
Potential Therapeutic Targets of Quercetin, a Plant Flavonol, and Its Role in the Therapy of Various Types of Cancer through the Modulation of Various Cell Signaling Pathways

 , , , ,
, , , ,  , and
, and
Abstract
:1. Introduction
2. Major Mechanism of Qu in Management of Cancer
2.1. Inflammation
2.2. Reactive Oxygen Species (ROS)
2.3. Angiogenesis
2.4. Apoptosis
2.5. Cell Cycle
2.6. Tumor Suppressor Gene
2.7. Phosphatidylinositide-3-Kinase (PI3K)/Akt Pathways
2.8. Signal Transducer and Activator of Transcription 3 (STAT3)
2.9. Epidermal Growth Factor Receptor (EGFR)
3. Role of Qu in Prevention and Inhibition of Various Types of Cancer
3.1. Cervical Cancer
3.2. Breast Cancer
3.3. Ovarian Cancer
3.4. Endometrial Cancer
3.5. Pancreatic Cancer
3.6. Gastric Cancer
3.7. Liver Cancer
3.8. Colon Cancer
3.9. Renal Cancer
3.10. Prostate Cancer
3.11. Urinary Bladder Cancer
3.12. Leukemia
3.13. Lymphoma
3.14. Lung Cancer
3.15. Osteosarcoma
3.16. Brain Tumor
3.17. Head and Neck Cancer
3.18. Oral Cancer
3.19. Esophageal Cancer
3.20. Melanoma
3.21. Myeloma
3.22. Retinoblastoma
4. Bioavailability of Qu
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Bray, F.; Ferlay, J.; Soerjomataram, I.; Siegel, R.L.; Torre, L.A.; Jemal, A. Global cancer statistics 2018: GLOBOCAN estimates of incidence and mortality worldwide for 36 cancers in 185 countries. CA Cancer J. Clin. 2018, 68, 394–424. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- World Health Organization. Cancer 2020. Available online: https://www.who.int/news-room/fact-sheets/detail/cancer (accessed on 5 October 2019).
- Wu, S.; Zhu, W.; Thompson, P.; Hannun, Y.A. Evaluating intrinsic and non-intrinsic cancer risk factors. Nat. Commun. 2018, 9, 3490. [Google Scholar] [CrossRef]
- Ames, B.N. Mutagenesis and carcinogenesis: Endogenous and exogenous factors. Environ. Mol. Mutagen. 1989, 14, 66–77. [Google Scholar] [CrossRef] [PubMed]
- Hussain, S.; Harris, C.C. Molecular epidemiology and carcinogenesis: Endogenous and exogenous carcinogens. Mutat. Res. Mutat. Res. 2000, 462, 311–322. [Google Scholar] [CrossRef]
- Hanahan, D.; Weinberg, R.A. The Hallmarks of Cancer. Cell 2000, 100, 57–70. [Google Scholar] [CrossRef] [Green Version]
- Hanahan, D.; Weinberg, R.A. Hallmarks of Cancer: The Next Generation. Cell 2011, 144, 646–674. [Google Scholar] [CrossRef] [Green Version]
- Pollack, L.A.; Rowland, J.H.; Crammer, C.; Stefanek, M. Introduction: Charting the landscape of cancer survivors’ health-related outcomes and care. Cancer 2009, 115, 4265–4269. [Google Scholar] [CrossRef] [PubMed]
- Baskar, R.; Lee, K.A.; Yeo, R.; Yeoh, K.-W. Cancer and Radiation Therapy: Current Advances and Future Directions. Int. J. Med. Sci. 2012, 9, 193–199. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kashyap, D.; Garg, V.K.; Tuli, H.S.; Yerer, M.B.; Sak, K.; Sharma, A.K.; Kumar, M.; Aggarwal, V.; Sandhu, S.S. Fisetin and Quercetin: Promising Flavonoids with Chemopreventive Potential. Biomolecules 2019, 9, 174. [Google Scholar] [CrossRef] [Green Version]
- Wu, H.; Xie, J.; Pan, Q.; Wang, B.; Hu, D.; Hu, X. Anticancer Agent Shikonin Is an Incompetent Inducer of Cancer Drug Resistance. PLoS ONE 2013, 8, e52706. [Google Scholar] [CrossRef]
- Verellen, D.; De Ridder, M.; Linthout, N.; Tournel, K.; Soete, G.; Storme, G. Innovations in image-guided radiotherapy. Nat. Rev. Cancer 2007, 7, 949–960. [Google Scholar] [CrossRef]
- Chabner, B.A.; Roberts, T.G. Chemotherapy and the war on cancer. Nat. Rev. Cancer 2005, 5, 65–72. [Google Scholar] [CrossRef] [PubMed]
- Palucka, K.; Banchereau, J. Cancer immunotherapy via dendritic cells. Nat. Rev. Cancer 2012, 12, 265–277. [Google Scholar] [CrossRef] [PubMed]
- Tang, S.-M.; Deng, X.-T.; Zhou, J.; Li, Q.-P.; Ge, X.-X.; Miao, L. Pharmacological basis and new insights of quercetin action in respect to its anti-cancer effects. Biomed. Pharmacother. 2020, 121, 109604. [Google Scholar] [CrossRef]
- Weatherall, D.; Greenwood, B.; Chee, H.L.; Wasi, P. Science and Technology for Disease Control: Past, Present, and Future. In Disease Control Priorities in Developing Countries, 2nd ed.; Jamison, D.T., Breman, J.G., Measham, A.R., Alleyne, G., Claeson, M., Evans, D.B., Jha, P., Mills, A., Musgrove, P., Eds.; World Bank: Washington, DC, USA, 2006. [Google Scholar]
- Kennedy, D.A.; Hart, J.; Seely, D. Cost Effectiveness of Natural Health Products: A Systematic Review of Randomized Clinical Trials. Evid. Based Complementary Altern. Med. 2009, 6, 297–304. [Google Scholar] [CrossRef]
- Downer, S.M.; Cody, M.M.; McCluskey, P.; Wilson, P.D.; Arnott, S.J.; Lister, T.A.; Slevin, M.L. Pursuit and practice of complementary therapies by cancer patients receiving conventional treatment. BMJ 1994, 309, 86–89. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yates, J.S.; Mustian, K.M.; Morrow, G.R.; Gillies, L.J.; Padmanaban, D.; Atkins, J.N.; Issell, B.; Kirshner, J.J.; Colman, L.K. Prevalence of complementary and alternative medicine use in cancer patients during treatment. Support. Care Cancer 2005, 13, 806–811. [Google Scholar] [CrossRef]
- Greenwald, P. Chemoprevention of Cancer. Sci. Am. 1996, 275, 96–99. [Google Scholar] [CrossRef] [PubMed]
- Danaei, G.; Hoorn, S.V.; Lopez, A.D.; Murray, C.J.; Ezzati, M. Causes of cancer in the world: Comparative risk assessment of nine behavioural and environmental risk factors. Lancet 2005, 366, 1784–1793. [Google Scholar] [CrossRef] [Green Version]
- Knekt, P.; Kumpulainen, J.; Järvinen, R.; Rissanen, H.; Heliövaara, M.; Reunanen, A.; Hakulinen, T.; Aromaa, A. Flavonoid intake and risk of chronic diseases. Am. J. Clin. Nutr. 2002, 76, 560–568. [Google Scholar] [CrossRef] [Green Version]
- Anand, P.; Kunnumakkara, A.B.; Sundaram, C.; Harikumar, K.B.; Tharakan, S.T.; Lai, O.S.; Sung, B.; Aggarwal, B.B. Cancer is a Preventable Disease that Requires Major Lifestyle Changes. Pharm. Res. 2008, 25, 2200. [Google Scholar] [CrossRef] [Green Version]
- Irigaray, P.; Newby, J.; Clapp, R.; Hardell, L.; Howard, V.; Montagnier, L.; Epstein, S.; Belpomme, D. Lifestyle-related factors and environmental agents causing cancer: An overview. Biomed. Pharmacother. 2007, 61, 640–658. [Google Scholar] [CrossRef] [PubMed]
- Flores, I.R.; Vásquez-Murrieta, M.S.; Franco-Hernández, M.O.; Márquez-Herrera, C.E.; Ponce-Mendoza, A.; López-Cortéz, M.D.S. Bioactive compounds in tomato (Solanum lycopersicum) variety saladette and their relationship with soil mineral content. Food Chem. 2021, 344, 128608. [Google Scholar] [CrossRef] [PubMed]
- Torres, N.; Martínez-Lüscher, J.; Porte, E.; Yu, R.; Kurtural, S.K. Impacts of leaf removal and shoot thinning on cumulative daily light intensity and thermal time and their cascading effects of grapevine (Vitis vinifera L.) berry and wine chemistry in warm climates. Food Chem. 2021, 343, 128447. [Google Scholar] [CrossRef] [PubMed]
- Giuffrè, A.M.; Zappia, C.; Capocasale, M. Physico-chemical stability of blood orange juice during frozen storage. Int. J. Food Prop. 2017, 20, 1930–1943. [Google Scholar]
- Santiago, B.; Calvo, A.A.; Gullón, B.; Feijoo, G.; Moreira, M.T.; González-García, S. Production of flavonol quercetin and fructooligosaccharides from onion (Allium cepa L.) waste: An environmental life cycle approach. Chem. Eng. J. 2020, 392, 123772. [Google Scholar] [CrossRef]
- Ribes-Moya, A.M.; Adalid, A.M.; Raigón, M.D.; Hellín, P.; Fita, A.; Rodríguez-Burruezo, A. Variation in flavonoids in a collection of peppers (Capsicum sp.) under organic and conventional cultivation: Effect of the genotype, ripening stage, and growing system. J. Sci. Food Agric. 2020, 100, 2208–2223. [Google Scholar] [CrossRef]
- Sun, J.; Janisiewicz, W.J.; Takeda, F.; Evans, B.; Ii, W.M.J.; Zhang, M.; Yu, L.; Chen, P. Effect of nighttime UV-C irradiation of strawberry plants on phenolic content of fruit: Targeted and non-targeted metabolomic analysis. J. Berry Res. 2020, 10, 365–380. [Google Scholar] [CrossRef]
- Zhou, W.; Liang, X.; Dai, P.; Chen, Y.; Zhang, Y.; Zhang, M.; Lu, L.; Jin, C.; Lin, X. Alteration of Phenolic Composition in Lettuce (Lactuca sativa L.) by Reducing Nitrogen Supply Enhances its Anti-Proliferative Effects on Colorectal Cancer Cells. Int. J. Mol. Sci. 2019, 20, 4205. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Formica, J.; Regelson, W. Review of the biology of quercetin and related bioflavonoids. Food Chem. Toxicol. 1995, 33, 1061–1080. [Google Scholar] [CrossRef]
- Nugroho, A.; Heryani, H.; Choi, J.S.; Park, H.-J. Identification and quantification of flavonoids in Carica papaya leaf and peroxynitrite-scavenging activity. Asian Pac. J. Trop. Biomed. 2017, 7, 208–213. [Google Scholar] [CrossRef]
- Bolling, B.W.; McKay, D.L.; Blumberg, J.B. The phytochemical composition and antioxidant actions of tree nuts. Asia Pac. J. Clin. Nutr. 2010, 19, 117–123. [Google Scholar]
- Kris-Etherton, P.M.; Hecker, K.D.; Bonanome, A.; Coval, S.M.; E Binkoski, A.; Hilpert, K.F.; E Griel, A.; Etherton, T.D. Bioactive compounds in foods: Their role in the prevention of cardiovascular disease and cancer. Am. J. Med. 2002, 113, 71–88. [Google Scholar] [CrossRef]
- Falcão, S.I.; Vale, N.; Gomes, P.; Domingues, M.R.M.; Freire, C.; Cardoso, S.M.; Vilas-Boas, M. Phenolic Profiling of Portuguese Propolis by LC-MS Spectrometry: Uncommon Propolis Rich in Flavonoid Glycosides. Phytochem. Anal. 2013, 24, 309–318. [Google Scholar] [CrossRef] [Green Version]
- Beecher, G.R.; Warden, B.A.; Merken, H. Analysis of Tea Polyphenols. Proc. Soc. Exp. Boil. Med. 1999, 220, 267–270. [Google Scholar] [CrossRef]
- Williamson, G.; Day, A.J.; Plumb, G.W.; Couteau, D. Human metabolic pathways of dietary flavonoids and cinnamates. Biochem. Soc. Trans. 2000, 28, 16–22. [Google Scholar] [CrossRef] [PubMed]
- Rafiq, R.A.; Quadri, A.; Nazir, L.A.; Peerzada, K.; Ganai, B.A.; Tasduq, S.A. A Potent Inhibitor of Phosphoinositide 3-Kinase (PI3K) and Mitogen Activated Protein (MAP) Kinase Signalling, Quercetin (3, 3’, 4’, 5, 7-Pentahydroxyflavone) Promotes Cell Death in Ultraviolet (UV)-B-Irradiated B16F10 Melanoma Cells. PLoS ONE 2015, 10, e0131253. [Google Scholar] [CrossRef] [Green Version]
- Lakhanpal, P.; Rai, D.K. Quercetin: A Versatile Flavonoid. Internet J. Med. Updat. EJOURNAL 2007, 2. [Google Scholar] [CrossRef] [Green Version]
- Hirpara, K.V.; Aggarwal, P.; Mukherjee, A.J.; Joshi, N.; Burman, A.C. Quercetin and Its Derivatives: Synthesis, Pharmacological Uses with Special Emphasis on Anti-Tumor Properties and Prodrug with Enhanced Bio-Availability. Anti-Cancer Agents Med. Chem. 2009, 9, 138–161. [Google Scholar] [CrossRef]
- Boots, A.W.; Haenen, G.R.; Bast, A. Health effects of quercetin: From antioxidant to nutraceutical. Eur. J. Pharmacol. 2008, 585, 325–337. [Google Scholar] [CrossRef]
- Vafadar, A.; Shabaninejad, Z.; Movahedpour, A.; Fallahi, F.; Taghavipour, M.; Ghasemi, Y.; Akbari, M.; Shafiee, A.; Hajighadimi, S.; Moradizarmehri, S.; et al. Quercetin and cancer: New insights into its therapeutic effects on ovarian cancer cells. Cell Biosci. 2020, 10, 1–17. [Google Scholar] [CrossRef] [Green Version]
- Shanmugam, M.K.; Lee, J.H.; Chai, E.Z.P.; Kanchi, M.M.; Kar, S.; Arfuso, F.; Dharmarajan, A.; Kumar, A.P.; Ramar, P.S.; Looi, C.Y.; et al. Cancer prevention and therapy through the modulation of transcription factors by bioactive natural compounds. Semin. Cancer Biol. 2016, 40–41, 35–47. [Google Scholar] [CrossRef]
- Shanmugam, M.K.; Kannaiyan, R.; Sethi, G. Targeting Cell Signaling and Apoptotic Pathways by Dietary Agents: Role in the Prevention and Treatment of Cancer. Nutr. Cancer 2011, 63, 161–173. [Google Scholar] [CrossRef] [PubMed]
- Van Ginkel, P.R.; Yan, M.B.; Bhattacharya, S.; Polans, A.S.; Kenealey, J.D. Natural products induce a G protein-mediated calcium pathway activating p53 in cancer cells. Toxicol. Appl. Pharmacol. 2015, 288, 453–462. [Google Scholar] [CrossRef] [Green Version]
- Lugli, E.; Ferraresi, R.; Roat, E.; Troiano, L.; Pinti, M.; Nasi, M.; Nemes, E.; Bertoncelli, L.; Gibellini, L.; Salomoni, P.; et al. Quercetin inhibits lymphocyte activation and proliferation without inducing apoptosis in peripheral mononuclear cells. Leuk. Res. 2009, 33, 140–150. [Google Scholar] [CrossRef]
- Kashyap, D.; Tuli, H.S.; Garg, V.K.; Goel, N.; Bishayee, A. Oncogenic and Tumor-Suppressive Roles of MicroRNAs with Special Reference to Apoptosis: Molecular Mechanisms and Therapeutic Potential. Mol. Diagn. Ther. 2018, 22, 179–201. [Google Scholar] [CrossRef] [PubMed]
- Zhuo, W.; Zhang, L.; Zhu, Y.; Zhu, B.; Chen, Z. Fisetin, a dietary bioflavonoid, reverses acquired Cisplatin-resistance of lung adenocarcinoma cells through MAPK/Survivin/Caspase pathway. Am. J. Transl. Res. 2015, 7, 2045–2052. [Google Scholar] [PubMed]
- Darband, S.G.; Sadighparvar, S.; Yousefi, B.; Kaviani, M.; Ghaderi-Pakdel, F.; Mihanfar, A.; Rahimi, Y.; Mobaraki, K.; Majidinia, M. Quercetin attenuated oxidative DNA damage through NRF2 signaling pathway in rats with DMH induced colon carcinogenesis. Life Sci. 2020, 253, 117584. [Google Scholar] [CrossRef]
- Ferrero-Miliani, L.; Nielsen, O.H.; Andersen, P.S.; Girardin, S.E. Chronic inflammation: Importance of NOD2 and NALP3 in interleukin-1? generation. Clin. Exp. Immunol. 2006, 147, 227–235. [Google Scholar] [CrossRef]
- Warren, C.A.; Paulhill, K.J.; Davidson, L.A.; Lupton, J.R.; Taddeo, S.S.; Hong, M.Y.; Carroll, R.J.; Chapkin, R.S.; Turner, N.D. Quercetin May Suppress Rat Aberrant Crypt Foci Formation by Suppressing Inflammatory Mediators That Influence Proliferation and Apoptosis. J. Nutr. 2008, 139, 101–105. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Xiao, X.; Shi, D.; Liu, L.; Wang, J.; Xie, X.; Kang, T.; Deng, W. Quercetin Suppresses Cyclooxygenase-2 Expression and Angiogenesis through Inactivation of P300 Signaling. PLoS ONE 2011, 6, e22934. [Google Scholar] [CrossRef]
- Wu, Y.; Antony, S.; Meitzler, J.L.; Doroshow, J.H. Molecular mechanisms underlying chronic inflammation-associated cancers. Cancer Lett. 2014, 345, 164–173. [Google Scholar] [CrossRef] [Green Version]
- Todoric, J.; Antonucci, L.; Karin, M. Targeting Inflammation in Cancer Prevention and Therapy. Cancer Prev. Res. 2016, 9, 895–905. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chien, S.-Y.; Wu, Y.-C.; Chung, J.-G.; Yang, J.-S.; Lu, H.-F.; Tsou, M.-F.; Wood, W.G.; Kuo, S.-J.; Chen, D.-R. Quercetin-induced apoptosis acts through mitochondrial- and caspase-3-dependent pathways in human breast cancer MDA-MB-231 cells. Hum. Exp. Toxicol. 2009, 28, 493–503. [Google Scholar] [CrossRef] [PubMed]
- Lin, Y.; Yngve, A.; Lagergren, J.; Lu, Y. A dietary pattern rich in lignans, quercetin and resveratrol decreases the risk of oesophageal cancer. Br. J. Nutr. 2014, 112, 2002–2009. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Li, C.; Zhang, W.-J.; Frei, B. Quercetin inhibits LPS-induced adhesion molecule expression and oxidant production in human aortic endothelial cells by p38-mediated Nrf2 activation and antioxidant enzyme induction. Redox Biol. 2016, 9, 104–113. [Google Scholar] [CrossRef] [Green Version]
- Li, Y.; Yao, J.; Han, C.; Yang, J.; Chaudhry, M.T.; Wang, S.; Liu, H.; Yin, Y. Quercetin, Inflammation and Immunity. Nutrients 2016, 8, 167. [Google Scholar] [CrossRef]
- Priego, S.; Feddi, F.; Ferrer, P.; Mena, S.; Benlloch, M.; Ortega, A.; Carretero, J.; Obrador, E.; Asensi, M.; Estrela, J.M. Natural polyphenols facilitate elimination of HT-29 colorectal cancer xenografts by chemoradiotherapy: A Bcl-2- and superoxide dismutase 2-dependent mechanism. Mol. Cancer Ther. 2008, 7, 3330–3342. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Manna, S.K.; Mukhopadhyay, A.; Van, N.T.; Aggarwal, B.B. Silymarin Suppresses TNF-Induced Activation of NF-κB, c-Jun N-Terminal Kinase, and Apoptosis. J. Immunol. 1999, 163, 6800–6809. [Google Scholar]
- Hoesel, B.; A Schmid, J. The complexity of NF-κB signaling in inflammation and cancer. Mol. Cancer 2013, 12, 86. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Moore-Carrasco, R.; Busquets, S.; Figueras, M.; Palanki, M.; López-Soriano, F.J.; Argilés, J.M. Both AP-1 and NF-κB Seem to be Involved in Tumour Growth in an Experimental Rat Hepatoma. Anticancer Res. 2009, 29, 1315–1317. [Google Scholar]
- Park, S. Polyphenol Compound as a Transcription Factor Inhibitor. Nutrients 2015, 7, 8987–9004. [Google Scholar] [CrossRef] [Green Version]
- Youn, H.; Jeong, J.-C.; Jeong, Y.S.; Kim, E.-J.; Um, S.-J. Quercetin potentiates apoptosis by inhibiting nuclear factor-κB signaling in H460 lung cancer cells. Biol. Pharm. Bull. 2013, 36, 944–951. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Boots, A.W.; Wilms, L.C.; Swennen, E.L.; Kleinjans, J.C.; Bast, A.; Haenen, G.R. In vitro and ex vivo anti-inflammatory activity of quercetin in healthy volunteers. Nutrition 2008, 24, 703–710. [Google Scholar] [CrossRef] [PubMed]
- García-Mediavilla, M.V.; Crespo, I.; Collado, P.S.; Esteller, A.; Sánchez-Campos, S.; Tuñón, M.J.; González-Gallego, J. The anti-inflammatory flavones quercetin and kaempferol cause inhibition of inducible nitric oxide synthase, cyclooxygenase-2 and reactive C-protein, and down-regulation of the nuclear factor κB pathway in Chang Liver cells. Eur. J. Pharmacol. 2007, 557, 221–229. [Google Scholar] [CrossRef] [PubMed]
- McCann, S.E.; Ambrosone, C.B.; Moysich, K.B.; Brasure, J.; Marshall, J.R.; Freudenheim, J.L.; Wilkinson, G.S.; Graham, S. Intakes of Selected Nutrients, Foods, and Phytochemicals and Prostate Cancer Risk in Western New York. Nutr. Cancer 2005, 53, 33–41. [Google Scholar] [CrossRef] [PubMed]
- Nieman, D.C.; Williams, A.S.; Shanely, R.A.; Jin, F.; Mcanulty, S.R.; Triplett, N.T.; Austin, M.D.; Henson, D.A. Quercetin’s Influence on Exercise Performance and Muscle Mitochondrial Biogenesis. Med. Sci. Sports Exerc. 2010, 42, 338–345. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Guardia, T.; Rotelli, A.E.; Juarez, A.O.; Pelzer, L.E. Anti-inflammatory properties of plant flavonoids. Effects of rutin, quercetin and hesperidin on adjuvant arthritis in rat. Il Farm. 2001, 56, 683–687. [Google Scholar] [CrossRef]
- Mamani-Matsuda, M.; Kauss, T.; Al-Kharrat, A.; Rambert, J.; Fawaz, F.; Thiolat, D.; Moynet, D.; Coves, S.; Malvy, D.; Mossalayi, M.D. Therapeutic and preventive properties of quercetin in experimental arthritis correlate with decreased macrophage inflammatory mediators. Biochem. Pharmacol. 2006, 72, 1304–1310. [Google Scholar] [CrossRef] [PubMed]
- Rhee, S.G. H2O2, a Necessary Evil for Cell Signaling. Science 2006, 312, 1882–1883. [Google Scholar] [CrossRef]
- Lambeth, J.D. NOX enzymes and the biology of reactive oxygen. Nat. Rev. Immunol. 2004, 4, 181–189. [Google Scholar] [CrossRef]
- Xu, D.; Hu, M.-J.; Wang, Y.-Q.; Cui, Y.-L. Antioxidant Activities of Quercetin and Its Complexes for Medicinal Application. Molecules 2019, 24, 1123. [Google Scholar] [CrossRef] [Green Version]
- Ward, A.B.; Mir, H.; Kapur, N.; Gales, D.N.; Carriere, P.P.; Singh, S. Quercetin inhibits prostate cancer by attenuating cell survival and inhibiting anti-apoptotic pathways. World J. Surg. Oncol. 2018, 16, 1–12. [Google Scholar] [CrossRef] [Green Version]
- Abarikwu, S.O.; Pant, A.B.; Farombi, E.O. Dietary Antioxidant, Quercetin, Protects Sertoli-Germ Cell Coculture from Atrazine-Induced Oxidative Damage. J. Biochem. Mol. Toxicol. 2012, 26, 477–485. [Google Scholar] [CrossRef]
- Xiao, L.; Luo, G.; Tang, Y.; Yao, P. Quercetin and iron metabolism: What we know and what we need to know. Food Chem. Toxicol. 2018, 114, 190–203. [Google Scholar] [CrossRef] [PubMed]
- Gibellini, L.; Pinti, M.; Nasi, M.; De Biasi, S.; Roat, E.; Bertoncelli, L.; Cossarizza, A. Interfering with ROS Metabolism in Cancer Cells: The Potential Role of Quercetin. Cancers 2010, 2, 1288–1311. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Folkman, J. Angiogenesis: An organizing principle for drug discovery? Nat. Rev. Drug Discov. 2007, 6, 273–286. [Google Scholar] [CrossRef]
- Pratheeshkumar, P.; Budhraja, A.; Son, Y.-O.; Wang, X.; Zhang, Z.; Ding, S.; Wang, L.; Hitron, A.; Lee, J.-C.; Xu, M.; et al. Quercetin Inhibits Angiogenesis Mediated Human Prostate Tumor Growth by Targeting VEGFR- 2 Regulated AKT/mTOR/P70S6K Signaling Pathways. PLoS ONE 2012, 7, e47516. [Google Scholar] [CrossRef] [Green Version]
- Gao, X.; Wang, B.; Wei, X.; Men, K.; Zheng, F.; Zhou, Y.; Zheng, Y.; Gou, M.; Huang, M.; Guo, G.; et al. Anticancer effect and mechanism of polymer micelle-encapsulated quercetin on ovarian cancer. Nanoscale 2012, 4, 7021–7030. [Google Scholar] [CrossRef] [PubMed]
- Wong, M.L.; Prawira, A.; Kaye, A.H.; Hovens, C.M. Tumour angiogenesis: Its mechanism and therapeutic implications in malignant gliomas. J. Clin. Neurosci. 2009, 16, 1119–1130. [Google Scholar] [CrossRef]
- Adams, R.H.; Alitalo, K. Molecular regulation of angiogenesis and lymphangiogenesis. Nat. Rev. Mol. Cell Biol. 2007, 8, 464–478. [Google Scholar] [CrossRef]
- Lupo, G.; Cambria, M.T.; Olivieri, M.; Rocco, C.; Caporarello, N.; Longo, A.; Zanghì, G.; Salmeri, M.; Foti, M.C.; Anfuso, C.D. Anti-angiogenic effect of quercetin and its 8-methyl pentamethyl ether derivative in human microvascular endothelial cells. J. Cell. Mol. Med. 2019, 23, 6565–6577. [Google Scholar] [CrossRef] [Green Version]
- Balakrishnan, S.; Bhat, F.A.; Singh, P.R.; Mukherjee, S.; Elumalai, P.; Das, S.; Patra, C.R.; Arunakaran, J. Gold nanoparticle-conjugated quercetin inhibits epithelial-mesenchymal transition, angiogenesis and invasiveness via EGFR/VEGFR-2-mediated pathway in breast cancer. Cell Prolif. 2016, 49, 678–697. [Google Scholar] [CrossRef]
- Wang, L.; Wu, H.; Xiong, L.; Liu, X.; Yang, N.; Luo, L.; Qin, T.; Zhu, X.; Shen, Z.; Jing, H.; et al. Quercetin Downregulates Cyclooxygenase-2 Expression and HIF-1α/VEGF Signaling-Related Angiogenesis in a Mouse Model of Abdominal Aortic Aneurysm. BioMed Res. Int. 2020, 2020, 1–11. [Google Scholar] [CrossRef] [PubMed]
- Long, Q.; Xiel, Y.; Huang, Y.; Wu, Q.; Zhang, H.; Xiong, S.; Liu, Y.; Chen, L.; Wei, Y.; Zhao, X.; et al. Induction of Apoptosis and Inhibition of Angiogenesis by PEGylated Liposomal Quercetin in Both Cisplatin-Sensitive and Cisplatin-Resistant Ovarian Cancers. J. Biomed. Nanotechnol. 2013, 9, 965–975. [Google Scholar] [CrossRef]
- Lei, C.-S.; Hou, Y.-C.; Pai, M.-H.; Lin, M.-T.; Yeh, S.-L. Effects of quercetin combined with anticancer drugs on metastasis-associated factors of gastric cancer cells: In vitro and in vivo studies. J. Nutr. Biochem. 2018, 51, 105–113. [Google Scholar] [CrossRef] [PubMed]
- Fernald, K.; Kurokawa, M. Evading apoptosis in cancer. Trends Cell Biol. 2013, 23, 620–633. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ghobrial, I.M.; Witzig, T.E.; A Adjei, A. Targeting Apoptosis Pathways in Cancer Therapy. CA Cancer J. Clin. 2005, 55, 178–194. [Google Scholar] [CrossRef] [PubMed]
- Primikyri, A.; Sayyad, N.; Quilici, G.; Vrettos, E.I.; Lim, K.; Chi, S.; Musco, G.; Gerothanassis, I.P.; Tzakos, A.G. Probing the interaction of a quercetin bioconjugate with Bcl-2 in living human cancer cells with in-cell NMR spectroscopy. FEBS Lett. 2018, 592, 3367–3379. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Shen, X.; Si, Y.; Wang, Z.; Wang, J.; Guo, Y.; Zhang, X. Quercetin inhibits the growth of human gastric cancer stem cells by inducing mitochondrial-dependent apoptosis through the inhibition of PI3K/Akt signaling. Int. J. Mol. Med. 2016, 38, 619–626. [Google Scholar] [CrossRef] [PubMed]
- Psahoulia, F.H.; Drosopoulos, K.G.; Doubravska, L.; Andera, L.; Pintzas, A. Quercetin enhances TRAIL-mediated apoptosis in colon cancer cells by inducing the accumulation of death receptors in lipid rafts. Mol. Cancer Ther. 2007, 6, 2591–2599. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chen, W.; Wang, X.; Zhuang, J.; Zhang, L.; Lin, Y. Induction of death receptor 5 and suppression of survivin contribute to sensitization of TRAIL-induced cytotoxicity by quercetin in non-small cell lung cancer cells. Carcinogenesis 2007, 28, 2114–2121. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Meeran, S.M.; Katiyar, S.K. Cell cycle control as a basis for cancer chemoprevention through dietary agents. Front. Biosci. 2008, 13, 2191–2202. [Google Scholar] [CrossRef] [Green Version]
- Gescher, A.; Pastorino, U.; Plummer, S.M.; Manson, M.M. Suppression of tumour development by substances derived from the diet—Mechanisms and clinical implications. Br. J. Clin. Pharmacol. 1998, 45, 1–12. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Milner, J.A.; McDonald, S.S.; Anderson, D.E.; Greenwald, P.R. Reviews: Molecular Targets for Nutrients Involved With Cancer Prevention. Nutr. Cancer 2001, 41, 1–16. [Google Scholar] [CrossRef]
- Yuan, Z.; Long, C.; Junming, T.; Qihuan, L.; Youshun, Z.; Chan, Z. Quercetin-induced apoptosis of HL-60 cells by reducing PI3K/Akt. Mol. Biol. Rep. 2012, 39, 7785–7793. [Google Scholar] [CrossRef]
- Yoshida, M.; Yamamoto, M.; Nikaido, T. Quercetin arrests human leukemic T-cells in late G1 phase of the cell cycle. Cancer Res. 1992, 52, 6676–6681. [Google Scholar]
- Richter, M.; Ebermann, R.; Marian, B. Quercetin-Induced Apoptosis in Colorectal Tumor Cells: Possible Role of EGF Receptor Signaling. Nutr. Cancer 1999, 34, 88–99. [Google Scholar] [CrossRef]
- Srivastava, S.; Somasagara, R.R.; Hegde, M.; Nishana, M.; Tadi, S.K.; Srivastava, M.; Choudhary, B.; Raghavan, S.C. Quercetin, a Natural Flavonoid Interacts with DNA, Arrests Cell Cycle and Causes Tumor Regression by Activating Mitochondrial Pathway of Apoptosis. Sci. Rep. 2016, 6, 24049. [Google Scholar] [CrossRef] [Green Version]
- Choi, J.-A.; Kim, J.-Y.; Lee, J.-Y.; Kang, C.-M.; Kwon, H.-J.; Yoo, Y.-D.; Kim, T.-W.; Lee, Y.-S.; Lee, S.-J. Induction of cell cycle arrest and apoptosis in human breast cancer cells by quercetin. Int. J. Oncol. 2001, 19, 837–844. [Google Scholar] [CrossRef]
- Lee, T.-J.; Kim, O.H.; Kim, Y.H.; Lim, J.H.; Kim, S.; Park, J.-W.; Kwon, T.K. Quercetin arrests G2/M phase and induces caspase-dependent cell death in U937 cells. Cancer Lett. 2006, 240, 234–242. [Google Scholar] [CrossRef]
- Zhang, Q.; Zhao, X.-H.; Wang, Z.-J. Flavones and flavonols exert cytotoxic effects on a human oesophageal adenocarcinoma cell line (OE33) by causing G2/M arrest and inducing apoptosis. Food Chem. Toxicol. 2008, 46, 2042–2053. [Google Scholar] [CrossRef]
- He, D.; Guo, X.; Zhang, E.; Zi, F.; Chen, J.; Chen, Q.; Lin, X.; Yang, L.; Li, Y.; Wu, W.; et al. Quercetin induces cell apoptosis of myeloma and displays a synergistic effect with dexamethasone in vitro and in vivo xenograft models. Oncotarget 2016, 7, 45489–45499. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ma, Y.; Yao, C.; Liu, H.; Yu, F.; Lin, J.; Lu, K.; Liao, C.; Chueh, F.; Chung, J. Quercetin induced apoptosis of human oral cancer SAS cells through mitochondria and endoplasmic reticulum mediated signaling pathways. Oncol. Lett. 2018, 15, 9663–9672. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chang, J.-H.; Lai, S.-L.; Chen, W.-S.; Hung, W.-Y.; Chow, J.-M.; Hsiao, M.; Lee, W.-J.; Chien, M.-H. Quercetin suppresses the metastatic ability of lung cancer through inhibiting Snail-dependent Akt activation and Snail-independent ADAM9 expression pathways. Biochim. Biophys. Acta Bioenerg. 2017, 1864, 1746–1758. [Google Scholar] [CrossRef] [PubMed]
- Nguyen, L.T.; Lee, Y.-H.; Sharma, A.R.; Park, J.-B.; Jagga, S.; Sharma, G.; Lee, S.-S.; Nam, J.-S. Quercetin induces apoptosis and cell cycle arrest in triple-negative breast cancer cells through modulation of Foxo3a activity. Korean J. Physiol. Pharmacol. 2017, 21, 205–213. [Google Scholar] [CrossRef] [Green Version]
- Jeong, J.-H.; An, J.Y.; Kwon, Y.T.; Rhee, J.G.; Lee, Y.J. Effects of low dose quercetin: Cancer cell-specific inhibition of cell cycle progression. J. Cell. Biochem. 2008, 106, 73–82. [Google Scholar] [CrossRef] [Green Version]
- Kooshyar, M.M. A Randomized Placebo- Controlled Double Blind Clinical Trial of Quercetin in the Prevention and Treatment of Chemotherapy-Induced Oral Mucositis. J. Clin. Diagn. Res. 2017, 11, ZC46–ZC50. [Google Scholar] [CrossRef]
- Asea, G.A.A. Effects of the flavonoid drug Quercetin on the response of human prostate tumours to hyperthermia in vitro and in vivo. Int. J. Hyperth. 2001, 17, 347–356. [Google Scholar] [CrossRef]
- Kok, K.; Geering, B.; Vanhaesebroeck, B. Regulation of phosphoinositide 3-kinase expression in health and disease. Trends Biochem. Sci. 2009, 34, 115–127. [Google Scholar] [CrossRef]
- Vogt, P.K.; Hart, J.R.; Gymnopoulos, M.; Jiang, H.; Kang, S.; Bader, A.G.; Zhao, L.; Denley, A. Phosphatidylinositol 3-Kinase: The Oncoprotein. Curr. Top. Microbiol. Immunol. 2010, 347, 79–104. [Google Scholar] [CrossRef] [Green Version]
- Maurya, A.K.; Vinayak, M. PI-103 and Quercetin Attenuate PI3K-AKT Signaling Pathway in T-Cell Lymphoma Exposed to Hydrogen Peroxide. PLoS ONE 2016, 11, e0160686. [Google Scholar] [CrossRef] [PubMed]
- Michaud-Levesque, J.; Bousquet-Gagnon, N.; Béliveau, R. Quercetin abrogates IL-6/STAT3 signaling and inhibits glioblastoma cell line growth and migration. Exp. Cell Res. 2012, 318, 925–935. [Google Scholar] [CrossRef]
- Alvarez, J.V.; Frank, D.A. Genome-wide analysis of STAT target genes: Elucidating the mechanism of STAT-mediated oncogenesis. Cancer Biol. Ther. 2004, 3, 1045–1050. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wang, Y.; Billouin-Frazier, S.S.; Windham, J.P.; Jones, D. Detection of DDR2 mutations in malignant melanoma: A potentially targetable feature of advanced stage disease. J. Clin. Oncol. 2013, 31, 9100. [Google Scholar] [CrossRef]
- Senggunprai, L.; Kukongviriyapan, V.; Prawan, A.; Kukongviriyapan, U. Quercetin and EGCG Exhibit Chemopreventive Effects in Cholangiocarcinoma Cells via Suppression of JAK/STAT Signaling Pathway. Phytother. Res. 2013, 28, 841–848. [Google Scholar] [CrossRef]
- Xiong, A.; Yang, Z.; Shen, Y.; Zhou, J.; Shen, Q. Transcription Factor STAT3 as a Novel Molecular Target for Cancer Prevention. Cancers 2014, 6, 926–957. [Google Scholar] [CrossRef] [Green Version]
- Chen, Y.-W.; Chou, H.-C.; Lin, S.-T.; Chen, Y.-H.; Chang, Y.-J.; Chen, L.; Chan, H.-L. Cardioprotective Effects of Quercetin in Cardiomyocyte under Ischemia/Reperfusion Injury. Evid. Based Complement. Altern. Med. 2013, 2013, 1–16. [Google Scholar] [CrossRef] [Green Version]
- Labbé, D.; Provençal, M.; Lamy, S.; Boivin, D.; Gingras, D.; Béliveau, R. The Flavonols Quercetin, Kaempferol, and Myricetin Inhibit Hepatocyte Growth Factor-Induced Medulloblastoma Cell Migration. J. Nutr. 2009, 139, 646–652. [Google Scholar] [CrossRef] [Green Version]
- Lee, W.-J.; Wu, L.-F.; Chen, W.-K.; Wang, C.-J.; Tseng, T.-H. Inhibitory effect of luteolin on hepatocyte growth factor/scatter factor-induced HepG2 cell invasion involving both MAPK/ERKs and PI3K–Akt pathways. Chem. Biol. Interact. 2006, 160, 123–133. [Google Scholar] [CrossRef]
- Mukherjee, A.; Khuda-Bukhsh, A.R. Quercetin Down-regulates IL-6/STAT-3 Signals to Induce Mitochondrial-mediated Apoptosis in a Nonsmall-cell Lung-cancer Cell Line, A549. J. Pharmacopunct. 2015, 18, 19–26. [Google Scholar] [CrossRef] [PubMed]
- Rauf, A.; Imran, M.; Khan, I.A.; Ur-Rehman, M.-; Gilani, S.A.; Mehmood, Z.; Mubarak, M.S. Anticancer potential of quercetin: A comprehensive review. Phytother. Res. 2018, 32, 2109–2130. [Google Scholar] [CrossRef] [PubMed]
- Seo, H.-S.; Ku, J.M.; Choi, H.-S.; Choi, Y.K.; Woo, J.-K.; Kim, M.; Kim, I.; Na, C.H.; Hur, H.; Jang, B.-H.; et al. Quercetin induces caspase-dependent extrinsic apoptosis through inhibition of signal transducer and activator of transcription 3 signaling in HER2-overexpressing BT-474 breast cancer cells. Oncol. Rep. 2016, 36, 31–42. [Google Scholar] [CrossRef] [Green Version]
- Lamson, D.W.; Brignall, M.S. Antioxidants and cancer, part 3: Quercetin. Altern. Med. Rev. 2000, 5, 196–208. [Google Scholar]
- Nicholson, K.M.; Anderson, N.G. The protein kinase B/Akt signalling pathway in human malignancy. Cell. Signal. 2002, 14, 381–395. [Google Scholar] [CrossRef]
- Mendez, M.G.; Kojima, S.; Goldman, R.D. Vimentin induces changes in cell shape, motility, and adhesion during the epithelial to mesenchymal transition. FASEB J. 2010, 24, 1838–1851. [Google Scholar] [CrossRef] [Green Version]
- Naryzhny, S.N. Proliferating cell nuclear antigen: A proteomics view. Cell. Mol. Life Sci. 2008, 65, 3789–3808. [Google Scholar] [CrossRef] [PubMed]
- Senthilkumar, K.; Arunkumar, R.; Elumalai, P.; Sharmila, G.; Gunadharini, D.N.; Banudevi, S.; Krishnamoorthy, G.; Benson, C.S.; Arunakaran, J. Quercetin inhibits invasion, migration and signalling molecules involved in cell survival and proliferation of prostate cancer cell line (PC-3). Cell Biochem. Funct. 2011, 29, 87–95. [Google Scholar] [CrossRef] [PubMed]
- Sharmila, G.; Bhat, F.; Arunkumar, R.; Elumalai, P.; Singh, P.R.; Senthilkumar, K.; Arunakaran, J. Chemopreventive effect of quercetin, a natural dietary flavonoid on prostate cancer in in vivo model. Clin. Nutr. 2014, 33, 718–726. [Google Scholar] [CrossRef]
- Arunakaran, J. Quercetin, a Natural Dietary Flavonoid Inhibits, Reverses, Retards Tumorigenesis in Prostate and Breast Cancer. Ann. Pharmacol. Pharm. 2017, 2, 1085. [Google Scholar]
- Sharmila, G.; Athirai, T.; Kiruthiga, B.; Senthilkumar, K.; Elumalai, P.; Arunkumar, R.; Arunakaran, J. Chemopreventive Effect of Quercetin in MNU and Testosterone Induced Prostate Cancer of Sprague-Dawley Rats. Nutr. Cancer 2013, 66, 38–46. [Google Scholar] [CrossRef] [PubMed]
- Stuart-Harris, R.; Caldas, C.; Pinder, S.; Pharoah, P. Proliferation markers and survival in early breast cancer: A systematic review and meta-analysis of 85 studies in 32,825 patients. Breast 2008, 17, 323–334. [Google Scholar] [CrossRef] [PubMed]
- Bhat, F.A.; Sharmila, G.; Balakrishnan, S.; Singh, P.R.; Srinivasan, N.; Arunakaran, J. Epidermal growth factor-induced prostate cancer (PC3) cell survival and proliferation is inhibited by quercetin, a plant flavonoid through apoptotic machinery. Biomed. Prev. Nutr. 2014, 4, 459–468. [Google Scholar] [CrossRef]
- Weng, L.-P.; Smith, W.M.; Brown, J.L.; Eng, C. PTEN inhibits insulin-stimulated MEK/MAPK activation and cell growth by blocking IRS-1 phosphorylation and IRS-1/Grb-2/Sos complex formation in a breast cancer model. Hum. Mol. Genet. 2001, 10, 605–616. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cardone, M.H.; Roy, N.; Stennicke, H.R.; Salvesen, G.S.; Franke, T.F.; Stanbridge, E.; Frisch, S.; Reed, J.C. Regulation of Cell Death Protease Caspase-9 by Phosphorylation. Science 1998, 282, 1318–1321. [Google Scholar] [CrossRef] [PubMed]
- Heiden, M.G.V.; Thompson, C.B. Bcl-2 proteins: Regulators of apoptosis or of mitochondrial homeostasis? Nat. Cell Biol. 1999, 1, E209–E216. [Google Scholar] [CrossRef]
- Ali, A.; Kim, M.Y.; Lee, H.J.; Roh, G.S.; Kim, H.J.; Cho, G.J.; Choi, W.S. Quercetin induces cell death in cervical cancer by reducing O-GlcNAcylation of adenosine monophosphate-activated protein kinase. Anat. Cell Biol. 2018, 51, 274–283. [Google Scholar] [CrossRef]
- Sundaram, M.K.; Raina, R.; Afroze, N.; Bajbouj, K.; Hamad, M.; Haque, S.; Hussain, A. Quercetin modulates signaling pathways and induces apoptosis in cervical cancer cells. Biosci. Rep. 2019, 39. [Google Scholar] [CrossRef] [Green Version]
- Liu, Y.; Gong, W.; Yang, Z.Y.; Zhou, X.S.; Gong, C.; Zhang, T.R.; Wei, X.; Ma, D.; Ye, F.; Gao, Q.L. Quercetin induces protective autophagy and apoptosis through ER stress via the p-STAT3/Bcl-2 axis in ovarian cancer. Apoptosis 2017, 22, 544–557. [Google Scholar] [CrossRef]
- Gong, C.; Yang, Z.; Zhang, L.; Wang, Y.; Gong, W.; Liu, Y. Quercetin suppresses DNA double-strand break repair and enhances the radiosensitivity of human ovarian cancer cells via p53-dependent endoplasmic reticulum stress pathway. Onco Targets Ther. 2017, 11, 17–27. [Google Scholar] [CrossRef] [Green Version]
- Liu, L.; Zhang, J.; Yang, X.; Fang, C.; Xu, H.; Xi, X. SALL4 as an Epithelial-Mesenchymal Transition and Drug Resistance Inducer through the Regulation of c-Myc in Endometrial Cancer. PLoS ONE 2015, 10, e0138515. [Google Scholar] [CrossRef] [PubMed]
- Herr, I.; Zhou, W.; Kallifatidis, G.; Baumann, B.; Rausch, V.; Mattern, J.; Gladkich, J.; Giese, N.; Moldenhauer, G.; Wirth, T.; et al. Dietary polyphenol quercetin targets pancreatic cancer stem cells. Int. J. Oncol. 2010, 37, 551–561. [Google Scholar] [CrossRef] [Green Version]
- Angst, E.; Park, J.L.; Moro, A.; Lu, Q.-Y.; Lu, X.; Li, G.; King, J.; Chen, M.; Reber, H.A.; Go, V.L.W.; et al. The Flavonoid Quercetin Inhibits Pancreatic Cancer Growth In Vitro and In Vivo. Pancreas 2013, 42, 223–229. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pang, B.; Xu, X.; Lu, Y.; Jin, H.; Yang, R.; Jiang, C.; Shao, D.; Liu, Y.; Shi, J. Prediction of new targets and mechanisms for quercetin in the treatment of pancreatic cancer, colon cancer, and rectal cancer. Food Funct. 2019, 10, 5339–5349. [Google Scholar] [CrossRef]
- Rubinsztein, D.C.; Cuervo, A.M.; Ravikumar, B.; Sarkar, S.; Korolchuk, V.I.; Kaushik, S.; Klionsky, D.J. In search of an “autophagomometer”. Autophagy 2009, 5, 585–589. [Google Scholar] [CrossRef]
- Li, H.; Chen, C. Quercetin Has Antimetastatic Effects on Gastric Cancer Cells via the Interruption of uPA/uPAR Function by Modulating NF-κb, PKC-δ, ERK1/2, and AMPKα. Integr. Cancer Ther. 2017, 17, 511–523. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Srisa-Nga, K.; Mankhetkorn, S.; Okonogi, S.; Khonkarn, R. Delivery of Superparamagnetic Polymeric Micelles Loaded With Quercetin to Hepatocellular Carcinoma Cells. J. Pharm. Sci. 2019, 108, 996–1006. [Google Scholar] [CrossRef]
- Wu, L.; Li, J.; Liu, T.; Li, S.; Feng, J.; Yu, Q.; Zhang, J.; Chen, J.; Zhou, Y.; Ji, J.; et al. Quercetin shows anti-tumor effect in hepatocellular carcinoma LM3 cells by abrogating JAK2/STAT3 signaling pathway. Cancer Med. 2019, 8, 4806–4820. [Google Scholar] [CrossRef]
- Hisaka, T.; Sakai, H.; Sato, T.; Goto, Y.; Nomura, Y.; Fukutomi, S.; Fujita, F.; Mizobe, T.; Nakashima, O.; Tanigawa, M.; et al. Quercetin Suppresses Proliferation of Liver Cancer Cell Lines In Vitro. Anticancer. Res. 2020, 40, 4695–4700. [Google Scholar] [CrossRef]
- Yang, L.; Liu, Y.; Wang, M.; Qian, Y.; Dong, X.; Gu, H.; Wang, H.; Guo, S.; Hisamitsu, T. Quercetin-induced apoptosis of HT-29 colon cancer cells via inhibition of the Akt-CSN6-Myc signaling axis. Mol. Med. Rep. 2016, 14, 4559–4566. [Google Scholar] [CrossRef] [Green Version]
- Buonerba, C.; De Placido, P.; Bruzzese, D.; Pagliuca, M.; Ungaro, P.; Bosso, D.; Ribera, D.; Iaccarino, S.; Scafuri, L.; Liotti, A.; et al. Isoquercetin as an Adjunct Therapy in Patients With Kidney Cancer Receiving First-Line Sunitinib (QUASAR): Results of a Phase I Trial. Front. Pharmacol. 2018, 9. [Google Scholar] [CrossRef] [Green Version]
- Xing, N.; Chen, Y.; Mitchell, S.H.; Young, C.Y.F. Quercetin inhibits the expression and function of the androgen receptor in LNCaP prostate cancer cells. Carcinogenesis 2001, 22, 409–414. [Google Scholar] [CrossRef] [Green Version]
- Su, Q.; Peng, M.; Zhang, Y.; Xu, W.; Darko, K.O.; Tao, T.; Huang, Y.; Tao, X.; Yang, X. Quercetin induces bladder cancer cells apoptosis by activation of AMPK signaling pathway. Am. J. Cancer Res. 2016, 6, 498–508. [Google Scholar]
- Oršolić, N.; Karač, I.; Sirovina, D.; Kukolj, M.; Kunštić, M.; Gajski, G.; Garaj-Vrhovac, V.; Štajcar, D. Chemotherapeutic potential of quercetin on human bladder cancer cells. J. Environ. Sci. Health A 2016, 51, 776–781. [Google Scholar] [CrossRef] [PubMed]
- Russo, G.L.R.; Usso, M.; Spagnuolo, C.; Moccia, S. Ins and Outs of Flavonoids in Cancer Prevention vs. Cancer Therapy: A Lesson from Quercetin in Leukemia. Proceedings 2017, 1, 977. [Google Scholar] [CrossRef] [Green Version]
- Naimi, A.; Entezari, A.; Hagh, M.F.; Hassanzadeh, A.; Saraei, R.; Solali, S. Quercetin sensitizes human myeloid leukemia KG-1 cells against TRAIL-induced apoptosis. J. Cell. Physiol. 2019, 234, 13233–13241. [Google Scholar] [CrossRef]
- Jacquemin, G.; Granci, V.; Gallouet, A.S.; Lalaoui, N.; Morlé, A.; Iessi, E.; Morizot, A.; Garrido, C.; Guillaudeux, T.; Micheau, O. Quercetin-mediated Mcl-1 and survivin downregulation restores TRAIL-induced apoptosis in non-Hodgkin’s lymphoma B cells. Haematology 2011, 97, 38–46. [Google Scholar] [CrossRef] [Green Version]
- Xingyu, Z.; Peijie, M.; Dan, P.; Youg, W.; Daojun, W.; Xinzheng, C.; Xijun, Z.; Yangrong, S. Quercetin suppresses lung cancer growth by targeting Aurora B kinase. Cancer Med. 2016, 5, 3156–3165. [Google Scholar] [CrossRef] [Green Version]
- Lan, H.; Hong, W.; Fan, P.; Qian, D.; Zhu, J.; Bai, B. Quercetin Inhibits Cell Migration and Invasion in Human Osteosarcoma Cells. Cell. Physiol. Biochem. 2017, 43, 553–567. [Google Scholar] [CrossRef] [Green Version]
- Li, S.; Pei, Y.; Wang, W.; Liu, F.; Zheng, K.; Zhang, X. Quercetin suppresses the proliferation and metastasis of metastatic osteosarcoma cells by inhibiting parathyroid hormone receptor 1. Biomed. Pharmacother. 2019, 114, 108839. [Google Scholar] [CrossRef] [PubMed]
- Pozsgai, E.; Bellyei, S.; Cseh, A.; Boronkai, A.; Racz, B.; Szabo, A.; Sumegi, B.; Hocsak, E. Quercetin Increases the Efficacy of Glioblastoma Treatment Compared to Standard Chemoradiotherapy by the Suppression of PI-3-Kinase-Akt Pathway. Nutr. Cancer 2013, 65, 1059–1066. [Google Scholar] [CrossRef] [Green Version]
- Kiekow, C.J.; Figueiró, F.; Dietrich, F.; Vechia, L.D.; Pires, E.N.; Jandrey, E.H.; Gnoatto, S.C.; Salbego, C.G.; Battastini, A.M.O.; Gosmann, G. Quercetin derivative induces cell death in glioma cells by modulating NF-κB nuclear translocation and caspase-3 activation. Eur. J. Pharm. Sci. 2016, 84, 116–122. [Google Scholar] [CrossRef] [PubMed]
- Chan, C.-Y.; Lien, C.-H.; Lee, M.-F.; Huang, C.-Y. Quercetin suppresses cellular migration and invasion in human head and neck squamous cell carcinoma (HNSCC). Biomedicine 2016, 6, 1–6. [Google Scholar] [CrossRef]
- Castillo, M.H.; Perkins, I.; Campbell, J.H.; Doerr, R.; Hassett, J.M.; Kandaswami, C.; Middleton, E. The effects of the bioflavonoid quercetin on squamous cell carcinoma of head and neck origin. Am. J. Surg. 1989, 158, 351–355. [Google Scholar] [CrossRef]
- Lai, W.-W.; Hsu, S.-C.; Chueh, F.-S.; Chen, Y.-Y.; Yang, Y.-S.; Lin, J.-P.; Lien, J.-C.; Tsai, C.-H.; Chung, J.-G. Quercetin inhibits migration and invasion of SAS human oral cancer cells through inhibition of NF-κB and matrix metalloproteinase-2/-9 signaling pathways. Anticancer Res. 2013, 33, 1941–1950. [Google Scholar] [PubMed]
- Min, Y.S.; Lee, S.E.; Hong, S.T.; Kim, H.S.; Choi, B.-C.; Sim, S.S.; Whang, W.K.; Sohn, U.D. The Inhibitory Effect of Quercetin-3-O-β-d-Glucuronopyranoside on Gastritis and Reflux Esophagitis in Rats. Korean J. Physiol. Pharmacol. 2009, 13, 295–300. [Google Scholar] [CrossRef] [Green Version]
- Harris, Z.; Donovan, M.G.; Branco, G.M.; Limesand, K.H.; Burd, R. Quercetin as an Emerging Anti-Melanoma Agent: A Four-Focus Area Therapeutic Development Strategy. Front. Nutr. 2016, 3. [Google Scholar] [CrossRef] [PubMed]
- Marback, E.F.; Arias, V.E.A.; A Paranhos, J.; Soares, F.A.; Murphree, A.L.; Erwenne, C.M. Tumour angiogenesis as a prognostic factor for disease dissemination in retinoblastoma. Br. J. Ophthalmol. 2003, 87, 1224–1228. [Google Scholar] [CrossRef]
- Marth, C.; Landoni, F.; Mahner, S.; McCormack, M.; Gonzalez-Martin, A.; Colombo, N. Cervical cancer: ESMO Clinical Practice Guidelines for diagnosis, treatment and follow-up. Ann. Oncol. 2017, 28, iv72–iv83. [Google Scholar] [CrossRef] [PubMed]
- Durst, M.; Gissmann, L.; Ikenberg, H.; Hausen, H.Z. A papillomavirus DNA from a cervical carcinoma and its prevalence in cancer biopsy samples from different geographic regions. Proc. Natl. Acad. Sci. USA 1983, 80, 3812–3815. [Google Scholar] [CrossRef] [Green Version]
- Luo, C.-L.; Liu, Y.-Q.; Wang, P.; Song, C.-H.; Wang, K.-J.; Dai, L.-P.; Zhang, J.-Y.; Ye, H. The effect of quercetin nanoparticle on cervical cancer progression by inducing apoptosis, autophagy and anti-proliferation via JAK2 suppression. Biomed. Pharmacother. 2016, 82, 595–605. [Google Scholar] [CrossRef]
- Clemente-Soto, A.F.; Salas-Vidal, E.; Milan-Pacheco, C.; Peralta-Zaragoza, O. Quercetin induces G2 phase arrest and apoptosis with the activation of p53 in an E6 expression-independent manner in HPV-positive human cervical cancer-derived cells. Mol. Med. Rep. 2019, 19, 2097–2106. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Fan, J.-J.; Hsu, W.-H.; Lee, K.-H.; Chen, K.-C.; Lin, C.-W.; A Lee, Y.-L.; Ko, T.-P.; Lee, L.-T.; Lee, M.-T.; Chang, M.-S.; et al. Dietary Flavonoids Luteolin and Quercetin Inhibit Migration and Invasion of Squamous Carcinoma through Reduction of Src/Stat3/S100A7 Signaling. Antioxidants 2019, 8, 557. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wang, R.; Yang, L.; Li, S.; Ye, D.; Yang, L.; Liu, Q.; Zhao, Z.; Cai, Q.; Tan, J.; Li, X. Quercetin Inhibits Breast Cancer Stem Cells via Downregulation of Aldehyde Dehydrogenase 1A1 (ALDH1A1), Chemokine Receptor Type 4 (CXCR4), Mucin 1 (MUC1), and Epithelial Cell Adhesion Molecule (EpCAM). Med. Sci. Monit. 2018, 24, 412–420. [Google Scholar] [CrossRef] [Green Version]
- Nikolaou, K.C.; Talianidis, I. Hepatic cancer stem cells may arise from adult ductal progenitors. Mol. Cell. Oncol. 2015, 3, e1021946. [Google Scholar] [CrossRef] [Green Version]
- Noolu, B.; Gogulothu, R.; Bhat, M.; Qadri, S.S.; Reddy, V.S.; Reddy, G.B.; Ismail, A. In Vivo Inhibition of Proteasome Activity and Tumour Growth by Murraya koenigii Leaf Extract in Breast Cancer Xenografts and by Its Active Flavonoids in Breast Cancer Cells. Anti-Cancer Agents Med. Chem. 2016, 16, 1605–1614. [Google Scholar] [CrossRef]
- Pradhan, S.J.; Mishra, R.; Sharma, P.; Kundu, G.C. Quercetin and sulforaphane in combination suppress the progression of melanoma through the down-regulation of matrix metalloproteinase-9. Exp. Ther. Med. 2010, 1, 915–920. [Google Scholar] [CrossRef] [Green Version]
- Ezzati, M.; Yousefi, B.; Velaei, K.; Safa, A. A review on anti-cancer properties of Quercetin in breast cancer. Life Sci. 2020, 248, 117463. [Google Scholar] [CrossRef]
- Maciejczyk, A.; Surowiak, P. Quercetin inhibits proliferation and increases sensitivity of ovarian cancer cells to cisplatin and paclitaxel. Ginekol. Polska 2013, 84, 590–595. [Google Scholar] [CrossRef]
- Jayson, G.C.; Kohn, E.C.; Kitchener, H.C.; A Ledermann, J. Ovarian cancer. Lancet 2014, 384, 1376–1388. [Google Scholar] [CrossRef]
- Rafie, N.; Hamedani, S.G.; Ghiasvand, R.; Miraghajani, M. Kefir and Cancer: A Systematic Review of Literatures. Arch. Iran. Med. 2015, 18, 852–857. [Google Scholar]
- McCann, S.E.; Freudenheim, J.L.; Marshall, J.R.; Graham, S. Risk of Human Ovarian Cancer Is Related to Dietary Intake of Selected Nutrients, Phytochemicals and Food Groups. J. Nutr. 2003, 133, 1937–1942. [Google Scholar] [CrossRef]
- Seidman, J.D.; Kurman, R.J. Pathology of ovarian carcinoma. Hematol. Clin. N. Am. 2003, 17, 909–925. [Google Scholar] [CrossRef]
- Fathalla, M. Incessant Ovulation—A Factor In Ovarian Neoplasia? Lancet 1971, 298, 163. [Google Scholar] [CrossRef]
- Saed, G.M.; Diamond, M.P.; Fletcher, N.M. Updates of the role of oxidative stress in the pathogenesis of ovarian cancer. Gynecol. Oncol. 2017, 145, 595–602. [Google Scholar] [CrossRef]
- Gates, M.A.; Tworoger, S.S.; Hecht, J.L.; De Vivo, I.; Rosner, B.; Hankinson, S.E. A prospective study of dietary flavonoid intake and incidence of epithelial ovarian cancer. Int. J. Cancer 2007, 121, 2225–2232. [Google Scholar] [CrossRef]
- Scambia, G.; Ranelletti, F.; Panici, P.; Piantelli, M.; Bonanno, G.; De Vincenzo, R.; Ferrandina, G.; Maggiano, N.; Capelli, A.; Mancuso, S. Inhibitory effect of quercetin on primary ovarian and endometrial cancers and synergistic activity with cis-diamminedichloroplatinum(II). Gynecol. Oncol. 1992, 45, 13–19. [Google Scholar] [CrossRef]
- Nessa, M.U.; Beale, P.; Chan, C.; Yu, J.Q.; Huq, F. Synergism from combinations of cisplatin and oxaliplatin with Querce-tin and thymoquinone in human ovarian tumour models. Anticancer Res. 2011, 31, 3789–3797. [Google Scholar]
- Siegelin, M.D.; Reuss, D.E.; Habel, A.; Rami, A.; Von Deimling, A. Quercetin promotes degradation of survivin and thereby enhances death-receptor–mediated apoptosis in glioma cells. Neuro-Oncology 2009, 11, 122–131. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gupta, K.; Panda, D. Perturbation of Microtubule Polymerization by Quercetin through Tubulin Binding: A Novel Mechanism of Its Antiproliferative Activity†. Biochemistry 2002, 41, 13029–13038. [Google Scholar] [CrossRef] [PubMed]
- Yang, Z.; Liu, Y.; Liao, J.; Gong, C.; Sun, C.; Zhou, X.; Wei, X.; Zhang, T.; Gao, Q.; Ma, D.; et al. Quercetin induces endoplasmic reticulum stress to enhance cDDP cytotoxicity in ovarian cancer: Involvement of STAT3 signaling. FEBS J. 2015, 282, 1111–1125. [Google Scholar] [CrossRef]
- Luo, H.; Jiang, B.-H.; King, S.M.; Chen, Y.C. Inhibition of Cell Growth and VEGF Expression in Ovarian Cancer Cells by Flavonoids. Nutr. Cancer 2008, 60, 800–809. [Google Scholar] [CrossRef]
- Parazzini, F.; La Vecchia, C.; Bocciolone, L.; Franceschi, S. The epidemiology of endometrial cancer. Gynecol. Oncol. 1991, 41, 1–16. [Google Scholar] [CrossRef]
- Kaaks, R.; Lukanova, A.; Kurzer, M.S. Obesity, endogenous hormones, and endometrial cancer risk: A synthetic review. Cancer Epidemiol. Biomark. Prev. 2002, 11, 1531–1543. [Google Scholar]
- Bandera, E.V.; Williams, M.G.; Sima, C.; Bayuga, S.; Pulick, K.; Wilcox, H.; Soslow, R.; Zauber, A.G.; Olson, S.H. Phytoestrogen consumption and endometrial cancer risk: A population-based case–control study in New Jersey. Cancer Causes Control. 2009, 20, 1117–1127. [Google Scholar] [CrossRef] [Green Version]
- Lin, C.-W.; Lai, G.-M.; Chen, K.-C.; Lin, T.-H.; Fan, J.-J.; Hsu, R.-L.; Chou, C.-M.; Lin, C.-M.; Kandaswami, C.C.; Lee, M.-T.; et al. RPS12 increases the invasiveness in cervical cancer activated by c-Myc and inhibited by the dietary flavonoids luteolin and quercetin. J. Funct. Foods 2015, 19, 236–247. [Google Scholar] [CrossRef]
- Rossi, M.; Edefonti, V.C.; Parpinel, M.; Lagiou, P.; Franchi, M.; Ferraroni, M.; DeCarli, A.; Zucchetto, A.; Serraino, D.; Maso, L.D.; et al. Proanthocyanidins and other flavonoids in relation to endometrial cancer risk: A case–control study in Italy. Br. J. Cancer 2013, 109, 1914–1920. [Google Scholar] [CrossRef] [Green Version]
- Klein, A.P.; Brune, K.A.; Petersen, G.M.; Goggins, M.; Tersmette, A.C.; Offerhaus, G.J.A.; Griffin, C.; Cameron, J.L.; Yeo, C.J.; Kern, S.; et al. Prospective Risk of Pancreatic Cancer in Familial Pancreatic Cancer Kindreds. Cancer Res. 2004, 64, 2634–2638. [Google Scholar] [CrossRef] [Green Version]
- Johnson, J.L.; Rupasinghe, S.G.; Stefani, F.; Schuler, M.A.; De Mejia, E.G. Citrus Flavonoids Luteolin, Apigenin, and Quercetin Inhibit Glycogen Synthase Kinase-3β Enzymatic Activity by Lowering the Interaction Energy Within the Binding Cavity. J. Med. Food 2011, 14, 325–333. [Google Scholar] [CrossRef]
- Jung, J.H.; Lee, J.O.; Kim, J.H.; Lee, S.K.; You, G.Y.; Park, S.H.; Park, J.M.; Kim, E.-K.; Suh, P.-G.; An, J.K.; et al. Quercetin suppresses HeLa cell viability via AMPK-induced HSP70 and EGFR down-regulation. J. Cell. Physiol. 2010, 223, 408–414. [Google Scholar] [CrossRef]
- Sun, Z.-J.; Chen, G.; Hu, X.; Zhang, W.; Liu, Y.; Zhu, L.-X.; Zhou, Q.; Zhao, Y.-F. Activation of PI3K/Akt/IKK-α/NF-κB signaling pathway is required for the apoptosis-evasion in human salivary adenoid cystic carcinoma: Its inhibition by quercetin. Apoptosis 2010, 15, 850–863. [Google Scholar] [CrossRef]
- Priyadarsini, R.V.; Murugan, R.S.; Maitreyi, S.; Ramalingam, K.; Karunagaran, D.; Nagini, S. The flavonoid quercetin induces cell cycle arrest and mitochondria-mediated apoptosis in human cervical cancer (HeLa) cells through p53 induction and NF-κB inhibition. Eur. J. Pharmacol. 2010, 649, 84–91. [Google Scholar] [CrossRef]
- Wang, K.; Liu, R.; Li, J.; Mao, J.; Lei, Y.; Wu, J.; Zeng, J.; Zhang, T.; Wu, H.; Chen, L.; et al. Quercetin induces protective autophagy in gastric cancer cells: Involvement of Akt-mTOR- and hypoxia-induced factor 1α-mediated signaling. Autophagy 2011, 7, 966–978. [Google Scholar] [CrossRef] [Green Version]
- Mouria, M.; Gukovskaya, A.S.; Jung, Y.; Buechler, P.; Hines, O.J.; Reber, H.A.; Pandol, S.J. Food-derived polyphenols inhibit pancreatic cancer growth through mitochondrial cytochrome C release and apoptosis. Int. J. Cancer 2002, 98, 761–769. [Google Scholar] [CrossRef] [PubMed]
- Fan, P.; Zhang, Y.; Liu, L.; Zhao, Z.; Yin, Y.; Xiao, X.; Bauer, N.; Gladkich, J.; Mattern, J.; Gao, C.; et al. Continuous exposure of pancreatic cancer cells to dietary bioactive agents does not induce drug resistance unlike chemotherapy. Cell Death Dis. 2016, 7, e2246. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kallifatidis, G.; Rausch, V.; Baumann, B.; Apel, A.; Beckermann, B.M.; Groth, A.; Mattern, J.; Li, Z.; Kolb, A.; Moldenhauer, G.; et al. Sulforaphane targets pancreatic tumour-initiating cells by NF-B-induced antiapoptotic signalling. Gut 2008, 58, 949–963. [Google Scholar] [CrossRef] [PubMed]
- Liu, R.; Li, Z.; Bai, S.; Zhang, H.; Tang, M.; Lei, Y.; Chen, L.; Liang, S.; Zhao, Y.-L.; Wei, Y.; et al. Mechanism of Cancer Cell Adaptation to Metabolic Stress. Mol. Cell. Proteom. 2009, 8, 70–85. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kim, D.-O.; Lee, C.Y. Comprehensive Study on Vitamin C Equivalent Antioxidant Capacity (VCEAC) of Various Polyphenolics in Scavenging a Free Radical and its Structural Relationship. Crit. Rev. Food Sci. Nutr. 2004, 44, 253–273. [Google Scholar] [CrossRef]
- Yang, C.S.; Landau, J.M.; Huang, M.-T.; Newmark, H.L. Inhibition of Carcinogenesis by Dietary Polyphenolic Compounds. Annu. Rev. Nutr. 2001, 21, 381–406. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ekström, A.M.; Serafini, M.; Nyrén, O.; Wolk, A.; Bosetti, C.; Bellocco, R. Dietary quercetin intake and risk of gastric cancer: Results from a population-based study in Sweden. Ann. Oncol. 2010, 22, 438–443. [Google Scholar] [CrossRef]
- Lee, K.H.; Kim, S.W.; Kim, J.-R. Reactive oxygen species regulate urokinase plasminogen activator expression and cell invasion via mitogen-activated protein kinase pathways after treatment with hepatocyte growth factor in stomach cancer cells. J. Exp. Clin. Cancer Res. 2009, 28, 73–79. [Google Scholar] [CrossRef] [Green Version]
- Shintani, T. Autophagy in Health and Disease: A Double-Edged Sword. Science 2004, 306, 990–995. [Google Scholar] [CrossRef] [Green Version]
- Wang, P.; Zhang, K.; Zhang, Q.; Mei, J.; Chen, C.J.; Feng, Z.Z.; Yu, D.H. Effects of quercetin on the apoptosis of the human gastric carcinoma cells. Toxicol. In Vitro 2012, 26, 221–228. [Google Scholar] [CrossRef]
- Kim, D.H.; Khan, H.; Ullah, H.; Hassan, S.T.; Šmejkal, K.; Efferth, T.; Mahomoodally, M.F.; Xu, S.; Habtemariam, S.; Filosa, R.; et al. MicroRNA targeting by quercetin in cancer treatment and chemoprotection. Pharmacol. Res. 2019, 147, 104346. [Google Scholar] [CrossRef] [PubMed]
- Zhang, J.-Y.; Lin, M.-T.; Zhou, M.-J.; Yi, T.; Tang, Y.-N.; Tang, S.-L.; Yang, Z.-J.; Zhao, Z.-Z.; Chen, H.-B. Combinational Treatment of Curcumin and Quercetin against Gastric Cancer MGC-803 Cells in Vitro. Molecules 2015, 20, 11524–11534. [Google Scholar] [CrossRef] [Green Version]
- Yang, L.; Hu, Z.; Zhu, J.; Liang, Q.; Zhou, H.; Li, J.; Fan, X.; Zhao, Z.; Pan, H.; Fei, B. Systematic Elucidation of the Mechanism of Quercetin against Gastric Cancer via Network Pharmacology Approach. BioMed Res. Int. 2020, 2020, 1–11. [Google Scholar] [CrossRef]
- Casella, M.L.; Parody, J.P.; Ceballos, M.P.; Quiroga, A.D.; Ronco, M.T.; Francés, D.E.; Monti, J.A.; Pisani, G.B.; Carnovale, C.E.; Carrillo, M.C.; et al. Quercetin prevents liver carcinogenesis by inducing cell cycle arrest, decreasing cell proliferation and enhancing apoptosis. Mol. Nutr. Food Res. 2013, 58, 289–300. [Google Scholar] [CrossRef] [Green Version]
- Dai, W.; Gao, Q.; Qiu, J.; Yuan, J.; Wu, G.; Shen, G. Quercetin induces apoptosis and enhances 5-FU therapeutic efficacy in hepatocellular carcinoma. Tumor Biol. 2015, 37, 6307–6313. [Google Scholar] [CrossRef] [PubMed]
- Wu, L.; Wang, C.; Li, J.; Li, S.; Feng, J.; Liu, T.; Xu, S.; Wang, W.; Lu, X.; Chen, K.; et al. Hepatoprotective effect of quercetin via TRAF6/JNK pathway in acute hepatitis. Biomed. Pharmacother. 2017, 96, 1137–1146. [Google Scholar] [CrossRef]
- Wu, L.; Zhang, Q.; Dai, W.; Li, S.; Feng, J.; Li, J.; Liu, T.; Xu, S.; Wang, W.; Lu, X.; et al. Quercetin Pretreatment Attenuates Hepatic Ischemia Reperfusion-Induced Apoptosis and Autophagy by Inhibiting ERK/NF-κB Pathway. Gastroenterol. Res. Pract. 2017, 2017, 1–15. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wu, L.; Zhang, Q.; Mo, W.; Feng, J.; Li, S.; Li, J.; Liu, T.; Xu, S.; Wang, W.; Lu, X.; et al. Quercetin prevents hepatic fibrosis by inhibiting hepatic stellate cell activation and reducing autophagy via the TGF-β1/Smads and PI3K/Akt pathways. Sci. Rep. 2017, 7, 1–13. [Google Scholar] [CrossRef] [Green Version]
- Granato, M.; Rizzello, C.; Montani, M.S.G.; Cuomo, L.; Vitillo, M.; Santarelli, R.; Gonnella, R.; D’Orazi, G.; Faggioni, A.; Cirone, M. Quercetin induces apoptosis and autophagy in primary effusion lymphoma cells by inhibiting PI3K/AKT/mTOR and STAT3 signaling pathways. J. Nutr. Biochem. 2017, 41, 124–136. [Google Scholar] [CrossRef]
- Bhardwaj, A.; Sethi, G.; Vadhan-Raj, S.; Bueso-Ramos, C.; Takada, Y.; Gaur, U.; Nair, A.S.; Shishodia, S.; Aggarwal, B.B. Resveratrol inhibits proliferation, induces apoptosis, and overcomes chemoresistance through down-regulation of STAT3 and nuclear factor-κB–regulated antiapoptotic and cell survival gene products in human multiple myeloma cells. Blood 2006, 109, 2293–2302. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Liu, Y.; Li, P.-K.; Li, C.; Lin, J. Inhibition of STAT3 Signaling Blocks the Anti-apoptotic Activity of IL-6 in Human Liver Cancer Cells. J. Biol. Chem. 2010, 285, 27429–27439. [Google Scholar] [CrossRef] [Green Version]
- Brito, A.F.; Ribeiro, M.; Abrantes, A.M.; Mamede, A.C.; Laranjo, M.; Casalta-Lopes, J.E.; Gonçalves, A.C.; Sarmento-Ribeiro, A.B.; Tralhão, J.G.; Botelho, M.F. New Approach for Treatment of Primary Liver Tumors: The Role of Quercetin. Nutr. Cancer 2016, 68, 250–266. [Google Scholar] [CrossRef] [PubMed]
- Carrasco-Torres, G.; Monroy-Ramírez, H.C.; Martínez-Guerra, A.A.; Baltiérrez-Hoyos, R.; Romero-Tlalolini, M.D.L. Ángeles; Villa-Treviño, S.; Sánchez-Chino, X.; Vásquez-Garzón, V.R. Quercetin Reverses Rat Liver Preneoplastic Lesions Induced by Chemical Carcinogenesis. Oxid. Med. Cell. Longev. 2017, 2017, 1–8. [Google Scholar] [CrossRef]
- Boateng, J.; Verghese, M.; Shackelford, L.; Walker, L.T.; Khatiwada, J.; Ogutu, S.; Williams, D.S.; Jones, J.; Guyton, M.; Asiamah, D.; et al. Selected fruits reduce azoxymethane (AOM)-induced aberrant crypt foci (ACF) in Fisher 344 male rats. Food Chem. Toxicol. 2007, 45, 725–732. [Google Scholar] [CrossRef]
- Zhang, X.-A.; Zhang, S.; Yin, Q.; Zhang, J. Quercetin induces human colon cancer cells apoptosis by inhibiting the nuclear factor-kappa B Pathway. Pharmacogn. Mag. 2015, 11, 404–409. [Google Scholar] [CrossRef] [Green Version]
- Janzen, N.K.; Perry, K.T.; Han, K.-R.; Kristo, B.; Raman, S.; Said, J.W.; Belldegrun, A.S.; Schulam, P.G. The Effects of Intentional Cryoablation and Radio Frequency Ablation Of Renal Tissue Involving The Collecting System In A Porcine Model. J. Urol. 2005, 173, 1368–1374. [Google Scholar] [CrossRef]
- Jemal, A.; Tiwari, R.C.; Murray, T.; Ghafoor, A.; Samuels, A.; Ward, E.; Feuer, E.J.; Thun, M.J. Cancer Statistics, 2004. CA Cancer J. Clin. 2004, 54, 8–29. [Google Scholar] [CrossRef] [PubMed]
- Meng, F.-D.; Li, Y.; Tian, X.; Ma, P.; Sui, C.-G.; Fu, L.-Y.; Jiang, Y.-H. Synergistic effects of snail and Quercetin on renal cell carcinoma Caki-2 by altering AKT/mTOR/ERK1/2 signaling pathways. Int. J. Clin. Exp. Pathol. 2015, 8, 6157–6168. [Google Scholar]
- Li, W.; Liu, M.; Xu, Y.-F.; Feng, Y.; Che, J.-P.; Wang, G.-C.; Zheng, J.-H. Combination of quercetin and hyperoside has anticancer effects on renal cancer cells through inhibition of oncogenic microRNA-27a. Oncol. Rep. 2013, 31, 117–124. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wei, R.; Zhu, G.; Jia, N.; Yang, W. Epigallocatechin-3-gallate Sensitizes Human 786-O Renal Cell Carcinoma Cells to TRAIL-Induced Apoptosis. Cell Biophys. 2014, 72, 157–164. [Google Scholar] [CrossRef]
- Wang, P.; Heber, D.; Henning, S.M. Quercetin increased bioavailability and decreased methylation of green tea polyphenols in vitro and in vivo. Food Funct. 2012, 3, 635–642. [Google Scholar] [CrossRef] [Green Version]
- Ren, J.; Li, J.; Liu, X.; Feng, Y.; Gui, Y.; Yang, J.; He, W.; Dai, C. Quercetin Inhibits Fibroblast Activation and Kidney Fibrosis Involving the Suppression of Mammalian Target of Rapamycin and β-catenin Signaling. Sci. Rep. 2016, 6, 23968. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wang, G.; Song, L.; Wang, H.; Xing, N. Quercetin synergizes with 2-methoxyestradiol inhibiting cell growth and inducing apoptosis in human prostate cancer cells. Oncol. Rep. 2013, 30, 357–363. [Google Scholar] [CrossRef]
- Firdous, A.B.; Sharmila, G.; Balakrishnan, S.; Rajasingh, P.; Suganya, S.; Srinivasan, N.; Arunakaran, J. Quercetin, a natural dietary flavonoid, acts as a chemopreventive agent against prostate cancer in an in vivo model by inhibiting the EGFR signaling pathway. Food Funct. 2014, 5, 2632–2645. [Google Scholar] [CrossRef]
- Piao, S.; Kang, M.; Lee, Y.J.; Choi, W.S.; Chun, Y.-S.; Kwak, C.; Kim, H.H. Cytotoxic Effects of Escin on Human Castration-resistant Prostate Cancer Cells Through the Induction of Apoptosis and G2/M Cell Cycle Arrest. Urology 2014, 84, 982.e1–982.e7. [Google Scholar] [CrossRef] [Green Version]
- Liu, K.-C.; Yen, C.-Y.; Wu, R.S.-C.; Yang, J.-S.; Lu, H.-F.; Lu, K.-W.; Lo, C.; Chen, H.-Y.; Tang, N.-Y.; Wu, C.-C.; et al. The roles of endoplasmic reticulum stress and mitochondrial apoptotic signaling pathway in quercetin-mediated cell death of human prostate cancer PC-3 cells. Environ. Toxicol. 2014, 29, 428–439. [Google Scholar] [CrossRef]
- Ferruelo, A.; Romero, I.; Cabrera, P.; Arance, I.; Andrés, G.; Angulo, J. Los efectos de resveratrol y otros polifenoles del vino sobre la proliferación, apoptosis y expresión de receptor androgénico en células LNCaP. Actas Urol. Esp. 2014, 38, 397–404. [Google Scholar] [CrossRef]
- Siegel, R.L.; Miller, K.D.; Jemal, A. Cancer statistics, 2016. CA Cancer J. Clin. 2016, 66, 7–30. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sternberg, C.N.; Bellmunt, J.; Sonpavde, G.; Siefker-Radtke, A.O.; Stadler, W.M.; Bajorin, D.F.; Dreicer, R.; George, D.J.; Milowsky, M.I.; Theodorescu, D.; et al. ICUD-EAU International Consultation on Bladder Cancer 2012: Chemotherapy for Urothelial Carcinoma—Neoadjuvant and Adjuvant Settings. Eur. Urol. 2013, 63, 58–66. [Google Scholar] [CrossRef]
- Lu, Y.; Liu, L.; Wang, Y.; Li, F.; Zhang, J.; Ye, M.; Zhao, H.; Zhang, X.; Zhang, M.; Zhao, J.; et al. siRNA delivered by EGFR-specific scFv sensitizes EGFR-TKI-resistant human lung cancer cells. Biomaterials 2016, 76, 196–207. [Google Scholar] [CrossRef] [Green Version]
- Tsai, T.-F.; Hwang, T.I.-S.; Lin, J.-F.; Chen, H.-E.; Yang, S.-C.; Lin, Y.-C.; Chou, K.-Y. Suppression of Quercetin-induced autophagy enhances cytotoxicity through elevating apoptotic cell death in human bladder cancer cells. Urol. Sci. 2019, 30, 58. [Google Scholar] [CrossRef]
- Melo, M.B.; Ahmad, N.N.; Lima, C.S.; Pagnano, K.B.; Bordin, S.; Lorand-Metze, I.; Saad, S.T.; Costa, F.F. Mutations in the p53 Gene in Acute Myeloid Leukemia Patients Correlate with Poor Prognosis. Hematology 2002, 7, 13–19. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Steensma, D.P. Myelodysplastic Syndromes. Mayo Clin. Proc. 2015, 90, 969–983. [Google Scholar] [CrossRef] [Green Version]
- Hatziapostolou, M.; Iliopoulos, D. Epigenetic aberrations during oncogenesis. Cell. Mol. Life Sci. 2011, 68, 1681–1702. [Google Scholar] [CrossRef]
- Galm, O.; Herman, J.G.; Baylin, S.B. The fundamental role of epigenetics in hematopoietic malignancies. Blood Rev. 2006, 20, 1–13. [Google Scholar] [CrossRef]
- Calgarotto, A.K.; Maso, V.; Junior, G.C.F.; Nowill, A.E.; Filho, P.L.; Vassallo, J.; Saad, S.T.O. Antitumor activities of Quercetin and Green Tea in xenografts of human leukemia HL60 cells. Sci. Rep. 2018, 8, 1–7. [Google Scholar] [CrossRef] [Green Version]
- Alvarez, M.C.; Maso, V.; Torello, C.O.; Ferro, K.P.; Saad, S.T.O. The polyphenol quercetin induces cell death in leukemia by targeting epigenetic regulators of pro-apoptotic genes. Clin. Epigenetics 2018, 10, 1–11. [Google Scholar] [CrossRef]
- Maso, V.; Calgarotto, A.K.; Franchi, G.C.; Nowill, A.E.; Filho, P.L.; Vassallo, J.; Saad, S.T.O. Multitarget Effects of Quercetin in Leukemia. Cancer Prev. Res. 2014, 7, 1240–1250. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Alexander, D.D.; Mink, P.J.; Adami, H.-O.; Chang, E.T.; Cole, P.; Mandel, J.S.; Trichopoulos, D. The non-Hodgkin lymphomas: A review of the epidemiologic literature. Int. J. Cancer 2007, 120, 1–39. [Google Scholar] [CrossRef]
- Jemal, A.; Siegel, R.; Ward, E.; Murray, T.; Xu, J.; Thun, M.J. Cancer Statistics, 2007. CA Cancer J. Clin. 2007, 57, 43–66. [Google Scholar] [CrossRef]
- Kawahara, T.; Kawaguchi-Ihara, N.; Okuhashi, Y.; Itoh, M.; Nara, N.; Tohda, S. Cyclopamine and quercetin suppress the growth of leukemia and lymphoma cells. Anticancer. Res. 2009, 29, 4629–4632. [Google Scholar]
- Feitelson, M.A.; Arzumanyan, A.; Kulathinal, R.J.; Blain, S.W.; Holcombe, R.F.; Mahajna, J.; Marino, M.; Martinez-Chantar, M.L.; Nawroth, R.; Sanchez-Garcia, I.; et al. Sustained proliferation in cancer: Mechanisms and novel therapeutic targets. Semin. Cancer Biol. 2015, 35, S25–S54. [Google Scholar] [CrossRef] [PubMed]
- Cai, H.; Sale, S.; Schmid, R.; Britton, R.G.; Brown, K.; Steward, W.P.; Gescher, A.J. Flavones as Colorectal Cancer Chemopreventive Agents—Phenol-O-Methylation Enhances Efficacy. Cancer Prev. Res. 2009, 2, 743–750. [Google Scholar] [CrossRef] [Green Version]
- Maurya, A.K.; Vinayak, M. Quercetin Regresses Dalton’s Lymphoma Growth via Suppression of PI3K/AKT Signaling Leading to Upregulation of p53 and Decrease in Energy Metabolism. Nutr. Cancer 2015, 67, 354–363. [Google Scholar] [CrossRef]
- Frankenfeld, C.L.; Cerhan, J.R.; Cozen, W.; Davis, S.; Schenk, M.; Morton, L.M.; Hartge, P.; Ward, M.H. Dietary flavonoid intake and non-Hodgkin lymphoma risk. Am. J. Clin. Nutr. 2008, 87, 1439–1445. [Google Scholar] [CrossRef] [Green Version]
- Allemani, C.; Weir, H.K.; Carreira, H.; Harewood, R.; Spika, D.; Wang, X.-S.; Bannon, F.; Ahn, J.V.; Johnson, C.J.; Bonaventure, A.; et al. Global surveillance of cancer survival 1995–2009: Analysis of individual data for 25 676 887 patients from 279 population-based registries in 67 countries (CONCORD-2). Lancet 2015, 385, 977–1010. [Google Scholar] [CrossRef] [Green Version]
- Herbst, R.S.; Heymach, J.V.; Lippman, S.M. Lung Cancer. N. Engl. J. Med. 2008, 359, 1367–1380. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Shivappa, N.; Wang, R.; Hébert, J.R.; Jin, A.; Koh, W.-P.; Yuan, J.M. Association between inflammatory potential of diet and risk of lung cancer among smokers in a prospective study in Singapore. Eur. J. Nutr. 2018, 58, 2755–2766. [Google Scholar] [CrossRef] [PubMed]
- Altorki, N.K.; Markowitz, G.J.; Gao, D.; Port, J.L.; Saxena, A.; Stiles, B.; McGraw, T.; Mittal, V. The lung microenvironment: An important regulator of tumour growth and metastasis. Nat. Rev. Cancer 2019, 19, 9–31. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Takahashi, T.; Tateishi, A.; Bychkov, A.; Fukuoka, J. Remarkable Alteration of PD-L1 Expression after Immune Checkpoint Therapy in Patients with Non-Small-Cell Lung Cancer: Two Autopsy Case Reports. Int. J. Mol. Sci. 2019, 20, 2578. [Google Scholar] [CrossRef] [Green Version]
- Zanoaga, O.; Braicu, C.; Jurj, A.; Rusu, A.; Buiga, R.; Berindan-Neagoe, I. Progress in Research on the Role of Flavonoids in Lung Cancer. Int. J. Mol. Sci. 2019, 20, 4291. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Collins, I.; Workman, P. New approaches to molecular cancer therapeutics. Nat. Chem. Biol. 2006, 2, 689–700. [Google Scholar] [CrossRef] [PubMed]
- González-Loyola, A.; Fernández-Miranda, G.; Trakala, M.; Partida, D.; Samejima, K.; Ogawa, H.; Cañamero, M.; De Martino, A.; Martínez-Ramírez, Á.; De Cárcer, G.; et al. Aurora B Overexpression Causes Aneuploidy and p21Cip1Repression during Tumor Development. Mol. Cell. Biol. 2015, 35, 3566–3578. [Google Scholar] [CrossRef] [Green Version]
- Keizer, R.J.; Zandvliet, A.S.; Beijnen, J.H.; Schellens, J.H.M.; Huitema, A.D.R. Two-stage model-based design of cancer phase I dose escalation trials: Evaluation using the phase I program of barasertib (AZD1152). Investig. New Drugs 2012, 30, 1519–1530. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Dittrich, C.; Fridrik, M.A.; Koenigsberg, R.; Lee, C.; Goeldner, R.-G.; Hilbert, J.; Greil, R. A phase 1 dose escalation study of BI 831266, an inhibitor of Aurora kinase B, in patients with advanced solid tumors. Investig. New Drugs 2014, 33, 409–422. [Google Scholar] [CrossRef] [Green Version]
- Hasima, N.; Ozpolat, B. Regulation of autophagy by polyphenolic compounds as a potential therapeutic strategy for cancer. Cell Death Dis. 2014, 5, e1509. [Google Scholar] [CrossRef] [Green Version]
- Zhang, X.; Guo, Q.; Chen, J.; Chen, A.Z. Quercetin Enhances Cisplatin Sensitivity of Human Osteosarcoma Cells by Modulating microRNA-217-KRAS Axis. Mol. Cells 2015, 38, 638–642. [Google Scholar] [CrossRef] [Green Version]
- Dong, Y.; Yang, J.; Yang, L.; Li, P. Quercetin inhibits the proliferation and metastasis of human non-small cell lung cancer cell line: The key role of src-mediated fibroblast growth factor-inducible 14 (Fn14)/nuclear factor kappa B (NF-κB) pathway. Med. Sci. Monit. 2020, 26, e920537-1. [Google Scholar] [CrossRef]
- Meyers, P.A.; Gorlick, R. Osteosarcoma. Pediatr. Clin. N. Am. 1997, 44, 973–989. [Google Scholar] [CrossRef]
- Ostrom, Q.T.; Gittleman, H.; Fulop, J.; Liu, M.; Blanda, R.; Kromer, C.; Wolinsky, Y.; Kruchko, C.; Barnholtz-Sloan, J.S. CBTRUS Statistical Report: Primary Brain and Central Nervous System Tumors Diagnosed in the United States in 2008-2012. Neuro-Oncol. 2015, 17, iv1–iv62. [Google Scholar] [CrossRef]
- Afshari, A.R.; Jalili-Nik, M.; Soukhtanloo, M.; Ghorbani, A.; Sadeghnia, H.R.; Mollazadeh, H.; Roshan, M.K.; Rahmani, F.; Sabri, H.; Vahedi, M.M.; et al. Auraptene-induced cytotoxicity mechanisms in human malignant glioblastoma (U87) cells: Role of reac-tive oxygen species (ROS). EXCLI J. 2019, 18, 576–590. [Google Scholar] [CrossRef] [PubMed]
- Braganhol, E.; Zamin, L.L.; Canedo, A.D.; Horn, F.; Tamajusuku, A.S.; Wink, M.R.; Salbego, C.; Battastini, A.M. Antiproliferative effect of quercetin in the human U138MG glioma cell line. Anti-Cancer Drugs 2006, 17, 663–671. [Google Scholar] [CrossRef]
- Kim, E.J.; Choi, C.H.; Park, J.Y.; Kang, S.K.; Kim, Y.K. Underlying Mechanism of Quercetin-induced Cell Death in Human Glioma Cells. Neurochem. Res. 2008, 33, 971–979. [Google Scholar] [CrossRef]
- Tavana, E.; Mollazadeh, H.; Mohtashami, E.; Modaresi, S.M.S.; Hosseini, A.; Sabri, H.; Soltani, A.; Javid, H.; Afshari, A.R.; Sahebkar, A. Quercetin: A promising phytochemical for the treatment of glioblastoma multiforme. BioFactors 2019, 46, 356–366. [Google Scholar] [CrossRef]
- Pan, H.-C.; Jiang, Q.; Yu, Y.; Mei, J.-P.; Cui, Y.-K.; Zhao, W.-J. Quercetin promotes cell apoptosis and inhibits the expression of MMP-9 and fibronectin via the AKT and ERK signalling pathways in human glioma cells. Neurochem. Int. 2015, 80, 60–71. [Google Scholar] [CrossRef]
- Santos, B.L.; Oliveira, M.N.; Coelho, P.L.; Pitanga, B.P.; da Silva, A.B.; Adelita, T.; Silva, V.D.A.; Costa, M.D.F.; El-Bachá, R.S.; Tardy, M.; et al. Flavonoids suppress human glioblastoma cell growth by inhibiting cell metabolism, migration, and by regulating extracellular matrix proteins and metalloproteinases expression. Chem. Biol. Interact. 2015, 242, 123–138. [Google Scholar] [CrossRef]
- Liu, Y.; Tang, Z.-G.; Yang, J.-Q.; Zhou, Y.; Meng, L.-H.; Wang, H.; Li, C.-L. Low concentration of quercetin antagonizes the invasion and angiogenesis of human glioblastoma U251 cells. OncoTargets Ther. 2017, 10, 4023–4028. [Google Scholar] [CrossRef] [Green Version]
- Sheu, J.J.-C.; Hua, C.-H.; Wan, L.; Lin, Y.-J.; Lai, M.-T.; Tseng, H.-C.; Jinawath, N.; Tsai, M.-H.; Chang, N.-W.; Lin, C.-F.; et al. Functional Genomic Analysis Identified Epidermal Growth Factor Receptor Activation as the Most Common Genetic Event in Oral Squamous Cell Carcinoma. Cancer Res. 2009, 69, 2568–2576. [Google Scholar] [CrossRef] [Green Version]
- Chen, S.-F.; Nien, S.; Wu, C.-H.; Liu, C.-L.; Chang, Y.-C.; Lin, Y.-S. Reappraisal of the anticancer efficacy of quercetin in oral cancer cells. J. Chin. Med. Assoc. 2013, 76, 146–152. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Davoodvandi, A.; Varkani, M.S.; Clark, C.C.T.; Jafarnejad, S. Quercetin as an anticancer agent: Focus on esophageal cancer. J. Food Biochem. 2020, 44, 13374. [Google Scholar] [CrossRef]
- A Anderson, L.; Watson, R.P.; Murphy, S.J.; Johnston, B.T.; Comber, H.; Mc Guigan, J.; Reynolds, J.V.; Murray, L.J. Risk factors for Barrett’s oesophagus and oesophageal adenocarcinoma: Results from the FINBAR study. World J. Gastroenterol. 2007, 13, 1585–1594. [Google Scholar] [CrossRef] [Green Version]
- Thangasamy, T.; Sittadjody, S.; Lanza-Jacoby, S.; Wachsberger, P.R.; Limesand, K.H.; Burd, R. Quercetin Selectively Inhibits Bioreduction and Enhances Apoptosis in Melanoma Cells that Overexpress Tyrosinase. Nutr. Cancer 2007, 59, 258–268. [Google Scholar] [CrossRef]
- Vargas, A.J.; Burd, R. Hormesis and synergy: Pathways and mechanisms of quercetin in cancer prevention and management. Nutr. Rev. 2010, 68, 418–428. [Google Scholar] [CrossRef]
- Guo, X.; He, D.; Yang, L.; Li, Y.; Chen, Q.; Chen, J.; Zhang, E.; Lin, X.; He, J.; Yang, Y.; et al. Quercetin Displays Anti-Myeloma Activity and Synergistic Effect with Dexamethasone in Vitro and In Vivo Xenograft Models. Blood 2015, 126, 5339. [Google Scholar] [CrossRef]
- Ma, Y.; Jin, Z.; Huang, J.; Zhou, S.; Ye, H.; Jiang, S.; Yu, K. Quercetin suppresses the proliferation of multiple myeloma cells by down-regulating IQ motif-containing GTPase activating protein 1 expression and extracellular signal-regulated kinase activation. Leuk. Lymphoma 2014, 55, 2597–2604. [Google Scholar] [CrossRef]
- Vogel, F. Genetics of retinoblastoma. Qual. Life Res. 1979, 52, 1–54. [Google Scholar] [CrossRef]
- Shields, C.L.; Shields, J.A. Basic understanding of current classification and management of retinoblastoma. Curr. Opin. Ophthalmol. 2006, 17, 228–234. [Google Scholar] [CrossRef] [PubMed]
- Saxena, P.; Kaur, J. Differential expression of genes in retinoblastoma. Clin. Chim. Acta 2011, 412, 2015–2021. [Google Scholar] [CrossRef]
- Rössler, J.; Dietrich, T.; Pavlakovic, H.; Schweigerer, L.; Havers, W.; Schuler, A.; Bornfeld, N.; Schilling, H. Higher Vessel Densities in Retinoblastoma with Local Invasive Growth and Metastasis. Am. J. Pathol. 2004, 164, 391–394. [Google Scholar] [CrossRef] [Green Version]
- Murakami, A.; Ashida, H.; Terao, J. Multitargeted cancer prevention by quercetin. Cancer Lett. 2008, 269, 315–325. [Google Scholar] [CrossRef] [PubMed]
- Houston, S.K.; Murray, T.G.; Wolfe, S.Q.; Fernandes, C.E. Current Update on Retinoblastoma. Int. Ophthalmol. Clin. 2011, 51, 77–91. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Song, W.; Zhao, X.; Xu, J.; Zhang, H. Quercetin inhibits angiogenesis-mediated human retinoblastoma growth by targeting vascular endothelial growth factor receptor. Oncol. Lett. 2017, 14, 3343–3348. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Liu, H.; Zhou, M. Antitumor effect of Quercetin on Y79 retinoblastoma cells via activation of JNK and p38 MAPK pathways. BMC Complement. Altern. Med. 2017, 17, 531. [Google Scholar] [CrossRef] [Green Version]
- Gibellini, L.; Pinti, M.; Nasi, M.; Montagna, J.P.; De Biasi, S.; Roat, E.; Bertoncelli, L.; Cooper, E.L.; Cossarizza, A. Quercetin and Cancer Chemoprevention. Evid. Based Complement. Altern. Med. 2011, 2011, 1–15. [Google Scholar] [CrossRef] [Green Version]
- Ong, C.S.; Tran, E.; Nguyen, T.T.; Ong, C.K.; Lee, S.K.; Lee, J.J.; Ng, C.P.; Leong, C.; Huynh, H. Quercetin-induced growth inhibition and cell death in nasopharyngeal carcinoma cells are associated with increase in Bad and hypophosphorylated retinoblastoma expressions. Oncol. Rep. 2004, 11, 727–733. [Google Scholar] [CrossRef]
- Jackson, M.J. The assessment of bioavailability of micronutrients: Introduction. Eur. J. Clin. Nutr. 1997, 51, 1–2. [Google Scholar]
- Rein, M.J.; Renouf, M.; Cruz-Hernandez, C.; Actis-Goretta, L.; Thakkar, S.K.; Da Silva Pinto, M. Bioavailability of bioactive food compounds: A challenging journey to bioefficacy. Br. J. Clin. Pharmacol. 2013, 75, 588–602. [Google Scholar] [CrossRef] [Green Version]
- Erlund, I.; Kosonen, T.; Alfthan, G.; Mäenpää, J.; Perttunen, K.; Kenraali, J.; Parantainen, J.; Aro, A. Pharmacokinetics of quercetin from quercetin aglycone and rutin in healthy volunteers. Eur. J. Clin. Pharmacol. 2000, 56, 545–553. [Google Scholar] [CrossRef] [PubMed]
- Ader, P. Bioavailability and metabolism of the flavonol quercetin in the pig. Free Radic. Biol. Med. 2000, 28, 1056–1067. [Google Scholar] [CrossRef]
- Graefe, E.U.; Derendorf, H.; Veit, M. Pharmacokinetics and bioavailability of the flavonol quercetin in humans. Int. J. Clin. Pharmacol. Ther. 1999, 37, 219–233. [Google Scholar] [PubMed]
- Crespy, V.; Morand, C.; Besson, C.; Manach, C.; Demigne, C.; Remesy, C. Quercetin, but not Its Glycosides, Is Absorbed from the Rat Stomach. J. Agric. Food Chem. 2002, 50, 618–621. [Google Scholar] [CrossRef]
- Toutain, P.L.; Bousquet-Melou, A. Bioavailability and its assessment. J. Veter-Pharmacol. Ther. 2004, 27, 455–466. [Google Scholar] [CrossRef]
- Lee, J.; Ebeler, S.E.; Zweigenbaum, J.A.; Mitchell, A.E. UHPLC-(ESI)QTOF MS/MS Profiling of Quercetin Metabolites in Human Plasma Postconsumption of Applesauce Enriched with Apple Peel and Onion. J. Agric. Food Chem. 2012, 60, 8510–8520. [Google Scholar] [CrossRef] [PubMed]
- Hollman, P.C.; Van Trijp, J.M.; Buysman, M.N.; Gaag, M.S.V.; Mengelers, M.J.; De Vries, J.H.; Katan, M.B. Relative bioavailability of the antioxidant flavonoid quercetin from various foods in man. FEBS Lett. 1997, 418, 152–156. [Google Scholar] [CrossRef] [Green Version]
- Day, A.J.; Cañada, F.J.; Díaz, J.C.; Kroon, P.A.; Mclauchlan, R.; Faulds, C.B.; Plumb, G.W.; Morgan, M.R.; Williamson, G. Dietary flavonoid and isoflavone glycosides are hydrolysed by the lactase site of lactase phlorizin hydrolase. FEBS Lett. 2000, 468, 166–170. [Google Scholar] [CrossRef] [Green Version]
- Hollman, P.C.; De Vries, J.H.; Van Leeuwen, S.D.; Mengelers, M.J.; Katan, M.B. Absorption of dietary quercetin glycosides and quercetin in healthy ileostomy volunteers. Am. J. Clin. Nutr. 1995, 62, 1276–1282. [Google Scholar] [CrossRef] [Green Version]
- Hollman, P.C.; van Trijp, J.M.; Mengelers, M.J.; de Vries, J.H.; Katan, M.B. Bioavailability of the dietary antioxidant flavonol quercetin in man. Cancer Lett. 1997, 114, 139–140. [Google Scholar] [CrossRef]
- Russo, M.; Spagnuolo, C.; Tedesco, I.; Bilotto, S.; Russo, G.L. The flavonoid quercetin in disease prevention and therapy: Facts and fancies. Biochem. Pharmacol. 2012, 83, 6–15. [Google Scholar] [CrossRef]
- Dabeek, W.M.; Marra, M.V. Dietary Quercetin and Kaempferol: Bioavailability and Potential Cardiovascular-Related Bioactivity in Humans. Nutrients 2019, 11, 2288. [Google Scholar] [CrossRef] [Green Version]
- Jiang, H.; Engelhardt, U.H.; Thräne, C.; Maiwald, B.; Stark, J. Determination of flavonol glycosides in green tea, oolong tea and black tea by UHPLC compared to HPLC. Food Chem. 2015, 183, 30–35. [Google Scholar] [CrossRef]
- Geleijnse, J.M.; Launer, L.J.; Kuip, D.A.M.V.D.; Hofman, A.; Witteman, J.C.M. Inverse association of tea and flavonoid intakes with incident myocardial infarction: The Rotterdam Study. Am. J. Clin. Nutr. 2002, 75, 880–886. [Google Scholar] [CrossRef]
- Kaşıkcı, M.; Bağdatlıoğlu, N. Bioavailability of Quercetin. Curr. Res. Nutr. Food Sci. J. 2016, 4, 146–151. [Google Scholar] [CrossRef]
- Lesser, S.; Cermak, R.; Wolffram, S. Bioavailability of Quercetin in Pigs Is Influenced by the Dietary Fat Content. J. Nutr. 2004, 134, 1508–1511. [Google Scholar] [CrossRef] [PubMed]
- Guo, Y.; Mah, E.; Davis, C.G.; Jalili, T.; Ferruzzi, M.G.; Chun, O.K.; Bruno, R.S. Dietary fat increases quercetin bioavailability in overweight adults. Mol. Nutr. Food Res. 2013, 57, 896–905. [Google Scholar] [CrossRef] [PubMed]
- Rothwell, J.A.; Day, A.J.; Morgan, M.R.A. Experimental Determination of Octanol−Water Partition Coefficients of Quercetin and Related Flavonoids. J. Agric. Food Chem. 2005, 53, 4355–4360. [Google Scholar] [CrossRef] [PubMed]
- Guo, Y.; Bruno, R.S. Endogenous and exogenous mediators of quercetin bioavailability. J. Nutr. Biochem. 2015, 26, 201–210. [Google Scholar] [CrossRef]
- Almeida, A.F.; Borge, G.I.A.; Piskula, M.; Tudose, A.; Tudoreanu, L.; Valentová, K.; Williamson, G.; Santos, C.N. Bioavailability of Quercetin in Humans with a Focus on Interindividual Variation. Compr. Rev. Food Sci. Food Saf. 2018, 17, 714–731. [Google Scholar] [CrossRef] [Green Version]
- Quagliariello, V.; Armenia, E.; Aurilio, C.; Rosso, F.; Clemente, O.; De Sena, G.; Barbarisi, M.; Barbarisi, A. New Treatment of Medullary and Papillary Human Thyroid Cancer: Biological Effects of Hyaluronic Acid Hydrogel Loaded With Quercetin Alone or in Combination to an Inhibitor of Aurora Kinase. J. Cell. Physiol. 2016, 231, 1784–1795. [Google Scholar] [CrossRef] [PubMed]
- Riva, A.; Ronchi, M.; Petrangolini, G.; Bosisio, S.; Allegrini, P. Improved Oral Absorption of Quercetin from Quercetin Phytosome®, a New Delivery System Based on Food Grade Lecithin. Eur. J. Drug Metab. Pharmacokinet. 2019, 44, 169–177. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gee, J.M.; Dupont, M.S.; Day, A.J.; Plumb, G.W.; Williamson, G.; Johnson, I.T. Intestinal Transport of Quercetin Glycosides in Rats Involves Both Deglycosylation and Interaction with the Hexose Transport Pathway. J. Nutr. 2000, 130, 2765–2771. [Google Scholar] [CrossRef]
- Hollman, P.C.H.; Katan, M.B. Bioavailability and Health Effects of Dietary Flavonols in Man. Arch. Toxicol. 1998, 20, 237–248. [Google Scholar] [CrossRef] [Green Version]
- Jung, S.Y.; Choi, S.; Ko, Y.S.; Park, C.S.; Oh, S.; Koh, S.R.; Oh, U.; Oh, J.W.; Rhee, M.H.; Nah, S.Y. Effects of ginsenosides on vanilloid receptor (VR1) channels expressed in Xenopus oocytes. Mol. Cells 2001, 12, 342–346. [Google Scholar]





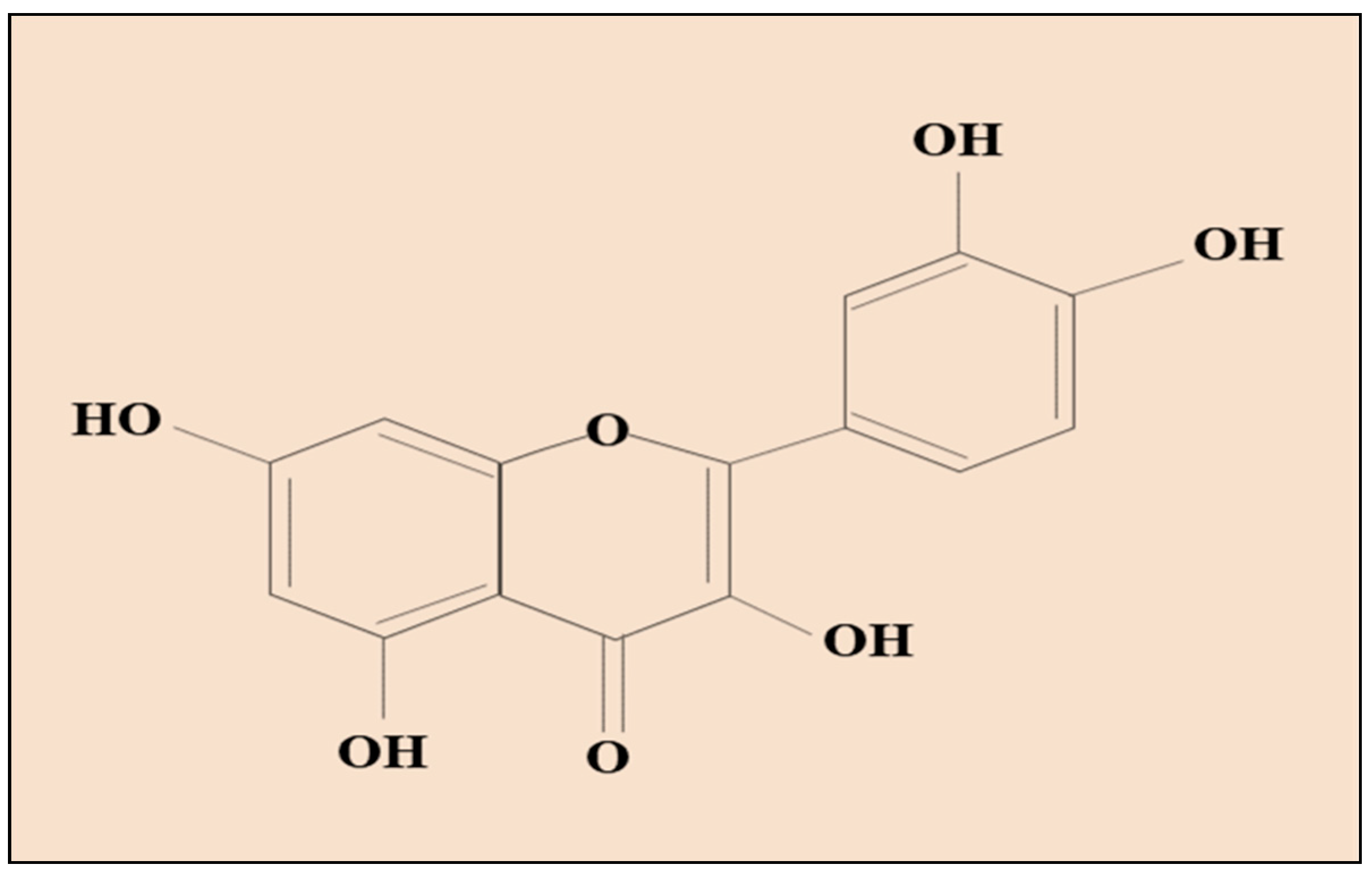{kind=link}
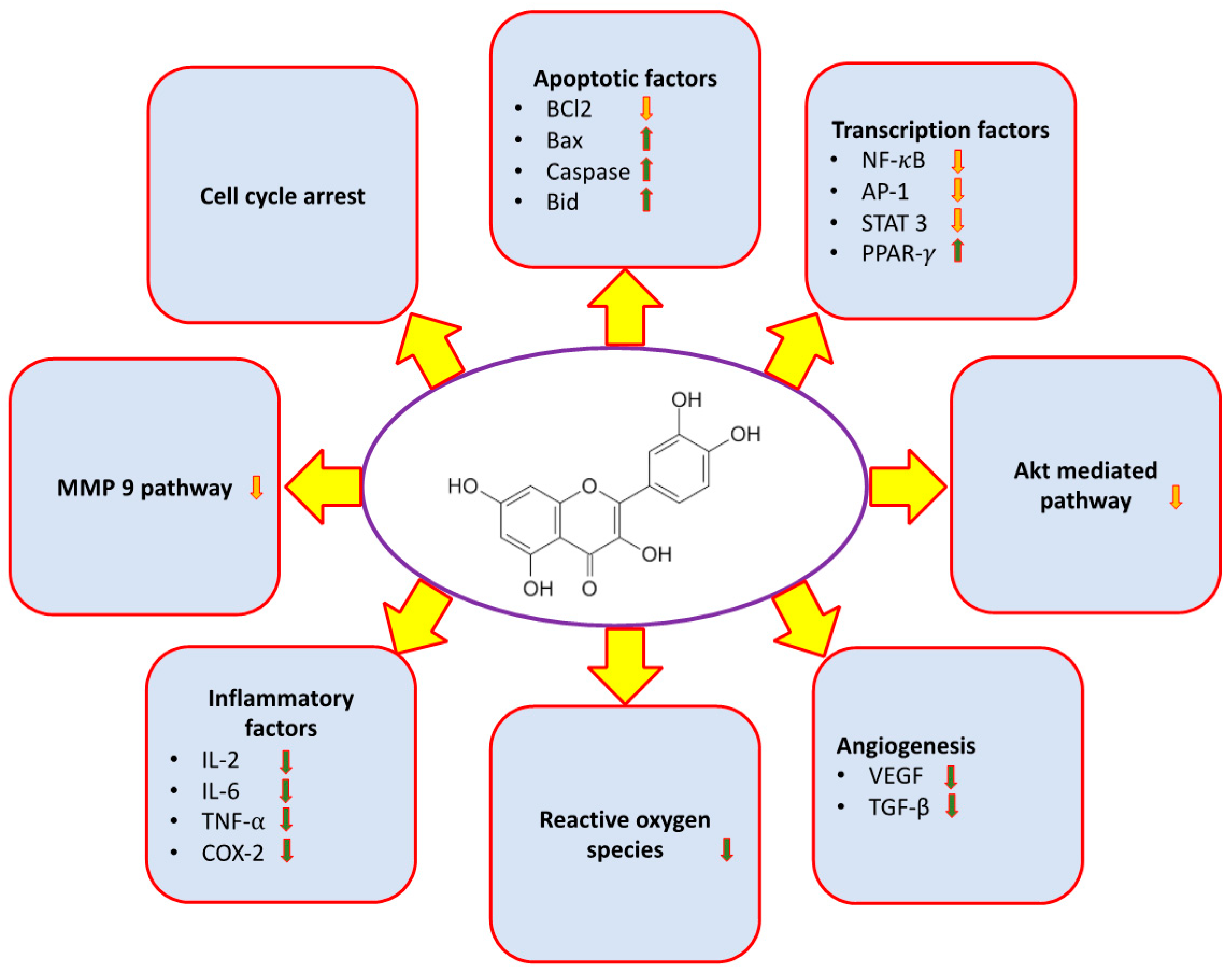{kind=link}
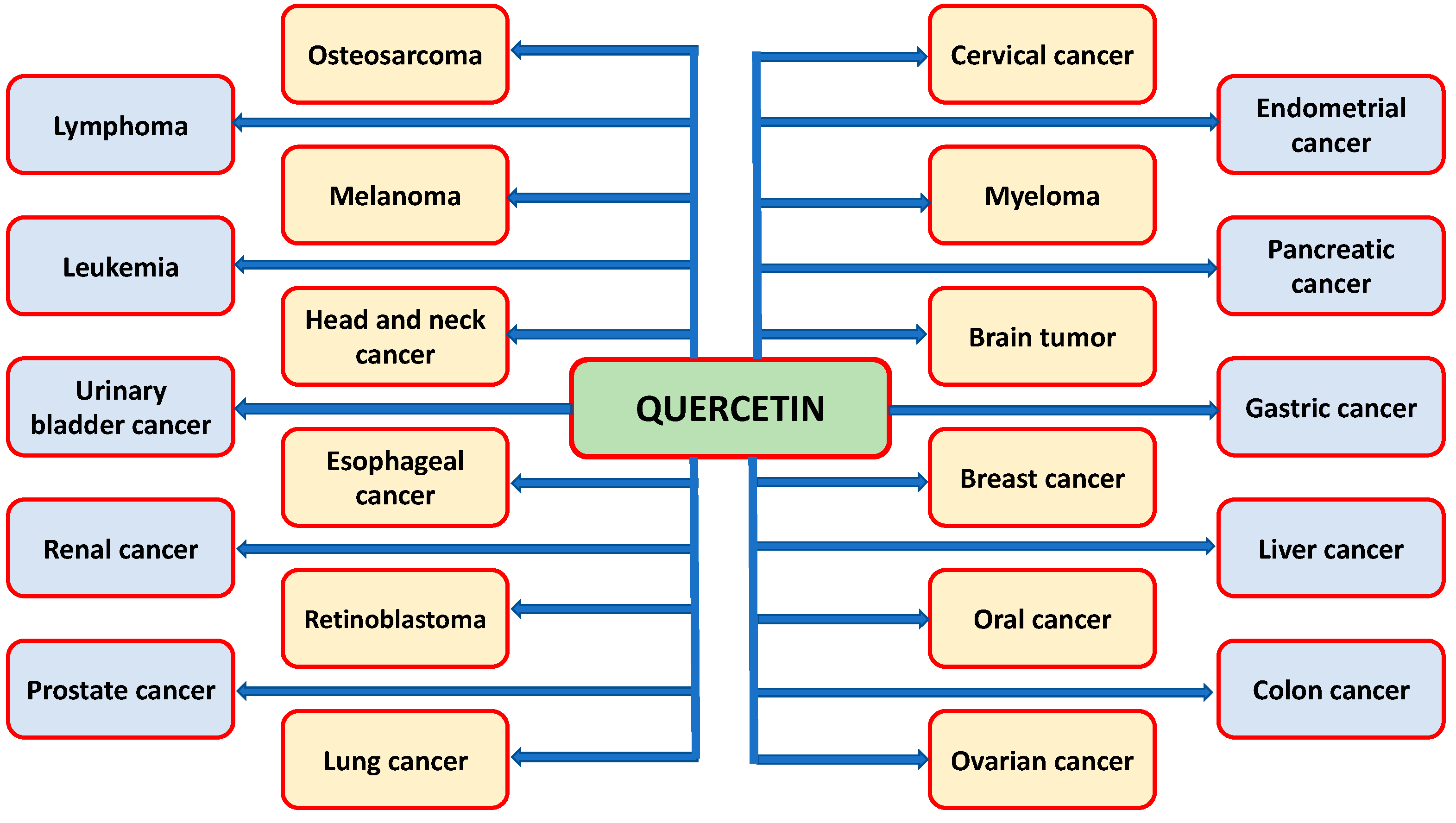{kind=link}
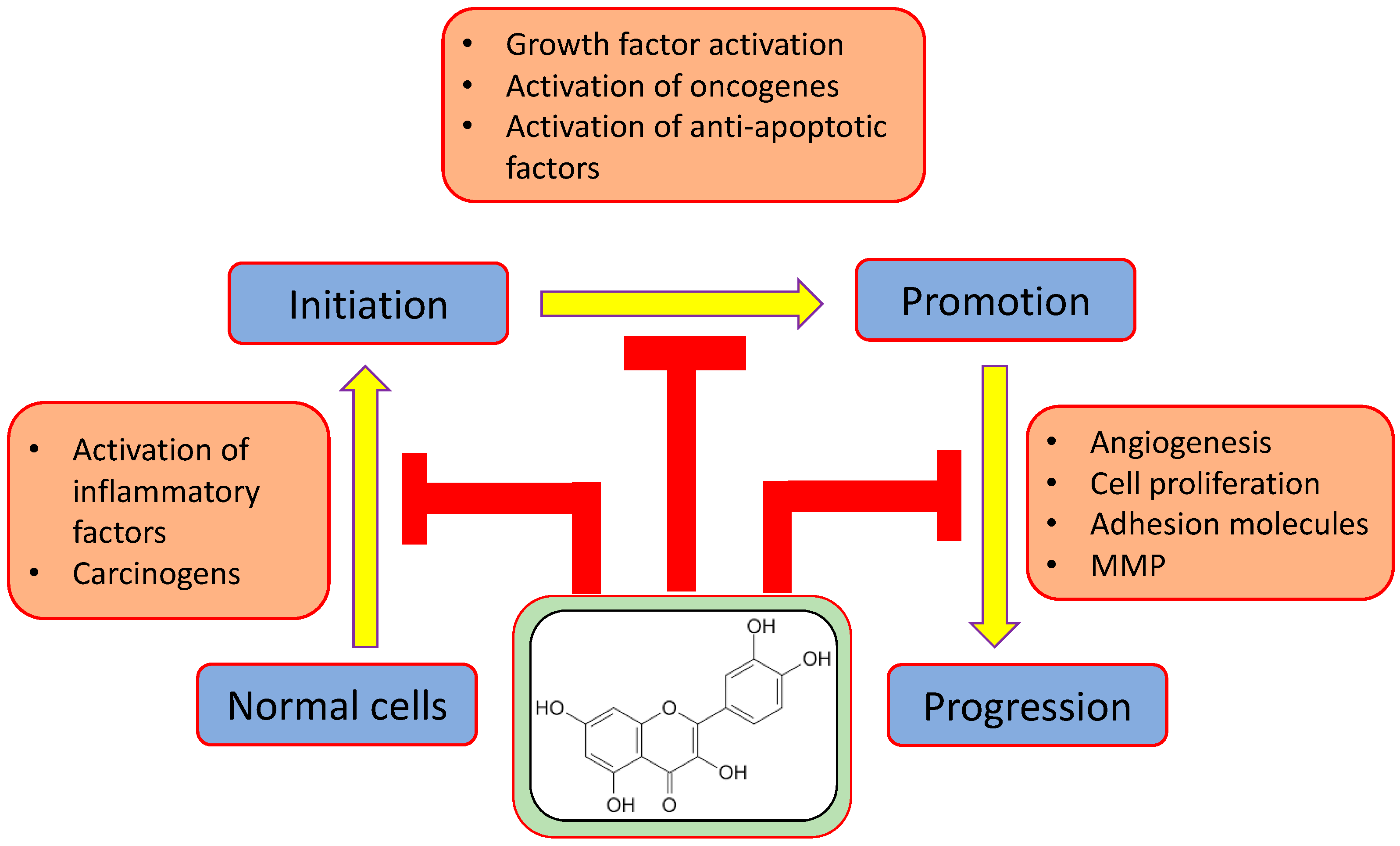{kind=link}
| Major Mechanism | Result of the Study | Refs |
|---|---|---|
| Inflammation | Quercetin discourages cancer development by hindering the development of inflammation producing enzymes (protein kinase C, cyclooxygenase, and lipoxygenase) and reducing proinflammatory-mediators production. | [59] |
| Inflammation | Inhibitory effects of Qu associated with prostate cancer have been demonstrated through several clinical studies, where it reduces inflammation and associated risk of prostate cancer. | [68,69] |
| Reactive Oxygen Species (ROS) | The ROS production in (prostate cancer) prostate cancer cells through its pronounced impact on mitochondrial integrity and its antioxidant has been confirmed by Qu treatment. | [75] |
| Reactive Oxygen Species (ROS) | Lipid oxidation and iron-catalyzed ROS production is inhibited by Qu treatment. | [77] |
| Angiogenesis | The antitumor effects of Qu has been explained through its anti-angiogenesis effects. | [80] |
| Angiogenesis | Suppression of ovarian cancer cell growth by intravenously administering QU/m-PEG-PCL micelles, which, amazingly, inhibit the growth of xenograft ovarian tumors by repressing angiogenesis, which reflect novel application of Qu in treatment of ovarian cancer. | [81] |
| Apoptosis | Qu is associated with stimulating intrinsic pathways and increases the release of cytochrome c from the mitochondria to the cytoplasm. Both processes are responsible for death of tumor cells and inhibit chances of cancer progression. | [91] |
| Apoptosis | The induction of apoptosis has been noticed in the Qu treatment gastric cancer stem cell. | [92] |
| Cell cycle | Qu therapy might lead to G0/G1 cell-cycle arrest in leukemia. | [98,99] |
| Cell cycle | The anticancer role of Qu is measured through the induction of apoptosis and G2-M cell cycle arrest. | [105] |
| Phosphatidylinositide-3-kinase (PI3k)/Protein kinase B (PKB) pathways | Qu mediates modulation of the PI3K/Akt signaling pathway and modulates suppression of the PI3/Akt signaling pathway survival signals in inhibition of growth of lymphoma. | [114] |
| Signal transducer and activator of transcription 3 (STAT3) | The anticancer activity of Qu has been identified through the reduction of glioblastoma cell proliferation and migration via suppressing activation of Signal transducer and activator of transcription 3 (STAT3). | [115,117] |
| Epidermal Growth Factor Receptor (EGFR) | In the in vitro model, Qu inhibits EGF-induced Epithelial-to-mesenchymal transition through the EGFR/PI3K/AKt/ERK-1/2 pathway and suppresses the transcriptional repressor slug, twist, and snail in prostrate cancerous cell line PC-3. Hence, Qu inhibits metastasis of cancer by aiming EMT. | [131] |
| Types of Cancer | Mechanisms/Results of the Study | Refs. |
|---|---|---|
| Cervix cancer | Qu causes cell death in cervix cancer by decreasing O-GlcNAcylation of adenosine monophosphate activated protein kinase. | [139] |
| Cervix cancer | Qu steadily alters the WNT, PI3K, and MAPK pathways by regulating the expressions of various proteins, which leads to the inhibition of cell cycle arrest, cell proliferation, apoptosis, and DNA damage in cervical cancerous cells. | [140] |
| Breast cancer | Human breast cancer MDA-MB-231 cell death via mitochondrial- and caspase-3 dependent pathways. | [56] |
| Breast cancer | Qu targets the VEGFR-2 mediated angiogenesis pathway, suppresses the expression of the downstream regulatory factor AKT, and inhibits growth of tumor. | [80,85] |
| Ovarian cancer | Qu interferes with intracellular signaling pathway induced apoptosis through mitochondrial intrinsic and caspase dependent pathways. | [141] |
| Ovarian cancer | The intervention of Qu leads to ER stress, increased expression of Bax, p21, and p53, reduced Bcl-2 expression, continued repair of DNA, and causes radio sensitization in ovarian cancerous cell. | [142] |
| Endometrial cancer | The flavonoids, including Qu, can repress the cancerous cells metastasis via reducing expression of c-Myc. | [143] |
| Pancreatic cancer | Self-renewal capability of putative pancreatic cancerous stem cells was inhibited by Qu. | [144] |
| Pancreatic cancer | Qu, alone and in combination with gemcitabine, robustly induced cell death, leading to a decrease in cell growth in pancreatic cancer cells. | [145] |
| Pancreatic cancer | Cluster of differentiation (CD)36 is targeted that decreases the death rate caused by pancreatic cancer by increasing cell adhesion, facilitating fatty acids uptake, regulating thrombospondin 1, and increasing immune response. | [146] |
| Gastric cancer | LC3 turnover induced by Qu and activated autophagy related genes in gastric cancer cells. | [147] |
| Gastric cancer | Qu reduces AGS and BGC823 cell invasion and migration, accompanied by decreased uPAR and uPA protein expression, blocking signaling pathways, finally repressing GC cells. | [148] |
| Hepatocellular carcinoma | The anticancer functions of Qu has been noticed in hepatocellular carcinoma. | [149] |
| Hepatocellular carcinoma | The role of Qu has been noted as it suppress proliferation of HCC and encourage apoptosis. Qu prevented metastasis of LM3 cells by regulating expression of vimentin, N-cadherin, MMP9, and E-cadherin. | [150] |
| Hepatocellular carcinoma | Qu prevents the liver cancerous cells proliferation through initiation of apoptosis and cell cycle arrest. | [151] |
| Colon cancer | Induction of apoptosis was noticed in colon cancer through the Qu treatment. | [152] |
| Renal cancer | Qu mitigates the adverse impact of sunitinib (a primary renal cancer treating drug); Qu-3-O-β-d-glucopyranoside was observed—that is, iso-Qu has an enhanced pharmacokinetic report. | [153] |
| Prostate cancer | Qu provokes prostate cancer by decreasing expression of androgen receptor (AR), by causing apoptosis and by repressing proliferation. | [154] |
| Prostate cancer | A total of 150 mg per kg was administered intraperitoneally in DU-145 and PC-3 xenograft tumor models; it repressed growth of xenograft tumor ascribed to the antagonization of expression of HSP72. | [111] |
| Bladder cancer | Dose dependent administration of Qu caused induction of apoptosis. | [155] |
| Bladder cancer | Cell proliferation and formation of colony of cancerous cells was inhibited by Qu; Qu may be an efficient chemopreventive and chemotherapeutical agent. | [156] |
| Leukemia | Qu synergistically sensitizes to apoptosis numerous leukemic cell lines and B-cells isolated from patients of CLL, when associated with other proapoptotic agents. | [157] |
| Leukemia | It can be used as a warning factor, along with TRAIL, promoting the impact of TRAIL-induced apoptosis in KG-1 cells. | [158] |
| Lymphoma | Qu restores TRAIL induced cell death in resistant transformed follicular lymphoma. | [159] |
| Lung cancer | Antigrowth of cancer was noted in Qu treatment. | [160] |
| Lung cancer | Qu plays a role in enhancement of the gene expression linked with death receptor signaling and inhibition of cell cycle, but reduces the expression of genes involved in activation of NF-κB. | [65] |
| Osteosarcoma | Qu attenuates invasion and migration of cells in MG63 and HOS cells in comparison with treatment with control medium and may have potential as a therapy for human osteosarcoma. | [161] |
| Osteosarcoma | It significantly reduced the mRNA expression levels of PTHR1 and consequent knockdown of PTHR1, and enhanced Qu-inhibited proliferation and invasion. | [162] |
| Glioblastoma | The anticancer activity of Qu was proven through repressing the PI3K-Akt pathway. | [163] |
| Glioblastoma | The growth of glioblastoma cells and induction of apoptosis was measured in glioblastoma cells. | [164] |
| Head and Neck cancer | Qu is an efficient anticancer agent in EGFR-overexpressing HNSCC. | [165] |
| Head and Neck cancer | The in vivo and in vitro studies have revealed that Qu has anticancer activity. | [166] |
| Oral cancer | The invasion and migration of human oral cancer cells is inhibited by Qu via inhibition of matrix metalloproteinase-2/-9 and NF-κB signaling pathways. | [167] |
| Esophagus cancer | Qu-3-O-β-d-glucopyranoside on gastritis and reflux esophagitis in rats have been studied by Min et al. (2009), and findings demonstrate that Qu-3-O-β-d-3-O-β-d-glucopyranoside can prevent gastritis and reflux esophagitis in rats. | [168] |
| Melanoma | Qu quickly metabolizes tyrosinase into several compounds, which encourage anticancer activity. It suggests that Qu can be used to target melanoma. | [169] |
| Myeloma | Qu encourages apoptosis of myeloma cells and shows a synergetic effect with dexamethasone in vitro and in vivo xenograft models. | [105] |
| Retinoblastoma | The antitumor effects of Qu occur by inhibiting metastasis progression, cell proliferation, and angiogenesis, modifying cell cycle progression, encouraging apoptosis, and influencing autophagy, including retinoblastoma. | [15,170] |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Almatroodi, S.A.; Alsahli, M.A.; Almatroudi, A.; Verma, A.K.; Aloliqi, A.; Allemailem, K.S.; Khan, A.A.; Rahmani, A.H. Potential Therapeutic Targets of Quercetin, a Plant Flavonol, and Its Role in the Therapy of Various Types of Cancer through the Modulation of Various Cell Signaling Pathways. Molecules 2021, 26, 1315. https://doi.org/10.3390/molecules26051315
Almatroodi SA, Alsahli MA, Almatroudi A, Verma AK, Aloliqi A, Allemailem KS, Khan AA, Rahmani AH. Potential Therapeutic Targets of Quercetin, a Plant Flavonol, and Its Role in the Therapy of Various Types of Cancer through the Modulation of Various Cell Signaling Pathways. Molecules. 2021; 26(5):1315. https://doi.org/10.3390/molecules26051315
Chicago/Turabian StyleAlmatroodi, Saleh A., Mohammed A. Alsahli, Ahmad Almatroudi, Amit Kumar Verma, Abdulaziz Aloliqi, Khaled S. Allemailem, Amjad Ali Khan, and Arshad Husain Rahmani. 2021. "Potential Therapeutic Targets of Quercetin, a Plant Flavonol, and Its Role in the Therapy of Various Types of Cancer through the Modulation of Various Cell Signaling Pathways" Molecules 26, no. 5: 1315. https://doi.org/10.3390/molecules26051315