
Melatonin Administration Accelerates Puberty Onset in Mice by Promoting FSH Synthesis

 , ,
, , 
Abstract
:1. Introduction
2. Results
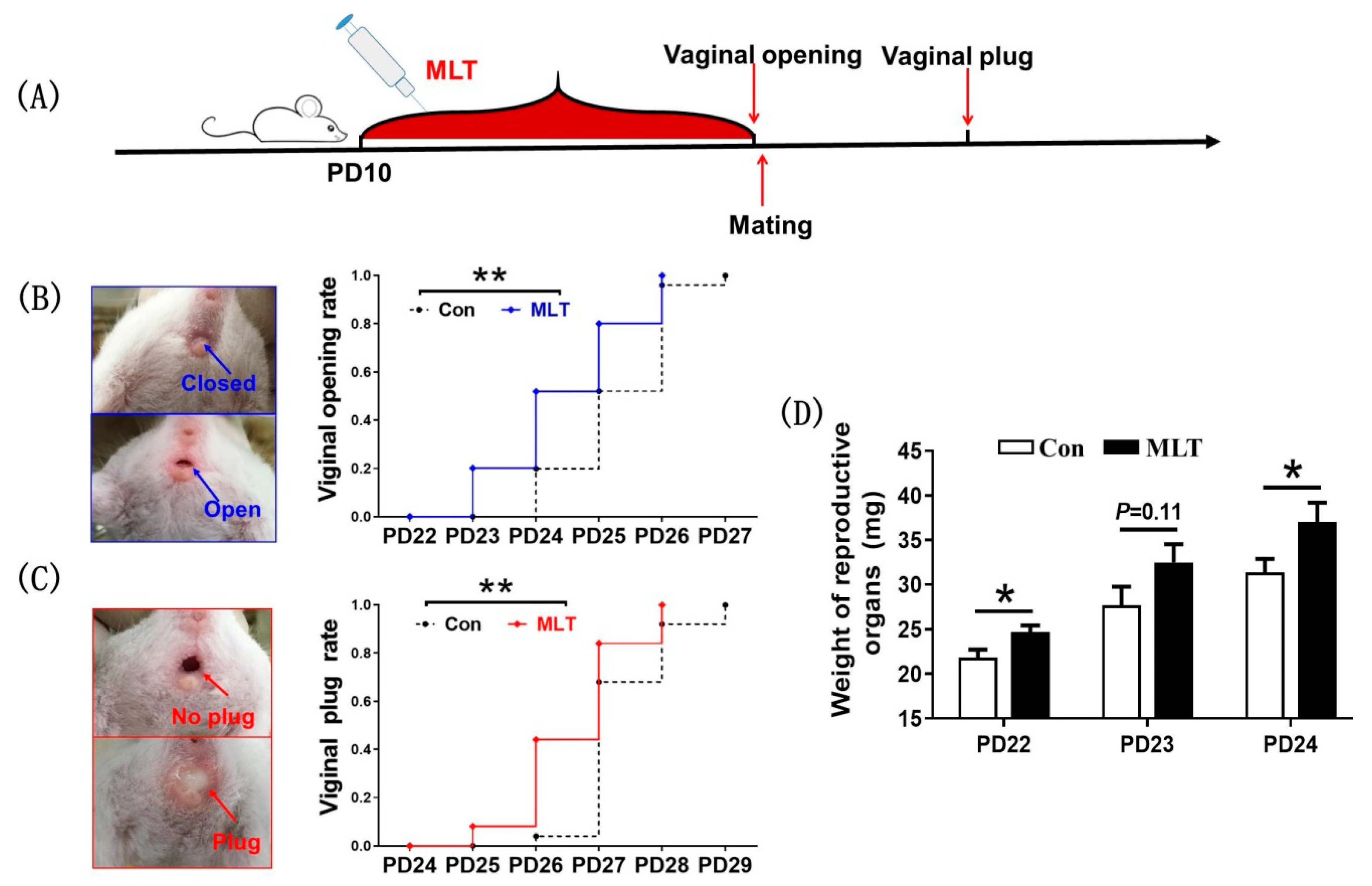
2.1. Melatonin Administration during Juvenile Stage Accelerated the Onset of Puberty
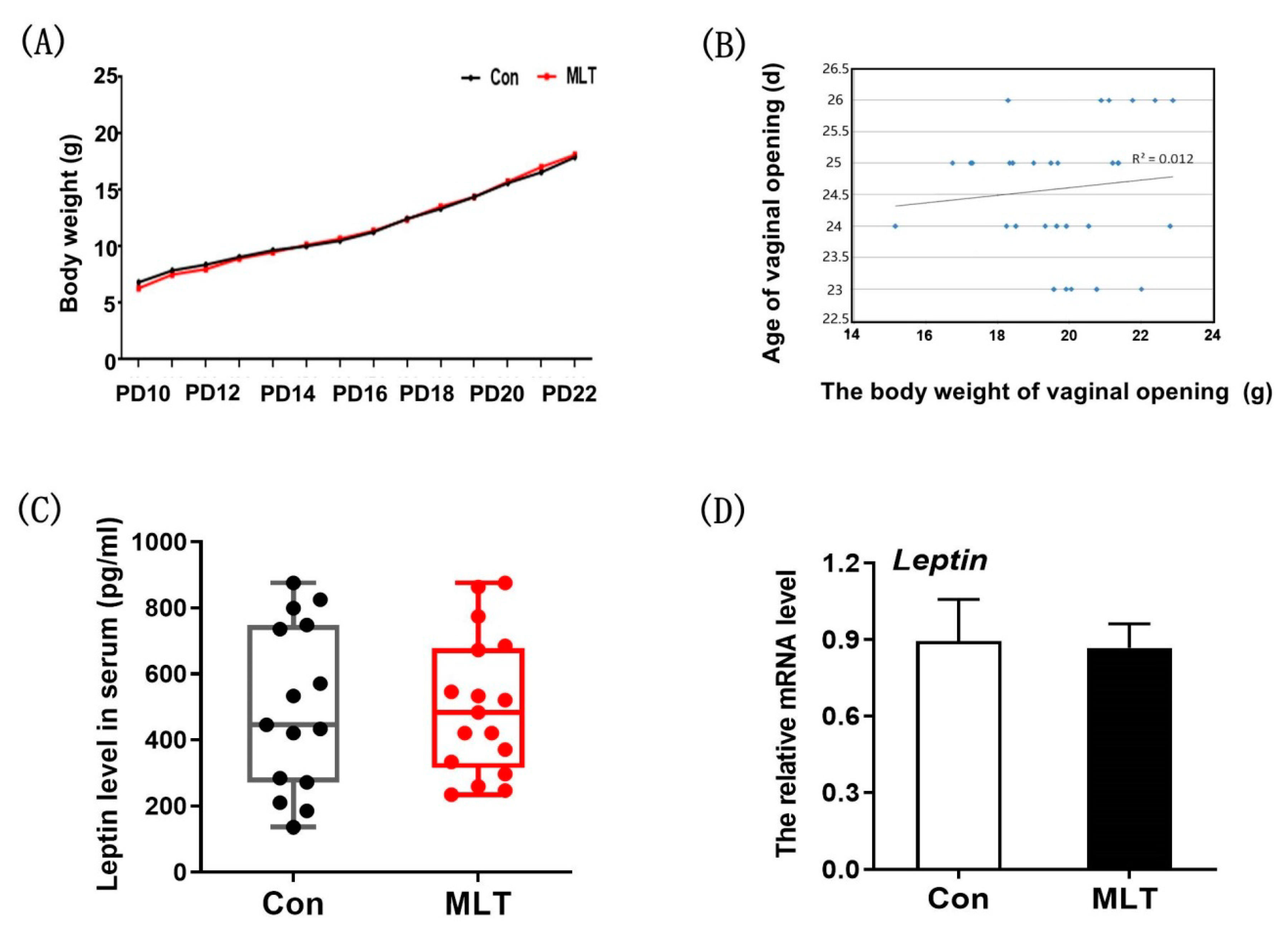
2.2. Melatonin Did Not Accelerate the Onset of Puberty by Physical Development and Leptin Level
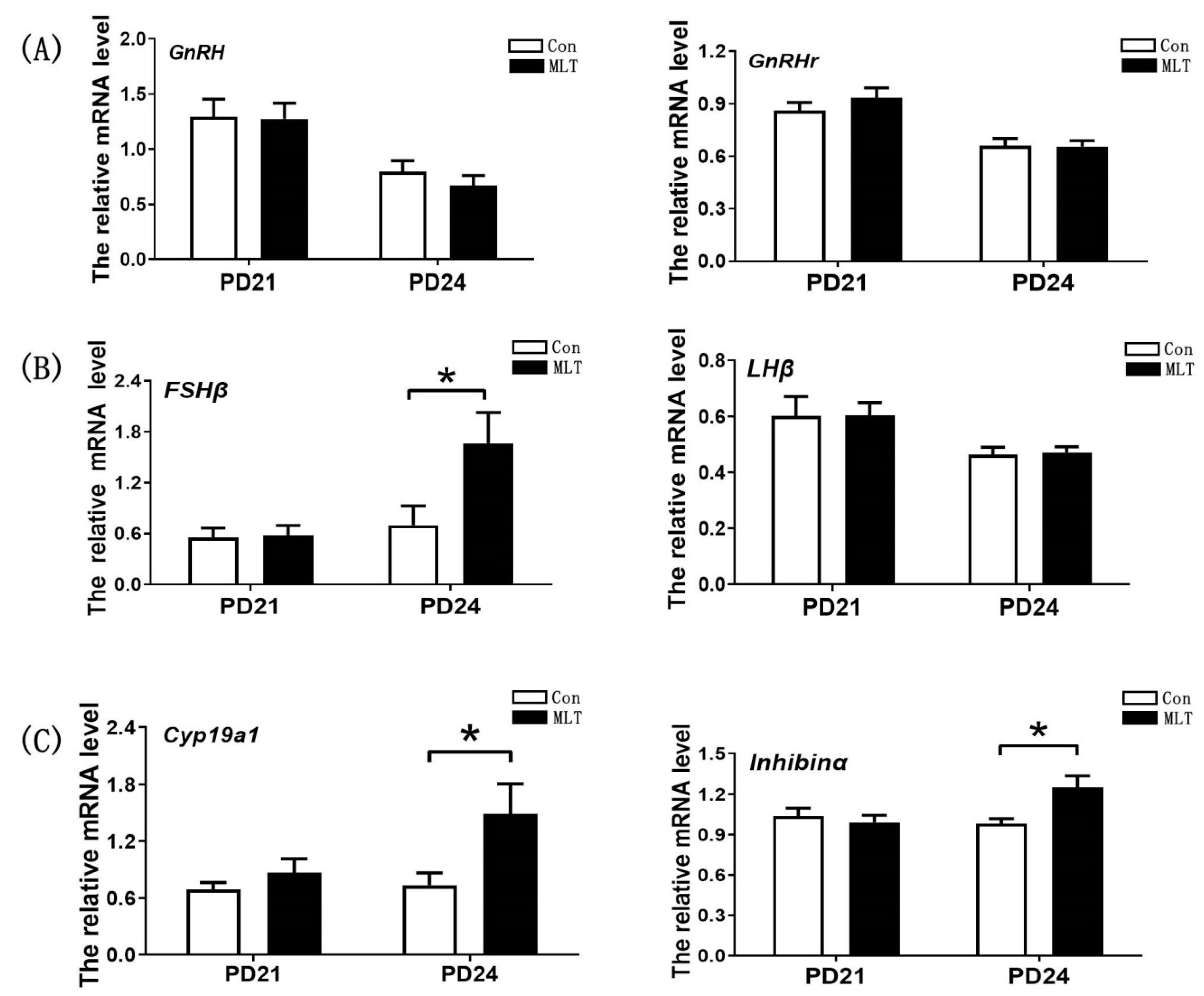
2.3. Melatonin Accelerated the Onset of Puberty by Enhancing the Secretion of FSH and Estrogen
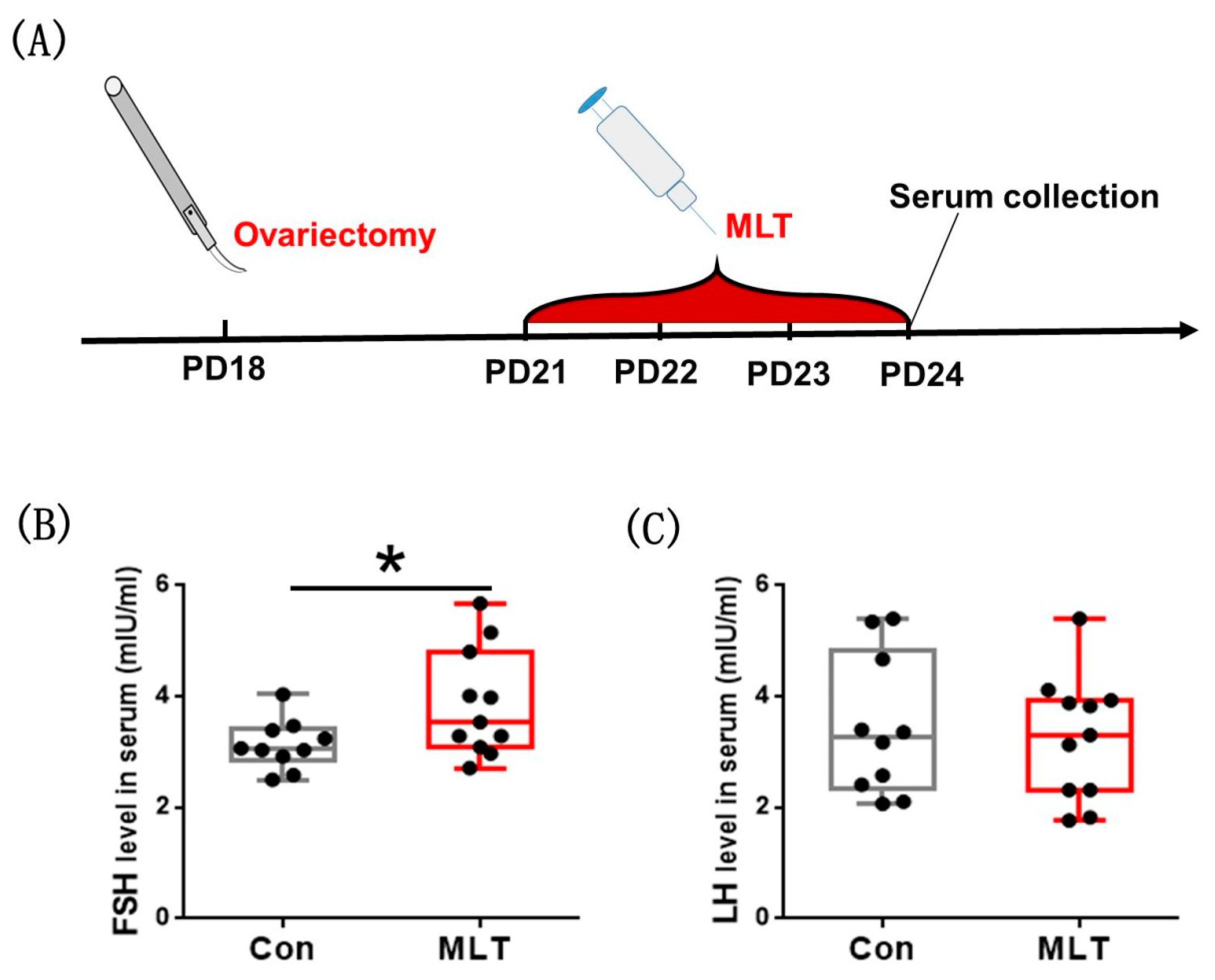
2.4. Melatonin Accelerated the Onset of Puberty by the Pituitary
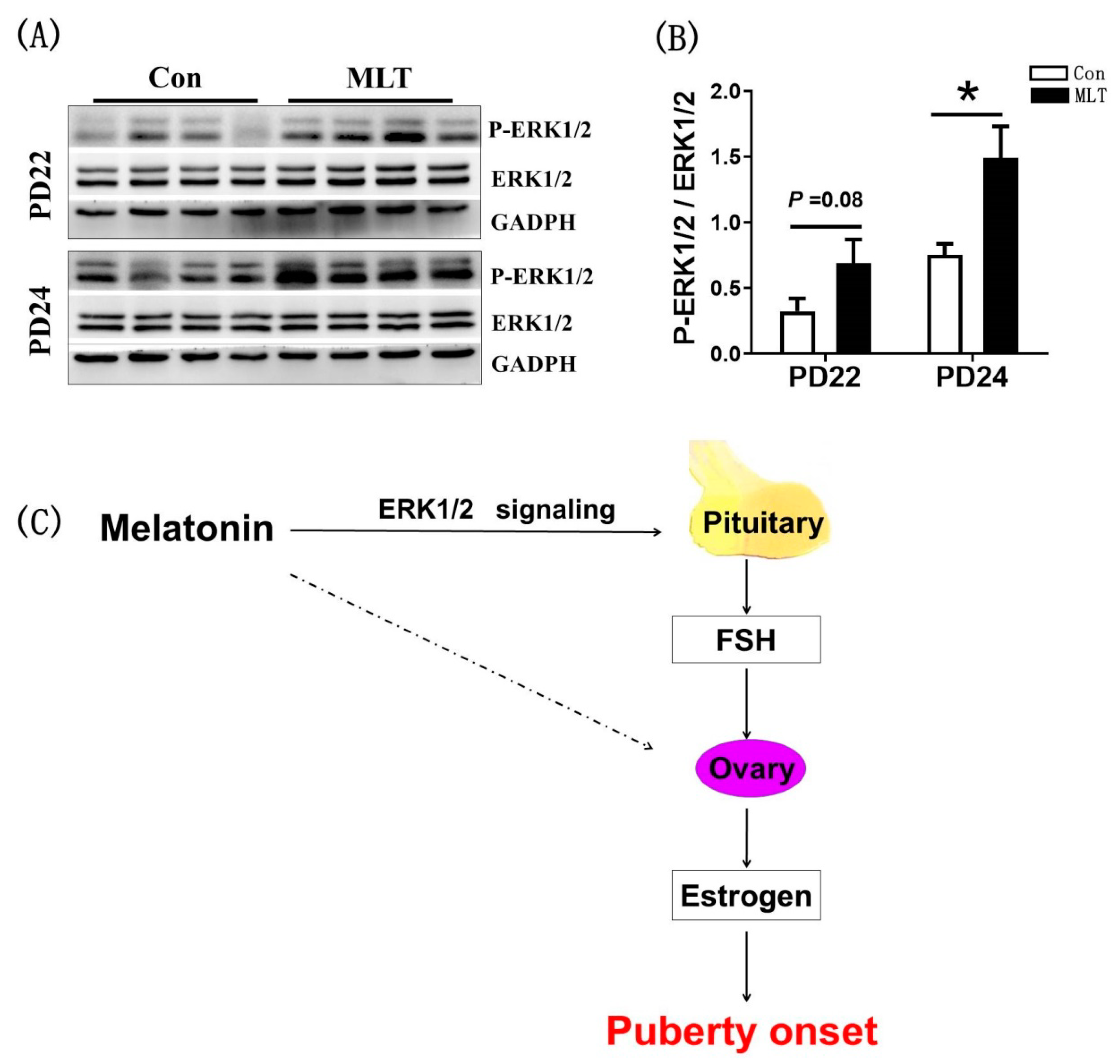
2.5. Melatonin Promoted the FSH Synthesis through the Pituitary ERK1/2 Signaling Pathway
3. Discussion
4. Materials and Methods
4.1. Ethics
4.2. Experimental Design and Animal Treatment
4.3. Hormone Determination
4.4. Real-Time Quantitative PCR Analysis
4.5. Western Blotting
4.6. Statistics Analysis
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
Sample Availability
References
- Gillette, M.U.; McArthur, A.J. Circadian actions of melatonin at the suprachiasmatic nucleus. Behav. Brain Res. 1996, 73, 135–139. [Google Scholar] [CrossRef] [Green Version]
- Reiter, R.J. The pineal and its hormones in the control of reproduction in mammals. Endocr. Rev. 1980, 1, 109–131. [Google Scholar] [CrossRef]
- Galano, A.; Tan, D.X.; Reiter, R.J. On the free radical scavenging activities of melatonin’s metabolites, AFMK and AMK. J. Pineal Res. 2013, 54, 245–257. [Google Scholar] [CrossRef]
- Day, D.; Burgess, C.M.; Kircik, L.H. Assessing the Potential Role for Topical Melatonin in an Antiaging Skin Regimen. J. Drugs Dermatol. 2018, 17, 966–969. [Google Scholar] [PubMed]
- Reiter, R.J.; Rosales-Corral, S.A.; Tan, D.X.; Acuna-Castroviejo, D.; Qin, L.; Yang, S.F.; Xu, K. Melatonin, a Full Service Anti-Cancer Agent: Inhibition of Initiation, Progression and Metastasis. Int. J. Mol. Sci. 2017, 18, 843. [Google Scholar] [CrossRef]
- Auld, F.; Maschauer, E.L.; Morrison, I.; Skene, D.J.; Riha, R.L. Evidence for the efficacy of melatonin in the treatment of primary adult sleep disorders. Sleep Med. Rev. 2017, 34, 10–22. [Google Scholar] [CrossRef] [PubMed]
- Tamura, H.; Takasaki, A.; Taketani, T.; Tanabe, M.; Lee, L.; Tamura, I.; Maekawa, R.; Aasada, H.; Yamagata, Y.; Sugino, N. Melatonin and female reproduction. J. Obstet. Gynaecol. Res. 2014, 40, 1–11. [Google Scholar] [CrossRef]
- Tamura, H.; Nakamura, Y.; Korkmaz, A.; Manchester, L.C.; Tan, D.X.; Sugino, N.; Reiter, R.J. Melatonin and the ovary: Physiological and pathophysiological implications. Fertil. Steril. 2009, 92, 328–343. [Google Scholar] [CrossRef] [PubMed]
- Woo, M.M.; Tai, C.J.; Kang, S.K.; Nathwani, P.S.; Pang, S.F.; Leung, P.C. Direct action of melatonin in human granulosa-luteal cells. J. Clin. Endocrinol. Metab. 2001, 86, 4789–4797. [Google Scholar] [CrossRef]
- Fang, L.; Li, Y.; Wang, S.; Yu, Y.; Li, Y.; Guo, Y.; Yan, Y.; Sun, Y.P. Melatonin induces progesterone production in human granulosa-lutein cells through upregulation of StAR expression. Aging 2019, 11, 9013–9024. [Google Scholar] [CrossRef]
- Dubocovich, M.L.; Yun, K.; Al-Ghoul, W.M.; Benloucif, S.; Masana, M.I. Selective MT2 melatonin receptor antagonists block melatonin-mediated phase advances of circadian rhythms. FASEB J. 1998, 12, 1211–1220. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jan, J.E.; Reiter, R.J.; Wong, P.K.; Bax, M.C.; Ribary, U.; Wasdell, M.B. Melatonin has membrane receptor-independent hypnotic action on neurons: An hypothesis. J. Pineal Res. 2011, 50, 233–240. [Google Scholar] [CrossRef] [PubMed]
- Suofu, Y.; Li, W.; Jean-Alphonse, F.G.; Jia, J. Dual role of mitochondria in producing melatonin and driving GPCR signaling to block cytochrome c release. Proc. Natl. Acad. Sci. USA 2017, 114, E7997–E8006. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- He, C.J.; Ma, T.; Shi, J.M.; Zhang, Z.Z.; Wang, J.; Zhu, K.F.; Li, Y.; Yang, M.H.; Song, Y.K.; Liu, G.S. Melatonin and its receptor MT1 are involved in the downstream reaction to luteinizing hormone and participate in the regulation of luteinization in different species. J. Pineal Res. 2016, 61, 279–290. [Google Scholar] [CrossRef]
- Adriaens, I.; Jacquet, P.; Cortvrindt, R.; Janssen, K.; Smitz, J. Melatonin has dose-dependent effects on folliculogenesis, oocyte maturation capacity and steroidogenesis. Toxicology 2006, 228, 333–343. [Google Scholar] [CrossRef]
- Cavallari, F.C.; Leal, C.L.V.; Zvi, R.; Hansen, P.J. Effects of melatonin on production of reactive oxygen species and developmental competence of bovine oocytes exposed to heat shock and oxidative stress during in vitro maturation. Zygote 2019, 27, 180–186. [Google Scholar] [CrossRef] [PubMed]
- Tian, X.Z.; Wang, F.; He, C.J.; Zhang, L.; Tan, D.X.; Reiter, R.J.; Xu, J.; Ji, P.Y.; Liu, G.S. Beneficial effects of melatonin on bovine oocytes maturation: A mechanistic approach. J. Pineal Res. 2014, 57, 239–247, Erratum in Zygote 2019, 27, 262. [Google Scholar] [CrossRef]
- Herbison, A.E. Control of puberty onset and fertility by gonadotropin-releasing hormone neurons. Nat. Rev. Endocrinol. 2016, 12, 452–466. [Google Scholar] [CrossRef] [PubMed]
- Waldhauser, F.; Weiszenbacher, G.; Tatzer, E.; Gisinger, B.; Waldhauser, M.; Schemper, M.; Frisch, H. Alterations in nocturnal serum melatonin levels in humans with growth and aging. J. Clin. Endocrinol. Metab. 1988, 66, 648–652. [Google Scholar] [CrossRef]
- Aleandri, V.; Spina, V.; Ciardo, A. Il ruolo della pineale nel controllo endocrino della pubertà [The role of the pineal body in the endocrine control of puberty. Minerva Ginecol. 1997, 49, 43–48. [Google Scholar]
- Vazquez, M.J.; Velasco, I.; Tena-Sempere, M. Novel mechanisms for the metabolic control of puberty: Implications for pubertal alterations in early-onset obesity and malnutrition. J. Endocrinol. 2019, 242, R51–R65. [Google Scholar] [CrossRef]
- Vogel, G. Leptin: A trigger for puberty? Science 1996, 274, 1466–1467. [Google Scholar] [CrossRef]
- Lv, D.Y.; Tan, T.; Zhu, T.Q.; Wang, J.; Zhang, S.H.; Zhang, L.; Hu, X.X.; Liu, G.S.; Xing, Y.M. Leptin mediates the effects of melatonin on female reproduction in mammals. J. Pineal Res. 2019, 66, e12559. [Google Scholar] [CrossRef] [PubMed]
- Van Geijlswijk, I.M.; Mol, R.H.; Egberts, T.C.; Smits, M.G. Evaluation of sleep, puberty and mental health in children with long-term melatonin treatment for chronic idiopathic childhood sleep onset insomnia. Psychopharmacology 2011, 216, 111–120. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Diekman, M.A.; Arthington, J.A.; Clapper, J.A.; Green, M.L. Failure of melatonin implants to alter onset of puberty in gilts. Anim. Reprod. Sci. 1997, 46, 283–288. [Google Scholar] [CrossRef]
- Pool, K.R.; Rickard, J.P.; de Graaf, S.P. Overcoming neuroendocrine and metabolic barriers to puberty: The role of melatonin in advancing puberty in ewe lambs. Domest. Anim. Endocrinol. 2020, 72, 106457. [Google Scholar] [CrossRef]
- Carcangiu, V.; Mura, M.C.; Bini, P.P.; Vacca, G.M.; Daga, C.; Luridiana, S. Can advance of first lambing induced by melatonin implants influence the next lambing time in Sarda breed sheep? Can. J. Anim. Sci. 2011, 92, 67–71. [Google Scholar] [CrossRef]
- Buchanan, K.L.; Yellon, S.M. Delayed puberty in the male djungarian hamster: Effect of short photoperiod or melatonin treatment on the gnrh neuronal system. Neuroendocrinology 1991, 54, 96–102. [Google Scholar] [CrossRef]
- Baker, E.R. Body weight and the initiation of puberty. Clin. Obstet. Gynecol. 1985, 28, 573–579. [Google Scholar] [CrossRef]
- Tomova, A.; Robeva, R.; Kumanov, P. Influence of the body weight on the onset and progression of puberty in boys. J. Pediatr. Endocr. Met. 2015, 28, 859–865. [Google Scholar] [CrossRef]
- Gimenez, F.; Stornelli, M.C.; Tittarelli, C.M.; Savignone, C.A.; Dorna, I.V.; de la Sota, R.L.; Stornelli, M.A. Suppression of estrus in cats with melatonin implants. Theriogenology 2009, 72, 493–499. [Google Scholar] [CrossRef]
- Chan, J.L.; Mantzoros, C.S. Leptin and the hypothalamic-pituitary regulation of the gonadotropin-gonadal axis. Pituitary 2001, 4, 87–92. [Google Scholar] [CrossRef]
- Pankov, Y.A. Kisspeptin and leptin in the regulation of fertility. Mol. Biol. 2015, 49, 707–715. [Google Scholar] [CrossRef]
- Jones, R.E.; Lopez, K.H. Endocrinology, Brain and Pituitary Gland. In Human Reproductive Biology, 3rd ed.; Maragioglio, N., Ed.; Academic Press: Cambridge, MA, USA, 2006; pp. 3–29. [Google Scholar]
- Nagirnaja, L.; Rull, K.; Uusküla, L.; Hallast, P.; Grigorova, M.; Laan, M. Genomics and genetics of gonadotropin beta-subunit genes: Unique FSHB and duplicated LHB/CGB loci. Mol. Cell. Endocrinol. 2010, 329, 4–16. [Google Scholar] [CrossRef]
- McNeilly, A.S.; Tsonis, C.G.; Baird, D.T. Inhibin. Hum. Reprod. 1988, 3, 45–49. [Google Scholar] [CrossRef] [PubMed]
- Simpson, E.R.; Mahendroo, M.S.; Means, G.D.; Kilgore, M.W.; Hinshelwood, M.M.; Graham, L.S.; Amarneh, B.; Ito, Y.; Fisher, C.R.; Michael, M.D.; et al. Aromatase cytochrome P450, the enzyme responsible for estrogen biosynthesis. Endocr. Rev. 1994, 15, 342–355. [Google Scholar] [PubMed]
- Kahnamouyi, S.; Nouri, M.; Farzadi, L.; Darabi, M.; Hosseini, V.; Mehdizadeh, A. The role of mitogen-activated protein kinase-extracellular receptor kinase pathway in female fertility outcomes: A focus on pituitary gonadotropins regulation. Ther. Adv. Endocrinol. Metab. 2018, 9, 209–215. [Google Scholar] [CrossRef]
- Huffman, L.J.; Inskeep, E.K.; Goodman, R.L. Changes in episodic luteinizing hormone secretion leading to puberty in the lamb. Biol. Reprod. 1987, 37, 755–761. [Google Scholar] [CrossRef] [Green Version]
- Mahdi, D.; Khallili, K. Relationship between follicle growth and circulating gonadotrophin levels during postnatal development of sheep. Anim. Reprod. Sci. 2008, 106, 100–112. [Google Scholar] [CrossRef] [PubMed]
- Jang, H.; Lee, O.H.; Lee, Y.; Yoon, H.; Chang, E.M.; Park, M.; Lee, J.W.; Hong, K.; Kim, J.O.; Kim, N.K.; et al. Melatonin prevents cisplatin-induced primordial follicle loss via suppression of PTEN/AKT/FOXO3a pathway activation in the mouse ovary. J. Pineal Res. 2016, 60, 336–347. [Google Scholar] [CrossRef] [PubMed]
- Jimenez-Caliani, A.J.; Jimenez-Jorge, S.; Molinero, P.; Rubio, A.; Guerrero, J.M.; Osuna, C. Treatment with testosterone or estradiol in melatonin treated females and males MRL/MpJ-Faslpr mice induces negative effects in developing systemic lupus erythematosus. J. Pineal Res. 2008, 45, 204–211. [Google Scholar] [CrossRef] [PubMed]
- Turkoz, Y.; Celik, O.; Hascalik, S.; Cigremis, Y.; Hascalik, M.; Mizrak, B.; Yologlu, S. Melatonin reduces torsion-detorsion injury in rat ovary: Biochemical and histopathologic evaluation. J. Pineal Res. 2004, 37, 137–141. [Google Scholar] [CrossRef] [PubMed]
- He, C.J.; Wang, J.; Li, Y.; Zhu, K.F.; Xu, Z.Y.; Song, Y.L.; Song, Y.K.; Liu, G.S. Melatonin-related genes expressed in the mouse uterus during early gestation promote embryo implantation. J. Pineal Res. 2015, 58, 300–309. [Google Scholar] [CrossRef] [PubMed]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Hormones | Postnatal Days | Con | MLT | df/t |
|---|---|---|---|---|
| FSH mIU/mL | PD21 | 1.37 ± 0.232 a | 1.33 ± 0.171 a | 38/0.151 |
| PD22 | 0.93 ± 0.156 a | 1.07 ± 0.170 a | 36/0.592 | |
| PD23 | 1.22 ± 0.138 a | 1.64 ± 0.118 b | 37/2.295 | |
| PD24 | 1.26 ± 0.130 a | 1.60 ± 0.096 b | 38/2.158 | |
| LH mIU/mL | PD21 | 4.82 ± 0.643 a | 4.17 ± 0.559 a | 36/0.768 |
| PD22 | 5.41 ± 0.682 a | 5.30 ± 0.496 a | 37/0.121 | |
| PD23 | 4.84 ± 0.455 a | 5.20 ± 0.592 a | 35/0.489 | |
| PD24 | 4.11 ± 0.598 a | 3.32 ± 0.450 a | 38/1.051 | |
| Estrogen pg/mL | PD21 | 11.96 ± 1.641 a | 18.08 ± 1.307 b | 34/2.942 |
| PD22 | 14.04 ± 1.109 a | 22.39 ± 2.128 b | 37/3.340 | |
| PD23 | 14.82 ± 1.959 a | 20.51 ± 2.370 a | 35/1.807 | |
| PD24 | 10.82 ± 1.103 a | 13.97 ± 1.074 b | 36/2.040 |
| Genes | Primer seq (5′–3′) | Size (bp) |
|---|---|---|
| Actb | F: CCAGCCTTCCTTCTTGGGTAT R: AGGTCTTTACGGATGTCAACG | 93 |
| Leptin | F: CAAGCAGTGCCTATCCAGAAA | 162 |
| R: GGACAAACTCAGAATGGGGTG | ||
| GnRH | F: AGGAAGCCAGGCAGAAGAAG | 100 |
| R: GAGCCATTAACAGGTCACAAGC | ||
| GnRHr | F: GATGGTGGTGATTAGCCTGGAC | 192 |
| R:CATTGCGAGAAGACTGTGGG | ||
| FSHβ | F: ACCACTCATCCCTCCATCCA | 166 |
| R: CCACTTTCCTTTCCTCCCTCTA | ||
| LHβ | F: CCCATAGTCTCCTTTCCTGTAGC | 97 |
| R: AGGCCATTGGTTGAGTCCTG | ||
| Cyp19a1 | F: GACACATCATGCTGGACACC | 179 |
| R: CAAGTCCTTGACGGATCGTT | ||
| Inhibinα | F: CTTTCCCTCTGCTGACCCA | 184 |
| R: AAAGCCGCAGGAGACCAA |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Yang, C.; Ran, Z.; Liu, G.; Hou, R.; He, C.; Liu, Q.; Chen, Y.; Liu, Y.; Wang, X.; Ling, C.; et al. Melatonin Administration Accelerates Puberty Onset in Mice by Promoting FSH Synthesis. Molecules 2021, 26, 1474. https://doi.org/10.3390/molecules26051474
Yang C, Ran Z, Liu G, Hou R, He C, Liu Q, Chen Y, Liu Y, Wang X, Ling C, et al. Melatonin Administration Accelerates Puberty Onset in Mice by Promoting FSH Synthesis. Molecules. 2021; 26(5):1474. https://doi.org/10.3390/molecules26051474
Chicago/Turabian StyleYang, Chan, Zaohong Ran, Guoshi Liu, Rong Hou, Changjiu He, Qinghua Liu, Yingjun Chen, Yuliang Liu, Xiaodong Wang, Chuqi Ling, and et al. 2021. "Melatonin Administration Accelerates Puberty Onset in Mice by Promoting FSH Synthesis" Molecules 26, no. 5: 1474. https://doi.org/10.3390/molecules26051474