
Bis-3-Chloropiperidines Targeting TAR RNA as A Novel Strategy to Impair the HIV-1 Nucleocapsid Protein

 , ,
, ,
Abstract
:1. Introduction
2. Results and Discussion
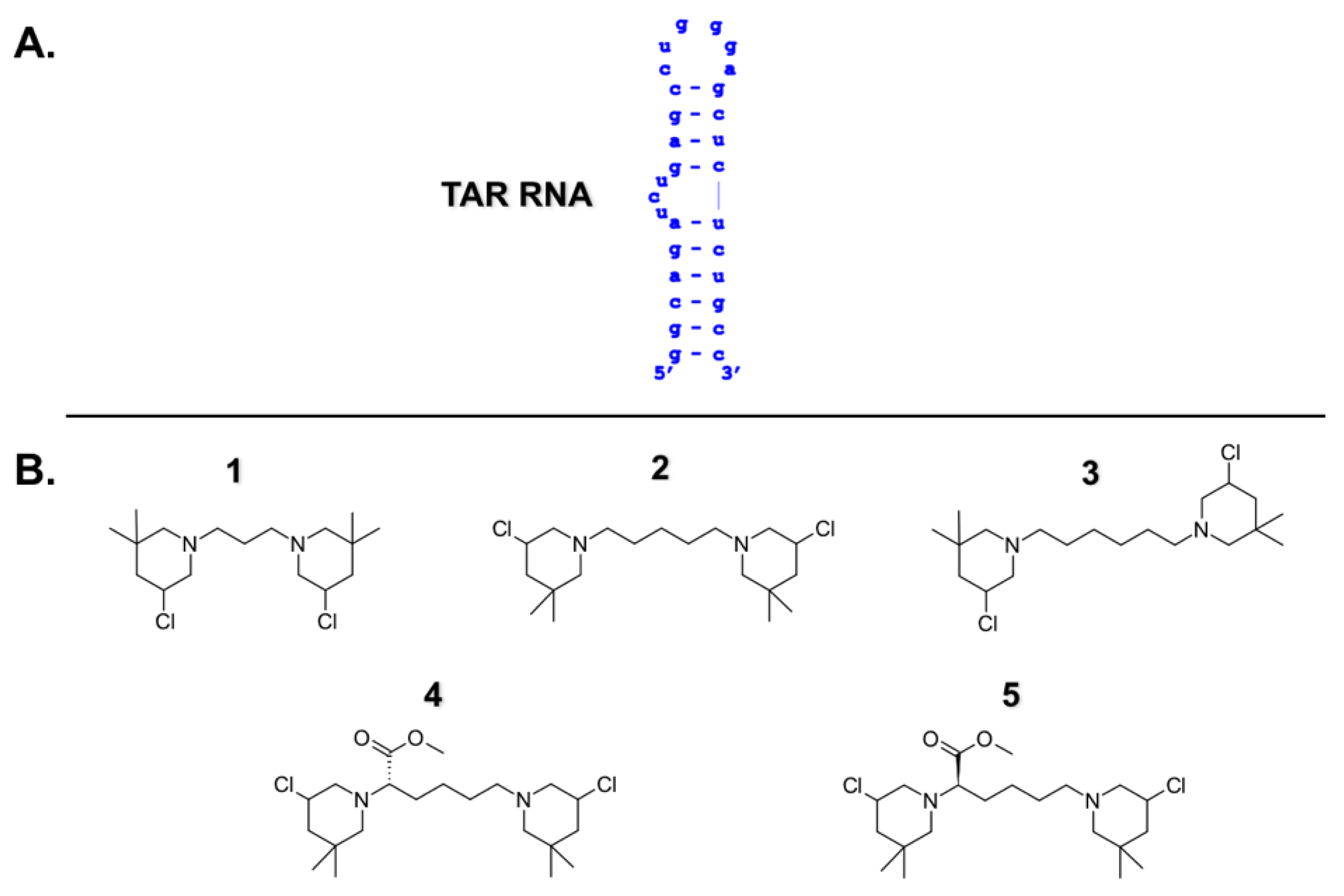
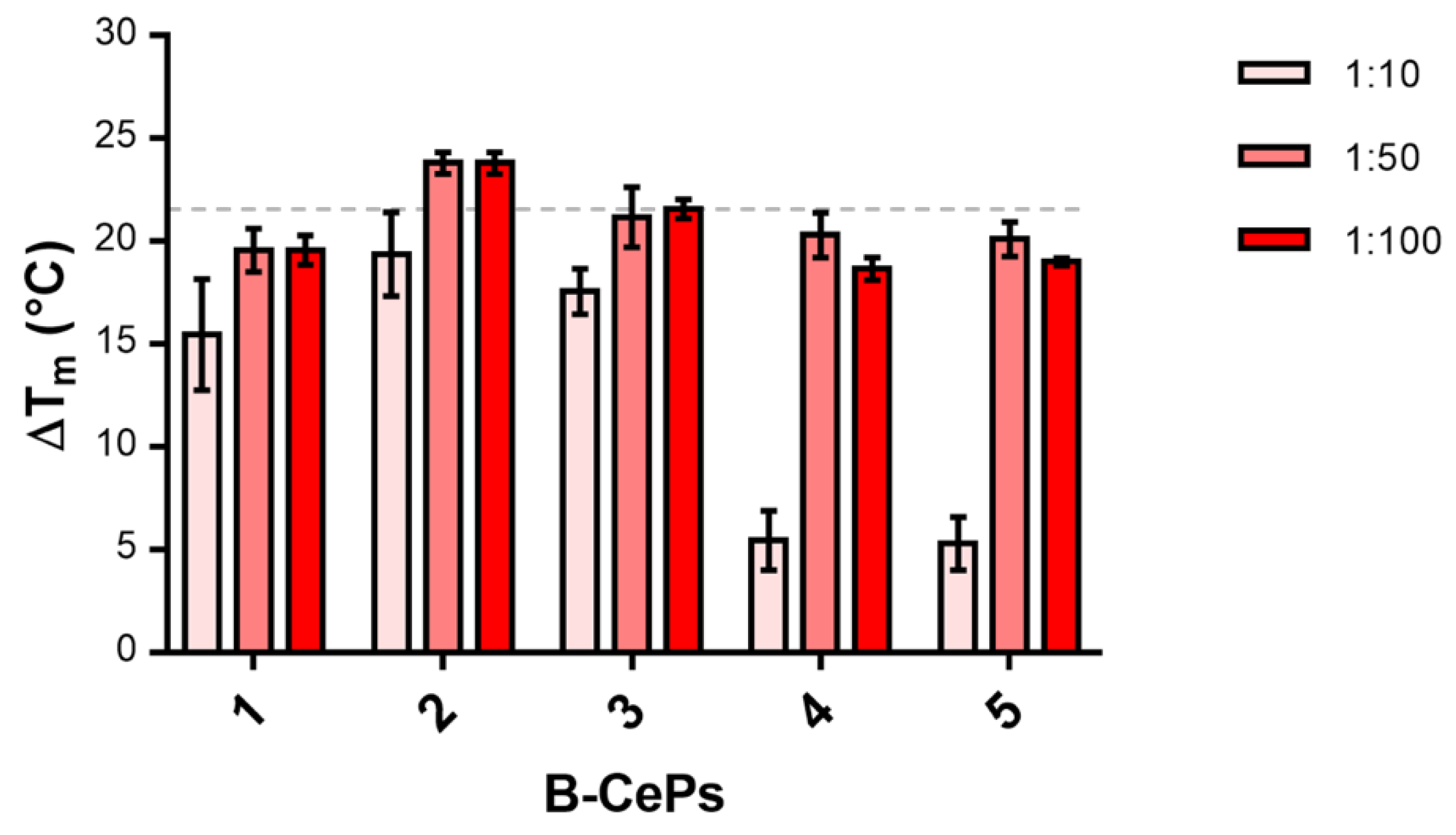
2.1. B-CePs Stabilize TAR RNA Hairpin Structure
2.2. B-CePs Directly Interact with TAR RNA
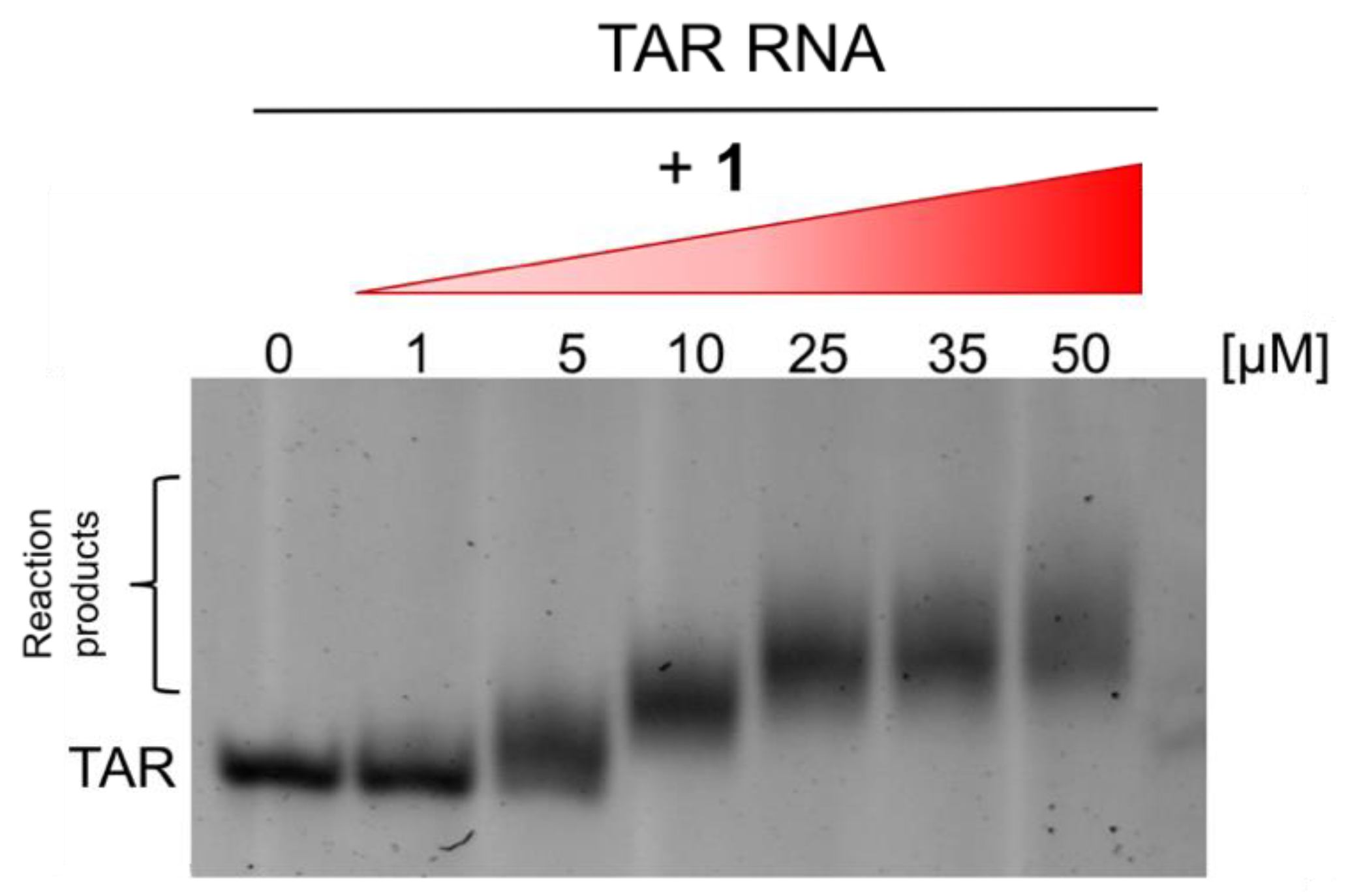
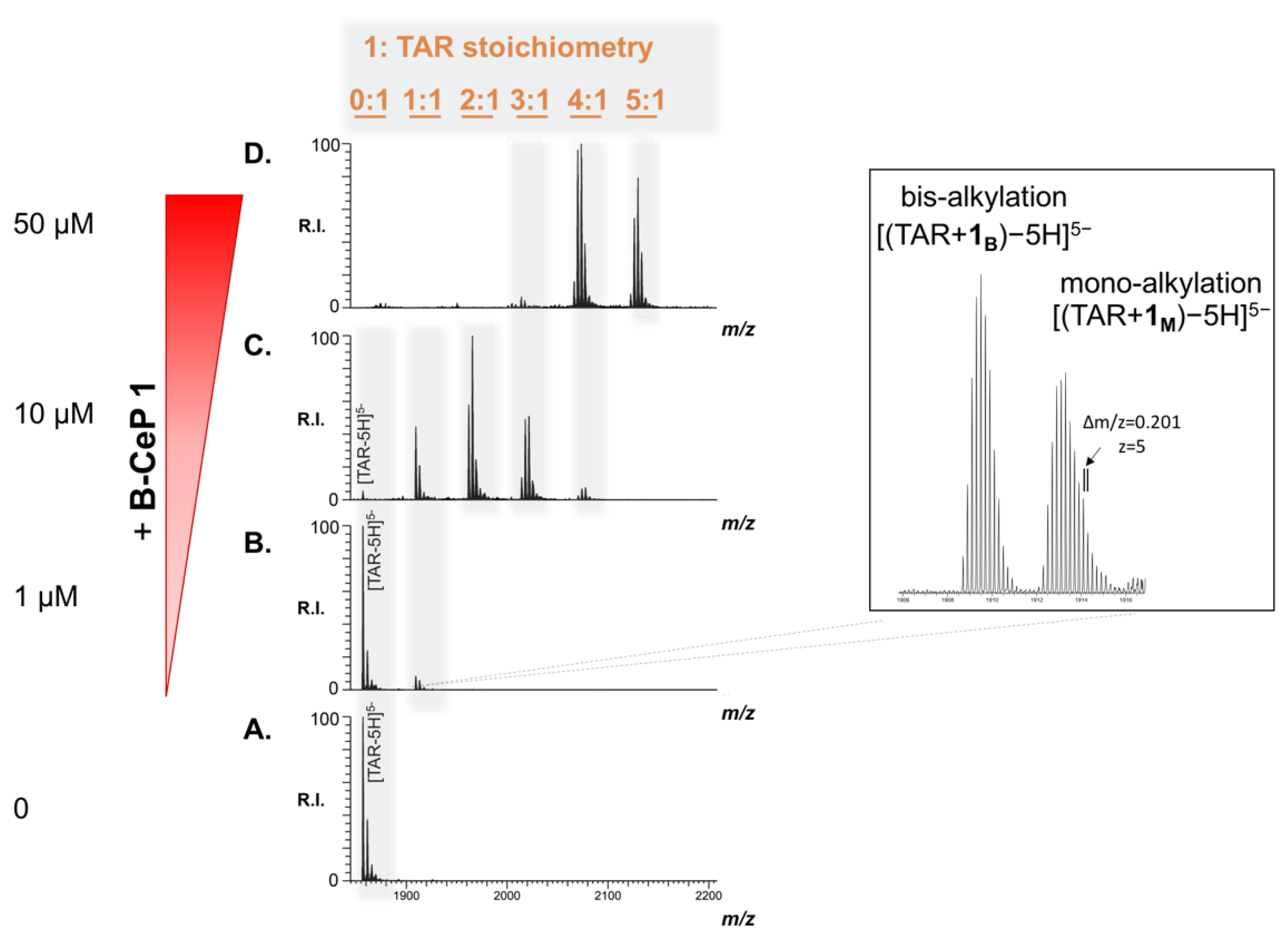
2.3. B-CePs Covalently React with TAR RNA
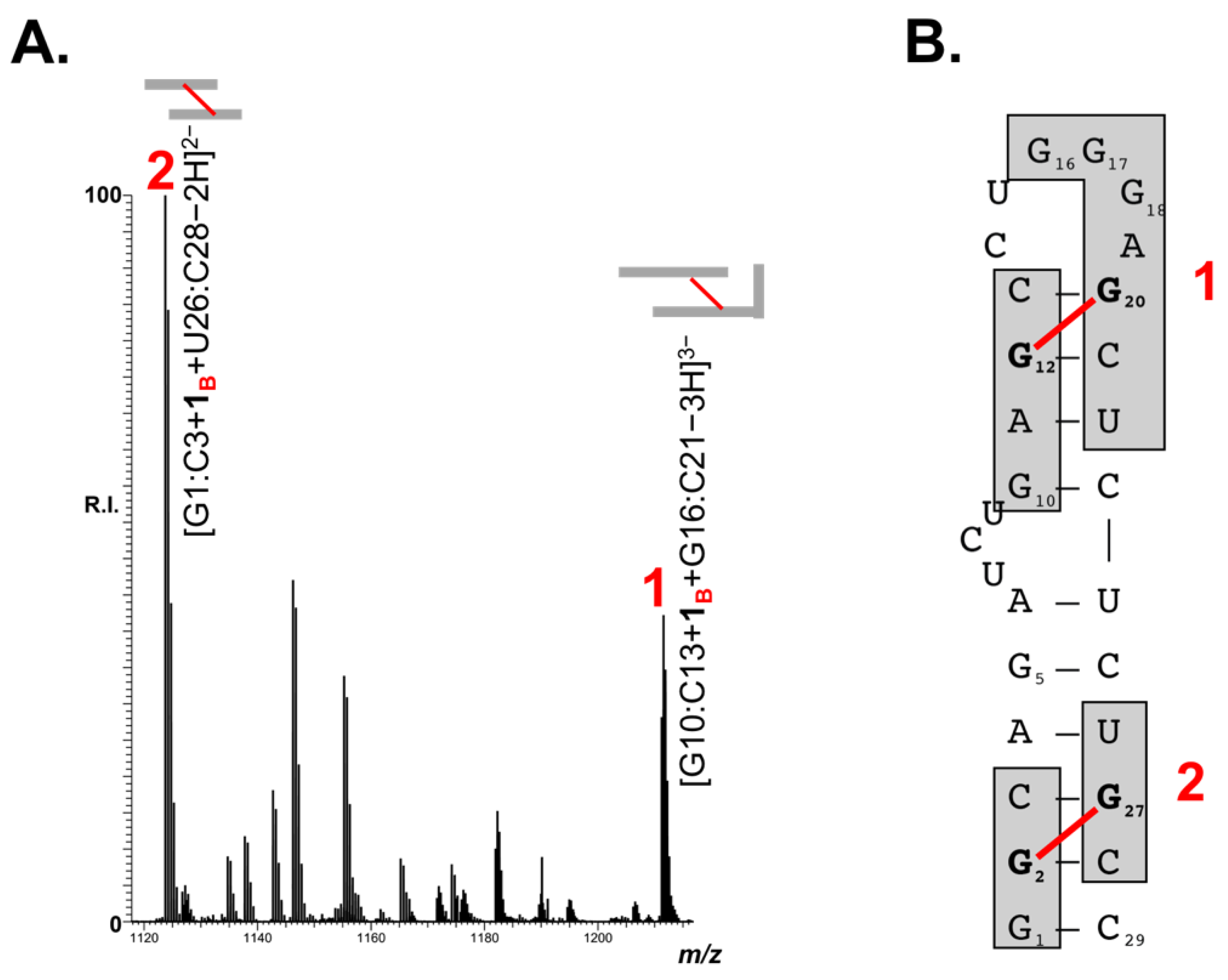
2.4. Adducts Mapping on TAR RNA
2.5. B-CePs Inhibit NC-Mediated Melting of TAR RNA In Vitro
3. Materials and Methods
3.1. RNA Substrate and Protein
3.2. Chemical Reagents
3.3. Fluorescence Quenching Assay (FQA)
3.4. Gel Electrophoretic Analysis
3.5. Mass Spectrometric Analysis
3.6. Enzymatic Digestion of TAR RNA
3.7. High-Throughput Screening (HTS)
4. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Acknowledgments
Conflicts of Interest
Sample Availability
References
- Chavali, S.S.; Bonn-Breach, R.; Wedekind, J.E. Face-time with TAR: Portraits of an HIV-1 RNA with diverse modes of effector recognition relevant for drug discovery. J. Biol. Chem. 2019, 294, 9326–9341. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Disney, M.D. Targeting RNA with Small Molecules to Capture Opportunities at the Intersection of Chemistry, Biology, and Medicine. J. Am. Chem. Soc. 2019, 141, 6776–6790. [Google Scholar] [CrossRef]
- Hermann, T. Viral RNA Targets and Their Small Molecule Ligands. In RNA Therapeutics. Topics in Medicinal Chemistry; Springer: Cham, Swithzerland, 2017; Volume 27. [Google Scholar]
- Gallego, J.; Varani, G. Targeting RNA with small-molecule drugs: Therapeutic promise and chemical challenges. Acc. Chem. Res. 2001, 34, 836–843. [Google Scholar] [CrossRef] [PubMed]
- Foreyen, M.; Herdewijn, P. RNA as a target for drug design, the example of Tat-TAR interaction. Curr. Top. Med. Chem. 2002, 2, 1123–1145. [Google Scholar] [CrossRef] [PubMed]
- Connelly, C.M.; Moon, M.H.; Schneekloth, J.S., Jr. The Emerging Role of RNA as a Therapeutic Target for Small Molecules. Cell Chem. Biol. 2016, 23, 1077–1090. [Google Scholar] [CrossRef] [PubMed]
- Le Grice, S.F. Targeting the HIV RNA genome: High-hanging fruit only needs a longer ladder. Curr. Top. Microbiol. Immunol. 2015, 389, 147–169. [Google Scholar] [PubMed]
- Belfetmi, A.; Zargarian, L.; Tisne, C.; Sleiman, D.; Morellet, N.; Lescop, E.; Maskri, O.; Rene, B.; Mely, Y.; Fosse, P.; et al. Insights into the mechanisms of RNA secondary structure destabilization by the HIV-1 nucleocapsid protein. RNA 2016, 22, 506–517. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kanevsky, I.; Chaminade, F.; Ficheux, D.; Moumen, A.; Gorelick, R.; Negroni, M.; Darlix, J.L.; Fosse, P. Specific interactions between HIV-1 nucleocapsid protein and the TAR element. J. Mol. Biol. 2005, 348, 1059–1077. [Google Scholar] [CrossRef] [PubMed]
- Rana, T.M.; Jeang, K.T. Biochemical and functional interactions between HIV-1 Tat protein and TAR RNA. Arch. Biochem. Biophys. 1999, 365, 175–185. [Google Scholar] [CrossRef] [PubMed]
- Boudier, C.; Storchak, R.; Sharma, K.K.; Didier, P.; Follenius-Wund, A.; Muller, S.; Darlix, J.L.; Mely, Y. The mechanism of HIV-1 Tat-directed nucleic acid annealing supports its role in reverse transcription. J. Mol. Biol. 2010, 400, 487–501. [Google Scholar] [CrossRef] [PubMed]
- Weeks, K.M.; Ampe, C.; Schltz, S.C.; Steitz, T.A.; Crothers, D.M. Fragments of the HIV-1 Tat protein specifically bind TAR RNA. Science 1990, 249, 1281–1285. [Google Scholar] [CrossRef] [PubMed]
- Berkhout, B.; Silverman, R.H.; Jeang, K.T. Tat trans-activates the human immunodeficiency virus through a nascent RNA target. Cell 1989, 59, 273–282. [Google Scholar] [CrossRef]
- Frecentese, F.; Sosic, A.; Saccone, I.; Gamba, E.; Link, K.; Miola, A.; Cappellini, M.; Cattelan, M.G.; Severino, B.; Fiorino, F.; et al. Synthesis and in Vitro Screening of New Series of 2,6-Dipeptidyl-anthraquinones: Influence of Side Chain Length on HIV-1 Nucleocapsid Inhibitors. J. Med. Chem. 2016, 59, 1914–1924. [Google Scholar] [CrossRef] [Green Version]
- Sosic, A.; Cappellini, M.; Scalabrin, M.; Gatto, B. Nucleocapsid Annealing-Mediated Electrophoresis (NAME) assay allows the rapid identification of HIV-1 nucleocapsid inhibitors. J. Visual Exp. JoVE 2015, 95, 52474. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sosic, A.; Cappellini, M.; Sinigaglia, L.; Jacquet, R.; Deffieux, D.; Fabris, D.; Quideau, S.; Gatto, B. Polyphenolic C-glucosidic ellagitannins present in oak-aged wine inhibit HIV-1 nucleocapsid protein. Tetrahedron 2015, 71, 3020–3026. [Google Scholar] [CrossRef]
- Sosic, A.; Frecentese, F.; Perissutti, E.; Sinigaglia, L.; Santagada, V.; Caliendo, G.; Magli, E.; Ciano, A.; Zagotto, G.; Parolin, C.; et al. Design, synthesis and biological evaluation of TAR and cTAR binders as HIV-1 nucleocapsid inhibitors. MedChemComm 2013, 4, 1388–1393. [Google Scholar] [CrossRef]
- Sosic, A.; Saccone, I.; Carraro, C.; Kenderdine, T.; Gamba, E.; Caliendo, G.; Corvino, A.; Di Vaio, P.; Fiorino, F.; Magli, E.; et al. Non-Natural Linker Configuration in 2,6-Dipeptidyl-Anthraquinones Enhances the Inhibition of TAR RNA Binding/Annealing Activities by HIV-1 NC and Tat Proteins. Bioconjug. Chem. 2018, 29, 2195–2207. [Google Scholar] [CrossRef] [PubMed]
- Sosic, A.; Sinigaglia, L.; Cappellini, M.; Carli, I.; Parolin, C.; Zagotto, G.; Sabatino, G.; Rovero, P.; Fabris, D.; Gatto, B. Mechanisms of HIV-1 Nucleocapsid Protein Inhibition by Lysyl-Peptidyl-Anthraquinone Conjugates. Bioconjug. Chem. 2016, 27, 247–256. [Google Scholar] [CrossRef]
- Mori, M.; Kovalenko, L.; Lyonnais, S.; Antaki, D.; Torbett, B.E.; Botta, M.; Mirambeau, G.; Mely, Y. Nucleocapsid Protein: A Desirable Target for Future Therapies Against HIV-1. Curr. Top. Microbiol Immunol. 2015, 389, 53–92. [Google Scholar]
- Helbing, T.; Carraro, C.; Francke, A.; Sosic, A.; De Franco, M.; Gandin, V.; Gottlich, R.; Gatto, B. Aromatic Linkers Unleash the Antiproliferative Potential of 3-Chloropiperidines against Pancreatic Cancer Cells. ChemMedChem 2020, 15, 2040–2051. [Google Scholar] [CrossRef] [PubMed]
- Sosic, A.; Zuravka, I.; Schmitt, N.K.; Miola, A.; Gottlich, R.; Fabris, D.; Gatto, B. Direct and Topoisomerase II Mediated DNA Damage by Bis-3-chloropiperidines: The Importance of Being an Earnest, G. ChemMedChem 2017, 12, 1471–1479. [Google Scholar] [CrossRef] [PubMed]
- Zuravka, I.; Roesmann, R.; Sosic, A.; Gottlich, R.; Gatto, B. Bis-3-chloropiperidines containing bridging lysine linkers: Influence of side chain structure on DNA alkylating activity. Bioorg. Med. Chem. 2015, 23, 1241–1250. [Google Scholar] [CrossRef] [PubMed]
- Zuravka, I.; Roesmann, R.; Sosic, A.; Wende, W.; Pingoud, A.; Gatto, B.; Gottlich, R. Synthesis and DNA cleavage activity of Bis-3-chloropiperidines as alkylating agents. ChemMedChem 2014, 9, 2178–2185. [Google Scholar] [CrossRef] [PubMed]
- Zuravka, I.; Sosic, A.; Gatto, B.; Gottlich, R. Synthesis and evaluation of a bis-3-chloropiperidine derivative incorporating an anthraquinone pharmacophore. Bioorg. Med. Chem. Lett. 2015, 25, 4606–4609. [Google Scholar] [CrossRef] [PubMed]
- Carraro, C.; Helbing, T.; Francke, A.; Zuravka, I.; Sosic, A.; De Franco, M.; Gandin, V.; Gatto, B.; Gottlich, D.R. Appended Aromatic Moieties in Flexible Bis-3-chloropiperidines Confer Tropism against Pancreatic Cancer Cells. ChemMedChem 2021, 16, 860–868. [Google Scholar] [CrossRef]
- Shvadchak, V.; Sanglier, S.; Rocle, S.; Villa, P.; Haiech, J.; Hibert, M.; Van Dorsselaer, A.; Mély, Y.; de Rocquigny, H. Identification by high throughput screening of small compounds inhibiting the nucleic acid destabilization activity of the HIV-1 nucleocapsid protein. Biochimie 2009, 91, 916–923. [Google Scholar] [CrossRef] [PubMed]
- Bernacchi, S.; Stoylov, S.; Piemont, E.; Ficheux, D.; Roques, B.P.; Darlix, J.L.; Mely, Y. HIV-1 nucleocapsid protein activates transient melting of least stable parts of the secondary structure of TAR and its complementary sequence. J. Mol. Biol. 2002, 317, 385–399. [Google Scholar] [CrossRef]
- Turner, K.B.; Hagan, N.A.; Kohlway, A.S.; Fabris, D. Mapping noncovalent ligand binding to stemloop domains of the HIV-1 packaging signal by tandem mass spectrometry. J. Am. Soc. Mass Spectrom. 2006, 17, 1402–1411. [Google Scholar] [CrossRef] [Green Version]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Name | Description | Mass (u) [b] | |
|---|---|---|---|
| Experimental | Calculated | ||
| TAR | TAR oligoribonucleotide construct | 9286.25 | 9286.24 |
| TAR + 1B | bis-alkylation product of 1 on TAR | 9548.49 | 9548.48 |
| TAR + 1M | mono-alkylation product of 1 on TAR | 9566.49 | 9566.48 |
| Label | Bridged RNA Fragments | Symbol | Exp. Mass (u) | Calc. Mass (u) |
|---|---|---|---|---|
| 1 | G10:C13 + 1B + G16:C21 |  | 3636.73 | 3636.73 |
| 2 | G1:C3 + 1B + U26:C28 |  | 2249.52 | 2249.52 |
| B-CePs | Linker | IC50 a [μM] |
|---|---|---|
| 1 | -(CH2)3- | 4.75 ± 0.15 |
| 2 | -(CH2)5- | 2.75 ± 0.70 |
| 3 | -(CH2)6- | 3.55 ± 0.37 |
| 4 | -(CH2)5COOCH3 | 5.09 ± 0.05 |
| 5 | -(CH2)5COOCH3 | 4.78 ± 0.71 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Sosic, A.; Olivato, G.; Carraro, C.; Göttlich, R.; Fabris, D.; Gatto, B. Bis-3-Chloropiperidines Targeting TAR RNA as A Novel Strategy to Impair the HIV-1 Nucleocapsid Protein. Molecules 2021, 26, 1874. https://doi.org/10.3390/molecules26071874
Sosic A, Olivato G, Carraro C, Göttlich R, Fabris D, Gatto B. Bis-3-Chloropiperidines Targeting TAR RNA as A Novel Strategy to Impair the HIV-1 Nucleocapsid Protein. Molecules. 2021; 26(7):1874. https://doi.org/10.3390/molecules26071874
Chicago/Turabian StyleSosic, Alice, Giulia Olivato, Caterina Carraro, Richard Göttlich, Dan Fabris, and Barbara Gatto. 2021. "Bis-3-Chloropiperidines Targeting TAR RNA as A Novel Strategy to Impair the HIV-1 Nucleocapsid Protein" Molecules 26, no. 7: 1874. https://doi.org/10.3390/molecules26071874