
Alginic Acid from Padina boryana Abate Particulate Matter-Induced Inflammatory Responses in Keratinocytes and Dermal Fibroblasts

 ,
,  ,
,  ,
,
Abstract
:1. Introduction
2. Results
2.1. Proximate Composition and Chemical Composition
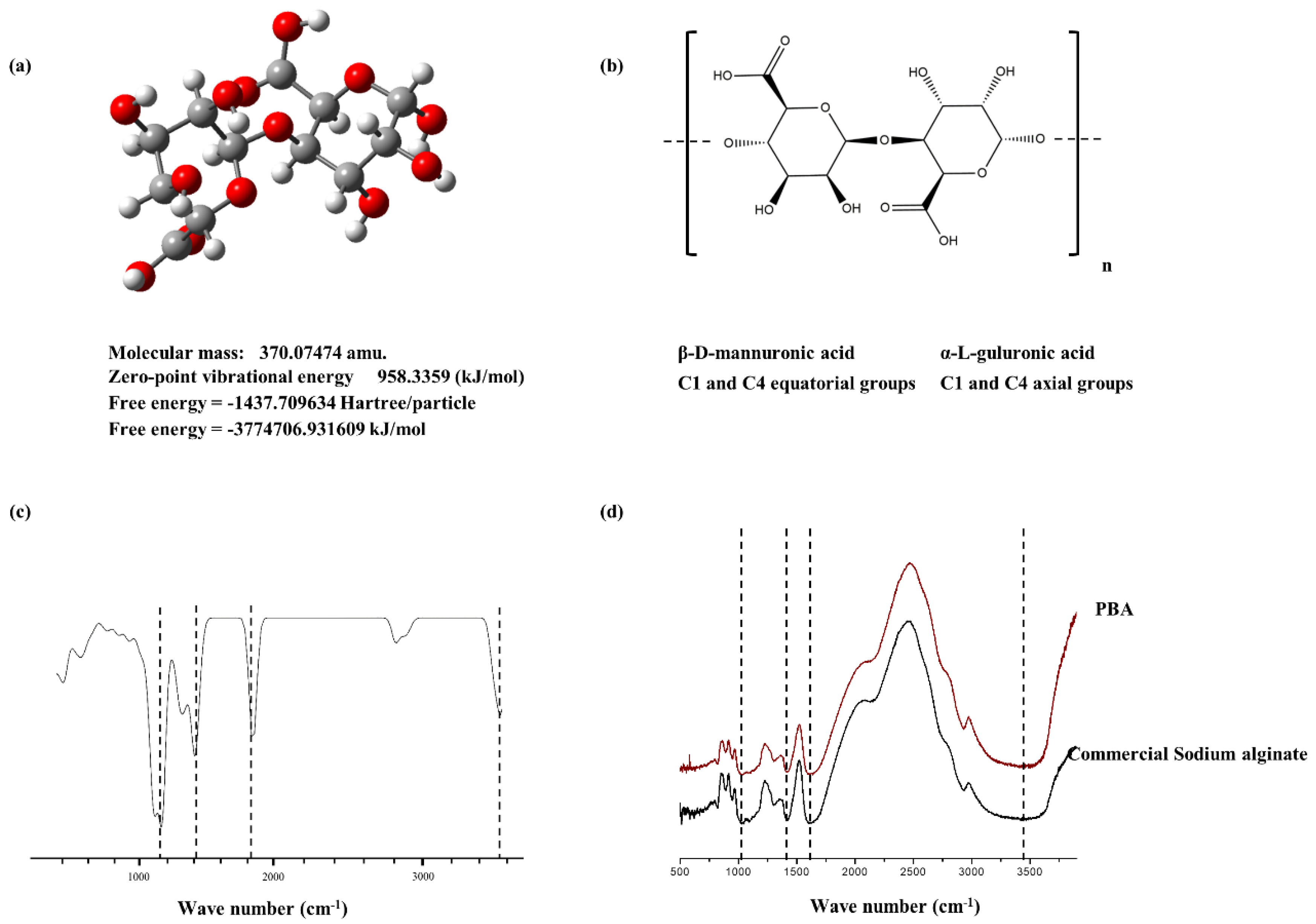
2.2. Structural Characterization of PBA
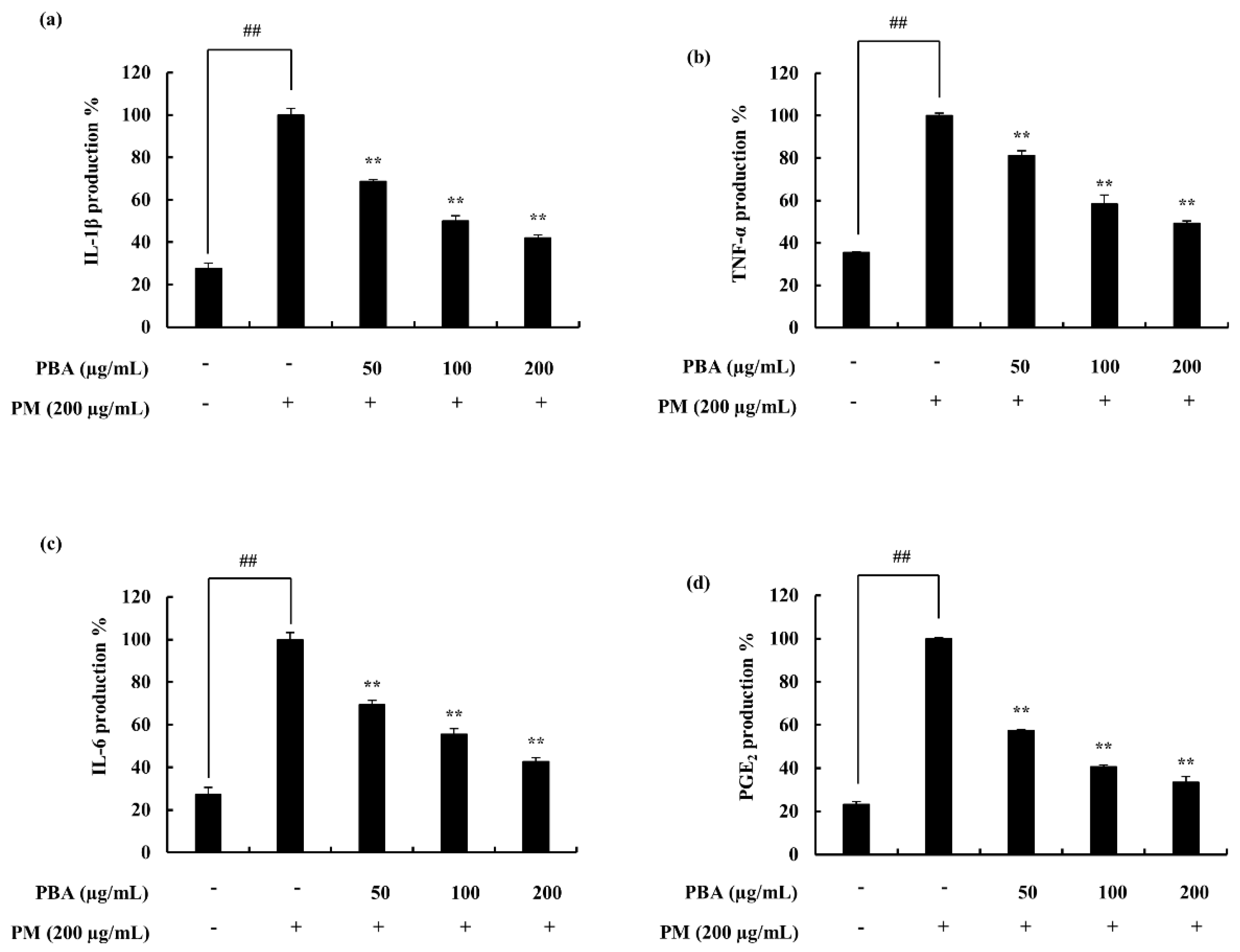
2.3. Potential of PBA to Reduce PM-Stimulated Inflammatory Responses in Keratinocytes and Fibroblasts
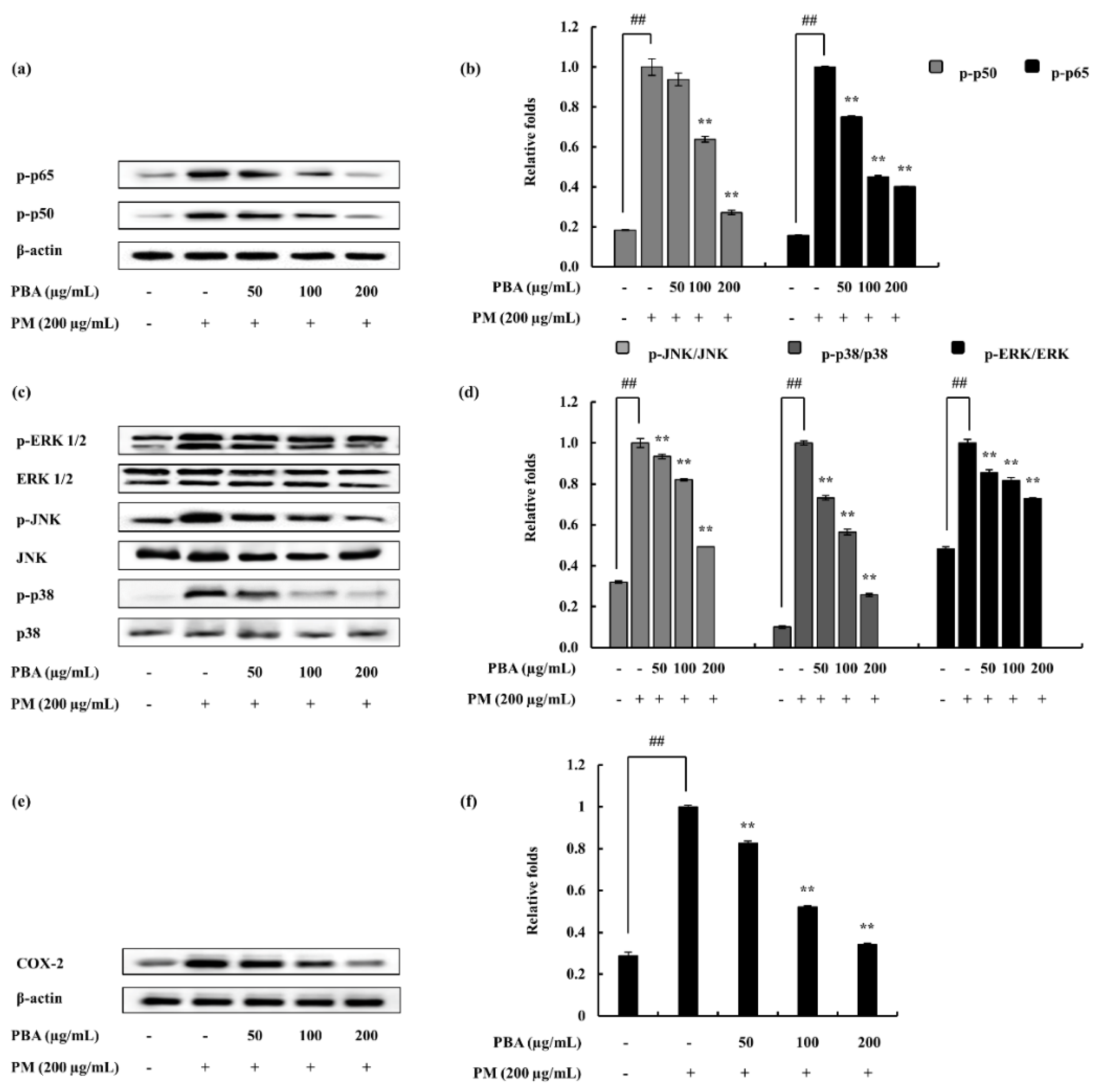
2.4. Potential of PBA to Abate PM-Induced Inflammatory Responses via NF-κB and MAPK Pathways in Keratinocytes
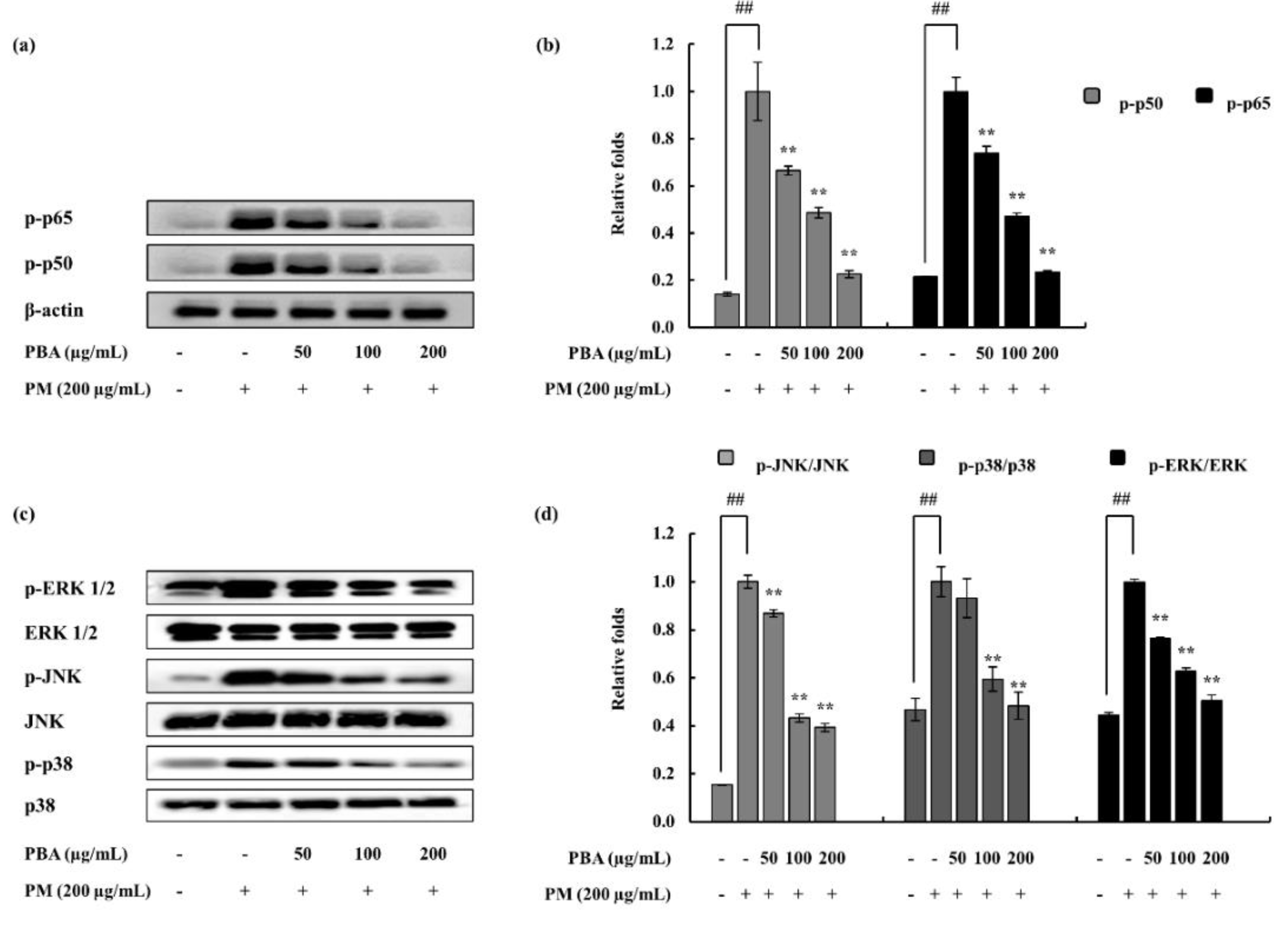
2.5. Effect of PBA on PM-Induced NF-κB and MAPK Proteins in Fibroblasts
2.6. Keratinocytes Stimulated with PM and Treated with PBA; Compositional Analysis
3. Discussion
4. Materials and Methods
4.1. Materials
4.2. Alginic Acid Purification from P. boryana
4.3. Analysis of the Proximate Composition of P. boryana
4.4. Evaluation of Chemical Composition of PBA
4.5. Functional Group Analysis of PBA Using FTIR
4.6. FTIR Spectra Interpretation Using Computational Calculations
4.7. Maintenance of Cell Lines
4.8. Analysis of Cell Viability and Intracellular ROS
4.9. PGE2 and Pro-Inflammatory Cytokine Production Level Assessment
4.10. Western Blot Analysis
4.11. Spectroscopic Analysis (ICP-OES)
4.12. Statistical Analysis
5. Conclusions
Author Contributions
Funding
Conflicts of Interest
References
- Turnbull, A.B.; Harrison, R.M. Major component contributions to PM10 composition in the UK atmosphere. Atmos. Environ. 2000, 34, 3129–3137. [Google Scholar] [CrossRef]
- Fernando, I.P.S.; Kim, H.-S.; Sanjeewa, K.K.A.; Oh, J.-Y.; Jeon, Y.-J.; Lee, W.W. Inhibition of inflammatory responses elicited by urban fine dust particles in keratinocytes and macrophages by diphlorethohydroxycarmalol isolated from a brown alga Ishige okamurae. Algae 2017, 32, 261–273. [Google Scholar] [CrossRef] [Green Version]
- Jayawardena, T.U.; Asanka Sanjeewa, K.K.; Shanura Fernando, I.P.; Ryu, B.M.; Kang, M.C.; Jee, Y.; Lee, W.W.; Jeon, Y.J. Sargassum horneri (Turner) C. Agardh ethanol extract inhibits the fine dust inflammation response via activating Nrf2/HO-1 signaling in RAW 264.7 cells. BMC Complement. Altern. Med. 2018, 18, 249. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gilmour, P.S.; Brown, D.M.; Lindsay, T.G.; Beswick, P.H.; MacNee, W.; Donaldson, K. Adverse health effects of PM10 particles: Involvement of iron in generation of hydroxyl radical. Occup. Environ. Med. 1996, 53, 817–822. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Knaapen, A.M.; Shi, T.; Borm, P.J.A.; Schins, R.P.F. Soluble metals as well as the insoluble particle fraction are involved in cellular DNA damage induced by particulate matter. In Oxygen/Nitrogen Radicals: Cell Injury and Disease; Vallyathan, V., Shi, X., Castranova, V., Eds.; Springer US: Boston, MA, USA, 2002; pp. 317–326. [Google Scholar] [CrossRef]
- Brown, D.M.; Wilson, M.R.; MacNee, W.; Stone, V.; Donaldson, K. Size-dependent proinflammatory effects of ultrafine polystyrene particles: A role for surface area and oxidative stress in the enhanced activity of ultrafines. Toxicol. Appl. Pharmacol. 2001, 175, 191–199. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Becker, S.; Soukup, J.M.; Gilmour, M.I.; Devlin, R.B. Stimulation of human and rat alveolar macrophages by urban air particulates: Effects on oxidant radical generation and cytokine production. Toxicol. Appl. Pharmacol. 1996, 141, 637–648. [Google Scholar] [CrossRef]
- Schins, R.P.; Lightbody, J.H.; Borm, P.J.; Shi, T.; Donaldson, K.; Stone, V. Inflammatory effects of coarse and fine particulate matter in relation to chemical and biological constituents. Toxicol. Appl. Pharmacol. 2004, 195, 1–11. [Google Scholar] [CrossRef]
- Williams, I.R.; Kupper, T.S. Immunity at the surface: Homeostatic mechanisms of the skin immune system. Life Sci. 1996, 58, 1485–1507. [Google Scholar] [CrossRef]
- Werner, S.; Peters, K.G.; Longaker, M.T.; Fuller-Pace, F.; Banda, M.J.; Williams, L.T. Large induction of keratinocyte growth factor expression in the dermis during wound healing. Proc. Natl. Acad. Sci. USA 1992, 89, 6896–6900. [Google Scholar] [CrossRef] [Green Version]
- Fernando, I.P.S.; Sanjeewa, K.K.A.; Kim, S.Y.; Lee, J.S.; Jeon, Y.J. Reduction of heavy metal (Pb2+) biosorption in zebrafish model using alginic acid purified from Ecklonia cava and two of its synthetic derivatives. Int. J. Biol. Macromol. 2018, 106, 330–337. [Google Scholar] [CrossRef]
- Sarithakumari, C.H.; Renju, G.L.; Kurup, G.M. Anti-inflammatory and antioxidant potential of alginic acid isolated from the marine algae, Sargassum wightii on adjuvant-induced arthritic rats. Inflammopharmacology 2013, 21, 261–268. [Google Scholar] [CrossRef] [PubMed]
- Fertah, M.; Belfkira, A.; Dahmane, E.M.; Taourirte, M.; Brouillette, F. Extraction and characterization of sodium alginate from Moroccan Laminaria digitata brown seaweed. Arab. J. Chem. 2017, 10, S3707–S3714. [Google Scholar] [CrossRef] [Green Version]
- Kim, K.E.; Cho, D.; Park, H.J. Air pollution and skin diseases: Adverse effects of airborne particulate matter on various skin diseases. Life Sci. 2016, 152, 126–134. [Google Scholar] [CrossRef] [PubMed]
- Hahn, T.; Lang, S.; Ulber, R.; Muffler, K. Novel procedures for the extraction of fucoidan from brown algae. Process. Biochem. 2012, 47, 1691–1698. [Google Scholar] [CrossRef]
- McHugh, D.J. Production, properties and uses of alginates. Prod. Util. Prod. Commer. Seaweeds. FAO Fish. Tech. Pap. 1987, 288, 58–115. [Google Scholar]
- Smidsrod, O.; Haug, A. Dependence upon the gel-sol state of the ion-exchange properties of alginates. Acta Chem. Scand. 1972, 26, 2063–2074. [Google Scholar] [CrossRef]
- Penman, A.; Sanderson, G.R. A method for the determination of uronic acid sequence in alginates. Carbohydr. Res. 1972, 25, 273–282. [Google Scholar] [CrossRef]
- Haug, A.; Larsen, B.; Smidsrod, O. Uronic Acid Sequence in Alginate from Different Sources. Carbohydr. Res. 1974, 32, 217–225. [Google Scholar] [CrossRef]
- Donati, I.; Vetere, A.; Gamini, A.; Skjak-Braek, G.; Coslovi, A.; Campa, C.; Paoletti, S. Galactose-substituted alginate: Preliminary characterization and study of gelling properties. Biomacromolecules 2003, 4, 624–631. [Google Scholar] [CrossRef]
- Papageorgiou, S.K.; Kouvelos, E.P.; Favvas, E.P.; Sapalidis, A.A.; Romanos, G.E.; Katsaros, F.K. Metal-carboxylate interactions in metal-alginate complexes studied with FTIR spectroscopy. Carbohydr. Res. 2010, 345, 469–473. [Google Scholar] [CrossRef]
- Schaumann, F.; Borm, P.J.; Herbrich, A.; Knoch, J.; Pitz, M.; Schins, R.P.; Luettig, B.; Hohlfeld, J.M.; Heinrich, J.; Krug, N. Metal-rich ambient particles (particulate matter 2.5) cause airway inflammation in healthy subjects. Am. J. Respir. Crit. Care Med. 2004, 170, 898–903. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Fu, F.; Wang, Q. Removal of heavy metal ions from wastewaters: A review. J. Environ. Manag. 2011, 92, 407–418. [Google Scholar] [CrossRef] [PubMed]
- Blue, L.Y.; Van Aelstyn, M.A.; Matlock, M.; Atwood, D.A. Low-level mercury removal from groundwater using a synthetic chelating ligand. Water Res. 2008, 42, 2025–2028. [Google Scholar] [CrossRef] [PubMed]
- King, R.B. Coordination number, electronic configuration, and ionic charge as discrete variables in coordination chemistry. Adv. Chem. Ser. 1967, 62, 203–220. [Google Scholar] [CrossRef]
- Jeon, C.; Park, J.Y.; Yoo, Y.J. Novel immobilization of alginic acid for heavy metal removal. Biochem. Eng. J. 2002, 11, 159–166. [Google Scholar] [CrossRef]
- Grant, G.T.; Morris, E.R.; Rees, D.A.; Smith, P.J.C.; Thom, D. Biological interactions between polysaccharides and divalent cations: The egg-box model. FEBS Lett. 1973, 32, 195–198. [Google Scholar] [CrossRef] [Green Version]
- Fernando, I.P.S.; Jayawardena, T.U.; Sanjeewa, K.K.A.; Wang, L.; Jeon, Y.J.; Lee, W.W. Anti-inflammatory potential of alginic acid from Sargassum horneri against urban aerosol-induced inflammatory responses in keratinocytes and macrophages. Ecotoxicol. Environ. Saf. 2018, 160, 24–31. [Google Scholar] [CrossRef]
- Kim, B.H.; Oh, I.; Kim, J.H.; Jeon, J.E.; Jeon, B.; Shin, J.; Kim, T.Y. Anti-inflammatory activity of compounds isolated from Astragalus sinicus L. in cytokine-induced keratinocytes and skin. Exp. Mol. Med. 2014, 46, e87. [Google Scholar] [CrossRef]
- Karin, M.; Delhase, M. The I kappa B kinase (IKK) and NF-kappa B: Key elements of proinflammatory signalling. Semin. Immunol. 2000, 12, 85–98. [Google Scholar] [CrossRef]
- Sanjeewa, K.K.A.; Jayawardena, T.U.; Kim, H.-S.; Kim, S.-Y.; Ahn, G.; Kim, H.-J.; Fu, X.; Jee, Y.; Jeon, Y.-J. Ethanol extract separated from Sargassum horneri (Turner) abate LPS-induced inflammation in RAW 264.7 macrophages. Fish. Aquat. Sci. 2019, 22, 6. [Google Scholar] [CrossRef] [Green Version]
- Akira, S. Toll-like Receptors and Innate Immunity. In Advances in Immunology; Dixon, F.J., Ed.; Academic Press: Cambridge, MA, USA, 2001; Volume 78, pp. 1–56. [Google Scholar]
- Kaminska, B. MAPK signalling pathways as molecular targets for anti-inflammatory therapy—From molecular mechanisms to therapeutic benefits. Biochim. Biophys. Acta (BBA) Proteins Proteom. 2005, 1754, 253–262. [Google Scholar] [CrossRef] [PubMed]
- Dubois, M.; Gilles, K.A.; Hamilton, J.K.; Rebers, P.T.; Smith, F. Colorimetric method for determination of sugars and related substances. Anal. Chem. 1956, 28, 350–356. [Google Scholar] [CrossRef]
- Chandler, S.F.; Dodds, J.H. The effect of phosphate, nitrogen and sucrose on the production of phenolics and solasodine in callus cultures of solanum laciniatum. Plant Cell Rep. 1983, 2, 205–208. [Google Scholar] [CrossRef] [PubMed]
- Cardenas-Jiron, G.; Leal, D.; Matsuhiro, B.; Osorio-Roman, I.O. Vibrational spectroscopy and density functional theory calculations of poly-D-mannuronate and heteropolymeric fractions from sodium alginate. J. Raman Spectrosc. 2011, 42, 870–878. [Google Scholar] [CrossRef]
- Mosmann, T. Rapid colorimetric assay for cellular growth and survival: Application to proliferation and cytotoxicity assays. J. Immunol. Methods 1983, 65, 55–63. [Google Scholar] [CrossRef]
- Wang, L.; Ryu, B.; Kim, W.S.; Kim, G.H.; Jeon, Y.J. Protective effect of gallic acid derivatives from the freshwater green alga Spirogyra sp against ultraviolet B-induced apoptosis through reactive oxygen species clearance in human keratinocytes and zebrafish. Algae 2017, 32, 379–388. [Google Scholar] [CrossRef] [Green Version]
- Jayawardena, T.U.; Kim, H.-S.; Sanjeewa, K.K.A.; Kim, S.-Y.; Rho, J.-R.; Jee, Y.; Ahn, G.; Jeon, Y.-J. Sargassum horneri and isolated 6-hydroxy-4,4,7a-trimethyl-5,6,7,7a-tetrahydrobenzofuran-2(4H)-one (HTT); LPS-induced inflammation attenuation via suppressing NF-κB, MAPK and oxidative stress through Nrf2/HO-1 pathways in RAW 264.7 macrophages. Algal Res. 2019, 40, 101513. [Google Scholar] [CrossRef]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Composition | Content (%) |
|---|---|
| Polysaccharide | 79.84 ± 1.32 |
| Ash | 3.42 ± 0.56 |
| Protein | 1.22 ± 0.18 |
| Polyphenol | 2.17 ± 0.69 |
| Yield | 16.85 ± 0.32 |
| Control | PM | PM + PBA (50) | PM + PBA (100) | PM + PBA (200) | |
|---|---|---|---|---|---|
| Mg | 54.67 ± 1.47 | 128.53 ± 2.51 | 137.44 ± 4.97 | 101.46 ± 6.14 | 84.95 ± 2.11 |
| Al | 0.91 | 125.08 ± 9.1 | 119.33 ± 8.21 | 79.31 ± 5.12 | 56.92 ± 3.52 |
| K | 452.98 ± 14.56 | 437.37 ± 18.47 | 522.65 ± 20.22 | 504.28 ± 31.58 | 466.12 ± 19.68 |
| Ca | 224.44 ± 12.56 | 344.37 ± 10.45 | 385.24 ± 9.56 | 321.07 ± 5.36 | 256.44 ± 7.48 |
| Fe | ND | 174.50 ± 4.57 | 163.62 ± 6.89 | 106.52 ± 7.58 | 77.27 ± 5.23 |
| Mn | ND | 80.2 ± 2.47 | 68.4 ± 3.93 | 45.2 ± 2.42 | 32.1 ± 2.9 |
| Cu | ND | 20.1 ± 1.44 | 10.84 ± 1.89 | 6.53 ± 2.01 | ND |
| Sr | 0.71 | 42.18 ± 1.32 | 31.94 ± 2.58 | 21.37 ± 1.22 | 11.86 ± 1.09 |
| Ba | ND | 92.92 ± 6.33 | 72.56 ± 4.39 | 51.99 ± 3.15 | 20.41 ± 1.02 |
| Pb | ND | 241.74 ± 12.46 | 168.41 ± 14.15 | 102.47 ± 6.27 | 58.96 ± 3.68 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Jayawardena, T.U.; Sanjeewa, K.K.A.; Wang, L.; Kim, W.-S.; Lee, T.-K.; Kim, Y.-T.; Jeon, Y.-J. Alginic Acid from Padina boryana Abate Particulate Matter-Induced Inflammatory Responses in Keratinocytes and Dermal Fibroblasts. Molecules 2020, 25, 5746. https://doi.org/10.3390/molecules25235746
Jayawardena TU, Sanjeewa KKA, Wang L, Kim W-S, Lee T-K, Kim Y-T, Jeon Y-J. Alginic Acid from Padina boryana Abate Particulate Matter-Induced Inflammatory Responses in Keratinocytes and Dermal Fibroblasts. Molecules. 2020; 25(23):5746. https://doi.org/10.3390/molecules25235746
Chicago/Turabian StyleJayawardena, Thilina U., K. K. Asanka Sanjeewa, Lei Wang, Won-Suk Kim, Tae-Ki Lee, Yong-Tae Kim, and You-Jin Jeon. 2020. "Alginic Acid from Padina boryana Abate Particulate Matter-Induced Inflammatory Responses in Keratinocytes and Dermal Fibroblasts" Molecules 25, no. 23: 5746. https://doi.org/10.3390/molecules25235746