
Antibacterial Activity of a Cationic Antimicrobial Peptide against Multidrug-Resistant Gram-Negative Clinical Isolates and Their Potential Molecular Targets

 , , ,
, , ,  , and
, and
Abstract
:1. Introduction
2. Results and Discussion
2.1. Clinical Isolates: Antibiotic Susceptibility and Resistance Genes
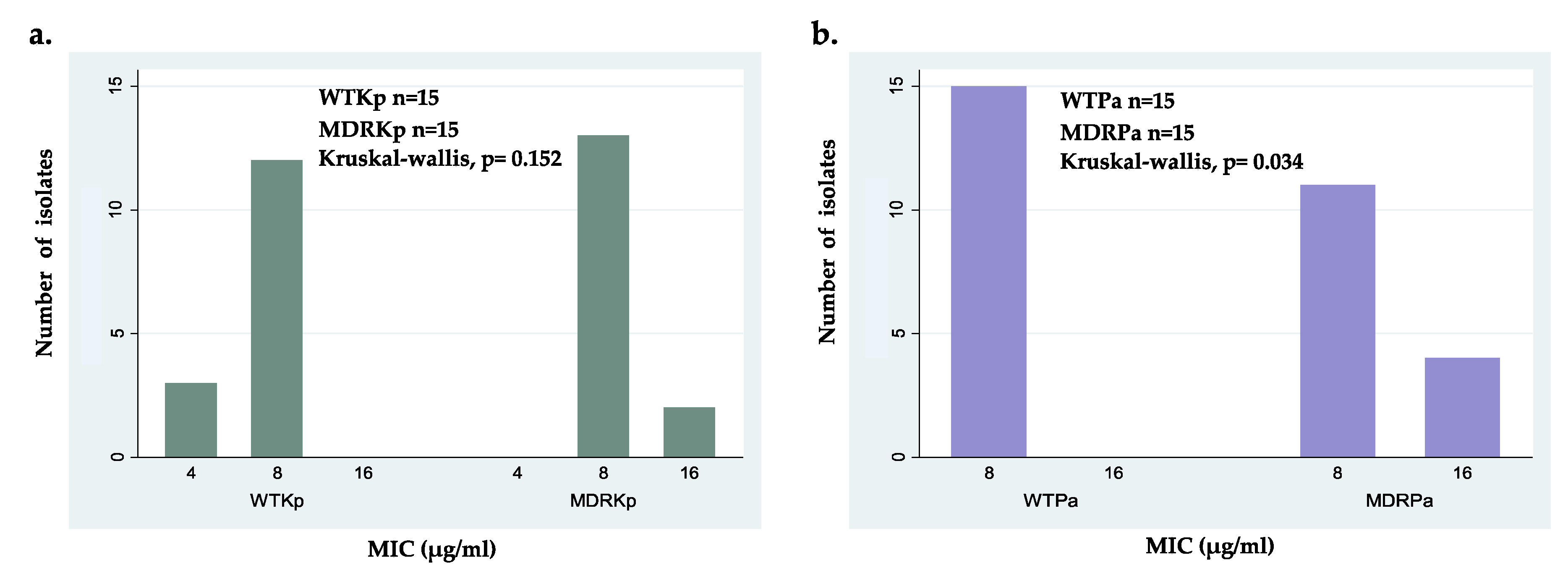
2.2. Antibacterial Activity of ΔM2 against Clinical Isolates
2.3. Interaction of ΔM2 with Potential Bacterial Targets
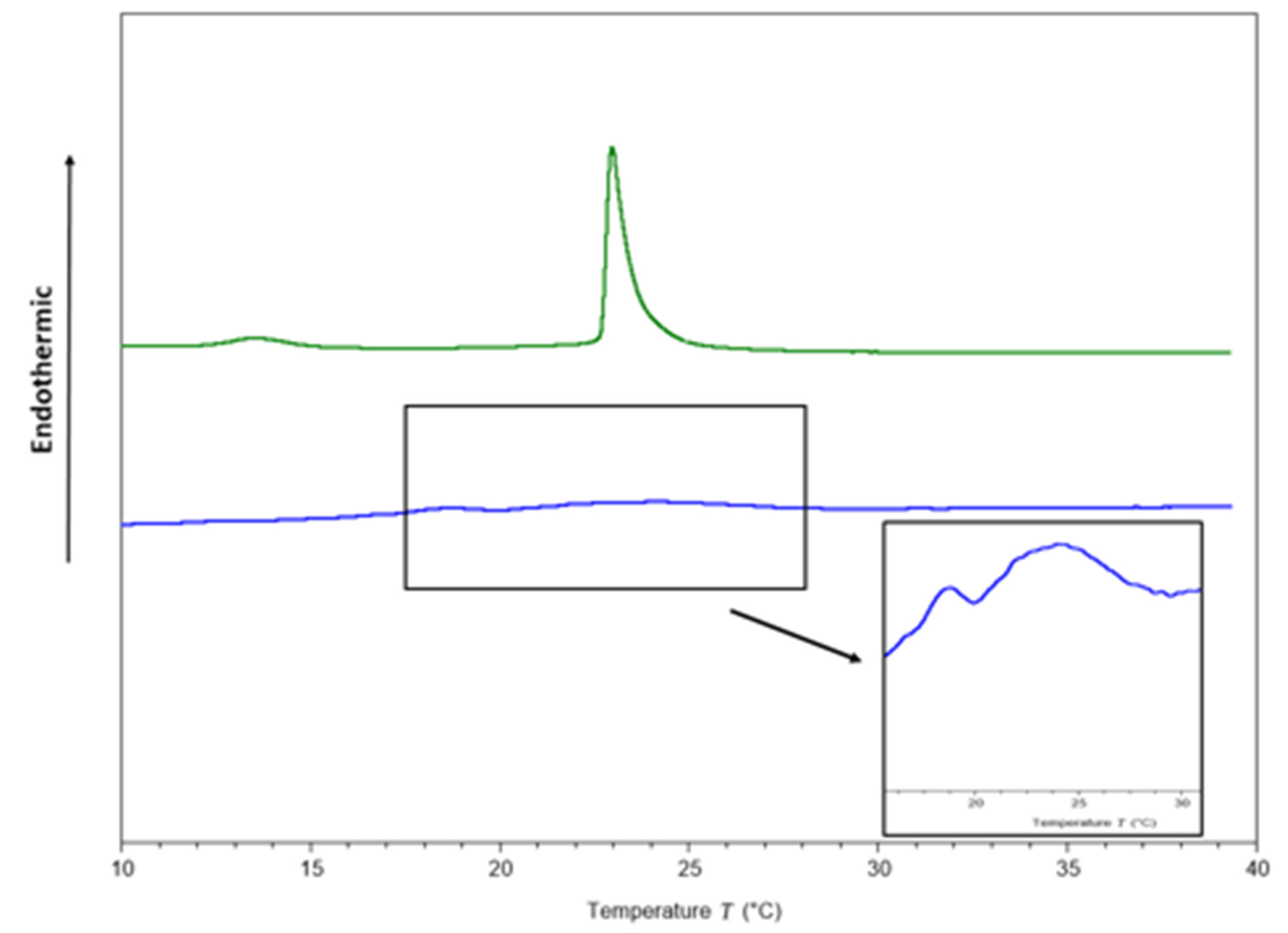
2.3.1. Interaction of ΔM2 with Model Bacterial Membrane
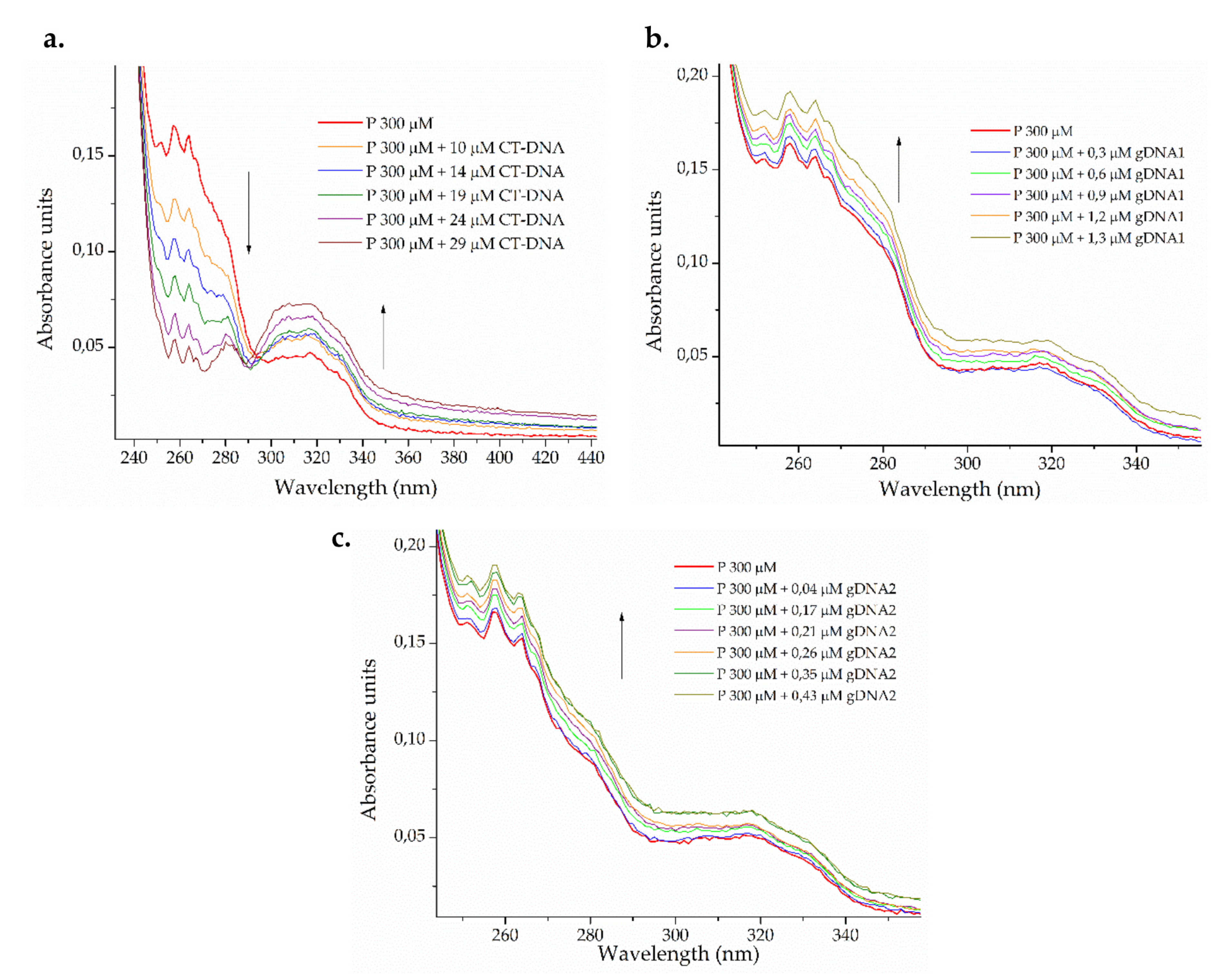
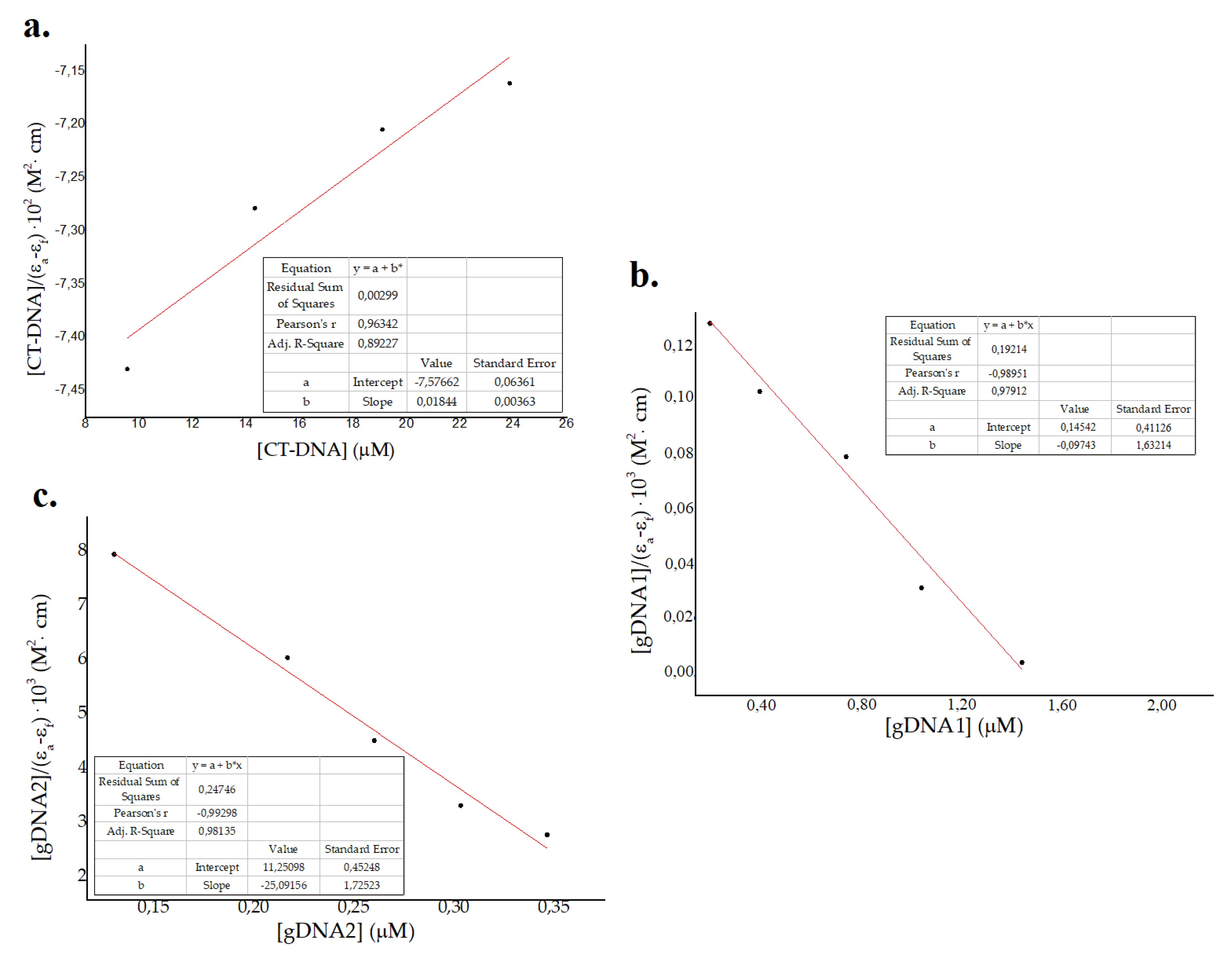
2.3.2. Interaction of ΔM2 with Bacterial DNA
Electronic Absorption Spectra
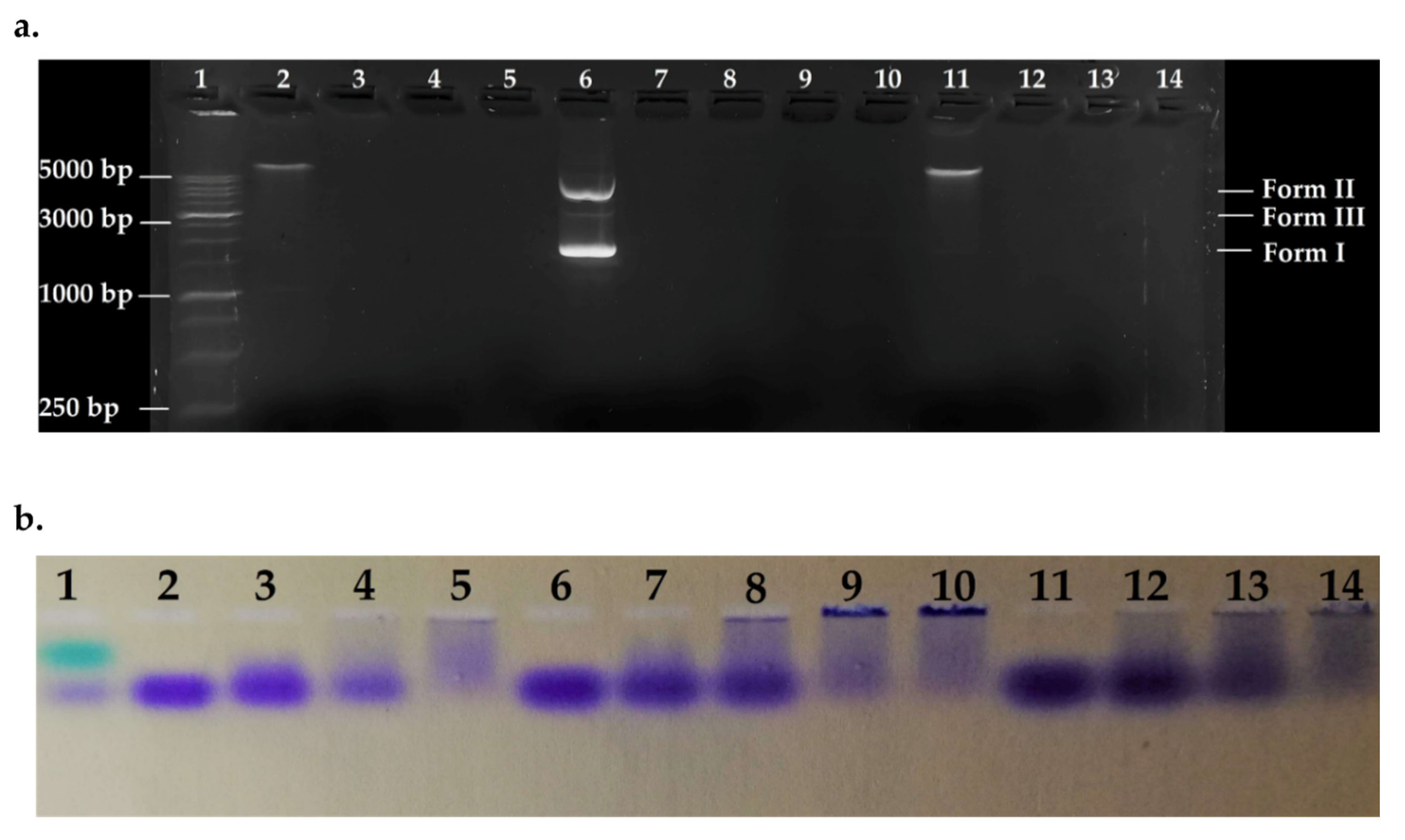
Agarose Gel Electrophoresis
3. Materials and Methods
3.1. Peptide Design and Synthesis
3.2. Identification of Clinical Isolates
3.3. Characterization of Clinical Isolates: Resistance Profile and Identification of Resistance Genes
3.4. Antimicrobial Assay of ΔM2
3.5. Interaction of Model Membranes
3.5.1. Membrane Preparation
3.5.2. Differential Scanning Calorimetry
3.6. DNA Interaction Assays
3.6.1. Electronic Absorption Monitoring Assays (UV-Vis)
3.6.2. Agarose Gel Electrophoresis
3.7. Statistical Analysis
4. Conclusions
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- World Health Organization. Antimicrobial Resistance. Available online: https://www.who.int/news-room/fact-sheets/detail/antimicrobial-resistance (accessed on 5 August 2020).
- World Health Organization. WHO Publishes List of Bacteria for Which New Antibiotics are Urgently Needed. Available online: https://www.who.int/news/item/27-02-2017-who-publishes-list-of-bacteria-for-which-new-antibiotics-are-urgently-needed (accessed on 5 August 2020).
- Weinstein, R.A.; Gaynes, R.; Edwards, J.R. Overview of Nosocomial Infections Caused by Gram-Negative Bacilli. Clin. Infect. Dis. 2005, 41, 848–854. [Google Scholar] [CrossRef] [PubMed]
- Levy, S.B.; Bonnie, M. Antibacterial resistance worldwide: Causes, challenges and responses. Nat. Med. 2004, 10, 122–129. [Google Scholar] [CrossRef]
- Klevens, R.M.; Edwards, J.R.; Gaynes, R.P. The Impact of Antimicrobial-Resistant, Health Care–Associated Infections on Mortality in the United States. Clin. Infect. Dis. 2008, 47, 927–930. [Google Scholar] [CrossRef] [PubMed]
- Magiorakos, A.P.; Burns, K.; Rodriguez Baño, J.; Borg, M.; Daikos, G.; Dumpis, U.; Lucet, J.C.; Moro, M.L.; Tacconelli, E.; Skov Simonsen, G.; et al. Infection prevention and control measures and tools for the prevention of entry of carbapenem-resistant Enterobacteriaceae into healthcare settings: Guidance from the European Centre for Disease Prevention and Control. Antimicrob. Resist. Infect. Control. 2017, 6, 113. [Google Scholar] [CrossRef] [PubMed]
- Pang, Z.; Raudonis, R.; Glick, B.R.; Lin, T.J.; Cheng, Z. Antibiotic resistance in Pseudomonas aeruginosa: Mechanisms and alternative therapeutic strategies. Biotechnol. Adv. 2019, 37, 177–192. [Google Scholar] [CrossRef]
- Dößelmann, B.; Willmann, M.; Steglich, M.; Bunk, B.; Nübel, U.; Peter, S.; Neher, R.A. Rapid and consistent evolution of colistin resistance in extensively drug-resistant pseudomonas aeruginosa during morbidostat culture. Antimicrob. Agents Chemother. 2017, 61. [Google Scholar] [CrossRef] [Green Version]
- Matos, E.C.; Matos, H.J.; Conceição, M.L.; Rodrigues, Y.C.; Carneiro, I.C.; Lima, K.V. Clinical and microbiological features of infections caused by pseudomonas aeruginosa in patients hospitalized in intensive care units. Rev. Soc. Bras. Med. Trop. 2016, 49, 305–311. [Google Scholar] [CrossRef]
- Fair, R.J.; Tor, Y. Antibiotics and bacterial resistance in the 21st century. Perspect. Med. Chem. 2014, 25–64. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Partridge, S.R. Resistance mechanisms in Enterobacteriaceae. Pathology 2015, 47, 276–284. [Google Scholar] [CrossRef]
- Ferreira, R.L.; Da Silva, B.C.M.; Rezende, G.S.; Nakamura-Silva, R.; Pitondo-Silva, A.; Campanini, E.B.; Brito, M.C.A.; Da Silva, E.M.L.; De Melo Freire, C.C.; Da Cunha, A.F.; et al. High prevalence of multidrug-resistant klebsiella pneumoniae harboring several virulence and β-lactamase encoding genes in a brazilian intensive care unit. Front. Microbiol. 2019, 10, 3198. [Google Scholar] [CrossRef] [Green Version]
- Ocampo, A.M.; Vargas, C.A.; Sierra, P.M.; Cienfuegos, A.V.; Jiménez, J.N. Caracterización molecular de un brote de Klebsiella pneumoniae resistente a carbapenémicos en un hospital de alto nivel de complejidad de Medellín, Colombia. Biomédica 2015, 35, 496–504. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ocampo, A.; Chen, L.; Cienfuegos, A.; Roncancio, G.; Chavda, K.; Kreiswirth, B.; Jiménez, J. A Two-Year Surveillance in Five Colombian Tertiary Care Hospitals Reveals High Frequency of Non-CG258 Clones of Carbapenem-Resistant Klebsiella pneumoniae with Distinct Clinical Characteristics. Antimicrob. Agents Chemother. 2016, 60, 332–342. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Prakash, V.; Mishra, P.; Premi, H.; Walia, A.; Dhawan, S.; Kumar, A. Increasing incidence of multidrug resistant Pseudomonas aeruginosa in inpatients of a tertiary care hospital. Int. J. Res. Med. Sci. 2014, 2, 1302. [Google Scholar] [CrossRef]
- WHO & PAHO. Epidemiological Alert: Enterobacteriacea with Plasmid-Mediated Transferable Colistin Resistance, Public Health Implications in the Americas; World Health Organization and Pan American Health Organization: Washington, DC, USA, 2016. [Google Scholar]
- Correa, A.; Del Campo, R.; Perenguez, M.; Blanco, V.M.; Rodríguez-Baños, M.; Perez, F.; Maya, J.J.; Rojas, L.; Cantón, R.; Arias, C.A.; et al. Dissemination of high-risk clones of extensively drug-resistant pseudomonas aeruginosa in Colombia. Antimicrob. Agents Chemother. 2015, 59, 2421–2425. [Google Scholar] [CrossRef] [Green Version]
- Henrichfreise, B.; Wiegand, I.; Pfister, W.; Wiedemann, B. Resistance mechanisms of multiresistant Pseudomonas aeruginosa strains from Germany and correlation with hypermutation. Antimicrob. Agents Chemother. 2007, 51, 4062–4070. [Google Scholar] [CrossRef] [Green Version]
- Hirsch, E.B.; Tam, V.H. Impact of multidrug-resistant Pseudomonas aeruginosa infection on patient outcomes. Expert Rev. Pharm. Outcomes Res. 2010, 10, 441–451. [Google Scholar] [CrossRef] [Green Version]
- Magiorakos, A.P.; Srinivasan, A.; Carey, R.B.; Carmeli, Y.; Falagas, M.E.; Giske, C.G.; Harbarth, S.; Hindler, J.F.; Kahlmeter, G.; Olsson-Liljequist, B.; et al. Multidrug-resistant, extensively drug-resistant and pandrug-resistant bacteria: An international expert proposal for interim standard definitions for acquired resistance. Clin. Microbiol. Infect. 2011, 18, 268–281. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sinha, R.; Shukla, P. Antimicrobial Peptides: Recent Insights on Biotechnological Interventions and Future Perspectives. Protein Pept. Lett. 2019, 79–87. [Google Scholar] [CrossRef] [PubMed]
- Aslam, B.; Wang, W.; Arshad, M.I.; Khurshid, M.; Muzammil, S.; Rasool, M.H.; Nisar, M.A.; Alvi, R.F.; Aslam, M.A.; Qamar, M.U.; et al. Antibiotic resistance: A rundown of a global crisis. Infect. Drug Resist. 2018, 11, 1645–1658. [Google Scholar] [CrossRef] [Green Version]
- Wang, S.; Zeng, X.; Yang, Q.; Qiao, S. Antimicrobial peptides as potential alternatives to antibiotics in food animal industry. Int. J. Mol. Sci. 2016, 17, 603. [Google Scholar] [CrossRef]
- Reddy, K.V.R.; Yedery, R.D.; Aranha, C. Antimicrobial peptides: Premises and promises. Int. J. Antimicrob. Agents 2004, 24, 536–547. [Google Scholar] [CrossRef] [PubMed]
- Fry, D.E. Antimicrobial peptides. Surg. Infect. (Larchmt) 2018, 19, 804–811. [Google Scholar] [CrossRef] [PubMed]
- Guaní-Guerra, E.; Santos-Mendoza, T.; Lugo-Reyes, S.O.; Terán, L.M. Antimicrobial peptides: General overview and clinical implications in human health and disease. Clin. Immunol. 2010, 135, 1–11. [Google Scholar] [CrossRef] [PubMed]
- Zhang, L.; Gallo, R.L. Antimicrobial peptides. Curr. Biol. 2016, 26, 14–19. [Google Scholar] [CrossRef] [PubMed]
- Sharma, S.; Sahoo, N.; Bhunia, A. Antimicrobial Peptides and their Pore/Ion Channel Properties in Neutralization of Pathogenic Microbes. Curr. Top. Med. Chem. 2015, 16, 46–53. [Google Scholar] [CrossRef] [PubMed]
- Mahlapuu, M.; Håkansson, J.; Ringstad, L.; Björn, C. Antimicrobial peptides: An emerging category of therapeutic agents. Front. Cell. Infect. Microbiol. 2016, 6, 194. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bahar, A.A.; Ren, D. Antimicrobial peptides. Pharmaceuticals 2013, 6, 1543–1575. [Google Scholar] [CrossRef] [Green Version]
- Geitani, R.; Ayoub Moubareck, C.; Touqui, L.; Karam Sarkis, D. Cationic antimicrobial peptides: Alternatives and/or adjuvants to antibiotics active against methicillin-resistant Staphylococcus aureus and multidrug-resistant Pseudomonas aeruginosa. BMC Microbiol. 2019, 19, 1–12. [Google Scholar] [CrossRef] [Green Version]
- Mandal, S.M.; Roy, A.; Ghosh, A.K.; Hazra, T.K.; Basak, A.; Franco, O.L. Challenges and future prospects of antibiotic therapy: From peptides to phages utilization. Front. Pharm. 2014, 5, 1–12. [Google Scholar] [CrossRef]
- Lei, J.; Sun, L.C.; Huang, S.; Zhu, C.; Li, P.; He, J.; Mackey, V.; Coy, D.H.; He, Q.Y. The antimicrobial peptides and their potential clinical applications. Am. J. Transl. Res. 2019, 11, 3919–3931. [Google Scholar]
- Zhang, D.; He, Y.; Ye, Y.; Ma, Y.; Zhang, P.; Zhu, H.; Xu, N.; Liang, S. Little antimicrobial peptides with big therapeutic roles. Protein Pept. Lett. 2019, 26, 564–578. [Google Scholar] [CrossRef] [PubMed]
- Alexander, J.L.; Thompson, Z.; Yu, Z.; Cowan, J.A. Cu-ATCUN Derivatives of Sub5 Exhibit Enhanced Antimicrobial Activity via Multiple Modes of Action. ACS Chem. Biol. 2019, 14, 449–458. [Google Scholar] [CrossRef] [PubMed]
- Libardo, M.D.; Cervantes, J.L.; Salazar, J.C.; Angeles-Boza, A.M. Improved bioactivity of antimicrobial peptides by addition of amino-terminal copper and nickel (ATCUN) binding motifs. ChemMedChem 2014, 9, 1892–1901. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Nguyen, L.T.; Haney, E.F.; Vogel, H.J. The expanding scope of antimicrobial peptide structures and their modes of action. Trends Biotechnol. 2011, 29, 464–472. [Google Scholar] [CrossRef] [PubMed]
- Marchand, C.; Krajewski, K.; Lee, H.F.; Antony, S.; Johnson, A.A.; Amin, R.; Roller, P.; Kvaratskhelia, M.; Pommier, Y. Covalent binding of the natural antimicrobial peptide indolicidin to DNA abasic sites. Nucleic Acids Res. 2006, 34, 5157–5165. [Google Scholar] [CrossRef]
- Park, C.B.; Kim, H.S.; Kim, S.C. Mechanism of Action of the Antimicrobial Peptide Buforin II: Buforin II Kills Microorganisms by Penetrating the Cell Membrane and Inhibiting Cellular Functions. Biochem. Biophys. Res. Commun. 1998, 244, 253–257. [Google Scholar] [CrossRef] [Green Version]
- Nam, J.; Yun, H.; Rajasekaran, G.; Kumar, S.D.; Kim, J., II; Min, H.J.; Shin, S.Y.; Lee, C.W. Structural and Functional Assessment of mBjAMP1, an Antimicrobial Peptide from Branchiostoma japonicum, Revealed a Novel α-Hairpinin-like Scaffold with Membrane Permeable and DNA Binding Activity. J. Med. Chem. 2018, 61, 11101–11113. [Google Scholar] [CrossRef]
- Zhong, C.; Zhu, N.; Zhu, Y.; Liu, T.; Gou, S.; Xie, J.; Yao, J.; Ni, J. Antimicrobial peptides conjugated with fatty acids on the side chain of D-amino acid promises antimicrobial potency against multidrug-resistant bacteria. Eur. J. Pharm. Sci. 2020, 141, 105123. [Google Scholar] [CrossRef]
- Ocampo-Ibáñez, I.D.; Liscano, Y.; Rivera-Sánchez, S.P.; Oñate-Garzón, J.; Lugo-Guevara, A.D.; Flórez-Elvira, L.J.; Lesmes, M.C. A Novel Cecropin d-Derived Short Cationic Antimicrobial Peptide Exhibits Antibacterial Activity Against Wild-Type and Multidrug-Resistant Strains of Klebsiella pneumoniae and Pseudomonas aeruginosa. Evol. Bioinform. 2020, 16. [Google Scholar] [CrossRef]
- Brady, D.; Grapputo, A.; Romoli, O.; Sandrelli, F. Insect cecropins, antimicrobial peptides with potential therapeutic applications. Int. J. Mol. Sci. 2019, 20, 5862. [Google Scholar] [CrossRef] [Green Version]
- Oñate-Garzón, J.; Manrique-Moreno, M.; Trier, S.; Leidy, C.; Torres, R.; Patiño, E. Antimicrobial activity and interactions of cationic peptides derived from Galleria mellonella cecropin D-like peptide with model membranes. J. Antibiot. 2016, 70, 238–245. [Google Scholar] [CrossRef]
- Romoli, O.; Mukherjee, S.; Mohid, S.A.; Dutta, A.; Montali, A.; Franzolin, E.; Brady, D.; Zito, F.; Bergantino, E.; Rampazzo, C.; et al. Enhanced Silkworm Cecropin B Antimicrobial Activity against Pseudomonas aeruginosa from Single Amino Acid Variation. ACS Infect. Dis. 2019, 5, 1200–1213. [Google Scholar] [CrossRef] [PubMed]
- Kaushal, A.; Gupta, K.; Shah, R.; van Hoek, M.L. Antimicrobial activity of mosquito cecropin peptides against Francisella. Dev. Comp. Immunol. 2016, 63, 171–180. [Google Scholar] [CrossRef]
- Lockey, T.D.; Ourth, D.D. Formation of pores in Escherichia coli cell membranes by a cecropin isolated from hemolymph of Heliothis virescens larvae. Eur. J. Biochem. 1996, 271, 263–271. [Google Scholar]
- Wang, J.; Ma, K.; Ruan, M.; Wang, Y.; Li, Y.; Fu, Y.V.; Song, Y.; Sun, H.; Wang, J. A novel cecropin B-derived peptide with antibacterial and potential anti-inflammatory properties. PeerJ 2018, 6, e5369. [Google Scholar] [CrossRef] [PubMed]
- Chen, H.L.; Su, P.Y.; Chang, Y.S.; Wu, S.Y.; Liao, Y.D.; Yu, H.M.; Lauderdale, T.L.; Chang, K.; Shih, C. Identification of a Novel Antimicrobial Peptide from Human Hepatitis B Virus Core Protein Arginine-Rich Domain (ARD). PLoS Pathog. 2013, 9, e1003425. [Google Scholar] [CrossRef] [Green Version]
- Lee, E.; Kim, J.K.; Jeon, D.; Jeong, K.W.; Shin, A.; Kim, Y. Functional roles of aromatic residues and helices of papiliocin in its antimicrobial and anti-inflammatory activities. Sci. Rep. 2015, 5, 12048. [Google Scholar] [CrossRef] [Green Version]
- Lee, E.; Shin, A.; Kim, Y. Anti-inflammatory activities of cecropin A and its mechanism of action. Arch. Insect Biochem. Physiol. 2015, 88, 31–44. [Google Scholar] [CrossRef]
- Saugar, J.M.; Rodríguez-Hernández, M.J.; De La Torre, B.G.; Pachón-Ibañez, M.E.; Fernández-Reyes, M.; Andreu, D.; Pachón, J.; Rivas, L. Activity of cecropin A-melittin hybrid peptides against colistin-resistant clinical strains of Acinetobacter baumannii: Molecular basis for the differential mechanisms of action. Antimicrob. Agents Chemother. 2006, 50, 1251–1256. [Google Scholar] [CrossRef] [Green Version]
- Jayamani, E.; Rajamuthiah, R.; Larkins-Ford, J.; Fuchs, B.B.; Conery, A.L.; Vilcinskas, A.; Ausubel, F.M.; Mylonakisa, E. Insect-derived cecropins display activity against Acinetobacter baumannii in a whole-animal high-throughput Caenorhabditis elegans model. Antimicrob. Agents Chemother. 2015, 59, 1728–1737. [Google Scholar] [CrossRef] [Green Version]
- Navon-Venezia, S.; Kondratyeva, K.; Carattoli, A. Klebsiella pneumoniae: A major worldwide source and shuttle for antibiotic resistance. Fems Microbiol. Rev. 2017, 41, 252–275. [Google Scholar] [CrossRef] [PubMed]
- Liu, Y.; Liu, P.P.; Wang, L.H.; Wei, D.D.; Wan, L.G.; Zhang, W. Capsular Polysaccharide Types and Virulence-Related Traits of Epidemic KPC-Producing Klebsiella pneumoniae Isolates in a Chinese University Hospital. Microb. Drug Resist. 2017, 23, 901–907. [Google Scholar] [CrossRef] [PubMed]
- Brumfitt, W. Nisin, alone and combined with peptidoglycan-modulating antibiotics: Activity against methicillin-resistant Staphylococcus aureus and vancomycin-resistant enterococci. J. Antimicrob. Chemother. 2002, 50, 731–734. [Google Scholar] [CrossRef] [Green Version]
- Hancock, R.E. Cationic antimicrobial peptides: Towards clinical applications. Expert Opin. Investig. Drugs 2000, 9, 1723–1729. [Google Scholar] [CrossRef]
- Aragón-Muriel, A.; Ausili, A.; Sánchez, K.; Rojasa, O.E.; Mosquera, J.L.; Polo-Cerón, D.; Oñate-Garzón, J. Studies on the interaction of alyteserin 1c peptideand its cationic analogue with model membranes imitating mammalian and bacterial membranes. Biomolecules 2019, 9, 527. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lohner, K.; Latal, A.; Lehrer, R.I.; Ganz, T. Differential scanning microcalorimetry indicates that human defensin, HNP-2, interacts specifically with biomembrane mimetic systems. Biochemistry 1997, 36, 1525–1531. [Google Scholar] [CrossRef]
- Wang, G. Determination of solution structure and lipid micelle location of an engineered membrane peptide by using one NMR experiment and one sample. Biochim. Biophys. Acta-Biomembr. 2007, 1768, 3271–3281. [Google Scholar] [CrossRef] [Green Version]
- Shinde, R.G.; Khan, A.A.; Barik, A. Exploring the interaction of copper-esculetin complex with ct-DNA: Insight from spectroscopic and docking studies. J. Mol. Struct. 2020, 1208, 127901. [Google Scholar] [CrossRef]
- Pawar, S.K.; Jaldappagari, S. Intercalation of a flavonoid, silibinin into DNA base pairs: Experimental and theoretical approach. J. Mol. Recognit. 2020, 33. [Google Scholar] [CrossRef]
- Aragón-muriel, A.; Polo-cerón, D.; Salcedo-reyes, J.C. Synthesis, antibacterial activity and DNA interactions of lanthanide (III) complexes of N (4) -substituted thiosemicarbazones. Univ. Sci. 2018, 23, 141–169. [Google Scholar] [CrossRef]
- Schmechel, D.E.V.; Crothers, D.M. Kinetic and hydrodynamic studies of the complex of proflavine with poly A·poly U. Biopolymers 1971, 10, 465–480. [Google Scholar] [CrossRef]
- Beebe, S.J.; Celestine, M.J.; Bullock, J.L.; Sandhaus, S.; Arca, J.F.; Cropek, D.M.; Ludvig, T.A.; Foster, S.R.; Clark, J.S.; Beckford, F.A.; et al. Synthesis, characterization, DNA binding, topoisomerase inhibition, and apoptosis induction studies of a novel cobalt (III) complex with a thiosemicarbazone ligand. J. Inorg. Biochem. 2020, 203, 110907. [Google Scholar] [CrossRef]
- Shahabadi, N.; Hakimi, M.; Morovati, T.; Fatahi, N. DNA binding affinity of a macrocyclic copper (II) complex: Spectroscopic and molecular docking studies. Nucleosides Nucleotides Nucleic Acids 2017, 36, 497–510. [Google Scholar] [CrossRef]
- Polo-Cerón, D. Cu (II) and Ni (II) complexes with new tridentate NNS thiosemicarbazones: Synthesis, characterisation, DNA interaction, and antibacterial activity. Bioinorg. Chem. Appl. 2019, 2019. [Google Scholar] [CrossRef] [PubMed]
- Dong, W.; Luo, X.; Sun, Y.; Li, Y.; Wang, C.; Guan, Y.; Shang, D. Binding Properties of DNA and Antimicrobial Peptide Chensinin-1b Containing Lipophilic Alkyl Tails. J. Fluoresc. 2020, 30, 131–142. [Google Scholar] [CrossRef] [PubMed]
- CLSI. Performance Standards for Antimicrobial Susceptibility Testing. Available online: https://clsi.org/media/2663/m100ed29_sample.pdf (accessed on 5 August 2020).
- CLSI. Methods for Dilution Antimicrobial Susceptibility Tests for Bacteria That Grow Aerobically. Available online: https://clsi.org/media/1928/m07ed11_sample.pdf (accessed on 5 August 2020).
- CLSI. Methods for Dilution Antimicrobial Susceptibility Tests for Bacteria That Grow Aerobically; Approved Standard-Tenth Edition. Available online: https://clsi.org/media/1632/m07a10_sample.pdf (accessed on 5 August 2020).
- Giacometti, A.; Cirioni, O.; Barchiesi, F.; Del Prete, M.S.; Fortuna, M.; Caselli, F.; Scalise, G. In vitro susceptibility tests for cationic peptides: Comparison of broth microdilution methods for bacteria that grow aerobically. Antimicrob. Agents Chemother. 2000, 44, 1694–1696. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Birnboim, H.C.; Doly, J. A rapid alkaline extraction procedure for screening recombinant plasmid DNA. Nucleic Acids Res. 1979, 7, 1513–1523. [Google Scholar] [CrossRef] [PubMed] [Green Version]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Strain ID | Carbapenemase Gene | MIC (μg/mL) for Antibiotic/Interpretative Categories | Carba NP Result | ||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| SAM 1 | TZP 1 | FOX 2 | CAZ 3 | CRO 3 | FEP 3 | DOR 4 | ETP 4 | IPM 4 | MEM 4 | AMK 5 | GEN 5 | CIP 6 | |||
| Kp 01 | blaNDM, and blaVIM and/or blaIMP | ≥32/R | ≥128/R | ≥64/R | ≥64/R | ≥64/R | ≥64/R | ≥8/R | ≥8/R | ≥16/R | ≥16/R | 32/I | ≥16/R | =4/R | POS |
| Kp 02 | blaNDM | ≥32/R | ≥128/R | ≥64/R | ≥64/R | ≥64/R | 8/I | ≥8/R | ≥8/R | ≥16/R | ≥16/R | 16/S | 8/I | 2/R | POS |
| Kp 03 | blaKPC and blaNDM | ≥32/R | ≥128/R | ≥64/R | ≥64/R | ≥64/R | 4/I | ≥8/R | ≥8/R | ≥16/R | ≥16/R | 4/S | ≥16/R | 2/R | POS |
| Kp 04 | blaNDM | ≥32/R | ≥128/R | ≥64/R | ≥64/R | ≥64/R | ≥64/R | ≥8/R | ≥8/R | ≥16/R | ≥16/R | ≤2/S | ≥16/R | 1/R | POS |
| Kp 05 | blaKPC | ≥32/R | ≥128/R | ≥64/R | ≥64/R | ≥64/R | ≥64/R | ≥8/R | ≥8/R | ≥16/R | ≥16/R | ≥64/R | 4/S | ≥4/R | POS |
| Kp 06 | blaKPC | ≥32/R | ≥128/R | ≥64/R | ≥64/R | ≥64/R | ≤1/S | ≥8/R | 1/I | 8/I | 8/R | ≤2/S | ≤1/S | ≤0.25/S | POS |
| Kp 07 | blaKPC and blaNDM | ≥32/R | ≥128/R | ≥64/R | ≥64/R | ≥64/R | ≥64/R | ≥8/R | ≥8/R | ≥16/R | ≥16/R | 32/I | ≥16/R | ≥4/R | POS |
| Kp 08 | blaNDM | ≥32/R | ≥128/R | ≥64/R | ≥64/R | ≥64/R | ≥64/R | ≥8/R | ≥8/R | ≥16/R | ≥16/R | 16/S | 8/I | ≥4/R | POS |
| Kp 09 | blaNDM, and blaVIM and/or blaIMP | ≥32/R | ≥128/R | 8/S | 8/I | ≥64/R | 32/R | 0.25/S | ≤0.5/S | ≥16/R | 0.5/S | 16/S | 4/S | ≤0.25/S | POS |
| Kp 10 | blaKPC and blaNDM | ≥32/R | ≥128/R | ≥64/R | ≥64/R | ≥64/R | ≥64/R | ≥8/R | ≥8/R | ≥16/R | ≥16/R | 16/S | 8/I | ≥4/R | POS |
| Kp 11 | blaKPC and blaNDM | ≥32/R | ≥128/R | ≥64/R | ≥64/R | ≥64/R | ≥64/R | ≥8/R | ≥8/R | ≥16/R | ≥16/R | 8/S | ≥16/R | 2/R | POS |
| Kp 12 | blaKPC, blaNDM, and blaVIM and/or blaIMP | ≥32/R | ≥128/R | ≥64/R | ≥64/R | ≥64/R | ≥64/R | ≥8/R | ≥8/R | 8/I | ≥16/R | 16/S | ≤1/S | ≤0.25/S | POS |
| Kp 13 | blaKPC and blaNDM | ≥32/R | ≥128/R | ≥64/R | ≥64/R | ≥64/R | ≥64/R | ≥8/R | ≥8/R | ≥16/R | ≥16/R | 8/S | ≤1/S | ≤0.25/S | POS |
| Kp 14 | blaNDM | ≥32/R | ≥128/R | ≥64/R | ≥64/R | ≥64/R | ≥64/R | ≥8/R | ≥8/R | 8/I | ≥16/R | 16/S | ≥16/R | ≥4/R | POS |
| Kp 15 | blaKPC and blaNDM | ≥32/R | ≥128/R | ≥64/R | ≥64/R | ≥64/R | 32/R | ≥8/R | ≥8/R | ≥16/R | 8/R | 32/I | ≥16/R | ≥4/R | POS |
| Kp 16 | NEG | 4/S | ≤4/S | ≤4/S | ≤1/S | ≤1/S | ≤1/S | ≤0.12/S | ≤0.5/S | ≤0.25/S | ≤0.25/S | ≤2/S | ≤1/S | ≤0.25/S | NEG |
| Kp 17 | NEG | 8/S | ≤4/S | ≤4/S | ≤1/S | ≤1/S | ≤1/S | ≤0.12/S | ≤0.5/S | ≤0.25/S | ≤0.25/S | ≤2/S | ≤1/S | ≤0.25/S | NEG |
| Kp 18 | NEG | 4/S | ≤4/S | ≤4/S | ≤1/S | ≤1/S | ≤1/S | ≤0.12/S | ≤0.5/S | ≤0.25/S | ≤0.25/S | ≤2/S | ≤1/S | ≤0.25/S | NEG |
| Kp 19 | NEG | 4/S | ≤4/S | ≤4/S | ≤1/S | ≤1/S | ≤1/S | ≤0.12/S | ≤0.5/S | ≤0.25/S | ≤0.25/S | ≤2/S | ≤1/S | ≤0.25/S | NEG |
| Kp 20 | NEG | 4/S | ≤4/S | ≤4/S | ≤1/S | ≤1/S | ≤1/S | ≤0.12/S | ≤0.5/S | ≤0.25/S | ≤0.25/S | ≤2/S | ≤1/S | ≤0.25/S | NEG |
| Kp 21 | NEG | 8/S | ≤4/S | ≤4/S | ≤1/S | ≤1/S | ≤1/S | ≤0.12/S | ≤0.5/S | ≤0.25/S | ≤0.25/S | ≤2/S | ≤1/S | ≤0.25/S | NEG |
| Kp 22 | NEG | 8/S | ≤4/S | ≤4/S | ≤1/S | ≤1/S | ≤1/S | ≤0.12/S | ≤0.5/S | ≤0.25/S | ≤0.25/S | ≤2/S | ≤1/S | ≤0.25/S | NEG |
| Kp 23 | NEG | 4/S | ≤4/S | ≤4/S | ≤1/S | ≤1/S | ≤1/S | ≤0.12/S | ≤0.5/S | 0.5/S | ≤0.25/S | ≤2/S | ≤1/S | ≤0.25/S | NEG |
| Kp 24 | NEG | 8/S | ≤4/S | ≤4/S | ≤1/S | ≤1/S | ≤1/S | ≤0.12/S | ≤0.5/S | ≤0.25/S | ≤0.25/S | ≤2/S | ≤1/S | ≤0.25/S | NEG |
| Kp 25 | NEG | 4/S | ≤4/S | ≤4/S | ≤1/S | ≤1/S | ≤1/S | ≤0.12/S | ≤0.5/S | ≤0.25/S | ≤0.25/S | ≤2/S | ≤1/S | ≤0.25/S | NEG |
| Kp 26 | NEG | 4/S | ≤4/S | ≤4/S | ≤1/S | ≤1/S | ≤1/S | ≤0.12/S | ≤0.5/S | ≤0.25/S | ≤0.25/S | ≤2/S | ≤1/S | ≤0.25/S | NEG |
| Kp 27 | NEG | 4/S | ≤4/S | ≤4/S | ≤1/S | ≤1/S | ≤1/S | ≤0.12/S | ≤0.5/S | ≤0.25/S | ≤0.25/S | ≤2/S | ≤1/S | ≤0.25/S | NEG |
| Kp 28 | NEG | 4/S | ≤4/S | ≤4/S | ≤1/S | ≤1/S | ≤1/S | ≤0.12/S | ≤0.5/S | ≤0.25/S | ≤0.25/S | ≤2/S | ≤1/S | ≤0.25/S | NEG |
| Kp 29 | NEG | 4/S | ≤4/S | ≤4/S | ≤1/S | ≤1/S | ≤1/S | ≤0.12/S | ≤0.5/S | ≤0.25/S | ≤0.25/S | ≤2/S | ≤1/S | ≤0.25/S | NEG |
| Kp 30 | NEG | 4/S | ≤4/S | ≤4/S | ≤1/S | ≤1/S | ≤1/S | ≤0.12/S | ≤0.5/S | ≤0.25/S | ≤0.25/S | ≤2/S | ≤1/S | ≤0.25/S | NEG |
| Kp ATCC 2146 | blaNDM | ≥32/R | ≥128/R | ≥64/R | ≥64/R | ≥64/R | ≥64/R | ≥8/R | ≥8/R | ≥16/R | ≥16/R | ≥64/R | ≥16/R | ≥4/R | NEG |
| Ec ATCC 25922 | NEG | ≤2/S | ≤4/S | ≤4/S | ≤1/S | ≤1/S | ≤1/S | ≤0.12/S | ≤0.5/S | ≤0.25/S | ≤0.25/S | ≤2/S | ≤1/S | ≤0.25/S | NEG |
| Strain ID | Carbapenemase Gene | MIC (μg/mL) for Antibiotic/Interpretative Categories | Carba NP Result | |||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|
| TZP 1 | CAZ 2 | FEP 2 | DOR 3 | IPM 3 | MEM 3 | AMK 4 | GEN 4 | CIP 5 | CST 6 | |||
| Pa 01 | blaVIM and/or blaIMP | 64/R | 32/R | 4/S | ≥8/R | ≥16/R | 8/R | 32/I | 8/I | 2/R | ≤0.5/S | POS |
| Pa 02 | blaNDM, blaVIM and blaIMP | ≥128/R | ≥64/R | 32/R | ≥8/R | ≥16/R | ≥16/R | ≥64/R | ≥16/R | ≥4/R | ≥16/R | POS |
| Pa 03 | blaNDM, blaVIM and blaIMP | ≥128/R | ≥64/R | 32/R | ≥8/R | ≥16/R | 4/I | ≥64/R | ≥16/R | 2/R | ≤0.5/S | POS |
| Pa 04 | blaVIM and blaIMP | ≥128/R | 32/R | 32/R | ≥8/R | ≥16/R | 8/R | ≥64/R | 4/S | ≥4/R | ≥16/R | POS |
| Pa 05 | blaKPC, blaNDM, and blaVIM and/or blaIMP | ≥128/R | ≥64/R | 16/I | ≥8/R | ≥16/R | 8/R | ≥64/R | 4/S | 2/R | ≥16/R | POS |
| Pa 06 | blaNDM, and blaVIM and/or blaIMP | ≥128/R | ≥64/R | 32/R | ≥8/R | ≥16/R | ≥16/R | ≥64/R | 8/I | ≥4/R | ≥16/R | POS |
| Pa 07 | blaVIM and/or blaIMP | ≥128/R | 32/R | ≤1/S | ≥8/R | ≥16/R | 8/R | ≥64/R | ≥16/R | ≥4/R | ≥32/R | POS |
| Pa 08 | blaVIM and/or blaIMP | ≥128/R | ≥64/R | ≥64/R | ≥8/R | ≥16/R | ≥16/R | ≥64/R | ≤1/S | ≥4/R | ≤0.5/S | POS |
| Pa 09 | blaVIM and/or blaIMP | ≥128/R | ≥64/R | 32/R | ≥8/R | ≥16/R | ≥16/R | ≥64/R | 8/I | ≥4/R | ≥16/R | POS |
| Pa 10 | blaVIM and/or blaIMP | ≥128/R | ≥64/R | 32/R | ≥8/R | ≥16/R | ≥16/R | ≥64/R | ≥16/R | ≥4/R | ≤0.5/S | POS |
| Pa 11 | blaNDM, and blaVIM and/or blaIMP | ≥128/R | ≥64/R | ≥64/R | ≥8/R | ≥16/R | ≥16/R | 8/S | 4/S | 0.5/S | ≥16/R | POS |
| Pa 12 | blaKPC, blaNDM, and blaVIM and/or blaIMP | ≥128/R | 32/R | 16/I | ≥8/R | ≥16/R | 4/I | 32/I | ≥16/R | ≥4/R | ≤0.5/S | POS |
| Pa 13 | blaVIM and/or blaIMP | ≥128/R | ≥64/R | 32/R | ≥8/R | ≥16/R | ≥16/R | ≥64/R | ≥16/R | ≥4/R | ≥16/R | POS |
| Pa 14 | blaVIM and/or blaIMP | ≥128/R | ≥64/R | ≥64/R | 4/I | 1/S | 4/I | ≥64/R | ≥16/R | ≥4/R | ≤0.5/S | POS |
| Pa 15 | blaNDM, and blaVIM and/or blaIMP | ≥128/R | ≥64/R | 16/I | ≥8/R | ≥16/R | ≥16/R | ≥64/R | ≥16/R | ≥4/R | ≥16/R | POS |
| Pa 16 | NEG | 8/S | 4/S | 2/S | 0.5/S | 1/S | ≤0.25/S | ≤2/S | ≤1/S | ≤0.25/S | ≤0.5/S | NEG |
| Pa 17 | NEG | 8/S | 4/S | 2/S | ≤0.12/S | 2/S | ≤0.25/S | ≤2/S | 2/S | ≤0.25/S | ≤0.5/S | NEG |
| Pa 18 | NEG | 8/S | 4/S | 2/S | 1/S | 2/S | 1/S | ≤2/S | 2/S | ≤0.25/S | ≤0.5/S | NEG |
| Pa 19 | NEG | ≤4/S | 2/S | ≤1/S | 1/S | 1/S | 1/S | ≤2/S | ≤1/S | ≤0.25/S | ≤0.5/S | NEG |
| Pa 20 | NEG | ≤4/S | 2/S | ≤1/S | 1/S | 2/S | 1/S | ≤2/S | ≤1/S | ≤0.25/S | ≤0.5/S | NEG |
| Pa 21 | NEG | 8/S | 4/S | 2/S | ≤0.12/S | 1/S | ≤0.25/S | ≤2/S | ≤1/S | ≤0.25/S | ≤0.5/S | NEG |
| Pa 22 | NEG | 8/S | 4/S | 2/S | ≤0.12/S | ≤0.25/S | ≤0.25/S | ≤2/S | ≤1/S | ≤0.25/S | ≤0.5/S | NEG |
| Pa 23 | NEG | 8/S | 4/S | ≤1/S | ≤0.12/S | 2/S | ≤0.25/S | ≤2/S | ≤1/S | ≤0.25/S | ≤0.5/S | NEG |
| Pa 24 | NEG | 8/S | 4/S | 2/S | 1/S | 2/S | 0.5/S | ≤2/S | ≤1/S | ≤0.25/S | ≤0.5/S | NEG |
| Pa 25 | NEG | 8/S | 2/S | 2/S | 0.5/S | 1/S | 0.5/S | ≤2/S | 2/S | ≤0.25/S | ≤0.5/S | NEG |
| Pa 26 | NEG | 8/S | 4/S | 2/S | ≤0.12/S | 2/S | ≤0.25/S | ≤2/S | 2/S | ≤0.25/S | ≤0.5/S | NEG |
| Pa 27 | NEG | ≤4/S | 2/S | ≤1/S | ≤0.12/S | 1/S | ≤0.25/S | ≤2/S | ≤1/S | ≤0.25/S | ≤0.5/S | NEG |
| Pa 28 | NEG | 8/S | 4/S | 2/S | 0.25/S | 1/S | ≤0.25/S | ≤2/S | ≤1/S | ≤0.25/S | ≤0.5/S | NEG |
| Pa 29 | NEG | 8/S | 4/S | ≤1/S | 0.25/S | 2/S | ≤0.25/S | ≤2/S | ≤1/S | ≤0.25/S | ≤0.5/S | NEG |
| Pa 30 | NEG | 8/S | 4/S | 2/S | ≤0.12/S | 1/S | ≤0.25/S | ≤2/S | ≤1/S | ≤0.25/S | ≤0.5/S | NEG |
| Pa ATCC 27853 | NEG | ≤4/S | ≤1/S | ≤1/S | 0.25/S | 2/S | ≤0.25/S | ≤2/S | ≤1/S | ≤0.25/S | ≤0.5/S | NEG |
| K. pneumoniae (n) | MIC (μg/mL) | MBC (μg/mL) | P. aeruginosa (n) | MIC (μg/mL) | MBC (μg/mL) | p-Value 1 |
|---|---|---|---|---|---|---|
| WTKp (15) | 4–8 | 4–8 | WTPa (15) | 8 | 8 | 0.1833 |
| MDRKp (15) | 8–16 | 8–16 | MDRPa (15) | 8–16 | 8–16 | 0.0016 |
| Ec ATCC 25922 | 4–8 | 4–8 | Pa ATCC 27853 | 4–8 | 4–8 | |
| Kp ATCC 2146 | 8–16 | 8–16 |
| Peptide-Lipid Molar Ratio | Pre-Transition Temperature (°C) | Tm 1 (°C) | DH 2 (cal/g) | FWHM 3 (°C) |
|---|---|---|---|---|
| 0:1 | 13.55 | 22.98 | 0.32 | 0.55 |
| 1:10 | NA | 23.96 | 0.03 | 3.69 |
Sample Availability: Samples of the compounds are not available from the authors. |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Rivera-Sánchez, S.P.; Agudelo-Góngora, H.A.; Oñate-Garzón, J.; Flórez-Elvira, L.J.; Correa, A.; Londoño, P.A.; Londoño-Mosquera, J.D.; Aragón-Muriel, A.; Polo-Cerón, D.; Ocampo-Ibáñez, I.D. Antibacterial Activity of a Cationic Antimicrobial Peptide against Multidrug-Resistant Gram-Negative Clinical Isolates and Their Potential Molecular Targets. Molecules 2020, 25, 5035. https://doi.org/10.3390/molecules25215035
Rivera-Sánchez SP, Agudelo-Góngora HA, Oñate-Garzón J, Flórez-Elvira LJ, Correa A, Londoño PA, Londoño-Mosquera JD, Aragón-Muriel A, Polo-Cerón D, Ocampo-Ibáñez ID. Antibacterial Activity of a Cationic Antimicrobial Peptide against Multidrug-Resistant Gram-Negative Clinical Isolates and Their Potential Molecular Targets. Molecules. 2020; 25(21):5035. https://doi.org/10.3390/molecules25215035
Chicago/Turabian StyleRivera-Sánchez, Sandra Patricia, Helen Astrid Agudelo-Góngora, José Oñate-Garzón, Liliana Janeth Flórez-Elvira, Adriana Correa, Paola Andrea Londoño, Juan David Londoño-Mosquera, Alberto Aragón-Muriel, Dorian Polo-Cerón, and Iván Darío Ocampo-Ibáñez. 2020. "Antibacterial Activity of a Cationic Antimicrobial Peptide against Multidrug-Resistant Gram-Negative Clinical Isolates and Their Potential Molecular Targets" Molecules 25, no. 21: 5035. https://doi.org/10.3390/molecules25215035