
Suppressive Effect of Two Cucurbitane-Type Triterpenoids from Momordica charantia on Cutibacterium acnes-Induced Inflammatory Responses in Human THP-1 Monocytic Cell and Mouse Models

 ,
,  ,
,
Abstract
:1. Introduction
2. Results and Discussion
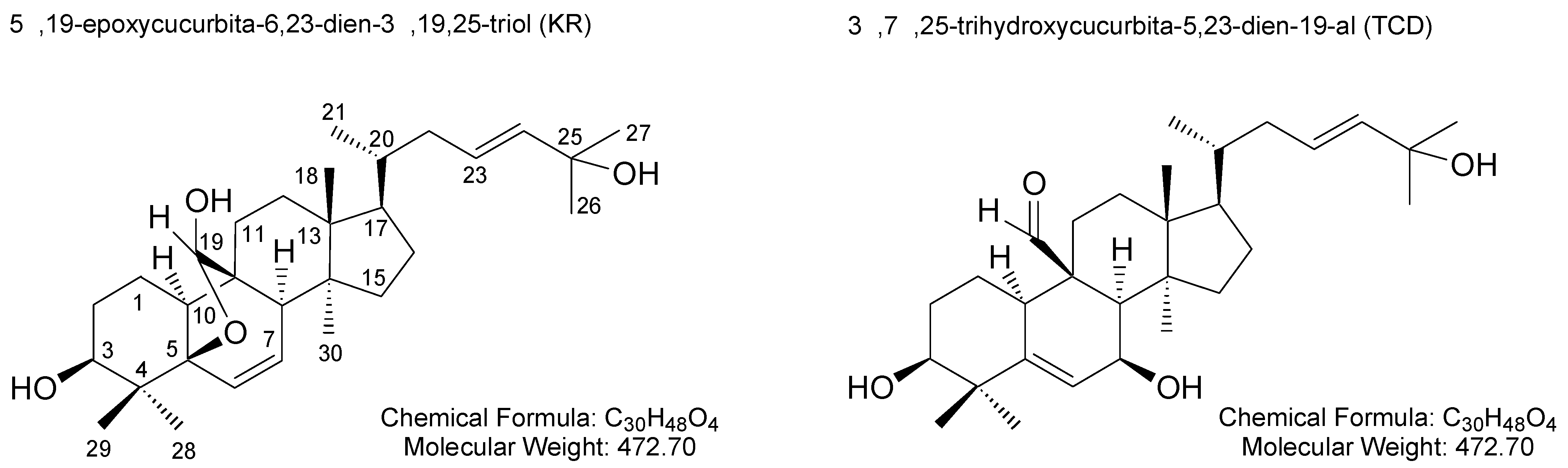
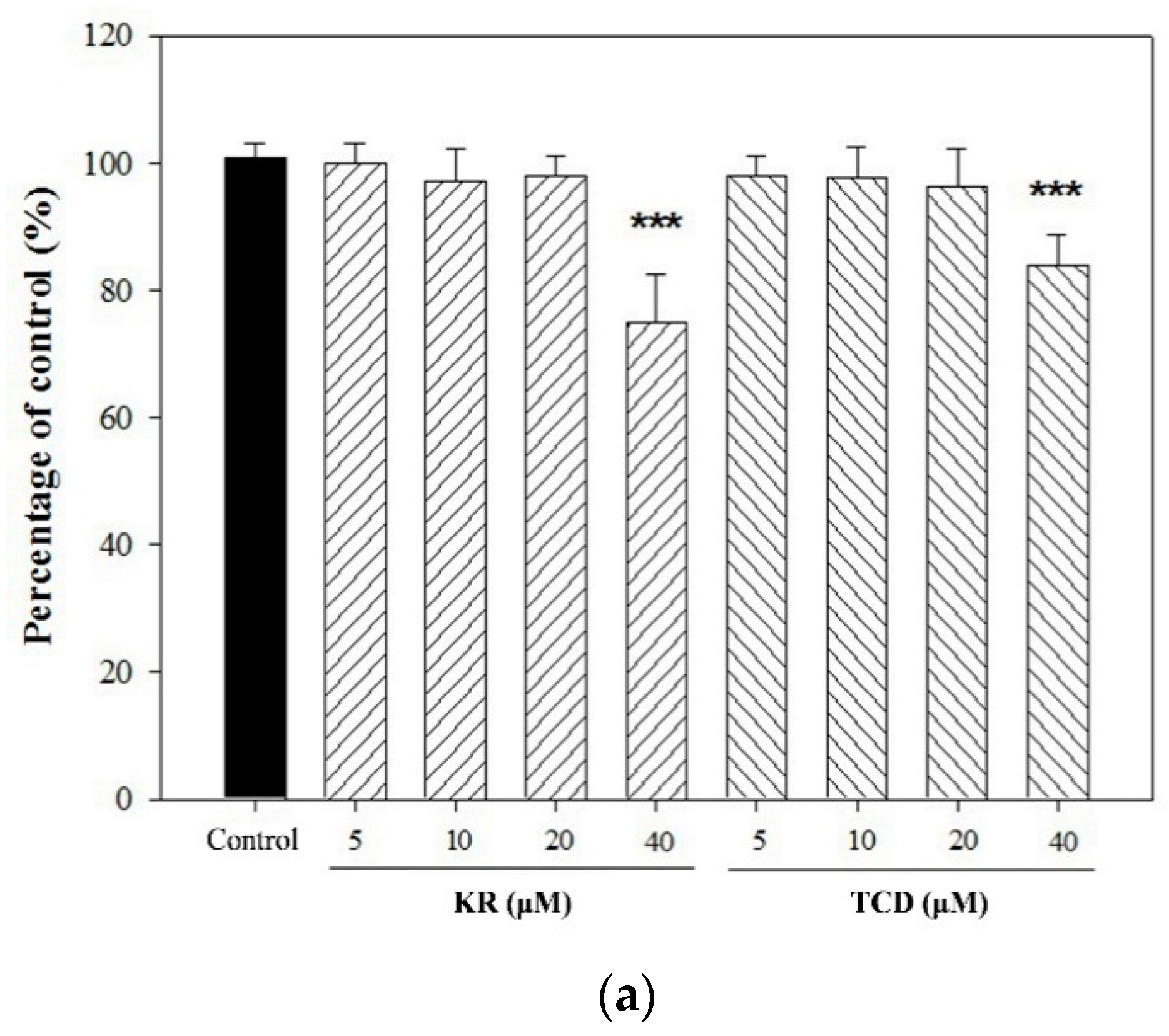
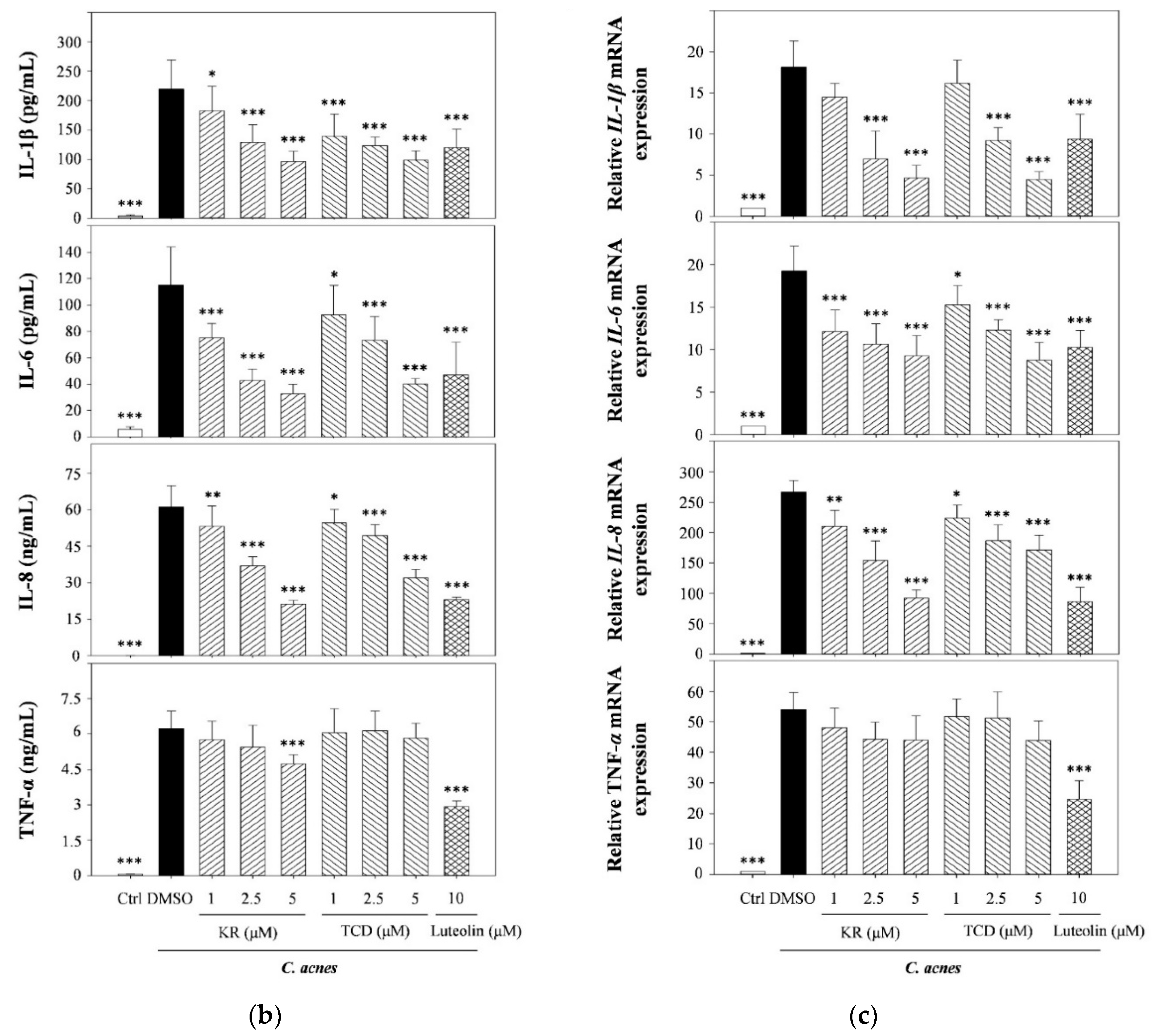
2.1. Effects of KR and TCD on C. acnes-Induced Cellular Cytokine Production and mRNA Expression
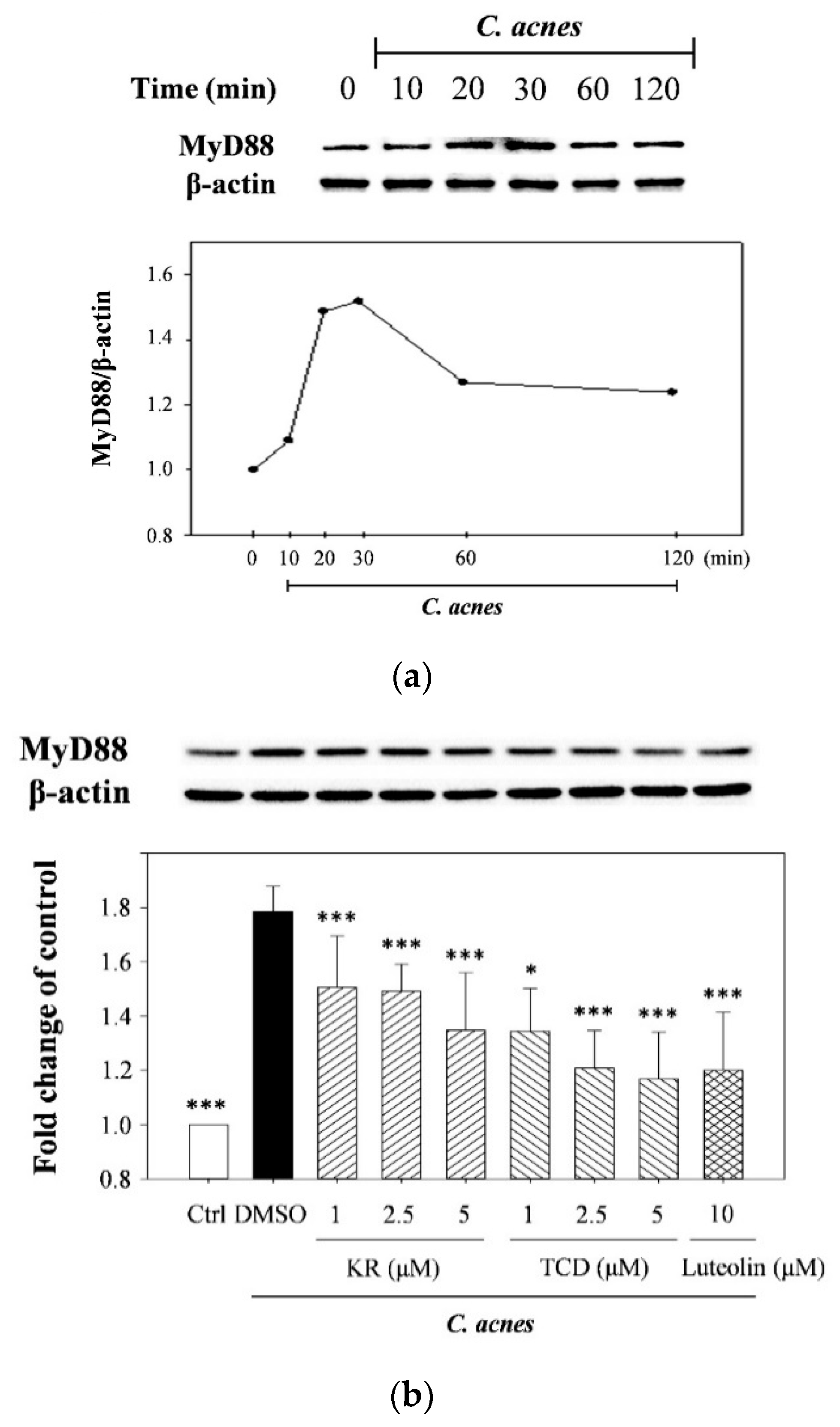
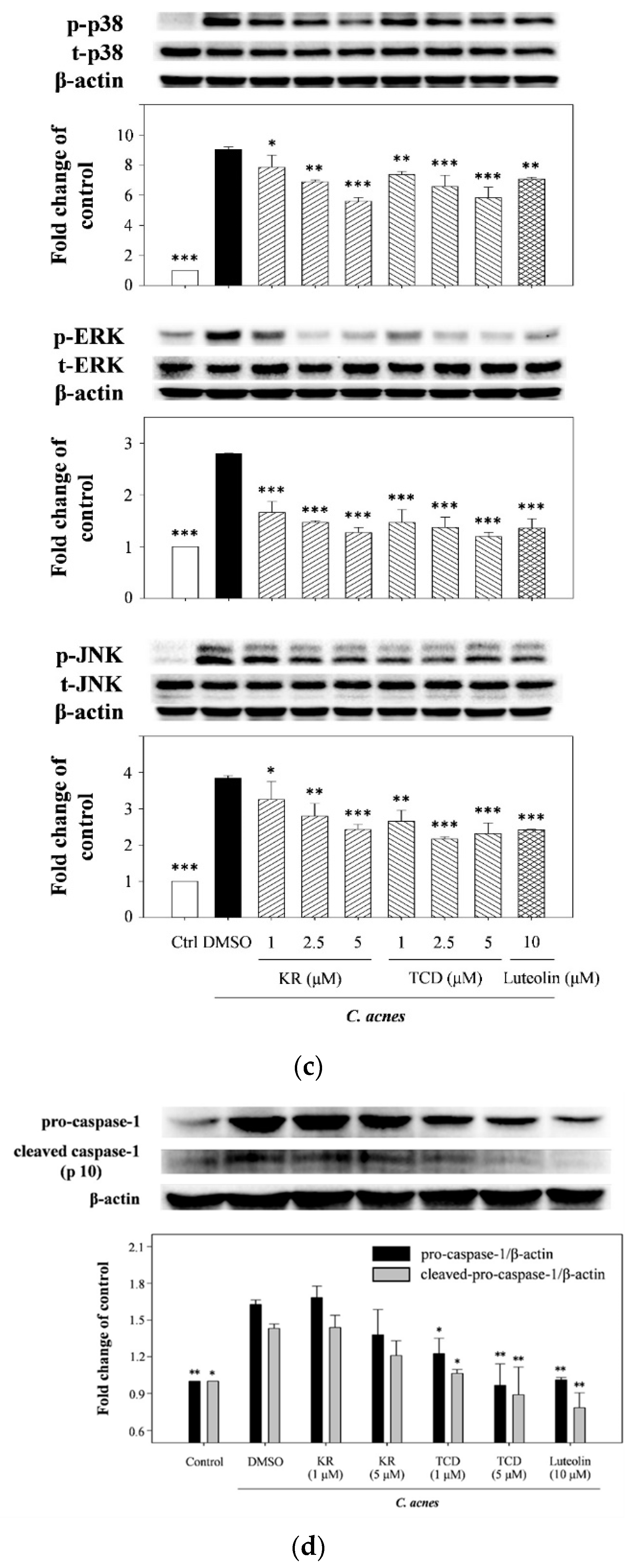
2.2. Effects of KR and TCD on Activation of MyD88, MAPK, and Caspase-1
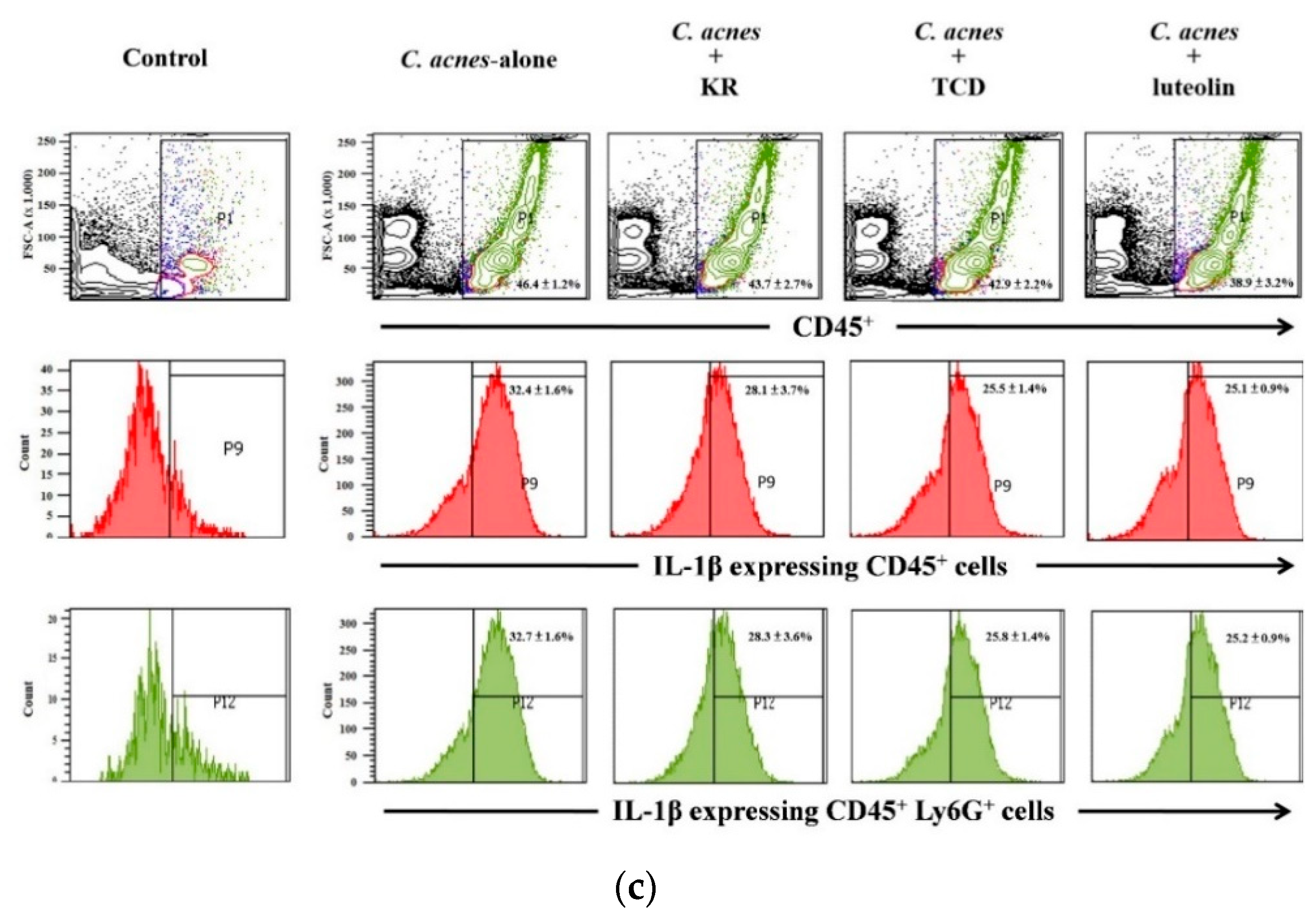
2.3. Effects of KR and TCD on C. acnes-Stimulated Mouse Ear Edema
3. Materials and Methods
3.1. Materials
3.2. Preparation of KR and TCD
3.3. Bacterial and Cell Cultures
3.4. Measurement of Cytokine Production in THP-1 Cells
3.5. Analysis of mRNA Levels by Quantitative Real-Time Reverse Transcription Polymerase Chain Reaction (PCR)
3.6. Western Blot Analysis
3.7. C. acnes-Induced Inflammation in Mouse Ears
3.8. Statistical Analysis
Author Contributions
Funding
Institutional Review Board Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
Sample Availability
References
- Tanghetti, E.A. The role of inflammation in the pathology of acne. J. Clin. Aesthet. Dermatol. 2013, 6, 27–35. [Google Scholar]
- Knor, T. The pathogenesis of acne. Acta Dermatovenerol. Croat. 2005, 13, 44–49. [Google Scholar]
- Kumar, B.; Pathak, R.; Mary, P.B.; Jha, D.; Sardana, K.; Gautam, H.K. New insights into acne pathogenesis: Exploring the role of acne-associated microbial populations. Dermatol. Sin. 2016, 34, 67–73. [Google Scholar]
- Vowels, B.R.; Yang, S.; Leyden, J.J. Induction of proinflammatory cytokines by a soluble factor of Propionibacterium acnes: Implications for chronic inflammatory acne. Infect. Immun. 1995, 63, 3158–3165. [Google Scholar]
- Nagarani, G.; Abirami, A.; Siddhuraju, P. Food prospects and nutraceutical attributes of Momordica species: A potential tropical bioresources—A review. Food Sci. Hum. Well. 2014, 3, 117–126. [Google Scholar]
- Tsai, T.H.; Huang, W.C.; Ying, H.T.; Kuo, Y.H.; Shen, C.C.; Lin, Y.K.; Tsai, P.J. Wild bitter melon leaf extract inhibits Porphyromonas gingivalis-induced inflammation: Identification of active compounds through bioassay-guided isolation. Molecules 2016, 21, 454. [Google Scholar] [PubMed] [Green Version]
- Bai, L.Y.; Chiu, C.F.; Chu, P.C.; Lin, W.Y.; Chiu, S.J.; Weng, J.R. A triterpenoid from wild bitter gourd inhibits breast cancer cells. Sci. Rep. 2016, 6, 22419. [Google Scholar] [PubMed] [Green Version]
- Seelinger, G.; Merfort, I.; Schempp, C.M. Anti-oxidant, anti-inflammatory and anti-allergic activities of luteolin. Planta Med. 2008, 74, 1667–1677. [Google Scholar]
- Gendrisch, F.; Esser, P.R.; Schempp, C.M.; Wölfle, U. Luteolin as a modulator of skin aging and inflammation. BioFactors 2021. [Google Scholar] [CrossRef]
- Lee, W.R.; Kim, K.H.; An, H.J.; Kim, J.Y.; Chang, Y.C.; Chung, H.; Park, Y.Y.; Lee, M.L.; Park, K.K. The protective effects of melittin on Propionibacterium acnes-induced inflammatory responses in vitro and in vivo. J. Investig. Dermatol. 2014, 134, 1922–1930. [Google Scholar]
- Zhang, B.; Choi, Y.M.; Lee, J.; An, I.L.; He, C.; Dong, Y.; Bae, S.; Meng, H. Toll-like receptor 2 plays a critical role in pathogenesis of acne vulgaris. Biomed. Dermatol. 2019, 3, 4. [Google Scholar]
- Krumm, B.; Xiang, Y.; Deng, J. Structural biology of the IL-1 superfamily: Key cytokines in the regulation of immune and inflammatory responses. Protein Sci. 2014, 23, 526–538. [Google Scholar] [PubMed] [Green Version]
- McLaughlin, J.; Watterson, S.; Layton, A.M.; Bjourson, A.J.; Barnard, E.; McDowell, A. Propionibacterium acnes and acne vulgaris: New insights from the integration of population genetic, multi-omic, biochemical and host-microbe studies. Microorganisms 2019, 7, 128. [Google Scholar]
- Deguine, J.; Barton, G.M. MyD88: A central player in innate immune signaling. F1000prime Rep. 2014, 6, 97. [Google Scholar]
- Huang, W.C.; Tsai, T.H.; Huang, C.J.; Li, Y.Y.; Chyuan, J.H.; Chuang, L.T.; Tsai, P.J. Inhibitory effects of wild bitter melon leaf extract on Propionibacterium acnes-induced skin inflammation in mice and cytokine production in vitro. Food Funct. 2015, 6, 2550–2560. [Google Scholar]
- Chuang, L.T.; Shih, Y.H.; Huang, W.C.; Lin, L.C.; Hsu, C.; Chyuan, J.H.; Tsai, T.H.; Tsai, P.J. In vitro and in vivo screening of wild bitter melon leaf for anti-inflammatory activity against Cutibacterium acnes. Molecules 2020, 25, 4277. [Google Scholar]
- Tsai, T.H.; Huang, W.C.; Lien, T.J.; Huang, Y.H.; Chang, H.; Yu, C.H.; Tsai, P.J. Clove extract and eugenol suppress inflammatory responses elicited by Propionibacterium acnes in vitro and in vivo. Food Agric. Immunol. 2017, 28, 916–931. [Google Scholar] [CrossRef] [Green Version]
- Rosales, C. Neutrophil: A cell with many roles in inflammation or several cell types? Front. Physiol. 2018, 9, 113. [Google Scholar]
- Sahdo, B.; Särndahl, E.; Elgh, F.; Söderquist, B. Propionibacterium acnes activates caspase-1 in human neutrophils. APMIS 2013, 121, 652–663. [Google Scholar]
- Li, Z.J.; Choi, D.K.; Sohn, K.C.; Seo, M.S.; Lee, H.E.; Lee, Y.; Seo, Y.J.; Lee, Y.H.; Shi, G.; Zouboulis, C.C.; et al. Propionibacterium acnes activates the NLRP3 inflammasome in human sebocytes. J. Investig. Dermatol. 2014, 134, 2747–2756. [Google Scholar]
- Qin, M.; Pirouz, A.; Kim, M.H.; Krutzik, S.R.; Garbán, H.J.; Kim, J. Propionibacterium acnes induces IL-1β secretion via the NLRP3 inflammasome in human monocytes. J. Investig. Dermatol. 2014, 134, 381–388. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rodriguez, A.E.; Bogart, C.; Gilbert, C.M.; McCullers, J.A.; Smith, A.M.; Kanneganti, T.D.; Lupfer, C.R. Enhanced IL-1β production is mediated by a TLR2-MYD88-NLRP3 signaling axis during coinfection with influenza A virus and Streptococcus pneumoniae. PLoS ONE 2019, 14, e0212236. [Google Scholar] [CrossRef] [PubMed]
- Chuang, L.T.; Tsai, T.H.; Lien, T.J.; Huang, W.C.; Liu, J.J.; Chang, H.; Chang, M.L.; Tsai, P.J. Ethanolic extract of Origanum vulgare suppresses Propionibacterium acnes-induced inflammatory responses in human monocyte and mouse ear edema models. Molecules 2018, 23, 1987. [Google Scholar] [CrossRef] [PubMed] [Green Version]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Protein | Clone | Source/Company (Catalog Number) | Assay |
|---|---|---|---|
| MyD88 | D80F5 | Cell Signaling Technology (4283) | WB |
| p38 | Polyclonal | Cell Signaling Technology (9212) | WB |
| ERK | 137F5 | Cell Signaling Technology (4695) | WB |
| JNK | 56G8 | Cell Signaling Technology (9258) | WB |
| p-p38 | D3F9 | Cell Signaling Technology (4511) | WB |
| p-ERK | 20G11 | Cell Signaling Technology (4376) | WB |
| p-JNK | 81E11 | Cell Signaling Technology (4668) | WB |
| pro-caspase-1 | EPR4321 | GeneTex (GTX62815) | WB |
| cleaved caspase-1 | Polyclonal | GeneTex (GTX134551) | WB |
| β-actin | AC-74 | Sigma (A5316) | WB |
| PerCP-anti-mouse CD45 1 | 30-F11 | BioLegend (103129) | Flow |
| FITC-anti-mouse Ly6G 2 | 1A8 | BioLegend (127605) | Flow |
| PE-anti-mouse F4/80 3 | 30-F11 | BioLegend (127605) | Flow |
| APC-anti-mouse-IL-1β 4 | NJTEN3 | BioLegend (127605) | Flow |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Chuang, L.-T.; Huang, W.-C.; Hou, Y.-C.; Chyuan, J.-H.; Chang, H.; Chang, C.-I.; Tsai, T.-H.; Tsai, P.-J. Suppressive Effect of Two Cucurbitane-Type Triterpenoids from Momordica charantia on Cutibacterium acnes-Induced Inflammatory Responses in Human THP-1 Monocytic Cell and Mouse Models. Molecules 2021, 26, 579. https://doi.org/10.3390/molecules26030579
Chuang L-T, Huang W-C, Hou Y-C, Chyuan J-H, Chang H, Chang C-I, Tsai T-H, Tsai P-J. Suppressive Effect of Two Cucurbitane-Type Triterpenoids from Momordica charantia on Cutibacterium acnes-Induced Inflammatory Responses in Human THP-1 Monocytic Cell and Mouse Models. Molecules. 2021; 26(3):579. https://doi.org/10.3390/molecules26030579
Chicago/Turabian StyleChuang, Lu-Te, Wen-Cheng Huang, Yu-Chen Hou, Jong-Ho Chyuan, Hsiang Chang, Chi-I Chang, Tsung-Hsien Tsai, and Po-Jung Tsai. 2021. "Suppressive Effect of Two Cucurbitane-Type Triterpenoids from Momordica charantia on Cutibacterium acnes-Induced Inflammatory Responses in Human THP-1 Monocytic Cell and Mouse Models" Molecules 26, no. 3: 579. https://doi.org/10.3390/molecules26030579