDH [34,35,36] | lower [29] | nucleus, mitochondria, marginal band, filaments [37,38] |
| Apparent Young’s Modulus (Ea; in Pa): | Percentile | ||||||
|---|---|---|---|---|---|---|---|
| Species | Medium | Temperature (°C) | 50% (Median) | 25% | 75% | p-Value (Comparing Temperatures) | RBCs Tested/Force-distance Curves Evaluated |
| human adult | 0.9% NaCl | 25 | 251 | 199 | 332 | 60/397 | |
| 32 | 169 | 125 | 226 | 0.001 | 60/413 | ||
| 37 | 179 | 136 | 238 | n.s. | 60/400 | ||
| autologous plasma | 25 | 144 | 101 | 224 | 60/187 | ||
| 32 | 126 | 91 | 162 | n.s. | 60/200 | ||
| 37 | 97 | 63 | 141 | 0.05 | 60/184 | ||
| human fetus | 0.9% NaCl | 25 | 754 | 527 | 954 | 50/357 | |
| 32 | 236 | 167 | 344 | 0.001 | 50/343 | ||
| 37 | 176 | 131 | 234 | 0.001 | 50/341 | ||
| autologous plasma | 25 | 217 | 138 | 299 | 30/122 | ||
| 32 | 133 | 103 | 177 | 0.01 | 30/113 | ||
| 37 | 100 | 80 | 175 | n.s. | 30/123 | ||
| horse | 0.9% NaCl | 25 | 1260 | 729 | 1753 | 60/397 | |
| 32 | 524 | 346 | 788 | 0.001 | 60/422 | ||
| 37 | 351 | 185 | 742 | n.s. | 60/406 | ||
| autologous plasma | 25 | 126 | 92 | 183 | 40/200 | ||
| 32 | 105 | 72 | 158 | n.s. | 40/160 | ||
| 37 | 96 | 71 | 138 | n.s. | 40/181 | ||
| camel | 0.9% NaCl | 25 | 825 | 565 | 1184 | 40/276 | |
| 32 | 489 | 320 | 697 | 0.001 | 40/296 | ||
| 37 | 439 | 345 | 600 | n.s. | 40/260 | ||
| autologous plasma | 25 | 583 | 272 | 863 | 40*/513 | ||
| 32 | 203 | 140 | 309 | 0.001 | 40*/455 | ||
| 37 | 164 | 109 | 221 | n.s. | 40*/463 | ||
| chicken | 0.9% NaCl | 25 | **2278/461 | 261 | 721 | 50/348 | |
| 32 | **1914/282 | 155 | 445 | 0.001 | 50/302 | ||
| 42 | **2384/183 | 91 | 427 | 0.01 | 50/274 | ||
| Adhesion (in aJ): | Percentiles | ||||||
|---|---|---|---|---|---|---|---|
| Species | Medium | Temperature (°C) | 50% (Median) | 25% | 75% | p-Value (Comparing Temperatures) | RBCs Tested/Force-Distance Curves Evaluated |
| human adult | 0.9% NaCl | 25 | 18 | 9 | 128 | 60/397 | |
| 32 | 31 | 15 | 63 | 0.001 | 60/413 | ||
| 37 | 44 | 21 | 149 | 0.05 | 60/400 | ||
| human fetus | 0.9% NaCl | 25 | 74 | 42 | 112 | 50/357 | |
| 32 | 108 | 57 | 168 | 0.001 | 50/343 | ||
| 37 | 137 | 89 | 221 | 0.01 | 50/341 | ||
| horse | 0.9% NaCl | 25 | 90 | 51 | 174 | 60/397 | |
| 32 | 68 | 42 | 175 | n.s. | 60/422 | ||
| 37 | 171 | 51 | 252 | 0.001 | 60/406 | ||
| camel | 0.9% NaCl | 25 | 143 | 55 | 194 | 40/276 | |
| 32 | 67 | 32 | 122 | 0.001 | 40/296 | ||
| 37 | 159 | 83 | 239 | 0.001 | 40/260 | ||
| chicken | 0.9% NaCl | 25 | 35 | 14 | 66 | 50/348 | |
| 32 | 32 | 2 | 97 | 0.01 | 50/302 | ||
| 42 | 7 | 1 | 23 | 0.001 | 50/274 | ||
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Baier, D.; Müller, T.; Mohr, T.; Windberger, U. Red Blood Cell Stiffness and Adhesion Are Species-Specific Properties Strongly Affected by Temperature and Medium Changes in Single Cell Force Spectroscopy. Molecules 2021, 26, 2771. https://doi.org/10.3390/molecules26092771
Baier D, Müller T, Mohr T, Windberger U. Red Blood Cell Stiffness and Adhesion Are Species-Specific Properties Strongly Affected by Temperature and Medium Changes in Single Cell Force Spectroscopy. Molecules. 2021; 26(9):2771. https://doi.org/10.3390/molecules26092771
Chicago/Turabian StyleBaier, Dina, Torsten Müller, Thomas Mohr, and Ursula Windberger. 2021. "Red Blood Cell Stiffness and Adhesion Are Species-Specific Properties Strongly Affected by Temperature and Medium Changes in Single Cell Force Spectroscopy" Molecules 26, no. 9: 2771. https://doi.org/10.3390/molecules26092771