
Participation of MicroRNAs in the Treatment of Cancer with Phytochemicals
by

Seung Wan Son
1,†, 
Han Yeoung Lee
1,†,
Sokviseth Moeng
1, 
Hyo Jeong Kuh
2,
Soo Young Choi
1 and
Jong Kook Park
1,* 1
Department of Biomedical Science and Research Institute for Bioscience & Biotechnology, Hallym University, Chunchon 24252, Korea
2
Department of Medical Life Sciences, College of Medicine, The Catholic University of Korea, Seoul 06591, Korea
*
Author to whom correspondence should be addressed.
†
These authors contributed equally to this work.
Molecules 2020, 25(20), 4701; https://doi.org/10.3390/molecules25204701
Submission received: 31 August 2020
/
Revised: 8 October 2020
/
Accepted: 13 October 2020
/
Published: 14 October 2020
(This article belongs to the Special Issue 25th Anniversary of Molecules—Hot Topics in Bioorganic Chemistry)
Abstract
:Cancer is a global health concern and one of the main causes of disease-related death. Even with considerable progress in investigations on cancer therapy, effective anti-cancer agents and regimens have thus far been insufficient. There has been compelling evidence that natural phytochemicals and their derivatives have potent anti-cancer activities. Plant-based anti-cancer agents, such as etoposide, irinotecan, paclitaxel, and vincristine, are currently being applied in medical treatments for patients with cancer. Further, the efficacy of plenty of phytochemicals has been evaluated to discover a promising candidate for cancer therapy. For developing more effective cancer therapy, it is required to apprehend the molecular mechanism deployed by natural compounds. MicroRNAs (miRNAs) have been realized to play a pivotal role in regulating cellular signaling pathways, affecting the efficacy of therapeutic agents in cancer. This review presents a feature of phytochemicals with anti-cancer activity, focusing mainly on the relationship between phytochemicals and miRNAs, with insights into the role of miRNAs as the mediators and the regulators of anti-cancer effects of phytochemicals.
1. Introduction
Cancer is generally incurable and life-threatening. According to the GLOBOCAN database, there were approximately 18.1 million new cancer cases and 9.6 million cancer deaths worldwide in 2018 [1]. Chemotherapy and radiotherapy are currently used in cancer treatments; however, they are moderately effective, leading to the insufficient regression and remission of cancer. Moreover, the adverse effects of chemotherapy, such as toxicity to normal cells and therapeutic resistance, restrict the utilization of anti-cancer agents [2,3,4]. Therefore, intensive efforts have been made to continuously search for and develop new agents and strategies for cancer therapy with better efficacy.
Natural compounds, procured from several resources, have significant therapeutic potentials for diverse types of diseases, including cancer, and provide critical guides for drug discovery. For example, psammaplin A and didemnin B from marine organisms show anti-cancer properties and effectively induce apoptosis [5]. Aphidicolin is a metabolite from fungi, such as Harziella entomophilla, and exhibits anti-cancer activity by inhibiting DNA replication and sensitizing cancer cells to therapeutic agents, such as fludarabine and cladribine in chronic lymphocytic leukemia [6]. In addition, various plant-derived phytochemicals and their derivatives possess a high potential as anti-cancer agents and are still under evaluation. Mounting evidence from preclinical studies demonstrated that phytochemical compounds effectively repress cancer progression and malignancy, such as cell proliferation, growth, invasion, and metastasis [7,8]. Additionally, anti-cancer agents currently used in clinical therapy are derived from plants. Well-known examples include etoposide, irinotecan, paclitaxel, and vincristine. Primary molecular targets of irinotecan and etoposide are topoisomerase I and topoisomerase II, respectively [9,10]. Paclitaxel and vincristine are microtubule inhibitors, displaying anti-cancer properties, such as cell cycle arrest and apoptosis [10,11].
MicroRNAs (miRNAs) are small regulatory non-coding RNAs that can affect multiple cellular signaling pathways by controlling the degradation and translation of their target messenger RNAs (mRNAs). Since miRNA levels can be epigenetically, transcriptionally, and/or post-transcriptionally regulated, miRNAs are able to mediate the effects of several stimuli such as a drug. For example, paclitaxel elevates the level of miR-512-3p that targets an anti-apoptotic gene, FADD-like apoptosis regulator (CFLAR, also known as c-FLIP), suggesting that miR-512-3p is responsible for paclitaxel-induced apoptosis to a degree in hepatocellular carcinoma cells [12]. In terms of cancer, miRNAs serve as oncogenic or tumor-suppressive factors depending on their target genes and cell types [4,13,14]. Additionally, dysregulated miRNAs can influence the sensitivity of cancer cells to anti-cancer treatments by modulating cellular events, such as apoptosis, autophagy, stemness, drug efflux, drug metabolism, and epithelial-mesenchymal transition (EMT) [4,15].
An improved understanding of the mode of action of phytochemicals at the molecular levels will be beneficial to establish novel and effective therapeutic strategies to strive against cancer. This article aims to present a review of the role of miRNAs in mediating and regulating the anti-cancer effects of phytochemicals. In the case of plant-derived compounds currently applied in clinical therapy, we focused on the involvement of miRNAs in the occurrence of resistance to these compounds. The phytochemical compounds and their derivatives mentioned in this review are summarized in Table 1 and Table 2.
2. Oncogenic MiRNAs Inhibited by Phytochemicals Currently Evaluated in Preclinical Studies and Clinical Trials
2.1. MiRNAs and Nitrogen-Containing Compounds
2.1.1. Berberine and Evodiamine
The miR-99a–125b cluster located at chromosome 21 consists of three miRNAs, namely miR-99a, let-7c, and miR-125b. These miRNAs have been validated as oncogenic or tumor-suppressive miRNAs depending on the type of cancer. For example, miR-99a can inhibit proliferation, migration, and invasion by directly regulating fibroblast growth factor receptor 3 (FGFR3) in breast cancer [87]. In multiple myeloma (MM), miR-125b is known to suppress apoptosis induced by dexamethasone via targeting tumor protein p53 (TP53) [88]. Recently, it was demonstrated that the levels of miR-99a–125b are downregulated by berberine treatments and that the knockdown of miR-99a–125b causes cell cycle arrest as well as apoptosis induction in MM [17] (Figure 1 and Table 3).
One of the critical miRNAs involved in the progression of colorectal cancer is miR-429. It has been noticed that this miRNA is overexpressed in colorectal cancer tissues compared to their normal counterparts and that miR-429 is able to augment EMT and metastasis of colorectal cancer by modulating the expression of homeobox A5 (HOXA5) [89]. Additionally, treatments of colorectal cancer with either berberine or evodiamine result in a decrease in miR-429 expression [90], suggesting that the anti-cancer activity of both phytochemicals is partly mediated by the modulation of miR-429 levels (Figure 1 and Table 3).
2.1.2. Matrine
Matrine has been reported to elicit anti-cancer effects on multiple cancer types. For example, matrine suppresses the migration and invasion capacities of lung, prostate, and breast cancer cells via downregulating the levels of C-X-C motif chemokine receptor 4 (CXCR4) [107]. In addition, it was demonstrated that matrine represses miR-182-5p and miR-93-5p levels in papillary thyroid and gastric cancer cells, respectively, and retards cancer growth in vivo [21,96] (Figure 1 and Table 3). In particular, the overexpression of miR-182-5p (a member of the miR-183-96-182 cluster) can block the matrine-mediated activation of caspase-3 in papillary thyroid cancer cells [21]. In addition, miR-93-5p (a member of the miR-106b-25 cluster) positively affects cell proliferation and the migration of gastric cancer cells via directly targeting AHNAK nucleoprotein (AHNAK), which is a negative regulator of the EMT process [96,108].
2.1.3. Neferine
Fibroblast growth factor receptor 2 (FGFR2) is known to be overexpressed in subsets of breast cancer tissues and is negatively correlated with the overall survival of breast cancer patients [109,110]. It was recently shown that treatments with neferine restrain the proliferation, migration, and invasion of breast cancer cells via downregulating miR-374a, which positively controls FGFR2 levels [22] (Figure 1 and Table 3). Neferine has been shown to sensitize cancer cells to oxaliplatin and tumor necrosis factor-related apoptosis-inducing ligand (TRAIL) in hepatocellular carcinoma and prostate cancer, respectively [111,112]. Since FGFR2 can confer tamoxifen resistance in estrogen-positive breast cancer cells [113], it is feasible that neferine also reverses FGFR2-mediated tamoxifen resistance.
2.1.4. Nitidine Chloride and Swainsonine
c-Myc, an oncogenic transcription factor, positively controls cell proliferation and decelerates the senescence and differentiation of leukemia [114,115,116]. The miR-17–92 cluster containing six different miRNAs (miR-17, miR-18a, miR-19a, miR-19b-1, miR-20a, and miR-92a) is transcriptionally induced by c-Myc, contributing to the maintenance of cell proliferation and survival via targeting multiple genes, including BCL2-like 11 (BCL2L11, also known as Bim) in leukemia [117]. A recent study demonstrated that nitidine chloride attenuates c-Myc expression levels via ubiquitin-mediated degradation and induces differentiation and apoptosis by upregulating cyclin-dependent kinase inhibitor 1A (CDKN1A, also known as p21Cip1) in leukemia. Indeed, nitidine chloride reduces miR-17 and miR-20a levels. The overexpression of these miRNAs reduces CDKN1A levels in leukemia cells, indicating that the anti-cancer activity of nitidine chloride is mediated by the c-Myc-miR-17–92 axis [91] (Figure 1 and Table 3).
In glioma, miR-92a has been identified to accelerate proliferation, cell survival, and metastasis through regulating BCL2L11, cadherin 1 (CDH1, also known as E-cadherin) and the Akt/mechanistic target of rapamycin kinase (mTOR) signaling [118,119,120]. It was shown that the treatment of glioma cells with swainsonine hinders cell proliferation, migration, and invasion via downregulating miR-92a levels and Akt/mTOR activities [28] (Figure 1 and Table 3).
2.1.5. Piperlongumine
Piperlongumine has been identified to induce reactive oxygen species (ROS), thus exhibiting intense anti-cancer activity and sensitizing cells to anti-cancer agents such as paclitaxel [24,121]. Increased ROS following a piperlongumine treatment diminishes the expression levels of both c-Myc and c-Myc-regulated miRNAs (miR-17, miR-20a, and miR-27a) [24] (Figure 1 and Table 3). Downregulation of these miRNAs further leads to the induction of their target mRNAs, zinc finger and BTB domain-containing protein 4 (ZBTB4) and ZBTB10. Both ZBTB4 and ZBTB10 are negative regulators of specificity protein (SP) transcription factors, such as SP1 and SP3. Thus, piperlongumine can inhibit several oncogenic factors, such as the epidermal growth factor receptor (EGFR), c-MET, and survivin, which are transcribed by SP transcription factors [24].
2.1.6. Sanguinarine
Sanguinarine has been shown to have anti-cancer activities against several types of cancer. For instance, sanguinarine can suppress proliferation and cell viability by regulating the levels of ROS and pro-apoptotic genes in cervical cancer cells [122]. Sanguinarine also impedes EMT by inactivating the Wingless (Wnt)/β-catenin and transforming growth factor-beta (TGF-β) signaling pathways in colorectal cancer and hepatocellular carcinoma, respectively [123,124]. In gastric cancer, sanguinarine blocks cell proliferation, together with a decrease in miR-29-3p and miR-96-5p levels [25] (Figure 1, Table 3 and Table 4). MiR-96-5p has been found to accelerate proliferation via negatively modulating forkhead box O3 (FOXO3), which can induce apoptotic cell death [125,126]. In the case of miR-29-3p, overexpression of miR-29-3p remarkably restrains the migration and invasion of gastric cancer cells by suppressing multiple oncogenes such as FGFR4 [127,128], implying that sanguinarine can unexpectedly downregulate tumor-suppressive miRNAs.
2.1.7. Sinomenine
In prostate cancer, sinomenine inactivates the phosphoinositide 3-kinase (PI3K)/Akt and Janus kinase (JAK)/signal transducer and activator of transcription 3 (STAT3) signal pathways, leading to the inhibition of cell viability, migration, and invasion [26]. In that study, it was noted that miR-23 is downregulated in sinomenine-treated cells and that the ectopic introduction of miR-23 reverses the anti-cancer effects of sinomenine on prostate cancer cells [26]. Another study also showed that the activity of nuclear factor kappa B (NF-κB) is dampened by sinomenine, thus reducing invasion and migration of breast cancer cells [104]. Furthermore, it is noteworthy that miR-324-5p levels are decreased by sinomenine treatments and that this miRNA directly targets the CUE domain-containing 2 (CUEDC2), a negative regulator of NF-κB [104] (Figure 1 and Table 3).
2.1.8. Sophocarpine
One of the most enormously explored oncogenic miRNAs is miR-21. The levels of miR-21 are upregulated in several cancers and are correlated with metastasis status [155,156]. In addition, miR-21 promotes EMT via negatively regulating several EMT-inhibiting factors, including phosphatase and tensin homolog (PTEN), SRY-box transcription factor 17 (SOX17), and leucine zipper transcription factor-like 1 (LZTFL1) [155,157,158]. Interestingly, cell-free assays indicated that sophocarpine binds to miR-21 precursors and blocks the Dicer-mediated maturation of miR-21; therefore, sophocarpine can inhibit the progression of head and neck cancer by negatively modulating EMT [27] (Figure 1 and Table 3).
2.2. MiRNAs and an Organosulfur Compound
Sulforaphane
Several studies demonstrated that miRNAs involve the regulation of cell cycle, senescence, and apoptosis in sulforaphane-treated cancer cells [30,93,94]. In glioblastoma, sulforaphane suppresses the activity of transcription factor 4 (TCF4), a downstream factor of Wnt/β-catenin signaling, thereby reducing the levels of miR-21 [93]. Such downregulation of miR-21 can potentiate temozolomide-induced apoptosis when combined with sulforaphane, suggesting that sulforaphane can be considered as a promising adjuvant candidate for glioblastoma therapy [93] (Figure 1 and Table 3).
Sulforaphane also shows anti-cancer efficacy by suppressing cell proliferation and inducing apoptosis in colorectal cancer. In that study, it was demonstrated that sulforaphane reduces the expression of both miR-21 and telomerase reverse transcriptase (TERT) [30]. TERT is generally recognized to exhibit multiple oncogenic activities, contributing to the regulation of angiogenesis, stemness, EMT, and metastasis [159]. Downregulation of TERT levels can be due to the ability of sulforaphane to turn off the transcription of TERT by recruiting methyl-CpG binding protein 2 (MECP2) to the TERT promoter [160]. Another possibility is that TERT levels can be positively controlled by miR-21 since TERT transcription is mediated by ERK1/2 signaling activated by this miRNA [161,162] (Figure 1 and Table 3).
In breast cancer, sulforaphane was noticed to promote cell cycle arrest, cellular senescence, and apoptosis induction by modulating global DNA methylation status and miRNA levels. For example, oncogenic miRNAs (miR-23 and miR-382) are downregulated by the treatment with sulforaphane [94]. It has been demystified that knockdown of miR-23 abates the growth of breast cancer in vivo and that miR-382 triggers breast cancer metastasis by activating Ras/ERK signaling pathways via targeting RAS-like estrogen-regulated growth inhibitor (RERG) [163,164] (Figure 1 and Table 3).
2.3. MiRNAs and Phenolic Compounds
2.3.1. Baicalin
Apoptotic cell death is induced by baicalin treatments, and further evidence showed that various oncogenic miRNAs, such as miR-23 and miR-217, can be depleted by baicalin in colorectal cancer [36,99] (Figure 2 and Table 3). Baicalin impedes the growth of colorectal cancer in vivo via repressing c-Myc expression in conjunction with a reduction of miR-23, which is one of the c-Myc-regulated miRNAs [36]. Additionally, baicalin can induce apoptosis through subduing Wnt/β-catenin signaling, accompanied by the upregulated levels of Dickkopf-related protein 1 (DKK1), an endogenous inhibitor of Wnt signaling [99]. In this study, it was noted that baicalin downregulates the expression of miR-217, which undeviatingly targets DKK1.
2.3.2. Curcumin
Reversion-inducing cysteine-rich protein with Kazal Motifs (RECK) is known to suppress proliferation, invasion, and metastasis primarily through inhibiting matrix metalloproteinases (MMPs), such as MMP-2 and MMP-9, in multiple cancer types [165,166,167]. A recent study demonstrated that the treatment of osteosarcoma cells with curcumin decelerates cell proliferation, along with a decrease in miR-21, which targets RECK [39]. The miR-21/RECK axis can support the previous finding that curcumin inhibits cancer metastasis [168,169] (Figure 2 and Table 3).
It is acknowledged that human growth hormone (hGH) facilitates proliferation, survival, EMT, etcetera, in cancer [170,171,172]. In breast cancer, hGH is capable of increasing the levels of the miR-183-96-182 cluster members via transcription factors, such as signal transducer and activator of transcription 3 (STAT3) and STAT5 [170]. This miR-183-96-182 cluster is overexpressed in breast cancer and prompts proliferation and migration [173]. Additionally, these cluster members facilitate cancer invasion, EMT, and metastasis through commonly targeting breast cancer metastasis-suppressor 1-like (BRMS1L) [170]. Recent evidence suggested that curcumin can block the expression of miR-183-96-182 cluster induced by hGH, ultimately preventing hGH-mediated breast cancer aggressiveness [97] (Figure 2 and Table 3).
Besides, curcumin diminishes the expression of miR-7641, which directly controls CDKN2A (also called p16Ink4A) [106]. This study showed that curcumin weakens bladder cancer invasion coupled with apoptosis induction by elevating CDKN2A levels [106]. CDKN2A has been recognized to negatively regulate metastasis-related signaling and induce apoptosis following cell cycle arrest [174,175] (Figure 2 and Table 3).
2.3.3. Epigallocatechin Gallate
Members of the miR-106b–25 cluster (miR-25, miR-93, and miR-106b) have been realized to contribute to the progression of breast cancer. For example, miR-106 potentiates the metastatic potential of breast cancer by increasing the activity of Rho-associated coiled-coil containing protein kinase 1 (ROCK1) [176]. Moreover, miR-25 can inhibit apoptotic cell death by suppressing B-cell translocation gene 2 (BTG2), a negative regulator of Akt and ERK [177]. Recently, it was found that miR-25 is suppressed by epigallocatechin gallate (EGCG) in breast cancer cells, contributing to the induction of apoptosis in vitro and the retardation of cancer growth in vivo [95] (Figure 2 and Table 3).
2.3.4. Formononetin, Galangin, Gambogic Acid, Honokiol, and Puerarin
By suppressing miR-21, diverse phytochemicals can exert anti-cancer effects. As stated in Section 2.1.8, miR-21 directly modulates PTEN, whose function is to inactivate PI3K/Akt signaling in cancer. As a consequence of the miR-21 inhibition, formononetin and galangin upregulate PTEN levels in bladder and cholangiocarcinoma, respectively, thereby inhibiting cancer aggressiveness [42,43]. In colorectal cancer, gambogic acid induces apoptosis by suppressing miR-21 expression, and the cytotoxicity of gambogic acid is reversed by overexpressing miR-21 [44]. Additionally, honokiol and puerarin act as miR-21 repressors and suppress multiple cellular events, such as proliferation, invasion, and EMT [48,57] (Figure 2 and Table 3).
2.3.5. Genistein
F-Box and WD repeat domain containing 7 (FBXW7), a tumor-suppressive E3 ligase, is known to degrade SHOC2 leucine-rich repeat scaffold protein (SHOC2), which activates Ras/ERK signaling in pancreatic cancer [178]. Recently, it was investigated that exposure to genistein attenuates pancreatic cancer proliferation and invasion by downregulating miR-223, which targets SHOC2 [101] (Figure 2 and Table 3). The transcription of miR-223 is enhanced by CCAAT/enhancer-binding protein beta (CEBPB), and genistein can inhibit CEBPB [179,180]. Therefore, it is feasible that genistein can reduce miR-223 levels via a CEBPB downregulation.
2.3.6. Icariin, Isoliquiritigenin, and Luteolin
The anti-cancer effects of icariin were assessed in thyroid cancer cells [49]. It showed that cell viability is suppressed in icariin-treated cells, concomitantly with a decrease in migration and invasion of cancer cells. After treatment with icariin, miR-625-3p levels are decreased, along with the inactivation of Akt and ERK (Figure 2 and Table 3). Ectopic introduction of miR-625-3p reactivates both Akt and ERK, indicating that miR-625-3p serves as an oncogenic factor [49]. However, miR-625-3p can negatively affect the proliferation and metastasis of gastric cancer cells [181], implying that the function of miR-625-3p is cell-type dependent.
Recent reports have demonstrated that anti-cancer activity of isoliquiritigenin is accomplished by downregulating oncogenic miRNAs. The expression of miR-32 is reduced by isoliquiritigenin in nasopharyngeal cancer cells, hence increasing the levels of large tumor suppressor kinase 2 (LATS2), a target mRNA of miR-32. LATS2 is responsible for the reduction of cancer growth by inhibiting Wnt/β-catenin signaling [50]. Further, it was observed that the proliferation and survival of melanoma cells are retarded by isoliquiritigenin and that one of the significantly downregulated miRNAs is miR-301-3p. The evaluation of miR-301-3p activity indicated that this miRNA functions as an oncogene by targeting leucine-rich repeats and immunoglobulin-like domains protein 1 (LRIG1) [102]. Furthermore, PTEN is upregulated by isoliquiritigenin, owing to the reduction of miR-374a levels, hence subduing the migration and invasion of breast cancer cells [105] (Figure 2 and Table 3).
Luteolin causes cell cycle arrest, induces apoptosis, and inhibits metastasis via repressing the expression of oncogenic factors, such as TERT, c-Myc, and MMPs [150,182]. Besides, luteolin can sensitize ovarian cancer and breast cancer cells to cisplatin and tamoxifen, respectively [183,184]. Moreover, luteolin diminishes the levels of miR-301-3p that targets caspase-8, ultimately sensitizing pancreatic cancer cells to the TNF superfamily member 10 (TNFSF10, also known as TNF-related apoptosis-inducing ligand (TRAIL)) [103] (Figure 2 and Table 3).
2.3.7. Physcion 8-O-β-Glucopyranoside and Procyanidin
Ferroptotic cell death can be induced by iron-dependent lipid peroxidation. Additionally, it was denoted that glutaminolysis catalyzed by glutaminase 2 (GLS2) facilitates ferroptotic cell death [185,186]. Interestingly, physcion 8-O-β-glucopyranoside (PG) is able to cause the ferroptotic cell death of gastric cancer cells by downregulating the levels of miR-103-3p and upregulating GLS2, a target of miR-103-3p [55] (Figure 2 and Table 3). Unlike wild-type TP53, mutant TP53 is incapable of regulating GLS2 transcription [89]. Since TP53 is widely mutated in cancer, it can be postulated that the miR-103-3p-GLS2 axis is an alternative way to modulate the ferroptotic cell death of cancer cells.
In lung cancer, miR-19a and miR-19b, members of the miR-17–92 cluster, were evaluated to be lowered by procyanidin. Also, procyanidin causes growth retardation of lung cancer in vivo, along with an increase in the level of PTEN, owing to the reduction of these miRNAs [56] (Figure 2 and Table 3). Indeed, miR-19a and miR-19b are validated miRNAs that target PTEN [187,188].
2.3.8. Resveratrol
Resveratrol has been shown to exhibit multitudinous anti-cancer effects owing to its ability to inhibit diverse signaling pathways involved in cell growth, apoptosis, and migration. For example, resveratrol has been known to inhibit c-Myc, thereby inducing cell cycle arrest and apoptotic cell death [189,190]. In breast cancer, resveratrol was found to promote the lysis of cancer cells by natural killer (NK) cells in vitro and in vivo via suppressing the expression of miR-17 in a c-Myc-dependent manner. In that study, it was unveiled that miR-17 directly targets MHC class I polypeptide-related sequence A (MICA) and MICB, which are ligands for NK cells [92]. In acute lymphoblastic leukemia, resveratrol inhibits cell proliferation and viability by downregulating the levels of miR-196b and miR-1290, both of which directly regulate the expression of insulin-like growth factor-binding protein 3 (IGFBP3) [98] (Figure 2 and Table 3).
Resveratrol obstructs the activation of NF-κB, thereby inhibiting the levels of anti-apoptotic and pro-proliferative genes, such as B-cell CLL/Lymphoma 2 (BCL-2) and cyclin D1 [191,192]. Moreover, in melanoma, the anti-cancer activity of resveratrol is mediated by downregulating the NF-κB activity and the levels of NF-κB-regulated miR-221, which targets the tropomyosin-receptor kinase fused gene (TFG) [59]. While TFG was supposed to be a tumor-suppressive gene in melanoma [193], its role in melanoma has been understudied. Therefore, it is still further required to investigate the function of TFG. In thyroid cancer, it was demonstrated that miR-222 is downregulated by resveratrol in a cell-line dependent manner [100]. The seed sequence of miR-222 is identical to miR-221; therefore, miR-221 and miR-222 can target the same mRNAs. For example, miR-221 and miR-222 can facilitate cell proliferation by targeting CDKN1B (also known as p27Kip1) in thyroid cancer cells [194] (Figure 2 and Table 3).
2.3.9. Silibinin
Both miR-21 and miR-155 can promote cancer aggressiveness via commonly targeting a tumor-suppressive gene such as cell adhesion molecule 1 (CADM1) [195,196]. Additionally, these two miRNAs were identified as possible biomarkers for breast cancer since their levels in plasma from cancer patients display the correlation with the degree of cancer progression [197]. Furthermore, it was recently revealed that the treatment of breast cancer cells with silibinin results in the induction of apoptosis and the attenuation of both miR-21 and miR-155 levels [61] (Figure 2 and Table 3). Silibinin and its derivative (e.g., glyco-conjugated silibinin) have antioxidant activities and can protect gastric and hepatocellular carcinoma cells from xanthine oxidase-induced oxidative stress [62,63]. Therefore, it is still further required to investigate the effects of silibinin on different cancer cell types.
2.4. MiRNAs and Terpenoid Compounds
2.4.1. Brucein D
CUGBP Elav-like family member 2 (CELF2, also called CUGBP2) is a tumor suppressor that can inhibit cancer growth and induce apoptosis. For instance, the knockdown of CELF2 leads to a decrease in apoptosis induced by ionizing radiation [198]. Also, CELF2 activates PTEN and suppresses the translation of oncogenes, such as vascular endothelial growth factor (VEGF) and cyclooxygenase-2 (COX-2) [199,200]. The efficacy of brucein D against cancer has been tested. For example, it was found that brucein D can inhibit pancreatic cancer growth by inducing ROS-mediated cell death [201]. Moreover, in hepatocellular carcinoma, it was noticed that the anti-cancer activity of brucein D is exerted by suppressing a CELF2-targeting miRNA, miR-95 [70] (Figure 3 and Table 3).
2.4.2. Celastrol
Celastrol has an efficient anti-cancer property against several cancer types. In ovarian cancer, apoptosis induced by celastrol is mediated by ROS generation [202]. Additionally, celastrol hinders gastric cancer growth in vivo by modulating a number of cell-cycle and apoptosis-related genes such as CDKN1B [203]. Further evidence demonstrated that celastrol shows an anti-proliferation effect by inactivating the PI3K/Akt pathway through the downregulation of miR-21 in colorectal cancer [72] (Figure 3 and Table 3).
2.4.3. Ginsenosides
Ginsenosides (G), such as G-Rd and G-Rh2, have been shown to possess anti-cancer properties through stimulating apoptosis, facilitating cell differentiation, and repressing cancer stemness, as well as angiogenesis [204,205,206]. Moreover, the association between ginsenosides and oncogenic miRNAs has been reported. In breast cancer, G-Rd shows an anti-metastatic efficacy via attenuating the levels of miR-18a, a member of the miR-17–92 cluster [75]. Their study also found that miR-18a can target SMAD family member 2 (SMAD2) [75]. It has been reported that reduced levels of SMAD2 expedite the growth and metastasis of breast cancer [207,208].
MiR-4295 was identified to accelerate cell proliferation and metastasis [209]. Also, this miRNA regulates PI3K/Akt signaling to confer resistance to cisplatin in gastric cancer cells [210]. In prostate cancer, it was demonstrated that G-Rh2 negatively regulates cell proliferation since G-Rh2 can suppress miR-4295, which directly interacts with the 3′ untranslated region (3′ UTR) of CDKN1A [76] (Figure 3 and Table 3).
2.4.4. Maytenin, 22-β-Hydroxymaytenin, and Oridonin
The expression of the miR-17–92 cluster is known to be downregulated by several phytochemicals. Recently, it was noticed that the treatment of head and neck cancer cells with either maytenin or 22-β-hydroxymaytenin shows cytotoxic effects, together with a profound decrease in miR-17 and miR-20a levels. Additionally, maytenin limits metastasis of cancer in vivo, with minimal toxicity to the kidneys [78]. Moreover, oridonin diminishes the expression of miR-17 and miR-20a and triggers cell death in both doxorubicin-sensitive and -resistant leukemia cells via derepressing BCL2L11, a target mRNA of miR-17 and miR-20a [80] (Figure 3 and Table 3).
In the case of maytenin, it also reduces the levels of miR-27a-3p [78] (Figure 3 and Table 3). However, this miRNA was recently identified to inhibit EMT via negatively regulating Yes1-associated transcriptional regulator (YAP1) in head and neck cancer cells [211]. These findings suggest that maytenin adversely affects the expression of a tumor-suppressive miRNA.
2.4.5. Triptolide
Similarly, in hepatocellular carcinoma cells, triptolide upregulates several tumor suppressors, including BCL2L11, PTEN, and CDKN1A, due to its capability to block the activation of ERCC excision repair 3, TFIIH core complex helicase subunit (ERCC3). Triptolide-mediated inactivation of ERCC3 represses the transcription of c-Myc and the miR-17–92 cluster [83]. In addition, the miR-106b–25 cluster levels are repressed by triptolide in a c-Myc-dependent manner [83], indicating that triptolide potentially inhibits miRNA clusters that play an oncogenic role (Figure 3 and Table 3).
2.4.6. α-Pinene
In hepatocellular carcinoma, miR-221 has been well known to promote cell survival and growth via the regulation of various tumor suppressors, such as PTEN and CDKN1B [212,213]. Moreover, the downregulation of miR-221 by α-pinene was shown to participate in the suppression of hepatocellular carcinoma progression via inducing cell cycle arrest and apoptosis through activating cellular factors, including CDKN1B [86] (Figure 3 and Table 3).
3. Tumor-Suppressive MiRNAs Induced by Phytochemicals Currently Tested in Preclinical Studies and Clinical Trials
3.1. MiRNAs and Nitrogen-Containing Compounds
3.1.1. 5-Aminolevulinic acid
5-Aminolevulinic acid (5-ALA) can be utilized as a sensitizer for photodynamic and sonodynamic therapies. The generation of ROS contributes to the anti-cancer effects of these 5-ALA-based therapies [214,215]. Moreover, the expression of miRNAs can be changed by 5-ALA-based therapies. 5-ALA-based photodynamic therapy increases the levels of miR-143 in cervical cancer, leading to enhanced apoptosis, along with an efficient decline of BCL-2 levels [142]. In melanoma, miR-34 is transcriptionally activated via TP53 following 5-ALA-based sonodynamic therapy [132] (Figure 1 and Table 4). This study also pointed out that miR-34 can promote TP53 activity by targeting sirtuin 1, a suppressor of TP53.
3.1.2. Coptisine
One of the significantly downregulated miRNAs in hepatocellular carcinoma is miR-122. By targeting ADAM metallopeptidase domain 7 (ADAM7) and pyruvate kinase M1/2 (PKM), miR-122 negatively affects in vitro invasion, migration, and survival of hepatocellular carcinoma cells [216,217]. Additionally, the recovery of miR-122 levels efficiently blocks angiogenesis and metastasis in vivo [216]. These results suggest that miR-122 can be a beneficial candidate for miRNA-replacement therapy to manage hepatocellular carcinoma. It was also discovered that the administration of coptisine strikingly reduces cancer growth, together with an enhanced level of miR-122 in a mouse xenograft model of hepatocellular carcinoma [18] (Figure 1 and Table 4).
3.1.3. Indole-3-Carbinol
Tumor-suppressive miR-34 can be transcribed in a TP53-dependent manner, thereby mediating the function of TP53. By regulating numerous target genes, such as BCL-2, miR-34 substantially blocks cell proliferation and induces apoptotic cell death, as well as cellular senescence [218]. Indole-3-carbinol has been proven to possess anti-cancer properties by regulating cell proliferation, apoptosis, and angiogenesis [219]. In addition, an experimental observation shows that indole-3-carbinol induces cell cycle arrest and TP53-dependent upregulation of miR-34 in breast cancer cells harboring wild-type TP53 [20] (Figure 1 and Table 4).
3.1.4. Matrine
The detection of differentially expressed miRNAs identified that miR-22 is comparably less abundant in colorectal cancer tissues [220]. This miRNA plays a role in decelerating in vitro cell proliferation, migration, and in vivo cancer growth by targeting multiple oncogenes, such as hypoxia-inducible factor 1α (HIF-1α) and ELAV-like RNA binding protein 1 (ELAVL1) [220,221]. Matrine can facilitate cell cycle arrest and prohibit cell survival by enhancing miR-22 levels in colorectal cancer (Figure 1 and Table 4). Indeed, it was also noticed that miR-22 targets Erb-B2 receptor tyrosine kinase 3 (ERBB3) and MDS1 and EVI1 complex locus (MECOM) in colorectal cancer [43]. However, given that miR-22 can target various transcripts with different functions, it is unsurprising that miR-22 acts as either an oncogene or tumor suppressor in a cellular context-dependent manner [222].
3.1.5. Sanguinarine
Another miRNA transcriptionally activated by TP53 is miR-16 that has adverse effects on cell survival due to the direct regulation of BCL-2 [223]. Additionally, transcription factor AP-4 (TFAP4), which is involved in the c-Myc-mediated EMT process, was identified as a target of miR-16 [224]. Moreover, miR-16 deters migration and invasion by inhibiting insulin-like growth factor 1 receptor (IGF1R) expression in hepatocellular carcinoma [225]. Besides, the anti-cancer effects of sanguinarine, such as cell cycle arrest and apoptosis, are caused by TP53-induced miR-16 in hepatocellular carcinoma [131] (Figure 1 and Table 4).
3.2. MiRNAs, Organosulfur, and Phytosterol Compounds
3.2.1. Allicin
The role of miR-383-5p has been investigated in cancer. For example, it has been shown that miR-383-5p interferes with oncogenic factors, such as cellular the inhibitor of PP2A (CIP2A), which is known to stabilize c-Myc and sustain cancer progression [226,227,228]. Furthermore, recent evidence revealed that miR-383-5p is upregulated in allicin-treated gastric cancer cells, leading to the suppression of ERBB4-related oncogenic signaling such as PI3K/Akt [29] (Figure 1 and Table 4).
3.2.2. Sulforaphane
In addition to the regulation of oncogenic miRNAs (Section 2.2), it was also shown that sulforaphane demethylates the promoter of miR-9 and restores the expression of miR-9 in lung cancer [129]. This study showed that sulforaphane inactivates DNA methyltransferases and attenuates the levels of histone deacetylases (HDACs) [129] (Figure 1 and Table 4). Since both sulforaphane and miR-9 are recognized to sensitize cancer cells to radiotherapy [229,230], it is feasible that miRNAs modulated by sulforaphane may contribute to enhance the effects of radiotherapy on cancer.
3.2.3. β-Sitosterol-d-Glucoside
In breast cancer, miR-10a was determined to interrupt PI3K/Akt activation via targeting phosphatidylinositol-4,5-bisphosphate 3-kinase catalytic subunit alpha (PIK3CA), hence acting as a tumor-suppressive miRNA to suppress breast cancer progression [231]. In line with this, new findings indicated that β-sitosterol-d-glucoside upregulates miR-10a levels and displays anti-cancer activity through inactivating PI3K/Akt signaling [31] (Figure 1 and Table 4). Treatment with β-sitosterol-d-glucoside significantly diminishes cancer growth in mouse xenograft models of breast cancer [31].
3.3. MiRNAs and Phenolic Compounds
3.3.1. Apigenin, Astragalin, and Baicalein
Multiple miRNAs acting as tumor-suppressive factors are regulated by phytochemicals in hepatocellular carcinoma. Apigenin has been proven to sensitize hepatocellular carcinoma through downregulating the levels of autophagy-related 7 (ATG7) and nuclear factor erythroid 2-related factor 2 (NFE2L2, also known as NRF2) [33,152]. Autophagy generally interrupts apoptosis induction and supports the property of cancer stem cells, thereby limiting the effects of anti-cancer drugs [4]. In addition, NFE2L2 can confer doxorubicin resistance in cancer cells via upregulating drug efflux proteins, such as P-glycoprotein (P-gp) [232]. ATG7 and NFE2L2 are downregulated by apigenin and directly targeted by miR-520b and miR-101, respectively, in hepatocellular carcinoma. Indeed, these miRNAs are increased in apigenin-treated cells, further sensitizing cancer cells to doxorubicin [33,152] (Figure 2 and Table 4).
The mechanism by which astragalin controls cancer growth was recently investigated in hepatocellular carcinoma. Astragalin increases the levels of miR-125b and impedes cancer growth in vivo (Figure 2 and Table 4). Further evidence showed that miR-125b targets hexokinase 2 (HK2) [34], which supports cell survival and the tumorigenesis of hepatocellular carcinoma [233].
In addition, treatments with baicalein show anti-cancer activity and modulate the expression of various miRNAs in hepatocellular carcinoma. One of the upregulated miRNAs by baicalein is miR-3127-5p, and the overexpression of this miRNA leads to the inhibition of cell proliferation, along with the inactivation of PI3K/Akt signaling [35] (Figure 2 and Table 4). In another study, it was demonstrated that miR-3127-5p negatively regulates Ras/ERK signaling through targeting ABL proto-oncogene 1 (ABL1) [234], supporting that miR-3127-5p acts as a tumor-suppressive miRNA.
3.3.2. Brazilein
Brazilein has been supposed to exert an anti-cancer activity in cancer. Brazilein induces cell cycle arrest [37], represses invasion and migration [235], and exhibits synergistic anti-cancer effects with cisplatin in colorectal cancer cells [236]. Additionally, miR-133a is upregulated in vestibular schwannoma cells treated with brazilein (Figure 2 and Table 4). In their study, it was indicated that the activation of caspases by brazilein is abrogated in miR-133a-silencing cells, demonstrating the apoptosis-promoting potential of miR-133a [138].
3.3.3. Chrysin, Curcumin, and Genistein
Curcumin and genistein exert their anti-cancer effects through modulating miR-34 levels in breast cancer and head and neck cancer, respectively [45,133] (Figure 2 and Table 4). EMT is negatively controlled by miR-34 that targets EMT-triggering factors, such as zinc finger E-box binding homeobox 1 (ZEB1) and snail family transcriptional repressor 1 (SNAI1) in breast cancer [237]. Curcumin elevates miR-34 levels, and EMT-related genes repressed by curcumin is restored by miR-34 knockdown, indicating that miR-34 mediates the effects of curcumin on EMT [133].
In addition, genistein attenuates the stemness and EMT traits of head and neck cancer. In particular, miR-34 induced by genistein directly targets RNA 2′,3′-cyclic phosphate and 5′-OH ligase (RTCB), contributing to the suppression of stemness features, including self-renewal potential [45]. This study shows that the overexpression of RTCB weakens the genistein-mediated suppression of cancer stemness (Figure 2 and Table 4).
Besides, both chrysin and curcumin can stimulate the expression of miR-132 in breast cancer cells (Figure 2 and Table 4). Additionally, the combinatorial treatment of chrysin with curcumin using nanoparticles boosts miR-132 expression and shows synergistic anti-cancer effects [38]. Through targeting forkhead box A1 (FOXA1), miR-132 shows an anti-proliferation effect on breast cancer cells [238]. Further, miR-132 negatively regulates cell proliferation and the metastasis of breast cancer via targeting Jupiter microtubule associated homolog 1 (JPT1, also called HN1). In that study, it was shown that JPT1-depleted cancer cells poorly metastasize to the lungs [239].
3.3.4. Delphinidin, Epigallocatechin Gallate, and Gossypol
Delphinidin was addressed to impede invasion and migration of colorectal cancer cells without affecting proliferation status [40]. Further analyses indicated that delphinidin increases miR-204-3p expression and diminishes the levels of integrins and the activation of focal adhesion kinase (FAK) signaling (Figure 2 and Table 4). The knockdown of miR-204-3p reverses the effects of delphinidin on cell invasion, integrin levels, and FAK activity. These results indicate that miR-204-3p plays a critical role in regulating signaling pathways following delphinidin treatments [40].
Epigallocatechin gallate (EGCG) has been demonstrated to suppress the migration and stemness of cancer cells by altering miRNA expression. In nasopharyngeal carcinoma, EGCG upregulates the levels of miR-296-3p that is involved in the inactivation of STAT3, resulting in the impairment of migratory property of anoikis-resistant cells [41]. Moreover, EGCG can upregulate miR-485, which targets CD44 in cisplatin-resistant lung cancer cells, thus weakening lung cancer stemness. The in vivo administration of EGCG also shows the reduction of cancer growth and the levels of stemness factors, such as SRY-box transcription factor 2 (SOX2), Octamer-binding protein 4 (OCT4), and Nanog homeobox (NANOG) [151] (Figure 2 and Table 4).
Gossypol directly interacts with and inhibits anti-apoptotic factors, BCL-2 and BCL-2-like protein 1 (BCL2L1, also named BCL-XL), enacting apoptotic cell death in cancer [46,240]. Besides, it was uncovered that gossypol induces apoptosis and upregulates miR-15a, a BCL-2-targeting miRNA, in pituitary cancer cells [130], suggesting that gossypol can also indirectly limit BCL-2 activity (Figure 2 and Table 4).
3.3.5. Hydroxygenkwanin, Isorhapontigenin, and Kaempferol
Forkhead Box M1 (FOXM1) is an EMT-promoting factor in cancer, and its knockdown hampers the invasion and migration of cancer cells [241,242]. A tumor-suppressive miRNA, miR-320a, has been known to inhibit hepatocellular carcinoma progression via targeting c-Myc and β-catenin [243,244]. Further evidence suggested that miR-320a induced by hydroxygenkwanin directly inhibits FOXM1 expression and mediates the EMT-suppressive effects of hydroxygenkwanin [47] (Figure 2 and Table 4).
Isorhapontigenin can elicit anti-cancer effects via increasing miR-137 and miR-145 [51,140,144]. It was shown that miR-137 targets SP1 and glycogen synthase kinase 3 beta (GSK3β) in bladder cancer and urothelial cancer, respectively. Thus, miR-137 suppresses anchorage-independent growth of bladder cancer and invasion of urothelial cancer [51,140]. Furthermore, isorhapontigenin escalates miR-145 levels and suppresses the sphere formation and anchorage-independent growth of patient-derived glioblastoma [144]. An additional analysis showed that miR-145 directly targets SOX2, which is involved in cyclin D1 transcription [144] (Figure 2 and Table 4).
Kaempferol was known to cause cell cycle arrest, autophagic cell death, and the inactivation of oncogenic signaling such as PI3K/Akt in cancer [245,246]. A further study on the molecular mechanism of kaempferol showed that miR-340 can be induced by kaempferol and that miR-340 knockdown abolishes the inactivation of PI3K/Akt in kaempferol-treated lung cancer cells [52] (Figure 2 and Table 4). Based on the clue that miR-340 targets sirtuin 7 (SIRT7) [247], an activator of Akt [248], it can be assumed that kaempferol negatively affects PI3K/Akt partly via the miR-340/SIRT7 axis.
3.3.6. Licochalcone A and Luteolin
Licochalcone A has shown to effectively inhibit cell migration as well as EMT and activate caspases [53]. In lung cancer, licochalcone A increases miR-144-3p expression and causes apoptosis induction [143] (Figure 2 and Table 4). In this study, it was also demonstrated that miR-144-3p targets nuclear factor erythroid 2-related factor (NFE2L2, also referred to as NRF2) [143], which can activate cytoprotective autophagy to inhibit apoptotic cell death in lung cancer [249]. In line with this, miR-144-3p has been realized to inhibit SRC proto-oncogene (SRC), thereby blocking TGF-β-induced invasion [250]. By targeting c-Met, miR-144-3p also induces apoptosis to inhibit cell survival [251].
It has been investigated that luteolin also increases tumor-suppressive miRNAs to carry out its anti-cancer activity. For example, miR-34 is related to the mechanism of action of luteolin (Figure 2 and Table 4). In lung cancer, miR-34 induced by luteolin can mediate the anti-proliferative and pro-apoptotic effects of luteolin on lung cancer cells by directly controlling the expression of mouse double minute 4 (MDM4), a repressor of TP53 [54].
Pleiotrophin (PTN) can interact with neuropilin-1 receptors, stimulating downstream signaling pathways, such as PI3K and focal adhesion kinase (FAK), to facilitate cancer growth and metastasis [252]. It was suggested that miR-384 augmented by luteolin targets PTN in colorectal cancer cells [150], suggesting that luteolin suppresses colorectal cancer metastasis, in part, via the miR-384/PTN axis. Moreover, in hepatocellular carcinoma, miR-6809-5p was noticed to be elevated by luteolin, contributing to the suppression of cancer growth [154]. This study showed that miR-6809-5p represses cancer growth via targeting flotillin-1 [154], which can activate oncogenic factors such as NF-κB [253] (Figure 2 and Table 4).
3.3.7. Physcion 8-O-β-Glucopyranoside and Quercetin
RalA-binding protein 1 (RALBP1, also called RLIP76) is a multifaceted drug transporter that mediates therapeutic resistance. Also, RALBP1 can facilitate cancer growth, angiogenesis, invasion, and metastasis [254,255,256,257]. A recent investigation showed that physcion 8-O-β-glucopyranoside (PG) raises the levels of miR-124, which has been validated as a tumor-suppressive miRNA in several cancer types [136] (Figure 2 and Table 4). RALBP1 can be targeted by miR-124 [136], implying a possibility that PG may sensitize cancer cells to anti-cancer agents whose efflux is catalyzed by RALBP1.
Quercetin is capable of affecting several signaling pathways that contribute to cell cycle progression, angiogenesis, and cell survival. For example, quercetin can inhibit the expression of heterogeneous nuclear ribonucleoprotein A1 (hnRNPA1), which controls mRNA exports of anti-apoptotic genes, thereby enhancing other anti-cancer agents, such as enzalutamide and JQ1 [258,259]. Moreover, recent studies pointed out that quercetin suppresses the stemness of pancreatic cancer [58,148]. The treatment of pancreatic cancer cells with quercetin leads to an increase in let-7c and NUMB endocytic adaptor protein (NUMB) levels [58] (Figure 2 and Table 4). In this study, it was proposed that the transcription of NUMB, a Notch inhibitory factor, is activated by let-7c, subsequently inhibiting the expression of Notch and stemness factors in pancreatic cancer cells. Since it was confirmed that NUMB is transcriptionally and post-transcriptionally inhibited by high mobility group AT-hook 1 (HMGA1) [260] and that Wnt/β-catenin signaling, which can increase HMGA1 levels, is repressed by let-7c [261,262], quercetin may affect Notch and stemness factor expression via the let-7c/Wnt/β-catenin/HMGA1/NUMB axis. Likewise, miR-200 induced by quercetin can contribute to repress the stemness property of pancreatic cancer cells via directly targeting Notch1 [148].
3.3.8. Resveratrol and Trans-3,5,4′-trimethoxystilbene
In breast cancer, resveratrol upregulates miR-34, miR-424, and miR-503 levels in a TP53-dependent way [134] (Figure 2 and Table 4). In that study, it was shown that the downregulation of hnRNPA1 by resveratrol is directly mediated by miR-424 and miR-503. In the case of miR-34, this miRNA also indirectly represses hnRNPA1 expression [134]. Since miR-34 was reported to downregulate c-Myc, a positive regulator of hnRNPA1 expression [263,264], resveratrol may reduce hnRNPA1 levels partly via the miR-34/c-Myc axis.
Resveratrol also increases the levels of other miRNAs, thereby enhancing the chemosensitivity and inhibiting the proliferation of breast cancer cells. Resveratrol can re-sensitize resistant cells to doxorubicin by upregulating miR-122 and downregulating BCL-2 levels [135]. Since miR-122 can negatively regulate the transcription of BCL-2 via suppressing c-Myc [265], resveratrol may regulate BCL-2 levels partly via the miR-122/c-Myc axis. In addition, resveratrol can show an anti-proliferation effect by upregulating both miR-663 and miR-744, which target eukaryotic translation elongation factor 1 alpha 2 (EEF1A2) [153] (Figure 2 and Table 4).
It was also addressed that resveratrol exerts apoptosis induction and EMT suppression by upregulating miR-139-5p and miR-200, respectively (Figure 2 and Table 4). In osteosarcoma, resveratrol induces apoptosis via upregulating miR-139-5p, which targets Notch1 [141]. Resveratrol also upregulates miR-200, thereby blocking EMT progression and inducing apoptosis in colorectal cancer cells [149].
A recent study demonstrated that trans-3,5,4′-trimethoxystilbene (TMS), which is a derivative of resveratrol, sensitizes lung cancer cells to gefitinib (a selective EGFR tyrosine kinase inhibitor). The expression of miR-345 and miR-498 is significantly downregulated in gefitinib-resistant cells, and the treatment of TMS can upregulate the levels of these miRNAs. In addition, the overexpression of miR-345 and miR-498 suppresses c-Fos (FOS) and BCL-2, respectively. Further investigation showed that miR-345 and miR-498 target mitogen-activated protein kinase 1 (MAPK1) and phosphoinositide-3-kinase regulatory subunit 1 (PIK3R1), respectively, in lung cancer cells [65] (Figure 2 and Table 4).
3.3.9. Silymarin
Silymarin attenuates the migration of lung cancer cells, together with the repression of HDAC activities and ZEB1 levels [64]. Further, it was demonstrated that silymarin restores the expression of miR-203, which is negatively regulated by ZEB1 [64] (Figure 2 and Table 4). These results suggest that miR-203 is one of the factors mediating the effects of silymarin on cancer cells. Indeed, it has been demonstrated that miR-203 shows its tumor-suppressive effects on lung cancer via enhancing let-7 maturation and targeting oncogenes such as SMAD3 [266,267].
3.4. MiRNAs and Terpenoid Compounds
3.4.1. Ailanthone and Andrographolide
Ailanthone effectively restrains the proliferation, invasion, and migration of cancer cells through increasing miR-148a and miR-449a levels in breast cancer and acute myeloid leukemia cells, respectively [66,145] (Figure 3 and Table 4). In particular, miR-148a suppresses Wnt/β-catenin signaling in breast cancer [145]. In the case of miR-449a, this miRNA is involved in the restriction of Notch and PI3K/Akt signaling pathways in ailanthone-treated acute myeloid leukemia cells [66].
B lymphoma Mo-MLV insertion region 1 homolog (BMI1) advances the EMT and stemness of cancer cells via promoting NF-κB-mediated NANOG expression [268]. Recently, it was presented that andrographolide suppresses oral cancer stemness and sensitizes cancer cells to radiotherapy. The mechanism underlying the anti-cancer activity of andrographolide involves the upregulation of miR-218 that directly regulates BMI1 [67] (Figure 3 and Table 4).
3.4.2. Artemisinin and Artesunate
As stated in Section 3.1.3, indole-3-carbinol upregulates miR-34 via wild-type TP53. In that study, both artemisinin and artesunate were also shown to increase miR-34 expression (Figure 3 and Table 4). The difference between indole-3-carbinol and artemisinin/artesunate is that miR-34 is increased by artemisinin and artesunate in a TP53-independent manner [20]. While further investigations are needed, it is plausible that artemisinin and artesunate may regulate miR-34 by inhibiting c-Myc, a repressor of miR-34 [68,218]. In the case of artesunate, this compound also positively regulates both miR-133a and miR-206 through ROS production in rhabdomyosarcoma cells [137] (Figure 3 and Table 4). Both miRNAs have been recognized as tumor suppressors and retard the proliferation of rhabdomyosarcoma cells [269,270].
3.4.3. Astragaloside IV and Cannabidiol
Astragaloside IV subdues cancer progression via regulating multiple factors. Astragaloside IV suppresses the levels of VEGF and MMPs. PI3K/Akt signaling can also be blocked by astragaloside IV [271]. Moreover, it was demonstrated that astragaloside IV blocks the phenotype conversion of macrophages into M2-like tumor-associated macrophages (TAMs) by suppressing AMP-activated protein kinase (AMPK) activation in macrophages, thus inhibiting cancer progression [69]. In addition, astragaloside IV can upregulate the levels of miR-134, which targets CAMP-responsive element-binding protein 1 (CREB1), consequently suppressing EMT and sensitizing cancer cells to oxaliplatin [139] (Figure 3 and Table 4).
Cannabidiol activates apoptosis in neuroblastoma cells. The levels of numerous miRNAs are altered by cannabidiol treatments, and one of the upregulated miRNAs is miR-1972 (Figure 3 and Table 4). Further investigation into the relationship between apoptosis and miR-1972 showed that anti-apoptotic factors, BCL2L1 and SIRT2, are targeted by miR-1972, contributing to apoptosis induction [71].
3.4.4. Cucurbitacin D, Curcumol, and Lycopene
Cucurbitacin D effectively shows anti-cancer activity toward cervical cancer. Both cell cycle arrest and apoptosis are induced by cucurbitacin D. Additionally, the expression of STAT3, c-Myc, and MMP9 are inhibited by cucurbitacin D. In addition, cucurbitacin D decelerates the growth of cervical cancer in vivo. Moreover, the levels of tumor-suppressive miRNAs, such as miR-34, miR-143, and miR-145, are enhanced by cucurbitacin D, suggesting that these miRNAs act as signaling mediators in cucurbitacin D-treated cells [73] (Figure 3 and Table 4).
ATP-binding cassette subfamily C member 3 (ABCC3) is one of the members of ABC transporter subfamily and is able to confer doxorubicin resistance in cancer cells by limiting intracellular concentrations of doxorubicin [74]. It was ascertained that miR-181-3p is upregulated by curcumol and that this miRNA targets ABCC3, eventually improving the efficacy of doxorubicin in breast cancer [74] (Figure 3 and Table 4). Since several anti-cancer agents are substrates for ABCC3, screening of other therapeutic agents that can be sensitized by curcumol will be beneficial to establish the new therapeutic strategy.
3.4.5. Oleanolic Acid and Pristimerin
Myocyte enhancer factor 2D (MEF2D) is a transcription factor that activates cell proliferation and migration in various cell types. For example, the knockdown of MEF2D leads to impaired proliferation and the migration of lung cancer cells [272]. Furthermore, it was noticed that the treatment of lung cancer cells with oleanolic acid upregulates miR-122 and downregulates two target mRNAs of this miRNA, namely MEF2D and cyclin G1, resulting in the suppression of cell proliferation [79] (Figure 3 and Table 4).
In breast cancer, pristimerin was observed to repress cell viability, cell cycle progression, and migration. Additional evidence demonstrated that pristimerin increases miR-542-5p levels and that this miRNA directly targets ubiquitin-specific-processing protease 17-like protein 2 (USP17L2, also referred to as DUB3) [81], which promotes migration, invasion, and metastasis of breast cancer by stabilizing SNAI1 [273] (Figure 3 and Table 4).
3.4.6. Toosendanin and Triptolide
Toosendanin impedes cell proliferation, migration, invasion, and TGF-β-induced EMT. In vivo administration of toosendanin leads to the reduction of cancer growth and liver metastasis in an orthotopic implantation model of gastric cancer [82]. The molecular mechanisms of toosendanin involve miR-200, which is upregulated by toosendanin and directly targets β-catenin [82] (Figure 3 and Table 4).
In the case of triptolide, it was demonstrated that PI3K/Akt signaling is inhibited by triptolide. An increment of miR-193-3p expression by triptolide causes the downregulation of kruppel-like factor 4 (KLF4) [147], which acts as an upstream activator of PI3K/Akt signaling [274] (Figure 3 and Table 4).
3.4.7. Tubeimoside-1 and Ursolic Acid
It was recently denoted that miR-126 mediates the anti-cancer effects of tubeimoside-1 by directly controlling the expression of vascular endothelial growth factor A (VEGFA) in lung cancer cells [84]. Through negatively regulating VEGFA, tubeimoside-1 inactivates ERK signaling, consequently hindering cell viability and metastasis of lung cancer cells [84] (Figure 3 and Table 4).
Myeloid differentiation primary response 88 (MyD88) is responsible for cell survival, DNA repair, and maintenance of stemness, thereby contributing to therapeutic resistance in cancer [275,276]. It was disclosed that ursolic acid reverses paclitaxel resistance by downregulating MyD88 levels in lung and breast cancer [85,146]. These studies also confirmed that miR-149-5p is upregulated in ursolic acid-treated cells and negatively affects MyD88 levels (Figure 3 and Table 4).
4. MiRNAs Affecting the Anti-Cancer Activity of Phytochemicals Currently Tested in Preclinical Studies and Clinical Trials
4.1. 1′S-1′-Acetoxychavicol Acetate
While cell survival is repressed by 1′S-1′-acetoxychavicol acetate (ACA), the induction of cytoprotective autophagy can be triggered by ACA in cancer cells [32]. The inhibition of autophagy augments the efficacy of ACA [32], suggesting that autophagy-regulating miRNAs can influence the effectiveness of ACA. In cervical cancer, it was noticed that the knockdown of either miR-210 or miR-629 sensitizes cells to ACA by upregulating SMAD family member 4 (SMAD4) or Ras suppressor protein 1 (RSU1), respectively. Additional findings showed that the pro-apoptotic effects of ACA are potentiated by the overexpression of either SMAD4 or RSU1 [277,278] (Figure 4 and Table 5).
4.2. Apigenin
As stated in Section 3.3.1, apigenin upregulates tumor-suppressive miRNAs and enhances the efficacy of doxorubicin. Besides, it was proven that a miRNA regulates the efficacy of apigenin. The knockdown of TERT leads to an increment of miR-138 levels in neuroblastoma cells. Furthermore, the overexpression of miR-138 augments apoptosis following apigenin treatments [283] (Figure 4 and Table 5). In addition, the anti-cancer effects of apigenin are more enhanced in miR-138-overexpressing cells than in TERT-silencing cells, implying that miR-138 can downregulate multiple oncogenic factors, which block the efficacy of apigenin [283].
4.3. Delphinidin
The expression of miR-137 is epigenetically silenced in glioblastoma, and the overexpression of miR-137 blocks the invasion of cancer cells, indicating that miR-137 is a tumor-suppressive miRNA in glioblastoma [282]. Apoptosis induced by delphinidin is effectively intensified by miR-137 overexpression. Further investigation showed that the combination of miR-137 and delphinidin remarkably suppresses various cellular factors involved in cellular survival, invasion, growth, and angiogenesis [282] (Figure 4 and Table 5).
4.4. Epigallocatechin Gallate
It was illustrated that the introduction of miR-126 sensitizes lung cancer cells to doxorubicin [284]. As mentioned in Section 3.4.7, miR-126 targets VEGFA. Moreover, miR-126 can target CFLAR, reversing TRAIL resistance in cervical cancer [285]. In addition, miR-126 augments epigallocatechin gallate-induced apoptosis in osteosarcoma cells [281], indicating a potential role of miR-126 in alleviating therapeutic resistance, especially in solid cancer (Figure 4 and Table 5). However, miR-126 acts as an oncogenic miRNA in acute myeloid leukemia. By maintaining the quiescence of leukemia stem cells, miR-126 can contribute to the emergence of therapeutic resistance to daunorubicin [286].
4.5. Luteolin and Silibinin
The efficacy of miR-7-3p was estimated in combination with luteolin or silibinin in glioblastoma. It was shown that the anti-growth effects of both luteolin and silibinin are significantly increased by the co-administration of miR-7-3p in vivo [279] (Figure 4 and Table 5). While it remains to be elucidated how miR-7-3p enhances the sensitivity of cancer cells to luteolin and silibinin, miR-7-3p is recognized to effectively reduce the level of anti-apoptotic factors, such as BCL-2 and X-linked inhibitor of apoptosis (XIAP) [279].
4.6. Oridonin
Oridonin shows its cytotoxic effects via downregulating miR-17 and miR-20a (Section 2.4.4). Moreover, it was noticed that the knockdown of miR-17 or miR-20a increases apoptosis following oridonin treatments, even at a low concentration [80] (Figure 4 and Table 5). These results suggest the possibility that miRNA-based therapy is attractive to improve the therapeutic efficacy of phytochemicals.
4.7. Physcion 8-O-β-Glucopyranoside
In ovarian cancer, physcion 8-O-β-glucopyranoside (PG) adequately exhibits its anti-cancer activities by inhibiting the growth, invasion, and migration of cancer cells. However, the overexpression of miR-25 was found to attenuate the effects of PG on ovarian cancer cells [280] (Figure 4 and Table 5). In line with this, miR-25, one of the miR-106b–25 cluster members, was reported to promote ovarian cancer proliferation and downregulates a pro-apoptotic factor, BCL2L11 [287].
4.8. Shikonin
Several studies presented that shikonin exerts its anti-cancer effects by inducing cell cycle arrest, mitochondrial dysfunction, apoptosis, and necroptosis [288,289,290]. While shikonin can effectively constrain the growth of lung cancer in vivo, cytoprotective autophagy is also induced by shikonin treatments [289]. Indeed, the pharmacological inhibition of autophagy augments the efficacy of shikonin [289]. It was also reported that miR-143, a tumor-suppressive miRNA, is downregulated in glioblastoma stem cells treated with shikonin [60]. It was further remarked that the ectopic introduction of miR-143 directly targets BCL2-binding athanogene 3 (BAG3), an anti-apoptotic factor, and enhances the cytotoxicity of shikonin [60] (Figure 4 and Table 5). These data suggest that it is required to overcome acquired resistance to shikonin for strengthening the therapeutic efficacy of shikonin.
5. MiRNAs Regulating the Sensitivity of Cancer Cells to Phytochemicals Currently Used in Cancer Therapy
5.1. Etoposide
5.1.1. MiRNAs Regulating Apoptosis and Autophagy
Recent studies demonstrated that miRNAs control the efficacy of etoposide by modulating cellular factors involved in apoptosis and cytoprotective autophagy. Etoposide resistance is advanced by miR-21 and miR-374a [291,292]. Cytoprotective autophagy is activated by miR-21; therefore, etoposide-induced apoptosis is abated by miR-21 in colorectal cancer cells [291]. In glioblastoma, miR-374a suppresses FOXO1 expression, thus blocking apoptosis induced by etoposide. The knockdown of miR-374a sensitizes cancer cells to etoposide by promoting the expression of FOXO1 [292] (Figure 5 and Table 6).
By contrast, tumor-suppressive miRNAs, such as miR-29-3p, miR-193-3p, and miR-196-5p, have been reported to target anti-apoptotic mRNAs and enhance the cytotoxicity of etoposide, showing the following: miR-29-3p sensitizes cervical cancer cells to etoposide by targeting myeloid cell leukemia sequence 1 (MCL1), a member of the BCL-2 family [293]; miR-193-3p targets insulin receptor substrate 2 (IRS2) and improves the effectiveness of etoposide in osteosarcoma cells [294]; miR-196-5p escalates etoposide-induced apoptosis via inhibiting insulin-like growth factor 2 mRNA-binding protein 1 (IGF2BP1) [295] (Figure 5 and Table 6).
5.1.2. MiRNAs Regulating EMT and Wnt/β-Catenin Signaling
The efficacy of etoposide can be regulated by miRNAs that control EMT and Wnt/β-catenin signaling. In lung cancer cells, both miR-192 and miR-662 potentiate cell invasion and anchorage-independent growth. Additionally, the expression of genes that activate EMT and Wnt/β-catenin signaling is enhanced by these miRNAs, hence contributing to the appearance of etoposide resistance [301] (Figure 5 and Table 6).
5.1.3. MiRNAs Regulating DNA Damage Repair
It was recognized that miR-302 directly targets DNA repair protein RAD52 homolog (RAD52) in leukemia cells [315]. Since one of the causes of therapeutic resistance is DNA damage repair pathways, the overexpression of miR-302 can sensitize cells to etoposide via repressing RAD52, which generally limits the efficacy of cancer therapies [315,326] (Figure 5 and Table 6).
5.1.4. A miRNA Regulating a Drug Transporter Level
ABCG2 (also referred to as breast cancer resistance protein (BCRP)) is responsible for therapeutic resistance by mediating the efflux of several anti-cancer agents, including etoposide [327]. It was recently reported that miR-3163 decelerates the proliferation of retinoblastoma cancer stem cells (RCSC), indicating that this miRNA has a tumor-suppressive property. Also, the anti-proliferative effects of etoposide are enhanced in miR-3163-overexpressing RCSC. Indeed, this miRNA was confirmed to target ABCG2 [323] (Figure 5 and Table 6).
5.2. Irinotecan
5.2.1. MiR-200
As described in Section 3.3 and Section 3.4, miR-200 suppresses cell viability, EMT, and stemness, suggesting that this miRNA can be used as a resistance-suppressive miRNA. In fact, it was shown that the combined treatment of irinotecan and miR-200 forcefully restrict the growth of colorectal cancer with a minimized systemic toxicity in mouse xenograft models [313] (Figure 5 and Table 6).
5.2.2. MiR-514b-5p
The metastasis of colorectal cancer is facilitated by miR-514b-5p. Functional experiments showed that miR-514b-5p triggers EMT and invasion of cancer cells via targeting cadherin 1 (CDH1, also named E-cadherin 1) and claudin 1 (CLDN1). Further, it was shown that the lentiviral delivery of miR-514b-5p decreases the efficacy of irinotecan in mouse xenograft models [302] (Figure 5 and Table 6).
5.2.3. MiR-627
Irinotecan is metabolized into inactive forms by cytochrome P450 family 3 subfamily A member 4 (CYP3A4) [328]. A recent study demonstrated that miR-627 can directly target CYP3A4, thereby enhancing the effects of irinotecan on growth inhibition and apoptosis induction in colorectal cancer cells [15] (Figure 5 and Table 6). In this study, it was also shown that calcitriol, a synthetic vitamin D, reduces CYP3A4 levels via upregulating miR-627 and enhances the anti-cancer efficacy of irinotecan.
5.2.4. MiR-4454
Moreover, it was recently found that miR-4454 expression is repressed in irinotecan-resistant colorectal cancer cells [324]. Further analyses indicated that the restoration of miR-4454 effectively sensitizes cells to irinotecan, hence obstructing cell growth, migration, and invasion of irinotecan-resistant cells (Figure 5 and Table 6). A mechanism underlying miR-4454-mediated sensitization of cancer cells involves the downregulation of guanine nucleotide-binding protein-like 3-like protein (GNL3L), an activator of NF-κB transcription activity [324].
5.3. Paclitaxel
5.3.1. MiRNAs Negatively Regulating Apoptosis
Multiple apoptosis-inhibiting miRNAs weaken paclitaxel activity toward cancer cells. In breast cancer, miR-21 and miR-125b target programmed cell death 4 (PDCD4) and BCL2-antagonist/killer 1 (BAK1), respectively, eventually aggravating paclitaxel resistance [296,299]. In addition, leucine zipper tumor suppressor 1 (LZTS1), a repressor of PI3K/Akt signaling [329], is known to be targeted by miR-1207-5p. Therefore, miR-1207-5p is capable of diminishing paclitaxel cytotoxicity toward breast cancer cells [304] (Figure 5 and Table 6).
Moreover, miR-27a-3p is increased by the hypoxic condition and advances resistance to paclitaxel by targeting apoptotic protease-activating Factor 1 (APAF1), a mediator of cytochrome c-dependent caspase activations, in ovarian cancer cells [297] (Figure 5 and Table 6).
The sensitivity of cancer cells to paclitaxel can be abolished by PTEN inhibition. In chordoma, both miR-140-3p and miR-155-5p contribute to exacerbating paclitaxel resistance via directly targeting PTEN [300]. It was also noted that miR-4262 expression is intensively augmented in paclitaxel-resistant lung cancer cells compared to paclitaxel-sensitive parental cells. By directly blocking PTEN, miR-4262 reduces the efficacy of paclitaxel. The co-treatment with paclitaxel and miR-4262 inhibitors synergistically inhibits the growth of cancer, along with the inactivation of Akt and GSK3β in xenograft models in which paclitaxel-resistant cells were subcutaneously injected [305] (Figure 5 and Table 6).
OTU domain-containing protein 3 (OTUD3) is recognized as a deubiquitinase of PTEN, thus acting as a tumor suppressor by stabilizing PTEN. By targeting OTUD3, miR-520h can contribute to paclitaxel resistance via indirectly suppressing PTEN. As a matter of fact, the silencing of miR-520h re-sensitizes resistant cells to paclitaxel by augmenting apoptosis induction [303] (Figure 5 and Table 6).
5.3.2. MiRNAs Positively Regulating Apoptosis
In breast cancer, apoptosis following paclitaxel treatments is boosted by miR-7-5p, miR-542-3p, miR-621, and miR-5195-3p via directly modulating BCL-2, survivin, F-box protein 11 (FBXO11), and eukaryotic translation initiation factor 4A2 (EIF4A2), respectively [307,321,325]. The paclitaxel-sensitizing effects of miR-7-5p are reversed by the restoration of BCL-2 in breast cancer cells [307]. By targeting survivin, miR-542-3p potentiates anti-cancer effects of paclitaxel in vitro and in vivo [319]. In addition, miR-621 increases TP53 activity by repressing FBXO11. Paclitaxel-induced apoptosis in miR-621-overexpressing cells is attenuated by the upregulation of FBXO11 [321]. In the case of miR-5195-3p, the expression of this miRNA is downregulated in paclitaxel-resistant cell lines and tissues derived from breast cancer patients. The overexpression of EIF4A2 abrogates apoptosis induction in paclitaxel-resistant cells [325] (Figure 5 and Table 6).
It has been demonstrated that integrin-mediated signaling impairs paclitaxel-induced apoptosis via inhibiting cytochrome c release [330]. A recent study uncovered that integrin β1 (ITGB1) and miR-29-3p levels are increased and decreased, respectively, in paclitaxel-resistant nasopharyngeal cancer cells. The overexpression of miR-29-3p re-sensitizes resistant cells to paclitaxel by repressing ITGB1. The knockdown of miR-29-3p was found to facilitate the growth of paclitaxel-resistant cells in vivo, indicating that miR-29-3p plays a critical role in regulating the effectiveness of paclitaxel [308] (Figure 5 and Table 6).
Tripartite motif-containing protein 27 (TRIM27) inhibits PTEN, hence activating PI3K/Akt signaling. The knockdown of TRIM27 is known to induce apoptosis in cancer cells [331]. TRIM27 is overexpressed in ovarian cancer tissues and correlated with the unfavorable prognosis of patients with ovarian cancer [317]. It was also noticed that miR-383-5p is downregulated in ovarian cancer tissues and is able to target TRIM27. The overexpression of miR-383-5p enhances paclitaxel efficacy via increasing apoptosis induction [317] (Figure 5 and Table 6).
5.3.3. MiRNAs Inhibiting EMT- and Stemness-Related Factors
Notch signaling is known to induce and maintain the stemness of cancer cells. Enhancer of zeste homolog 2 (EZH2) activates Notch signaling, thereby expanding cancer stem cell populations [332]. In addition, proto-oncogene c-Fos (FOS) promotes EMT and the expression of several stemness factors, such as Notch1 and SOX2 [333]. Both EZH2 and FOS were validated as target mRNAs of miR-365, and the upregulation of miR-365 inhibits cell invasion, migration, and survival. Additionally, the sensitivity of cells to paclitaxel is enhanced by miR-365 in endometrial cancer [316] (Figure 5 and Table 6).
SOX2, a stemness factor, can promote EMT through activating STAT3 signaling and maintain self-renewal potential [334]. SOX2 is directly regulated by miR-145-5p in breast cancer [312]. Both miR-145-5p overexpression and SOX2 knockdown reverse paclitaxel resistance. The efficacy of paclitaxel in miR-145-5p-overexpressing cells is abrogated by the restoration of SOX2 levels [312] (Figure 5 and Table 6).
5.3.4. MiRNAs Regulating the Level of Drug Transporters
In addition to BCL-2 (Section 5.3.2), miR-7-5p was also investigated to target ABCC1 (also called multidrug resistance protein 1, MRP1) [307]. These results demonstrated that miR-7-5p can sensitize cancer cells to paclitaxel via augmenting both pro-apoptotic pathways and intracellular paclitaxel levels (Figure 5 and Table 6).
As mentioned above, miR-27a-3p acts as an anti-apoptotic factor (Section 5.3.1). In addition to this role, miR-27a-3p was found to be upregulated in paclitaxel-resistant ovarian cancer cells. By targeting homeodomain-interacting protein kinase-2 (HIPK2), miR-27a-3p can indirectly stimulate P-gp levels, leading to paclitaxel resistance. The knockdown of miR-27a-3p makes paclitaxel-resistant cells more susceptible to paclitaxel [298] (Figure 5 and Table 6).
5.4. Vincristine
5.4.1. MiRNAs Regulating Apoptosis and Autophagy
The expression of miR-200 and miR-429 is downregulated in vincristine-resistant gastric cancer cells. These miRNAs are able to augment the induction of apoptosis following the treatment of resistant cells with vincristine. It was further pointed out that BCL-2 and XIAP are targeted by both miRNAs, suggesting that the efficiency of vincristine can be impeded by the alteration of miR-200 and miR-429 levels [314] (Figure 5 and Table 6).
The anti-cancer activity of vincristine can be augmented by autophagy-inhibiting miRNAs. In gastric cancer, miR-495-3p inhibits cytoprotective autophagy via targeting heat shock protein 70 family protein 5 (HSPA5, also called GRP78) [318]. The inhibition of HSPA5 by miR-495-3p suppresses autophagy via triggering mTOR signaling, improving the cytotoxicity of vincristine. Moreover, the lentiviral delivery of miR-495-3p efficiently decreases the growth of vincristine-resistant cells in vivo [318]. In addition, miR-874 inhibits autophagy via targeting ATG16L1, a positive regulator of autophagosome formation [322]. Both miR-874 overexpression and ATG16L1 knockdown elevate the efficacy of vincristine in gastric cancer cells [322] (Figure 5 and Table 6).
5.4.2. MiRNAs Inhibiting EMT- and Stemness-Related Factors
As stated in Section 5.1.2, the miR-34/MAGE-A axis affects etoposide cytotoxicity. In that study, it was also confirmed that the sensitivity of cells to vincristine is significantly enhanced and diminished in miR-34-overexpressing and -silencing retinoblastoma cells, respectively [309] (Figure 5 and Table 6).
In addition to miR-365 (see Section 5.3.3), EZH2 is also directly regulated by miR-126 in gastric cancer cells. It was remarked that the level of miR-126 is diminished in vincristine-resistant cells and that miR-126 overexpression can re-sensitize resistant cells to vincristine. The sensitization of cells to vincristine is also thoroughly observed in EZH2 silencing cells [310] (Figure 5 and Table 6).
The growth of medulloblastoma is known to be inhibited by miR-584-5p. In addition, the overexpression of miR-584-5p was identified to suppress the self-renewal of medulloblastoma cells owing to its ability to target EIF4E3 and HDAC1. Certainly, the silencing of EIF4E3 and HDAC1 also represses the self-renewal capacity of medulloblastoma. Further, the ectopic expression of miR-584-5p intensifies the anti-cancer efficacy of vincristine [320] (Figure 5 and Table 6).
5.4.3. MiRNAs Regulating the Intracellular Concentration of Vincristine
Sorcin was proven to induce the expression of P-gp and ABCC1, thus contributing to therapeutic resistance [335,336]. Notably, it was demonstrated that there is a negative correlation between sorcin and miR-1 levels in vincristine-resistant gastric cancer cells [306]. The levels of P-gp and ABCC1 are markedly upregulated in vincristine-resistant cells. The overexpression of miR-1 downregulates sorcin, consequently inhibiting the levels of P-gp and ABCC1 as well. Due to this ability, miR-1 can reverse vincristine-resistant in gastric cancer [306] (Figure 5 and Table 6).
Furthermore, ABCC1 is directly targeted by miR-133b. The intracellular concentration of vincristine is increased by miR-133b in colorectal cancer cells. Therefore, the inhibitory effects of vincristine on colony formation, cell survival, and cancer growth are significantly advanced by miR-133b [311] (Figure 5 and Table 6).
By suppressing ABCG2, miR-3163 sensitizes retinoblastoma cells not only to etoposide, but also to vincristine (also see Section 5.1.4). It was also validated that miR-3163 can enhance the efficacy of other anti-cancer agents, such as cisplatin, suggesting that miR-3163 plays a significant role in sensitizing cancer cells to therapeutic agents [323] (Figure 5 and Table 6).
6. Conclusions
The accumulating evidence presented here indicates that phytochemicals have valuable potentials as therapeutic agents against cancer. Single treatments of phytochemicals effectively regulate cellular signaling pathways to exert their anti-cancer activities. Additionally, phytochemicals sensitize cancer cells to other conventional drugs, such as doxorubicin, indicating a great possibility of their use in combination with other anti-cancer agents to maximize therapeutic responses for managing cancer.
Various anti-cancer activities of phytochemicals are mediated by altering the expression of miRNAs in cancer cells. As expected, oncogenic and tumor-suppressive miRNAs are generally downregulated and upregulated, respectively, by phytochemicals. This modulation of miRNA expression by phytochemicals plays a critical part in regulating the levels of cellular factors and restoring drug sensitivity to other anti-cancer agents.
Current challenges for cancer therapy with phytochemicals are, for example, the low solubility of compounds, cancer-specific delivery of compounds, and limited penetration of compounds into cancer. To overcome these impediments, the delivery of phytochemicals using nanoparticles has been investigated [38,337]. Moreover, the co-delivery of phytochemicals using nanoparticles displays synergistic effects on cancer cells (Section 3.3.3), demonstrating an opportunity to develop a new combination strategy for cancer therapy.
There is an unexpected modulation of miRNA levels (Section 2.1.6 and Section 2.4.4). Since there are over 2000 human miRNAs [338], it is feasible that a substantial number of miRNAs can be errantly modulated following cancer therapy, so alternative oncogenic factors can be activated, ultimately contributing to the acquired resistance to phytochemicals. Since drug resistance is another challenge in cancer therapy, comprehensive analyses of miRNA-mediated signaling pathways following the application of phytochemical compounds will enable the identification of the molecular targets to develop therapeutic strategies that prevent and overcome drug resistance. As mentioned in Section 4 and Section 5, the alteration of miRNA expression definitely regulates the sensitivity of cancer cells to phytochemicals and derivatives. From this perspective, a combination of miRNA modulators (mimics or inhibitors) and phytochemicals will also offer a new strategy for cancer therapy.
While miRNAs play critical roles in cancer, it has been realized that some miRNAs behave differently depending on the type of cancer. Examples of the dual role of miRNAs are discussed in this review (see Section 2.1.1 and Section 2.3.6). Additionally, as mentioned above, the treatment of cancer with therapeutic compounds can change miRNA levels in an unanticipated way. Therefore, it would be more promising if specific miRNAs are selectively targeted. Butylcycloheptyl prodiginine is a derivative of prodiginine, a natural compound from Gammaproteobacteria, and inhibits miR-21 levels by binding to miR-21 precursors and blocking Dicer-mediated maturation [339]. Additionally, we mentioned the ability of sophocarpine to target miR-21 precursors in Section 2.1.8. Unlike mature miRNAs, the sequence of miRNA precursors is typically not conserved among various miRNAs. Therefore, targeting miRNA precursors makes it possible to discriminate among miRNAs [340]. It suggests that phytochemicals (as well as natural compounds from other resources) can be developed as specific miRNA target agents to design a novel strategy for cancer therapy. Further investigations will be necessary to screen the effects of natural compounds on the Dicer-mediated maturation of other miRNAs. Advanced knowledge through further studies that uncover the role of phytochemicals and miRNAs will help to establish a clinically valuable therapeutic strategy against cancer.
Author Contributions
S.W.S., H.Y.L., and J.K.P. outlined the review; S.M., H.J.K., and S.Y.C. contributed to literature review; S.W.S. and H.Y.L. wrote a portion of manuscript; J.K.P. wrote and edited the manuscript. All authors have read and agreed to the published version of the manuscript.
Funding
This work was supported by a grant from the Basic Science Research Program through the National Research Foundation of Korea (NRF) funded by the Ministry of Education (2017R1D1A3B03035662) (J.K.P.) and (2019R1A6A1A11036849) (S.Y.C.); and the National Research Foundation of Korea (NRF) grant funded by the Korea government (MSIT) (2019R1A2C1089710) (J.K.P.) and (2019R1A5A2027588) (H.J.K.).
Conflicts of Interest
The authors declare no conflict of interest.
References
- Ferlay, J.; Colombet, M.; Soerjomataram, I.; Mathers, C.; Parkin, D.M.; Pineros, M.; Znaor, A.; Bray, F. Estimating the global cancer incidence and mortality in 2018: Globocan sources and methods. Int. J. Cancer 2019, 144, 1941–1953. [Google Scholar] [CrossRef] [Green Version]
- Saltalamacchia, G.; Frascaroli, M.; Bernardo, A.; Quaquarini, E. Renal and cardiovascular toxicities by new systemic treatments for prostate cancer. Cancers 2020, 12, 1750. [Google Scholar] [CrossRef] [PubMed]
- Nurgali, K.; Jagoe, R.T.; Abalo, R. Editorial: Adverse effects of cancer chemotherapy: Anything new to improve tolerance and reduce sequelae? Front. Pharmacol. 2018, 9, 245. [Google Scholar] [CrossRef] [PubMed]
- Seo, H.A.; Moeng, S.; Sim, S.; Kuh, H.J.; Choi, S.Y.; Park, J.K. Microrna-based combinatorial cancer therapy: Effects of micrornas on the efficacy of anti-cancer therapies. Cells 2019, 9, 29. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lichota, A.; Gwozdzinski, K. Anticancer activity of natural compounds from plant and marine environment. Int. J. Mol. Sci. 2018, 19, 3533. [Google Scholar] [CrossRef] [Green Version]
- Starczewska, E.; Beyaert, M.; Michaux, L.; Vekemans, M.C.; Saussoy, P.; Bol, V.; Arana Echarri, A.; Smal, C.; Van Den Neste, E.; Bontemps, F. Targeting DNA repair with aphidicolin sensitizes primary chronic lymphocytic leukemia cells to purine analogs. Oncotarget 2016, 7, 38367–38379. [Google Scholar] [CrossRef] [Green Version]
- Montane, X.; Kowalczyk, O.; Reig-Vano, B.; Bajek, A.; Roszkowski, K.; Tomczyk, R.; Pawliszak, W.; Giamberini, M.; Mocek-Plociniak, A.; Tylkowski, B. Current perspectives of the applications of polyphenols and flavonoids in cancer therapy. Molecules 2020, 25, 3342. [Google Scholar] [CrossRef]
- Mitra, T.; Bhattacharya, R. Phytochemicals modulate cancer aggressiveness: A review depicting the anticancer efficacy of dietary polyphenols and their combinations. J. Cell. Physiol. 2020, 235, 7696–7708. [Google Scholar] [CrossRef]
- Li, F.; Jiang, T.; Li, Q.; Ling, X. Camptothecin (cpt) and its derivatives are known to target topoisomerase i (top1) as their mechanism of action: Did we miss something in cpt analogue molecular targets for treating human disease such as cancer? Am. J. Cancer Res. 2017, 7, 2350–2394. [Google Scholar]
- Choudhari, A.S.; Mandave, P.C.; Deshpande, M.; Ranjekar, P.; Prakash, O. Phytochemicals in cancer treatment: From preclinical studies to clinical practice. Front. Pharmacol. 2019, 10, 1614. [Google Scholar] [CrossRef] [Green Version]
- Bergman, M.E.; Davis, B.; Phillips, M.A. Medically useful plant terpenoids: Biosynthesis, occurrence, and mechanism of action. Molecules 2019, 24, 3961. [Google Scholar] [CrossRef] [Green Version]
- Chen, F.; Zhu, H.H.; Zhou, L.F.; Wu, S.S.; Wang, J.; Chen, Z. Inhibition of c-flip expression by mir-512-3p contributes to taxol-induced apoptosis in hepatocellular carcinoma cells. Oncol. Rep. 2010, 23, 1457–1462. [Google Scholar] [CrossRef] [Green Version]
- Park, J.K.; Doseff, A.I.; Schmittgen, T.D. Micrornas targeting caspase-3 and -7 in panc-1 cells. Int. J. Mol. Sci. 2018, 19, 1206. [Google Scholar] [CrossRef] [Green Version]
- Moeng, S.; Son, S.W.; Lee, J.S.; Lee, H.Y.; Kim, T.H.; Choi, S.Y.; Kuh, H.J.; Park, J.K. Extracellular vesicles (evs) and pancreatic cancer: From the role of evs to the interference with ev-mediated reciprocal communication. Biomedicines 2020, 8, 267. [Google Scholar] [CrossRef]
- Sun, M.; Zhang, Q.; Yang, X.; Qian, S.Y.; Guo, B. Vitamin d enhances the efficacy of irinotecan through mir-627-mediated inhibition of intratumoral drug metabolism. Mol. Cancer Ther. 2016, 15, 2086–2095. [Google Scholar] [CrossRef] [Green Version]
- Wu, Y.; Jin, X.; Liao, W.; Hu, L.; Dawuda, M.M.; Zhao, X.; Tang, Z.; Gong, T.; Yu, J. 5-aminolevulinic acid (ala) alleviated salinity stress in cucumber seedlings by enhancing chlorophyll synthesis pathway. Front. Plant Sci. 2018, 9, 635. [Google Scholar] [CrossRef] [Green Version]
- Feng, M.; Luo, X.; Gu, C.; Li, Y.; Zhu, X.; Fei, J. Systematic analysis of berberine-induced signaling pathway between mirna clusters and mrnas and identification of mir-99a approximately 125b cluster function by seed-targeting inhibitors in multiple myeloma cells. RNA Biol. 2015, 12, 82–91. [Google Scholar] [CrossRef] [Green Version]
- Chai, F.N.; Ma, W.Y.; Zhang, J.; Xu, H.S.; Li, Y.F.; Zhou, Q.D.; Li, X.G.; Ye, X.L. Coptisine from rhizoma coptidis exerts an anti-cancer effect on hepatocellular carcinoma by up-regulating mir-122. Biomed. Pharmacother. 2018, 103, 1002–1011. [Google Scholar] [CrossRef]
- Jiang, J.; Hu, C. Evodiamine: A novel anti-cancer alkaloid from evodia rutaecarpa. Molecules 2009, 14, 1852–1859. [Google Scholar] [CrossRef]
- Hargraves, K.G.; He, L.; Firestone, G.L. Phytochemical regulation of the tumor suppressive microrna, mir-34a, by p53-dependent and independent responses in human breast cancer cells. Mol. Carcinog. 2016, 55, 486–498. [Google Scholar] [CrossRef] [Green Version]
- Fu, S.; Zhao, N.; Jing, G.; Yang, X.; Liu, J.; Zhen, D.; Tang, X. Matrine induces papillary thyroid cancer cell apoptosis in vitro and suppresses tumor growth in vivo by downregulating mir-182-5p. Biomed. Pharmacother. 2020, 128, 110327. [Google Scholar] [CrossRef]
- Liu, Z.; Zhang, S.; Wang, T.; Shao, H.; Gao, J.; Wang, Y.; Ge, Y. Neferine inhibits mda-mb-231cells growth and metastasis by regulating mir-374a/fgfr-2. Chem. Biol. Interact 2019, 309, 108716. [Google Scholar] [CrossRef]
- Mao, X.; Hu, Z.; Wang, Q.; Zhang, N.; Zhou, S.; Peng, Y.; Zheng, J. Nitidine chloride is a mechanism-based inactivator of cyp2d6. Drug Metab. Dispos. 2018, 46, 1137–1145. [Google Scholar] [CrossRef]
- Karki, K.; Hedrick, E.; Kasiappan, R.; Jin, U.H.; Safe, S. Piperlongumine induces reactive oxygen species (ros)-dependent downregulation of specificity protein transcription factors. Cancer Prev. Res. (Phila) 2017, 10, 467–477. [Google Scholar] [CrossRef] [Green Version]
- Dong, X.Z.; Song, Y.; Lu, Y.P.; Hu, Y.; Liu, P.; Zhang, L. Sanguinarine inhibits the proliferation of bgc-823 gastric cancer cells via regulating mir-96-5p/mir-29c-3p and the mapk/jnk signaling pathway. J. Nat. Med. 2019, 73, 777–788. [Google Scholar] [CrossRef]
- Xu, F.; Li, Q.; Wang, Z.; Cao, X. Sinomenine inhibits proliferation, migration, invasion and promotes apoptosis of prostate cancer cells by regulation of mir-23a. Biomed. Pharmacother. 2019, 112, 108592. [Google Scholar] [CrossRef]
- Liu, W.; Zhang, B.; Chen, G.; Wu, W.; Zhou, L.; Shi, Y.; Zeng, Q.; Li, Y.; Sun, Y.; Deng, X.; et al. Targeting mir-21 with sophocarpine inhibits tumor progression and reverses epithelial-mesenchymal transition in head and neck cancer. Mol. Ther. 2017, 25, 2129–2139. [Google Scholar] [CrossRef] [Green Version]
- Sun, L.; Jin, X.; Xie, L.; Xu, G.; Cui, Y.; Chen, Z. Swainsonine represses glioma cell proliferation, migration and invasion by reduction of mir-92a expression. BMC Cancer 2019, 19, 247. [Google Scholar] [CrossRef] [Green Version]
- Lv, Q.; Xia, Q.; Li, J.; Wang, Z. Allicin suppresses growth and metastasis of gastric carcinoma: The key role of microrna-383-5p-mediated inhibition of erbb4 signaling. Biosci. Biotechnol. Biochem. 2020, 84, 1997–2004. [Google Scholar] [CrossRef]
- Martin, S.L.; Kala, R.; Tollefsbol, T.O. Mechanisms for the inhibition of colon cancer cells by sulforaphane through epigenetic modulation of microrna-21 and human telomerase reverse transcriptase (htert) down-regulation. Curr. Cancer Drug Targets 2018, 18, 97–106. [Google Scholar] [CrossRef]
- Xu, H.; Li, Y.; Han, B.; Li, Z.; Wang, B.; Jiang, P.; Zhang, J.; Ma, W.; Zhou, D.; Li, X.; et al. Anti-breast-cancer activity exerted by beta-sitosterol-d-glucoside from sweet potato via upregulation of microrna-10a and via the pi3k-akt signaling pathway. J. Agric. Food Chem. 2018, 66, 9704–9718. [Google Scholar] [CrossRef]
- Sok, S.P.; Arshad, N.M.; Azmi, M.N.; Awang, K.; Ozpolat, B.; Hasima Nagoor, N. The apoptotic effect of 1’s-1’-acetoxychavicol acetate (aca) enhanced by inhibition of non-canonical autophagy in human non-small cell lung cancer cells. PLoS ONE 2017, 12, e0171329. [Google Scholar] [CrossRef] [Green Version]
- Gao, A.M.; Zhang, X.Y.; Ke, Z.P. Apigenin sensitizes bel-7402/adm cells to doxorubicin through inhibiting mir-101/nrf2 pathway. Oncotarget 2017, 8, 82085–82091. [Google Scholar] [CrossRef] [Green Version]
- Li, W.; Hao, J.; Zhang, L.; Cheng, Z.; Deng, X.; Shu, G. Astragalin reduces hexokinase 2 through increasing mir-125b to inhibit the proliferation of hepatocellular carcinoma cells in vitro and in vivo. J. Agric. Food Chem. 2017, 65, 5961–5972. [Google Scholar] [CrossRef]
- Bie, B.; Sun, J.; Li, J.; Guo, Y.; Jiang, W.; Huang, C.; Yang, J.; Li, Z. Baicalein, a natural anti-cancer compound, alters microrna expression profiles in bel-7402 human hepatocellular carcinoma cells. Cell. Physiol. Biochem. 2017, 41, 1519–1531. [Google Scholar] [CrossRef]
- Tao, Y.; Zhan, S.; Wang, Y.; Zhou, G.; Liang, H.; Chen, X.; Shen, H. Baicalin, the major component of traditional chinese medicine scutellaria baicalensis induces colon cancer cell apoptosis through inhibition of oncomirnas. Sci. Rep. 2018, 8, 14477. [Google Scholar] [CrossRef] [Green Version]
- Tao, L.Y.; Li, J.Y.; Zhang, J.Y. Brazilein, a compound isolated from caesalpinia sappan linn., induced growth inhibition in breast cancer cells via involvement of gsk-3beta/beta-catenin/cyclin d1 pathway. Chem. Biol. Interact. 2013, 206, 1–5. [Google Scholar] [CrossRef]
- Javan, N.; Khadem Ansari, M.H.; Dadashpour, M.; Khojastehfard, M.; Bastami, M.; Rahmati-Yamchi, M.; Zarghami, N. Synergistic antiproliferative effects of co-nanoencapsulated curcumin and chrysin on mda-mb-231 breast cancer cells through upregulating mir-132 and mir-502c. Nutr. Cancer 2019, 71, 1201–1213. [Google Scholar] [CrossRef]
- Zhou, L.; Lu, Y.; Liu, J.S.; Long, S.Z.; Liu, H.L.; Zhang, J.; Zhang, T. The role of mir-21/reck in the inhibition of osteosarcoma by curcumin. Mol. Cell. Probes 2020, 51, 101534. [Google Scholar] [CrossRef]
- Huang, C.C.; Hung, C.H.; Hung, T.W.; Lin, Y.C.; Wang, C.J.; Kao, S.H. Dietary delphinidin inhibits human colorectal cancer metastasis associating with upregulation of mir-204-3p and suppression of the integrin/fak axis. Sci. Rep. 2019, 9, 18954. [Google Scholar] [CrossRef] [Green Version]
- Lin, C.H.; Wang, H.H.; Chen, T.H.; Chiang, M.C.; Hung, P.H.; Chen, Y.J. Involvement of microrna-296 in the inhibitory effect of epigallocatechin gallate against the migratory properties of anoikis-resistant nasopharyngeal carcinoma cells. Cancers 2020, 12, 973. [Google Scholar] [CrossRef] [Green Version]
- Wu, Y.; Zhang, X.; Li, Z.; Yan, H.; Qin, J.; Li, T. Formononetin inhibits human bladder cancer cell proliferation and invasiveness via regulation of mir-21 and pten. Food Funct. 2017, 8, 1061–1066. [Google Scholar] [CrossRef]
- Zou, Y.; Li, R.; Kuang, D.; Zuo, M.; Li, W.; Tong, W.; Jiang, L.; Zhou, M.; Chen, Y.; Gong, W.; et al. Galangin inhibits cholangiocarcinoma cell growth and metastasis through downregulation of microrna-21 expression. Biomed. Res. Int. 2020, 2020, 5846938. [Google Scholar] [CrossRef]
- Gao, G.; Bian, Y.; Qian, H.; Yang, M.; Hu, J.; Li, L.; Yu, L.; Liu, B.; Qian, X. Gambogic acid regulates the migration and invasion of colorectal cancer via microrna-21-mediated activation of phosphatase and tensin homolog. Exp. Ther. Med. 2018, 16, 1758–1765. [Google Scholar] [CrossRef] [Green Version]
- Hsieh, P.L.; Liao, Y.W.; Hsieh, C.W.; Chen, P.N.; Yu, C.C. Soy isoflavone genistein impedes cancer stemness and mesenchymal transition in head and neck cancer through activating mir-34a/rtcb axis. Nutrients 2020, 12, 1924. [Google Scholar] [CrossRef]
- Lin, J.; Wu, Y.; Yang, D.; Zhao, Y. Induction of apoptosis and antitumor effects of a small molecule inhibitor of bcl-2 and bcl-xl, gossypol acetate, in multiple myeloma in vitro and in vivo. Oncol. Rep. 2013, 30, 731–738. [Google Scholar] [CrossRef] [Green Version]
- Chou, L.F.; Chen, C.Y.; Yang, W.H.; Chen, C.C.; Chang, J.L.; Leu, Y.L.; Liou, M.J.; Wang, T.H. Suppression of hepatocellular carcinoma progression through foxm1 and emt inhibition via hydroxygenkwanin-induced mir-320a expression. Biomolecules 2019, 10, 20. [Google Scholar] [CrossRef] [Green Version]
- Yang, J.; Zou, Y.; Jiang, D. Honokiol suppresses proliferation and induces apoptosis via regulation of the mir21/pten/pi3k/akt signaling pathway in human osteosarcoma cells. Int. J. Mol. Med. 2018, 41, 1845–1854. [Google Scholar]
- Fang, L.; Xu, W.; Kong, D. Icariin inhibits cell proliferation, migration and invasion by down-regulation of microrna-625-3p in thyroid cancer cells. Biomed. Pharmacother. 2019, 109, 2456–2463. [Google Scholar] [CrossRef]
- Wang, T.T.; Chen, Z.Z.; Xie, P.; Zhang, W.J.; Du, M.Y.; Liu, Y.T.; Zhu, H.Y.; Guo, Y.S. Isoliquiritigenin suppresses the proliferation and induced apoptosis via mir-32/lats2/wnt in nasopharyngeal carcinoma. Eur. J. Pharmacol. 2019, 856, 172352. [Google Scholar] [CrossRef]
- Zeng, X.; Xu, Z.; Gu, J.; Huang, H.; Gao, G.; Zhang, X.; Li, J.; Jin, H.; Jiang, G.; Sun, H.; et al. Induction of mir-137 by isorhapontigenin (iso) directly targets sp1 protein translation and mediates its anticancer activity both in vitro and in vivo. Mol. Cancer Ther. 2016, 15, 512–522. [Google Scholar] [CrossRef] [Green Version]
- Han, X.; Liu, C.F.; Gao, N.; Zhao, J.; Xu, J. Kaempferol suppresses proliferation but increases apoptosis and autophagy by up-regulating microrna-340 in human lung cancer cells. Biomed. Pharmacother. 2018, 108, 809–816. [Google Scholar] [CrossRef]
- Huang, W.C.; Su, H.H.; Fang, L.W.; Wu, S.J.; Liou, C.J. Licochalcone a inhibits cellular motility by suppressing e-cadherin and mapk signaling in breast cancer. Cells 2019, 8, 218. [Google Scholar] [CrossRef] [Green Version]
- Jiang, Z.Q.; Li, M.H.; Qin, Y.M.; Jiang, H.Y.; Zhang, X.; Wu, M.H. Luteolin inhibits tumorigenesis and induces apoptosis of non-small cell lung cancer cells via regulation of microrna-34a-5p. Int. J. Mol. Sci. 2018, 19, 447. [Google Scholar] [CrossRef] [Green Version]
- Niu, Y.; Zhang, J.; Tong, Y.; Li, J.; Liu, B. Physcion 8-o-beta-glucopyranoside induced ferroptosis via regulating mir-103a-3p/gls2 axis in gastric cancer. Life Sci. 2019, 237, 116893. [Google Scholar] [CrossRef]
- Mao, J.T.; Xue, B.; Smoake, J.; Lu, Q.Y.; Park, H.; Henning, S.M.; Burns, W.; Bernabei, A.; Elashoff, D.; Serio, K.J.; et al. Microrna-19a/b mediates grape seed procyanidin extract-induced anti-neoplastic effects against lung cancer. J. Nutr. Biochem. 2016, 34, 118–125. [Google Scholar] [CrossRef]
- Zhou, Y.; Xue, R.; Wang, J.; Ren, H. Puerarin inhibits hepatocellular carcinoma invasion and metastasis through mir-21-mediated pten/akt signaling to suppress the epithelial-mesenchymal transition. Braz. J. Med. Biol. Res. 2020, 53, e8882. [Google Scholar] [CrossRef] [Green Version]
- Nwaeburu, C.C.; Bauer, N.; Zhao, Z.; Abukiwan, A.; Gladkich, J.; Benner, A.; Herr, I. Up-regulation of microrna let-7c by quercetin inhibits pancreatic cancer progression by activation of numbl. Oncotarget 2016, 7, 58367–58380. [Google Scholar] [CrossRef] [Green Version]
- Wu, F.; Cui, L. Resveratrol suppresses melanoma by inhibiting nf-kappab/mir-221 and inducing tfg expression. Arch Dermatol. Res. 2017, 309, 823–831. [Google Scholar] [CrossRef]
- Liu, J.; Qu, C.B.; Xue, Y.X.; Li, Z.; Wang, P.; Liu, Y.H. Mir-143 enhances the antitumor activity of shikonin by targeting bag3 expression in human glioblastoma stem cells. Biochem. Biophys. Res. Commun. 2015, 468, 105–112. [Google Scholar] [CrossRef]
- Zadeh, M.M.; Motamed, N.; Ranji, N.; Majidi, M.; Falahi, F. Silibinin-induced apoptosis and downregulation of microrna-21 and microrna-155 in mcf-7 human breast cancer cells. J. Breast Cancer 2016, 19, 45–52. [Google Scholar] [CrossRef] [Green Version]
- Federico, A.; Dallio, M.; Di Fabio, G.; Zarrelli, A.; Zappavigna, S.; Stiuso, P.; Tuccillo, C.; Caraglia, M.; Loguercio, C. Silybin-phosphatidylcholine complex protects human gastric and liver cells from oxidative stress. In Vivo 2015, 29, 569–575. [Google Scholar]
- Zarrelli, A.; Romanucci, V.; Tuccillo, C.; Federico, A.; Loguercio, C.; Gravante, R.; Di Fabio, G. New silibinin glyco-conjugates: Synthesis and evaluation of antioxidant properties. Bioorg. Med. Chem. Lett. 2014, 24, 5147–5149. [Google Scholar] [CrossRef]
- Singh, T.; Prasad, R.; Katiyar, S.K. Therapeutic intervention of silymarin on the migration of non-small cell lung cancer cells is associated with the axis of multiple molecular targets including class 1 hdacs, zeb1 expression, and restoration of mir-203 and e-cadherin expression. Am. J. Cancer Res. 2016, 6, 1287–1301. [Google Scholar]
- Lu, M.; Liu, B.; Xiong, H.; Wu, F.; Hu, C.; Liu, P. Trans-3,5,4 -trimethoxystilbene reduced gefitinib resistance in nsclcs via suppressing mapk/akt/bcl-2 pathway by upregulation of mir-345 and mir-498. J. Cell. Mol. Med. 2019, 23, 2431–2441. [Google Scholar] [CrossRef] [Green Version]
- Zhang, Y.; Zhang, C.; Min, D. Ailanthone up-regulates mir-449a to restrain acute myeloid leukemia cells growth, migration and invasion. Exp. Mol. Pathol. 2019, 108, 114–120. [Google Scholar] [CrossRef]
- Yang, P.Y.; Hsieh, P.L.; Wang, T.H.; Yu, C.C.; Lu, M.Y.; Liao, Y.W.; Lee, T.H.; Peng, C.Y. Andrographolide impedes cancer stemness and enhances radio-sensitivity in oral carcinomas via mir-218 activation. Oncotarget 2017, 8, 4196–4207. [Google Scholar] [CrossRef] [Green Version]
- Konstat-Korzenny, E.; Ascencio-Aragon, J.A.; Niezen-Lugo, S.; Vazquez-Lopez, R. Artemisinin and its synthetic derivatives as a possible therapy for cancer. Med. Sci. 2018, 6, 19. [Google Scholar] [CrossRef] [Green Version]
- Xu, F.; Cui, W.Q.; Wei, Y.; Cui, J.; Qiu, J.; Hu, L.L.; Gong, W.Y.; Dong, J.C.; Liu, B.J. Astragaloside iv inhibits lung cancer progression and metastasis by modulating macrophage polarization through ampk signaling. J. Exp. Clin. Cancer Res. 2018, 37, 207. [Google Scholar] [CrossRef]
- Xiao, Z.; Ching Chow, S.; Han Li, C.; Chun Tang, S.; Tsui, S.K.; Lin, Z.; Chen, Y. Role of microrna-95 in the anticancer activity of brucein d in hepatocellular carcinoma. Eur. J. Pharmacol. 2014, 728, 141–150. [Google Scholar] [CrossRef]
- Alharris, E.; Singh, N.P.; Nagarkatti, P.S.; Nagarkatti, M. Role of mirna in the regulation of cannabidiol-mediated apoptosis in neuroblastoma cells. Oncotarget 2019, 10, 45–59. [Google Scholar] [CrossRef] [Green Version]
- Ni, H.; Han, Y.; Jin, X. Celastrol inhibits colon cancer cell proliferation by downregulating mir-21 and pi3k/akt/gsk-3beta pathway. Int. J. Clin. Exp. Pathol. 2019, 12, 808–816. [Google Scholar]
- Sikander, M.; Hafeez, B.B.; Malik, S.; Alsayari, A.; Halaweish, F.T.; Yallapu, M.M.; Chauhan, S.C.; Jaggi, M. Cucurbitacin d exhibits potent anti-cancer activity in cervical cancer. Sci. Rep. 2016, 6, 36594. [Google Scholar] [CrossRef]
- Zeng, C.; Fan, D.; Xu, Y.; Li, X.; Yuan, J.; Yang, Q.; Zhou, X.; Lu, J.; Zhang, C.; Han, J.; et al. Curcumol enhances the sensitivity of doxorubicin in triple-negative breast cancer via regulating the mir-181b-2-3p-abcc3 axis. Biochem. Pharmacol. 2020, 174, 113795. [Google Scholar] [CrossRef]
- Wang, P.; Du, X.; Xiong, M.; Cui, J.; Yang, Q.; Wang, W.; Chen, Y.; Zhang, T. Ginsenoside rd attenuates breast cancer metastasis implicating derepressing microrna-18a-regulated smad2 expression. Sci. Rep. 2016, 6, 33709. [Google Scholar] [CrossRef] [Green Version]
- Gao, Q.; Zheng, J. Ginsenoside rh2 inhibits prostate cancer cell growth through suppression of microrna-4295 that activates cdkn1a. Cell Prolif. 2018, 51, e12438. [Google Scholar] [CrossRef] [Green Version]
- Li, D.; Chen, L.; Zhao, W.; Hao, J.; An, R. Microrna-let-7f-1 is induced by lycopene and inhibits cell proliferation and triggers apoptosis in prostate cancer. Mol. Med. Rep. 2016, 13, 2708–2714. [Google Scholar] [CrossRef] [Green Version]
- Hernandes, C.; Miguita, L.; de Sales, R.O.; Silva, E.P.; Mendonca, P.O.R.; Lorencini da Silva, B.; Klingbeil, M.F.G.; Mathor, M.B.; Rangel, E.B.; Marti, L.C.; et al. Anticancer activities of the quinone-methide triterpenes maytenin and 22-beta-hydroxymaytenin obtained from cultivated maytenus ilicifolia roots associated with down-regulation of mirna-27a and mir-20a/mir-17-5p. Molecules 2020, 25, 760. [Google Scholar] [CrossRef] [Green Version]
- Zhao, X.; Liu, M.; Li, D. Oleanolic acid suppresses the proliferation of lung carcinoma cells by mir-122/cyclin g1/mef2d axis. Mol. Cell. Biochem. 2015, 400, 1–7. [Google Scholar] [CrossRef]
- Weng, H.; Huang, H.; Dong, B.; Zhao, P.; Zhou, H.; Qu, L. Inhibition of mir-17 and mir-20a by oridonin triggers apoptosis and reverses chemoresistance by derepressing bim-s. Cancer Res. 2014, 74, 4409–4419. [Google Scholar] [CrossRef] [Green Version]
- Cheng, S.; Zhang, Z.; Hu, C.; Xing, N.; Xia, Y.; Pang, B. Pristimerin suppressed breast cancer progression via mir-542-5p/dub3 axis. Onco Targets Ther. 2020, 13, 6651–6660. [Google Scholar] [CrossRef]
- Wang, G.; Huang, Y.X.; Zhang, R.; Hou, L.D.; Liu, H.; Chen, X.Y.; Zhu, J.S.; Zhang, J. Toosendanin suppresses oncogenic phenotypes of human gastric carcinoma sgc7901 cells partly via mir200amediated downregulation of beta-catenin pathway. Int. J. Oncol. 2017, 51, 1563–1573. [Google Scholar] [CrossRef] [Green Version]
- Li, S.G.; Shi, Q.W.; Yuan, L.Y.; Qin, L.P.; Wang, Y.; Miao, Y.Q.; Chen, Z.; Ling, C.Q.; Qin, W.X. C-myc-dependent repression of two oncogenic mirna clusters contributes to triptolide-induced cell death in hepatocellular carcinoma cells. J. Exp. Clin. Cancer Res. 2018, 37, 51. [Google Scholar] [CrossRef]
- Shi, H.; Bi, H.; Sun, X.; Dong, H.; Jiang, Y.; Mu, H.; Liu, G.; Kong, W.; Gao, R.; Su, J. Antitumor effects of tubeimoside-1 in nci-h1299 cells are mediated by microrna-126-5p-induced inactivation of vegf-a/vegfr-2/erk signaling pathway. Mol. Med. Rep. 2018, 17, 4327–4336. [Google Scholar] [CrossRef]
- Xiang, F.; Fan, Y.; Ni, Z.; Liu, Q.; Zhu, Z.; Chen, Z.; Hao, W.; Yue, H.; Wu, R.; Kang, X. Ursolic acid reverses the chemoresistance of breast cancer cells to paclitaxel by targeting mirna-149-5p/myd88. Front. Oncol. 2019, 9, 501. [Google Scholar] [CrossRef] [Green Version]
- Xu, Q.; Li, M.; Yang, M.; Yang, J.; Xie, J.; Lu, X.; Wang, F.; Chen, W. Alpha-pinene regulates mir-221 and induces g2/m phase cell cycle arrest in human hepatocellular carcinoma cells. Biosci. Rep. 2018, 38, BSR20180980. [Google Scholar] [CrossRef] [Green Version]
- Long, X.; Shi, Y.; Ye, P.; Guo, J.; Zhou, Q.; Tang, Y. Microrna-99a suppresses breast cancer progression by targeting fgfr3. Front. Oncol. 2019, 9, 1473. [Google Scholar] [CrossRef] [Green Version]
- Murray, M.Y.; Rushworth, S.A.; Zaitseva, L.; Bowles, K.M.; Macewan, D.J. Attenuation of dexamethasone-induced cell death in multiple myeloma is mediated by mir-125b expression. Cell Cycle 2013, 12, 2144–2153. [Google Scholar] [CrossRef]
- Yue, X.; Zhang, C.; Zhao, Y.; Liu, J.; Lin, A.W.; Tan, V.M.; Drake, J.M.; Liu, L.; Boateng, M.N.; Li, J.; et al. Gain-of-function mutant p53 activates small gtpase rac1 through sumoylation to promote tumor progression. Genes Dev. 2017, 31, 1641–1654. [Google Scholar] [CrossRef] [Green Version]
- Liu, H.; Huang, C.; Wu, L.; Wen, B. Effect of evodiamine and berberine on mir-429 as an oncogene in human colorectal cancer. Onco Targets Ther. 2016, 9, 4121–4127. [Google Scholar]
- Liu, N.; Li, P.; Zang, S.; Liu, Q.; Ma, D.; Sun, X.; Ji, C. Novel agent nitidine chloride induces erythroid differentiation and apoptosis in cml cells through c-myc-mirnas axis. PLoS ONE 2015, 10, e0116880. [Google Scholar] [CrossRef]
- Pan, J.; Shen, J.; Si, W.; Du, C.; Chen, D.; Xu, L.; Yao, M.; Fu, P.; Fan, W. Resveratrol promotes mica/b expression and natural killer cell lysis of breast cancer cells by suppressing c-myc/mir-17 pathway. Oncotarget 2017, 8, 65743–65758. [Google Scholar] [CrossRef] [Green Version]
- Lan, F.; Pan, Q.; Yu, H.; Yue, X. Sulforaphane enhances temozolomide-induced apoptosis because of down-regulation of mir-21 via wnt/beta-catenin signaling in glioblastoma. J. Neurochem. 2015, 134, 811–818. [Google Scholar] [CrossRef]
- Lewinska, A.; Adamczyk-Grochala, J.; Deregowska, A.; Wnuk, M. Sulforaphane-induced cell cycle arrest and senescence are accompanied by DNA hypomethylation and changes in microrna profile in breast cancer cells. Theranostics 2017, 7, 3461–3477. [Google Scholar] [CrossRef]
- Zan, L.; Chen, Q.; Zhang, L.; Li, X. Epigallocatechin gallate (egcg) suppresses growth and tumorigenicity in breast cancer cells by downregulation of mir-25. Bioengineered 2019, 10, 374–382. [Google Scholar] [CrossRef] [Green Version]
- Liu, Z.M.; Yang, X.L.; Jiang, F.; Pan, Y.C.; Zhang, L. Matrine involves in the progression of gastric cancer through inhibiting mir-93-5p and upregulating the expression of target gene ahnak. J. Cell Biochem. 2020, 121, 2467–2477. [Google Scholar] [CrossRef]
- Coker-Gurkan, A.; Bulut, D.; Genc, R.; Arisan, E.D.; Obakan-Yerlikaya, P.; Palavan-Unsal, N. Curcumin prevented human autocrine growth hormone (gh) signaling mediated nf-kappab activation and mir-183-96-182 cluster stimulated epithelial mesenchymal transition in t47d breast cancer cells. Mol. Biol. Rep. 2019, 46, 355–369. [Google Scholar] [CrossRef]
- Zhou, W.; Wang, S.; Ying, Y.; Zhou, R.; Mao, P. Mir-196b/mir-1290 participate in the antitumor effect of resveratrol via regulation of igfbp3 expression in acute lymphoblastic leukemia. Oncol. Rep. 2017, 37, 1075–1083. [Google Scholar] [CrossRef] [Green Version]
- Jia, Y.; Chen, L.; Guo, S.; Li, Y. Baicalin induced colon cancer cells apoptosis through mir-217/dkk1-mediated inhibition of wnt signaling pathway. Mol. Biol. Rep. 2019, 46, 1693–1700. [Google Scholar] [CrossRef]
- Allegri, L.; Rosignolo, F.; Mio, C.; Filetti, S.; Baldan, F.; Damante, G. Effects of nutraceuticals on anaplastic thyroid cancer cells. J. Cancer Res. Clin. Oncol. 2018, 144, 285–294. [Google Scholar] [CrossRef]
- Ma, J.; Cheng, L.; Liu, H.; Zhang, J.; Shi, Y.; Zeng, F.; Miele, L.; Sarkar, F.H.; Xia, J.; Wang, Z. Genistein down-regulates mir-223 expression in pancreatic cancer cells. Curr. Drug Targets 2013, 14, 1150–1156. [Google Scholar] [CrossRef]
- Xiang, S.; Chen, H.; Luo, X.; An, B.; Wu, W.; Cao, S.; Ruan, S.; Wang, Z.; Weng, L.; Zhu, H.; et al. Isoliquiritigenin suppresses human melanoma growth by targeting mir-301b/lrig1 signaling. J. Exp. Clin. Cancer Res. 2018, 37, 184. [Google Scholar] [CrossRef]
- Moeng, S.; Son, S.W.; Seo, H.A.; Lee, J.S.; Kim, C.K.; Kuh, H.J.; Park, J.K. Luteolin-regulated microrna-301-3p targets caspase-8 and modulates trail sensitivity in panc-1 cells. Anticancer Res. 2020, 40, 723–731. [Google Scholar] [CrossRef]
- Song, L.; Liu, D.; Zhao, Y.; He, J.; Kang, H.; Dai, Z.; Wang, X.; Zhang, S.; Zan, Y. Sinomenine inhibits breast cancer cell invasion and migration by suppressing nf-kappab activation mediated by il-4/mir-324-5p/cuedc2 axis. Biochem. Biophys. Res. Commun. 2015, 464, 705–710. [Google Scholar] [CrossRef]
- Peng, F.; Tang, H.; Liu, P.; Shen, J.; Guan, X.; Xie, X.; Gao, J.; Xiong, L.; Jia, L.; Chen, J.; et al. Isoliquiritigenin modulates mir-374a/pten/akt axis to suppress breast cancer tumorigenesis and metastasis. Sci. Rep. 2017, 7, 9022. [Google Scholar] [CrossRef] [Green Version]
- Wang, K.; Tan, S.L.; Lu, Q.; Xu, R.; Cao, J.; Wu, S.Q.; Wang, Y.H.; Zhao, X.K.; Zhong, Z.H. Curcumin suppresses microrna-7641-mediated regulation of p16 expression in bladder cancer. Am. J. Chin. Med. 2018, 46, 1357–1368. [Google Scholar] [CrossRef]
- Jung, Y.Y.; Um, J.Y.; Narula, A.S.; Namjoshi, O.A.; Blough, B.E.; Kumar, A.P.; Ahn, K.S. Identification of matrine as a novel regulator of the cxcr4 signaling axis in tumor cells. Int. J. Mol. Sci. 2020, 21, 4731. [Google Scholar] [CrossRef]
- Shen, E.; Wang, X.; Liu, X.; Lv, M.; Zhang, L.; Zhu, G.; Sun, Z. Microrna-93-5p promotes epithelial-mesenchymal transition in gastric cancer by repressing tumor suppressor ahnak expression. Cancer Cell Int. 2020, 20, 76. [Google Scholar] [CrossRef]
- Sobhani, N.; Ianza, A.; D’Angelo, A.; Roviello, G.; Giudici, F.; Bortul, M.; Zanconati, F.; Bottin, C.; Generali, D. Current status of fibroblast growth factor receptor-targeted therapies in breast cancer. Cells 2018, 7, 76. [Google Scholar] [CrossRef] [Green Version]
- Sun, S.; Jiang, Y.; Zhang, G.; Song, H.; Zhang, X.; Zhang, Y.; Liang, X.; Sun, Q.; Pang, D. Increased expression of fibroblastic growth factor receptor 2 is correlated with poor prognosis in patients with breast cancer. J. Surg. Oncol. 2012, 105, 773–779. [Google Scholar] [CrossRef]
- Deng, G.; Zeng, S.; Ma, J.; Zhang, Y.; Qu, Y.; Han, Y.; Yin, L.; Cai, C.; Guo, C.; Shen, H. The anti-tumor activities of neferine on cell invasion and oxaliplatin sensitivity regulated by emt via snail signaling in hepatocellular carcinoma. Sci. Rep. 2017, 7, 41616. [Google Scholar] [CrossRef] [Green Version]
- Nazim, U.M.; Yin, H.; Park, S.Y. Neferine treatment enhances the trailinduced apoptosis of human prostate cancer cells via autophagic flux and the jnk pathway. Int. J. Oncol. 2020, 56, 1152–1161. [Google Scholar]
- Turczyk, L.; Kitowska, K.; Mieszkowska, M.; Mieczkowski, K.; Czaplinska, D.; Piasecka, D.; Kordek, R.; Skladanowski, A.C.; Potemski, P.; Romanska, H.M.; et al. Fgfr2-driven signaling counteracts tamoxifen effect on eralpha-positive breast cancer cells. Neoplasia 2017, 19, 791–804. [Google Scholar] [CrossRef]
- Nakamura, S.; Yokota, D.; Tan, L.; Nagata, Y.; Takemura, T.; Hirano, I.; Shigeno, K.; Shibata, K.; Fujisawa, S.; Ohnishi, K. Down-regulation of thanatos-associated protein 11 by bcr-abl promotes cml cell proliferation through c-myc expression. Int. J. Cancer 2012, 130, 1046–1059. [Google Scholar] [CrossRef] [Green Version]
- Sodaro, G.; Cesaro, E.; Montano, G.; Blasio, G.; Fiorentino, F.; Romano, S.; Jacquel, A.; Aurberger, P.; Costanzo, P. Role of znf224 in c-myc repression and imatinib responsiveness in chronic myeloid leukemia. Oncotarget 2018, 9, 3417–3431. [Google Scholar] [CrossRef]
- Grandjenette, C.; Schnekenburger, M.; Karius, T.; Ghelfi, J.; Gaigneaux, A.; Henry, E.; Dicato, M.; Diederich, M. 5-aza-2’-deoxycytidine-mediated c-myc down-regulation triggers telomere-dependent senescence by regulating human telomerase reverse transcriptase in chronic myeloid leukemia. Neoplasia 2014, 16, 511–528. [Google Scholar] [CrossRef] [Green Version]
- Li, Y.; Choi, P.S.; Casey, S.C.; Dill, D.L.; Felsher, D.W. Myc through mir-17-92 suppresses specific target genes to maintain survival, autonomous proliferation, and a neoplastic state. Cancer Cell 2014, 26, 262–272. [Google Scholar] [CrossRef] [Green Version]
- Niu, H.; Wang, K.; Zhang, A.; Yang, S.; Song, Z.; Wang, W.; Qian, C.; Li, X.; Zhu, Y.; Wang, Y. Mir-92a is a critical regulator of the apoptosis pathway in glioblastoma with inverse expression of bcl2l11. Oncol. Rep. 2012, 28, 1771–1777. [Google Scholar] [CrossRef] [Green Version]
- Song, H.; Zhang, Y.; Liu, N.; Zhao, S.; Kong, Y.; Yuan, L. Mir-92a-3p exerts various effects in glioma and glioma stem-like cells specifically targeting cdh1/beta-catenin and notch-1/akt signaling pathways. Int. J. Mol. Sci. 2016, 17, 1799. [Google Scholar] [CrossRef]
- Liu, P.J.; Ye, Y.X.; Wang, Y.X.; Du, J.X.; Pan, Y.H.; Fang, X.B. Mirna-92a promotes cell proliferation and invasion through binding to klf4 in glioma. Eur. Rev. Med. Pharmacol. Sci. 2019, 23, 6612–6620. [Google Scholar]
- Rawat, L.; Hegde, H.; Hoti, S.L.; Nayak, V. Piperlongumine induces ros mediated cell death and synergizes paclitaxel in human intestinal cancer cells. Biomed. Pharmacother. 2020, 128, 110243. [Google Scholar] [CrossRef]
- Zhang, H.; Zhang, J.; Venkat, P.S.; Gu, C.; Meng, Y. Sanguinarine exhibits potent efficacy against cervical cancer cells through inhibiting the stat3 pathway in vitro and in vivo. Cancer Manag. Res. 2019, 11, 7557–7566. [Google Scholar] [CrossRef] [Green Version]
- Zhu, M.; Gong, Z.; Wu, Q.; Shi, X.; Su, Q.; Zhang, Y. Sanguinarine suppresses migration and metastasis in colorectal carcinoma associated with the inversion of emt through the wnt/beta-catenin signaling. Clin. Transl. Med. 2020, 10, 1–12. [Google Scholar] [CrossRef]
- Su, Q.; Fan, M.; Wang, J.; Ullah, A.; Ghauri, M.A.; Dai, B.; Zhan, Y.; Zhang, D.; Zhang, Y. Sanguinarine inhibits epithelial-mesenchymal transition via targeting hif-1alpha/tgf-beta feed-forward loop in hepatocellular carcinoma. Cell Death Dis. 2019, 10, 939. [Google Scholar] [CrossRef]
- He, X.; Zou, K. Mirna-96-5p contributed to the proliferation of gastric cancer cells by targeting foxo3. J. Biochem. 2020, 167, 101–108. [Google Scholar] [CrossRef]
- Gao, Y.; Qi, W.; Sun, L.; Lv, J.; Qiu, W.; Liu, S. Foxo3 inhibits human gastric adenocarcinoma (ags) cell growth by promoting autophagy in an acidic microenvironment. Cell Physiol. Biochem. 2018, 49, 335–348. [Google Scholar] [CrossRef]
- Wang, L.; Yu, T.; Li, W.; Li, M.; Zuo, Q.; Zou, Q.; Xiao, B. The mir-29c-kiaa1199 axis regulates gastric cancer migration by binding with wbp11 and ptp4a3. Oncogene 2019, 38, 3134–3150. [Google Scholar] [CrossRef]
- Liu, X.; Cai, J.; Sun, Y.; Gong, R.; Sun, D.; Zhong, X.; Jiang, S.; He, X.; Bao, E.; Yang, L.; et al. Microrna-29a inhibits cell migration and invasion via targeting roundabout homolog 1 in gastric cancer cells. Mol. Med. Rep. 2015, 12, 3944–3950. [Google Scholar] [CrossRef] [Green Version]
- Gao, L.; Cheng, D.; Yang, J.; Wu, R.; Li, W.; Kong, A.N. Sulforaphane epigenetically demethylates the cpg sites of the mir-9-3 promoter and reactivates mir-9-3 expression in human lung cancer a549 cells. J. Nutr. Biochem. 2018, 56, 109–115. [Google Scholar] [CrossRef]
- Tang, J.; Wang, Z.; Chen, L.; Huang, G.; Hu, X. Gossypol acetate induced apoptosis of pituitary tumor cells by targeting the bcl-2 via the upregulated microrna mir-15a. Int. J. Clin. Exp. Med. 2015, 8, 9079–9085. [Google Scholar]
- Zhang, B.; Wang, X.; Deng, J.; Zheng, H.; Liu, W.; Chen, S.; Tian, J.; Wang, F. P53-dependent upregulation of mir-16-2 by sanguinarine induces cell cycle arrest and apoptosis in hepatocellular carcinoma. Cancer Lett. 2019, 459, 50–58. [Google Scholar] [CrossRef]
- Hu, Z.; Fan, H.; Lv, G.; Zhou, Q.; Yang, B.; Zheng, J.; Cao, W. 5-aminolevulinic acid-mediated sonodynamic therapy induces anti-tumor effects in malignant melanoma via p53-mir-34a-sirt1 axis. J. Dermatol. Sci. 2015, 79, 155–162. [Google Scholar] [CrossRef]
- Gallardo, M.; Kemmerling, U.; Aguayo, F.; Bleak, T.C.; Munoz, J.P.; Calaf, G.M. Curcumin rescues breast cells from epithelialmesenchymal transition and invasion induced by antimir34a. Int. J. Oncol. 2020, 56, 480–493. [Google Scholar]
- Otsuka, K.; Yamamoto, Y.; Ochiya, T. Regulatory role of resveratrol, a microrna-controlling compound, in hnrnpa1 expression, which is associated with poor prognosis in breast cancer. Oncotarget 2018, 9, 24718–24730. [Google Scholar] [CrossRef] [Green Version]
- Zhang, W.; Jiang, H.; Chen, Y.; Ren, F. Resveratrol chemosensitizes adriamycin-resistant breast cancer cells by modulating mir-122-5p. J. Cell Biochem. 2019, 120, 16283–16292. [Google Scholar] [CrossRef]
- Zhang, D.; Han, Y.; Xu, L. Upregulation of mir-124 by physcion 8-o-beta-glucopyranoside inhibits proliferation and invasion of malignant melanoma cells via repressing rlip76. Biomed. Pharmacother. 2016, 84, 166–176. [Google Scholar] [CrossRef]
- Beccafico, S.; Morozzi, G.; Marchetti, M.C.; Riccardi, C.; Sidoni, A.; Donato, R.; Sorci, G. Artesunate induces ros- and p38 mapk-mediated apoptosis and counteracts tumor growth in vivo in embryonal rhabdomyosarcoma cells. Carcinogenesis 2015, 36, 1071–1083. [Google Scholar] [CrossRef] [Green Version]
- Mou, Z.; Wang, Y.; Li, Y. Brazilein induces apoptosis and g1/g0 phase cell cycle arrest by up-regulation of mir-133a in human vestibular schwannoma cells. Exp. Mol. Pathol. 2019, 107, 95–101. [Google Scholar] [CrossRef]
- Ye, Q.; Su, L.; Chen, D.; Zheng, W.; Liu, Y. Astragaloside iv induced mir-134 expression reduces emt and increases chemotherapeutic sensitivity by suppressing creb1 signaling in colorectal cancer cell line sw-480. Cell Physiol. Biochem. 2017, 43, 1617–1626. [Google Scholar] [CrossRef]
- Guo, X.; Huang, H.; Jin, H.; Xu, J.; Risal, S.; Li, J.; Li, X.; Yan, H.; Zeng, X.; Xue, L.; et al. Iso, via upregulating mir-137 transcription, inhibits gsk3beta-hsp70-mmp-2 axis, resulting in attenuating urothelial cancer invasion. Mol. Ther. Nucleic. Acids 2018, 12, 337–349. [Google Scholar] [CrossRef] [Green Version]
- Xiao, X.; Zhang, Y.; Pan, W.; Chen, F. Mir-139-mediated notch1 regulation is crucial for the inhibition of osteosarcoma progression caused by resveratrol. Life Sci. 2020, 242, 117215. [Google Scholar] [CrossRef]
- Guo, Q.; Dong, B.; Nan, F.; Guan, D.; Zhang, Y. 5-aminolevulinic acid photodynamic therapy in human cervical cancer via the activation of microrna-143 and suppression of the bcl-2/bax signaling pathway. Mol. Med. Rep. 2016, 14, 544–550. [Google Scholar] [CrossRef]
- Chen, G.; Ma, Y.; Jiang, Z.; Feng, Y.; Han, Y.; Tang, Y.; Zhang, J.; Ni, H.; Li, X.; Li, N. Lico a causes er stress and apoptosis via up-regulating mir-144-3p in human lung cancer cell line h292. Front. Pharmacol. 2018, 9, 837. [Google Scholar] [CrossRef]
- Xu, Z.; Zeng, X.; Xu, J.; Xu, D.; Li, J.; Jin, H.; Jiang, G.; Han, X.; Huang, C. Isorhapontigenin suppresses growth of patient-derived glioblastoma spheres through regulating mir-145/sox2/cyclin d1 axis. Neuro. Oncol. 2016, 18, 830–839. [Google Scholar] [CrossRef] [Green Version]
- Gao, W.; Ge, S.; Sun, J. Ailanthone exerts anticancer effect by up-regulating mir-148a expression in mda-mb-231 breast cancer cells and inhibiting proliferation, migration and invasion. Biomed. Pharmacother. 2019, 109, 1062–1069. [Google Scholar] [CrossRef]
- Chen, Q.; Luo, J.; Wu, C.; Lu, H.; Cai, S.; Bao, C.; Liu, D.; Kong, J. The mirna-149-5p/myd88 axis is responsible for ursolic acid-mediated attenuation of the stemness and chemoresistance of non-small cell lung cancer cells. Environ. Toxicol. 2020, 35, 561–569. [Google Scholar] [CrossRef]
- Hang, S.; Wang, X.; Li, H. Triptolide inhibits viability and migration while promotes apoptosis in nephroblastoma cells by regulation of mir-193b-3p. Exp. Mol. Pathol. 2019, 108, 80–88. [Google Scholar] [CrossRef]
- Nwaeburu, C.C.; Abukiwan, A.; Zhao, Z.; Herr, I. Quercetin-induced mir-200b-3p regulates the mode of self-renewing divisions in pancreatic cancer. Mol. Cancer 2017, 16, 23. [Google Scholar] [CrossRef] [Green Version]
- Karimi Dermani, F.; Saidijam, M.; Amini, R.; Mahdavinezhad, A.; Heydari, K.; Najafi, R. Resveratrol inhibits proliferation, invasion, and epithelial-mesenchymal transition by increasing mir-200c expression in hct-116 colorectal cancer cells. J. Cell. Biochem 2017, 118, 1547–1555. [Google Scholar] [CrossRef]
- Yao, Y.; Rao, C.; Zheng, G.; Wang, S. Luteolin suppresses colorectal cancer cell metastasis via regulation of the mir384/pleiotrophin axis. Oncol. Rep. 2019, 42, 131–141. [Google Scholar]
- Jiang, P.; Xu, C.; Chen, L.; Chen, A.; Wu, X.; Zhou, M.; Haq, I.U.; Mariyam, Z.; Feng, Q. Egcg inhibits csc-like properties through targeting mir-485/cd44 axis in a549-cisplatin resistant cells. Mol. Carcinog. 2018, 57, 1835–1844. [Google Scholar] [CrossRef]
- Gao, A.M.; Zhang, X.Y.; Hu, J.N.; Ke, Z.P. Apigenin sensitizes hepatocellular carcinoma cells to doxorubic through regulating mir-520b/atg7 axis. Chem. Biol. Interact. 2018, 280, 45–50. [Google Scholar] [CrossRef]
- Vislovukh, A.; Kratassiouk, G.; Porto, E.; Gralievska, N.; Beldiman, C.; Pinna, G.; El’skaya, A.; Harel-Bellan, A.; Negrutskii, B.; Groisman, I. Proto-oncogenic isoform a2 of eukaryotic translation elongation factor eef1 is a target of mir-663 and mir-744. Br. J. Cancer 2013, 108, 2304–2311. [Google Scholar] [CrossRef] [Green Version]
- Yang, P.W.; Lu, Z.Y.; Pan, Q.; Chen, T.T.; Feng, X.J.; Wang, S.M.; Pan, Y.C.; Zhu, M.H.; Zhang, S.H. Microrna-6809-5p mediates luteolin-induced anticancer effects against hepatoma by targeting flotillin 1. Phytomedicine 2019, 57, 18–29. [Google Scholar] [CrossRef]
- Wang, H.; Tan, Z.; Hu, H.; Liu, H.; Wu, T.; Zheng, C.; Wang, X.; Luo, Z.; Wang, J.; Liu, S.; et al. Microrna-21 promotes breast cancer proliferation and metastasis by targeting lztfl1. BMC Cancer 2019, 19, 738. [Google Scholar] [CrossRef] [Green Version]
- Nguyen, D.D.; Chang, S. Development of novel therapeutic agents by inhibition of oncogenic micrornas. Int. J. Mol. Sci. 2017, 19, 65. [Google Scholar] [CrossRef] [Green Version]
- Wang, C.; Li, Q.; He, Y. Microrna215p promotes epithelial to mesenchymal transition by targeting srybox 17 in endometrial cancer. Oncol. Rep. 2020, 43, 1897–1905. [Google Scholar]
- Han, M.; Liu, M.; Wang, Y.; Chen, X.; Xu, J.; Sun, Y.; Zhao, L.; Qu, H.; Fan, Y.; Wu, C. Antagonism of mir-21 reverses epithelial-mesenchymal transition and cancer stem cell phenotype through akt/erk1/2 inactivation by targeting pten. PLoS ONE 2012, 7, e39520. [Google Scholar] [CrossRef] [Green Version]
- Liu, N.; Guo, X.H.; Liu, J.P.; Cong, Y.S. Role of telomerase in the tumour microenvironment. Clin. Exp. Pharmacol. Physiol. 2020, 47, 357–364. [Google Scholar] [CrossRef] [Green Version]
- Abbas, A.; Hall, J.A.; Patterson, W.L., III; Ho, E.; Hsu, A.; Al-Mulla, F.; Georgel, P.T. Sulforaphane modulates telomerase activity via epigenetic regulation in prostate cancer cell lines. Biochem. Cell Biol. 2016, 94, 71–81. [Google Scholar] [CrossRef]
- Yang, Y.; Yang, J.J.; Tao, H.; Jin, W.S. Microrna-21 controls htert via pten in human colorectal cancer cell proliferation. J. Physiol. Biochem. 2015, 71, 59–68. [Google Scholar] [CrossRef]
- Li, Y.; Cheng, H.S.; Chng, W.J.; Tergaonkar, V. Activation of mutant tert promoter by ras-erk signaling is a key step in malignant progression of braf-mutant human melanomas. Proc. Natl. Acad. Sci. USA 2016, 113, 14402–14407. [Google Scholar] [CrossRef] [Green Version]
- Hannafon, B.N.; Cai, A.; Calloway, C.L.; Xu, Y.F.; Zhang, R.; Fung, K.M.; Ding, W.Q. Mir-23b and mir-27b are oncogenic micrornas in breast cancer: Evidence from a crispr/cas9 deletion study. BMC Cancer 2019, 19, 642. [Google Scholar] [CrossRef]
- Ho, J.Y.; Hsu, R.J.; Liu, J.M.; Chen, S.C.; Liao, G.S.; Gao, H.W.; Yu, C.P. Microrna-382-5p aggravates breast cancer progression by regulating the rerg/ras/erk signaling axis. Oncotarget 2017, 8, 22443–22459. [Google Scholar] [CrossRef] [Green Version]
- Clark, J.C.; Thomas, D.M.; Choong, P.F.; Dass, C.R. Reck—A newly discovered inhibitor of metastasis with prognostic significance in multiple forms of cancer. Cancer Metastasis Rev. 2007, 26, 675–683. [Google Scholar] [CrossRef]
- Zhang, J.; Kong, X.; Li, J.; Luo, Q.; Li, X.; Shen, L.; Chen, L.; Fang, L. Mir-96 promotes tumor proliferation and invasion by targeting reck in breast cancer. Oncol. Rep. 2014, 31, 1357–1363. [Google Scholar] [CrossRef] [Green Version]
- Kessenbrock, K.; Plaks, V.; Werb, Z. Matrix metalloproteinases: Regulators of the tumor microenvironment. Cell 2010, 141, 52–67. [Google Scholar] [CrossRef] [Green Version]
- Liao, H.; Wang, Z.; Deng, Z.; Ren, H.; Li, X. Curcumin inhibits lung cancer invasion and metastasis by attenuating glut1/mt1-mmp/mmp2 pathway. Int. J. Clin. Exp. Med. 2015, 8, 8948–8957. [Google Scholar]
- Mbese, Z.; Khwaza, V.; Aderibigbe, B.A. Curcumin and its derivatives as potential therapeutic agents in prostate, colon and breast cancers. Molecules 2019, 24, 4386. [Google Scholar] [CrossRef] [Green Version]
- Zhang, W.; Qian, P.; Zhang, X.; Zhang, M.; Wang, H.; Wu, M.; Kong, X.; Tan, S.; Ding, K.; Perry, J.K.; et al. Autocrine/paracrine human growth hormone-stimulated microrna 96-182-183 cluster promotes epithelial-mesenchymal transition and invasion in breast cancer. J. Biol. Chem. 2015, 290, 13812–13829. [Google Scholar] [CrossRef] [Green Version]
- Mukhina, S.; Mertani, H.C.; Guo, K.; Lee, K.O.; Gluckman, P.D.; Lobie, P.E. Phenotypic conversion of human mammary carcinoma cells by autocrine human growth hormone. Proc. Natl. Acad. Sci. USA 2004, 101, 15166–15171. [Google Scholar] [CrossRef] [Green Version]
- Zhu, T.; Starling-Emerald, B.; Zhang, X.; Lee, K.O.; Gluckman, P.D.; Mertani, H.C.; Lobie, P.E. Oncogenic transformation of human mammary epithelial cells by autocrine human growth hormone. Cancer Res. 2005, 65, 317–324. [Google Scholar]
- Li, P.; Sheng, C.; Huang, L.; Zhang, H.; Huang, L.; Cheng, Z.; Zhu, Q. Mir-183/-96/-182 cluster is up-regulated in most breast cancers and increases cell proliferation and migration. Breast Cancer Res. 2014, 16, 473. [Google Scholar] [CrossRef] [Green Version]
- Kim, M.; Katayose, Y.; Rojanala, L.; Shah, S.; Sgagias, M.; Jang, L.; Jung, Y.J.; Lee, S.H.; Hwang, S.G.; Cowan, K.H. Induction of apoptosis in p16ink4a mutant cell lines by adenovirus-mediated overexpression of p16ink4a protein. Cell Death Differ. 2000, 7, 706–711. [Google Scholar] [CrossRef] [Green Version]
- Silva, G.; Aboussekhra, A. P16(ink4a) inhibits the pro-metastatic potentials of osteosarcoma cells through targeting the erk pathway and tgf-beta1. Mol. Carcinog. 2016, 55, 525–536. [Google Scholar] [CrossRef]
- Wang, Z.; Li, T.E.; Chen, M.; Pan, J.J.; Shen, K.W. Mir-106b-5p contributes to the lung metastasis of breast cancer via targeting cnn1 and regulating rho/rock1 pathway. Aging (Albany NY) 2020, 12, 1867–1887. [Google Scholar] [CrossRef]
- Chen, H.; Pan, H.; Qian, Y.; Zhou, W.; Liu, X. Mir-25-3p promotes the proliferation of triple negative breast cancer by targeting btg2. Mol. Cancer 2018, 17, 4. [Google Scholar] [CrossRef]
- Xie, C.M.; Tan, M.; Lin, X.T.; Wu, D.; Jiang, Y.; Tan, Y.; Li, H.; Ma, Y.; Xiong, X.; Sun, Y. The fbxw7-shoc2-raptor axis controls the cross-talks between the ras-erk and mtorc1 signaling pathways. Cell Rep. 2019, 26, 3037–3050.e4. [Google Scholar] [CrossRef] [Green Version]
- Sun, D.; Wang, C.; Long, S.; Ma, Y.; Guo, Y.; Huang, Z.; Chen, X.; Zhang, C.; Chen, J.; Zhang, J. C/ebp-beta-activated microrna-223 promotes tumour growth through targeting rasa1 in human colorectal cancer. Br. J. Cancer 2015, 112, 1491–1500. [Google Scholar] [CrossRef] [Green Version]
- Harmon, A.W.; Patel, Y.M.; Harp, J.B. Genistein inhibits ccaat/enhancer-binding protein beta (c/ebpbeta) activity and 3t3-l1 adipogenesis by increasing c/ebp homologous protein expression. Biochem. J. 2002, 367, 203–208. [Google Scholar] [CrossRef]
- Li, Y.; Zhou, H.C.; Zhang, Y.; Huang, H. Microrna-625-3p inhibits gastric cancer metastasis through modulating ezh2. Eur. Rev. Med. Pharmacol. Sci. 2020, 24, 1177–1185. [Google Scholar]
- Huang, L.; Jin, K.; Lan, H. Luteolin inhibits cell cycle progression and induces apoptosis of breast cancer cells through downregulation of human telomerase reverse transcriptase. Oncol. Lett. 2019, 17, 3842–3850. [Google Scholar] [CrossRef] [Green Version]
- Wang, H.; Luo, Y.; Qiao, T.; Wu, Z.; Huang, Z. Luteolin sensitizes the antitumor effect of cisplatin in drug-resistant ovarian cancer via induction of apoptosis and inhibition of cell migration and invasion. J. Ovarian Res. 2018, 11, 93. [Google Scholar] [CrossRef]
- Tu, S.H.; Ho, C.T.; Liu, M.F.; Huang, C.S.; Chang, H.W.; Chang, C.H.; Wu, C.H.; Ho, Y.S. Luteolin sensitises drug-resistant human breast cancer cells to tamoxifen via the inhibition of cyclin e2 expression. Food Chem. 2013, 141, 1553–1561. [Google Scholar] [CrossRef]
- Stockwell, B.R.; Friedmann Angeli, J.P.; Bayir, H.; Bush, A.I.; Conrad, M.; Dixon, S.J.; Fulda, S.; Gascon, S.; Hatzios, S.K.; Kagan, V.E.; et al. Ferroptosis: A regulated cell death nexus linking metabolism, redox biology, and disease. Cell 2017, 171, 273–285. [Google Scholar] [CrossRef] [Green Version]
- Gao, M.; Monian, P.; Quadri, N.; Ramasamy, R.; Jiang, X. Glutaminolysis and transferrin regulate ferroptosis. Mol. Cell 2015, 59, 298–308. [Google Scholar] [CrossRef] [Green Version]
- Hou, C.; Chen, Y.; Huang, X.; Huang, Q.; Li, M.; Tan, X. Mir-19 targets pten and mediates high mobility group protein b1(hmgb1)-induced proliferation and migration of human airway smooth muscle cells. PLoS ONE 2019, 14, e0219081. [Google Scholar] [CrossRef] [Green Version]
- Liu, D.T.; Yao, H.R.; Li, Y.Y.; Song, Y.Y.; Su, M.Y. Microrna-19b promotes the migration and invasion of ovarian cancer cells by inhibiting the pten/akt signaling pathway. Oncol. Lett. 2018, 16, 559–565. [Google Scholar] [CrossRef]
- Zhang, P.; Li, H.; Wu, M.L.; Chen, X.Y.; Kong, Q.Y.; Wang, X.W.; Sun, Y.; Wen, S.; Liu, J. C-myc downregulation: A critical molecular event in resveratrol-induced cell cycle arrest and apoptosis of human medulloblastoma cells. J. Neurooncol. 2006, 80, 123–131. [Google Scholar] [CrossRef]
- Li, W.; Li, C.; Ma, L.; Jin, F. Resveratrol inhibits viability and induces apoptosis in the smallcell lung cancer h446 cell line via the pi3k/akt/cmyc pathway. Oncol. Rep. 2020, 44, 1821–1830. [Google Scholar]
- Bhardwaj, A.; Sethi, G.; Vadhan-Raj, S.; Bueso-Ramos, C.; Takada, Y.; Gaur, U.; Nair, A.S.; Shishodia, S.; Aggarwal, B.B. Resveratrol inhibits proliferation, induces apoptosis, and overcomes chemoresistance through down-regulation of stat3 and nuclear factor-kappab-regulated antiapoptotic and cell survival gene products in human multiple myeloma cells. Blood 2007, 109, 2293–2302. [Google Scholar] [CrossRef] [Green Version]
- Ko, J.H.; Sethi, G.; Um, J.Y.; Shanmugam, M.K.; Arfuso, F.; Kumar, A.P.; Bishayee, A.; Ahn, K.S. The role of resveratrol in cancer therapy. Int. J. Mol. Sci. 2017, 18, 2589. [Google Scholar] [CrossRef] [Green Version]
- Dutton-Regester, K.; Aoude, L.G.; Nancarrow, D.J.; Stark, M.S.; O’Connor, L.; Lanagan, C.; Pupo, G.M.; Tembe, V.; Carter, C.D.; O’Rourke, M.; et al. Identification of tfg (trk-fused gene) as a putative metastatic melanoma tumor suppressor gene. Genes Chromosomes Cancer 2012, 51, 452–461. [Google Scholar] [CrossRef]
- Visone, R.; Russo, L.; Pallante, P.; De Martino, I.; Ferraro, A.; Leone, V.; Borbone, E.; Petrocca, F.; Alder, H.; Croce, C.M.; et al. Micrornas (mir)-221 and mir-222, both overexpressed in human thyroid papillary carcinomas, regulate p27kip1 protein levels and cell cycle. Endocr. Relat. Cancer 2007, 14, 791–798. [Google Scholar] [CrossRef]
- Zheng, G.; Li, N.; Jia, X.; Peng, C.; Luo, L.; Deng, Y.; Yin, J.; Song, Y.; Liu, H.; Lu, M.; et al. Mycn-mediated mir-21 overexpression enhances chemo-resistance via targeting cadm1 in tongue cancer. J. Mol. Med. 2016, 94, 1129–1141. [Google Scholar] [CrossRef]
- Zhang, G.; Zhong, L.; Luo, H.; Wang, S. Microrna-155-3p promotes breast cancer progression through down-regulating cadm1. Onco Targets Ther. 2019, 12, 7993–8002. [Google Scholar] [CrossRef] [Green Version]
- Khalighfard, S.; Alizadeh, A.M.; Irani, S.; Omranipour, R. Plasma mir-21, mir-155, mir-10b, and let-7a as the potential biomarkers for the monitoring of breast cancer patients. Sci. Rep. 2018, 8, 17981. [Google Scholar] [CrossRef]
- Natarajan, G.; Ramalingam, S.; Ramachandran, I.; May, R.; Queimado, L.; Houchen, C.W.; Anant, S. Cugbp2 downregulation by prostaglandin e2 protects colon cancer cells from radiation-induced mitotic catastrophe. Am. J. Physiol. Gastrointest. Liver Physiol. 2008, 294, G1235–G1244. [Google Scholar] [CrossRef] [Green Version]
- Yeung, Y.T.; Fan, S.; Lu, B.; Yin, S.; Yang, S.; Nie, W.; Wang, M.; Zhou, L.; Li, T.; Li, X.; et al. Celf2 suppresses non-small cell lung carcinoma growth by inhibiting the prex2-pten interaction. Carcinogenesis 2020, 41, 377–389. [Google Scholar] [CrossRef]
- Subramaniam, D.; Ramalingam, S.; Linehan, D.C.; Dieckgraefe, B.K.; Postier, R.G.; Houchen, C.W.; Jensen, R.A.; Anant, S. Rna binding protein cugbp2/celf2 mediates curcumin-induced mitotic catastrophe of pancreatic cancer cells. PLoS ONE 2011, 6, e16958. [Google Scholar] [CrossRef] [Green Version]
- Lai, Z.Q.; Ip, S.P.; Liao, H.J.; Lu, Z.; Xie, J.H.; Su, Z.R.; Chen, Y.L.; Xian, Y.F.; Leung, P.S.; Lin, Z.X. Brucein d, a naturally occurring tetracyclic triterpene quassinoid, induces apoptosis in pancreatic cancer through ros-associated pi3k/akt signaling pathway. Front. Pharmacol. 2017, 8, 936. [Google Scholar] [CrossRef] [Green Version]
- Xu, L.N.; Zhao, N.; Chen, J.Y.; Ye, P.P.; Nan, X.W.; Zhou, H.H.; Jiang, Q.W.; Yang, Y.; Huang, J.R.; Yuan, M.L.; et al. Celastrol inhibits the growth of ovarian cancer cells in vitro and in vivo. Front. Oncol. 2019, 9, 2. [Google Scholar] [CrossRef]
- Lee, H.W.; Jang, K.S.; Choi, H.J.; Jo, A.; Cheong, J.H.; Chun, K.H. Celastrol inhibits gastric cancer growth by induction of apoptosis and autophagy. BMB Rep. 2014, 47, 697–702. [Google Scholar] [CrossRef] [Green Version]
- Tang, Y.C.; Zhang, Y.; Zhou, J.; Zhi, Q.; Wu, M.Y.; Gong, F.R.; Shen, M.; Liu, L.; Tao, M.; Shen, B.; et al. Ginsenoside rg3 targets cancer stem cells and tumor angiogenesis to inhibit colorectal cancer progression in vivo. Int. J. Oncol. 2018, 52, 127–138. [Google Scholar] [CrossRef]
- Li, T.; Sun, W.; Dong, X.; Yu, W.; Cai, J.; Yuan, Q.; Shan, L.; Efferth, T. Total ginsenosides of chinese ginseng induces cell cycle arrest and apoptosis in colorectal carcinoma ht-29 cells. Oncol. Lett. 2018, 16, 4640–4648. [Google Scholar] [CrossRef] [Green Version]
- Chung, K.S.; Cho, S.H.; Shin, J.S.; Kim, D.H.; Choi, J.H.; Choi, S.Y.; Rhee, Y.K.; Hong, H.D.; Lee, K.T. Ginsenoside rh2 induces cell cycle arrest and differentiation in human leukemia cells by upregulating tgf-beta expression. Carcinogenesis 2013, 34, 331–340. [Google Scholar] [CrossRef] [Green Version]
- Singha, P.K.; Pandeswara, S.; Geng, H.; Lan, R.; Venkatachalam, M.A.; Dobi, A.; Srivastava, S.; Saikumar, P. Increased smad3 and reduced smad2 levels mediate the functional switch of tgf-beta from growth suppressor to growth and metastasis promoter through tmepai/pmepa1 in triple negative breast cancer. Genes Cancer 2019, 10, 134–149. [Google Scholar] [CrossRef] [Green Version]
- Petersen, M.; Pardali, E.; van der Horst, G.; Cheung, H.; van den Hoogen, C.; van der Pluijm, G.; Ten Dijke, P. Smad2 and smad3 have opposing roles in breast cancer bone metastasis by differentially affecting tumor angiogenesis. Oncogene 2010, 29, 1351–1361. [Google Scholar] [CrossRef] [Green Version]
- Lu, S.; Zhou, C.; Zou, B.; Zhang, H.; Feng, M. Mir-4295 facilitates cell proliferation and metastasis in head and neck squamous cell carcinoma by targeting nptx1. Genes Immun. 2020, 21, 4–12. [Google Scholar] [CrossRef]
- Yan, R.; Li, K.; Yuan, D.W.; Wang, H.N.; Zhang, Y.; Dang, C.X.; Zhu, K. Downregulation of microrna-4295 enhances cisplatin-induced gastric cancer cell apoptosis through the egfr/pi3k/akt signaling pathway by targeting lrig1. Int. J. Oncol. 2018, 53, 2566–2578. [Google Scholar] [CrossRef] [Green Version]
- Zeng, G.; Xun, W.; Wei, K.; Yang, Y.; Shen, H. Microrna-27a-3p regulates epithelial to mesenchymal transition via targeting yap1 in oral squamous cell carcinoma cells. Oncol. Rep. 2016, 36, 1475–1482. [Google Scholar] [CrossRef] [Green Version]
- Kannan, M.; Jayamohan, S.; Moorthy, R.K.; Chabattula, S.C.; Ganeshan, M.; Arockiam, A.J.V. Aeg-1/mir-221 axis cooperatively regulates the progression of hepatocellular carcinoma by targeting pten/pi3k/akt signaling pathway. Int. J. Mol. Sci. 2019, 20, 5526. [Google Scholar] [CrossRef] [Green Version]
- Park, J.K.; Kogure, T.; Nuovo, G.J.; Jiang, J.; He, L.; Kim, J.H.; Phelps, M.A.; Papenfuss, T.L.; Croce, C.M.; Patel, T.; et al. Mir-221 silencing blocks hepatocellular carcinoma and promotes survival. Cancer Res. 2011, 71, 7608–7616. [Google Scholar] [CrossRef] [Green Version]
- Suehiro, S.; Ohnishi, T.; Yamashita, D.; Kohno, S.; Inoue, A.; Nishikawa, M.; Ohue, S.; Tanaka, J.; Kunieda, T. Enhancement of antitumor activity by using 5-ala-mediated sonodynamic therapy to induce apoptosis in malignant gliomas: Significance of high-intensity focused ultrasound on 5-ala-sdt in a mouse glioma model. J. Neurosurg. 2018, 129, 1416–1428. [Google Scholar] [CrossRef] [Green Version]
- Austin, E.; Koo, E.; Jagdeo, J. Thermal photodynamic therapy increases apoptosis and reactive oxygen species generation in cutaneous and mucosal squamous cell carcinoma cells. Sci. Rep. 2018, 8, 12599. [Google Scholar] [CrossRef]
- Tsai, W.C.; Hsu, P.W.; Lai, T.C.; Chau, G.Y.; Lin, C.W.; Chen, C.M.; Lin, C.D.; Liao, Y.L.; Wang, J.L.; Chau, Y.P.; et al. Microrna-122, a tumor suppressor microrna that regulates intrahepatic metastasis of hepatocellular carcinoma. Hepatology 2009, 49, 1571–1582. [Google Scholar] [CrossRef]
- Xu, Q.; Zhang, M.; Tu, J.; Pang, L.; Cai, W.; Liu, X. Microrna-122 affects cell aggressiveness and apoptosis by targeting pkm2 in human hepatocellular carcinoma. Oncol. Rep. 2015, 34, 2054–2064. [Google Scholar] [CrossRef] [Green Version]
- Slabakova, E.; Culig, Z.; Remsik, J.; Soucek, K. Alternative mechanisms of mir-34a regulation in cancer. Cell Death Dis. 2017, 8, e3100. [Google Scholar] [CrossRef]
- Katz, E.; Nisani, S.; Chamovitz, D.A. Indole-3-carbinol: A plant hormone combatting cancer. F1000Research 2018, 7, 689. [Google Scholar] [CrossRef] [Green Version]
- Liu, Y.; Chen, X.; Cheng, R.; Yang, F.; Yu, M.; Wang, C.; Cui, S.; Hong, Y.; Liang, H.; Liu, M.; et al. The jun/mir-22/hur regulatory axis contributes to tumourigenesis in colorectal cancer. Mol. Cancer 2018, 17, 11. [Google Scholar] [CrossRef]
- Yamakuchi, M.; Yagi, S.; Ito, T.; Lowenstein, C.J. Microrna-22 regulates hypoxia signaling in colon cancer cells. PLoS ONE 2011, 6, e20291. [Google Scholar] [CrossRef] [Green Version]
- Wang, J.; Li, Y.; Ding, M.; Zhang, H.; Xu, X.; Tang, J. Molecular mechanisms and clinical applications of mir-22 in regulating malignant progression in human cancer (review). Int. J. Oncol. 2017, 50, 345–355. [Google Scholar] [CrossRef]
- Cimmino, A.; Calin, G.A.; Fabbri, M.; Iorio, M.V.; Ferracin, M.; Shimizu, M.; Wojcik, S.E.; Aqeilan, R.I.; Zupo, S.; Dono, M.; et al. Mir-15 and mir-16 induce apoptosis by targeting bcl2. Proc. Natl. Acad. Sci. USA 2005, 102, 13944–13949. [Google Scholar] [CrossRef] [Green Version]
- Shi, L.; Jackstadt, R.; Siemens, H.; Li, H.; Kirchner, T.; Hermeking, H. P53-induced mir-15a/16-1 and ap4 form a double-negative feedback loop to regulate epithelial-mesenchymal transition and metastasis in colorectal cancer. Cancer Res. 2014, 74, 532–542. [Google Scholar] [CrossRef] [Green Version]
- Cheng, B.; Ding, F.; Huang, C.Y.; Xiao, H.; Fei, F.Y.; Li, J. Role of mir-16-5p in the proliferation and metastasis of hepatocellular carcinoma. Eur. Rev. Med. Pharmacol. Sci. 2019, 23, 137–145. [Google Scholar]
- Li, W.; Ge, Z.; Liu, C.; Liu, Z.; Bjorkholm, M.; Jia, J.; Xu, D. Cip2a is overexpressed in gastric cancer and its depletion leads to impaired clonogenicity, senescence, or differentiation of tumor cells. Clin. Cancer Res. 2008, 14, 3722–3728. [Google Scholar] [CrossRef] [Green Version]
- Li, X.; Yuan, J.; Cao, Q.; Xie, A.; Chen, J. Microrna3835p inhibits the proliferation and promotes the apoptosis of gastric cancer cells by targeting cancerous inhibitor of pp2a. Int. J. Mol. Med. 2020, 46, 397–405. [Google Scholar] [CrossRef]
- Zhang, S.; Wang, K. Mir-383 down-regulates the oncogene cip2a to influence glioma proliferation and invasion. Onco Targets Ther. 2020, 13, 4063–4074. [Google Scholar] [CrossRef]
- Naumann, P.; Liermann, J.; Fortunato, F.; Schmid, T.E.; Weber, K.J.; Debus, J.; Combs, S.E. Sulforaphane enhances irradiation effects in terms of perturbed cell cycle progression and increased DNA damage in pancreatic cancer cells. PLoS ONE 2017, 12, e0180940. [Google Scholar] [CrossRef] [Green Version]
- Xiong, K.; Shao, L.H.; Zhang, H.Q.; Jin, L.; Wei, W.; Dong, Z.; Zhu, Y.Q.; Wu, N.; Jin, S.Z.; Xue, L.X. Microrna-9 functions as a tumor suppressor and enhances radio-sensitivity in radio-resistant a549 cells by targeting neuropilin 1. Oncol. Lett. 2018, 15, 2863–2870. [Google Scholar] [CrossRef] [Green Version]
- Ke, K.; Lou, T. Microrna-10a suppresses breast cancer progression via pi3k/akt/mtor pathway. Oncol. Lett. 2017, 14, 5994–6000. [Google Scholar] [CrossRef] [Green Version]
- Ryoo, I.G.; Kim, G.; Choi, B.H.; Lee, S.H.; Kwak, M.K. Involvement of nrf2 signaling in doxorubicin resistance of cancer stem cell-enriched colonospheres. Biomol. Ther. (Seoul) 2016, 24, 482–488. [Google Scholar] [CrossRef] [Green Version]
- DeWaal, D.; Nogueira, V.; Terry, A.R.; Patra, K.C.; Jeon, S.M.; Guzman, G.; Au, J.; Long, C.P.; Antoniewicz, M.R.; Hay, N. Hexokinase-2 depletion inhibits glycolysis and induces oxidative phosphorylation in hepatocellular carcinoma and sensitizes to metformin. Nat. Commun. 2018, 9, 446. [Google Scholar] [CrossRef]
- Sun, Y.; Chen, C.; Zhang, P.; Xie, H.; Hou, L.; Hui, Z.; Xu, Y.; Du, Q.; Zhou, X.; Su, B.; et al. Reduced mir-3127-5p expression promotes nsclc proliferation/invasion and contributes to dasatinib sensitivity via the c-abl/ras/erk pathway. Sci. Rep. 2014, 4, 6527. [Google Scholar] [CrossRef] [Green Version]
- Hsieh, C.Y.; Tsai, P.C.; Chu, C.L.; Chang, F.R.; Chang, L.S.; Wu, Y.C.; Lin, S.R. Brazilein suppresses migration and invasion of mda-mb-231 breast cancer cells. Chem. Biol. Interact. 2013, 204, 105–115. [Google Scholar] [CrossRef]
- Handayani, S.; Susidarti, R.A.; Jenie, R.I.; Meiyanto, E. Two active compounds from caesalpinia sappan l. In combination with cisplatin synergistically induce apoptosis and cell cycle arrest on widr cells. Adv. Pharm. Bull. 2017, 7, 375–380. [Google Scholar] [CrossRef] [Green Version]
- Imani, S.; Wu, R.C.; Fu, J. Microrna-34 family in breast cancer: From research to therapeutic potential. J. Cancer 2018, 9, 3765–3775. [Google Scholar] [CrossRef] [Green Version]
- Wang, D.; Ren, J.; Ren, H.; Fu, J.L.; Yu, D. Microrna-132 suppresses cell proliferation in human breast cancer by directly targeting foxa1. Acta Pharmacol. Sin. 2018, 39, 124–131. [Google Scholar] [CrossRef]
- Zhang, Z.G.; Chen, W.X.; Wu, Y.H.; Liang, H.F.; Zhang, B.X. Mir-132 prohibits proliferation, invasion, migration, and metastasis in breast cancer by targeting hn1. Biochem. Biophys. Res. Commun. 2014, 454, 109–114. [Google Scholar] [CrossRef]
- Kitada, S.; Leone, M.; Sareth, S.; Zhai, D.; Reed, J.C.; Pellecchia, M. Discovery, characterization, and structure-activity relationships studies of proapoptotic polyphenols targeting b-cell lymphocyte/leukemia-2 proteins. J. Med. Chem. 2003, 46, 4259–4264. [Google Scholar] [CrossRef]
- Fei, B.Y.; He, X.; Ma, J.; Zhang, M.; Chai, R. Foxm1 is associated with metastasis in colorectal cancer through induction of the epithelial-mesenchymal transition. Oncol. Lett. 2017, 14, 6553–6561. [Google Scholar] [CrossRef]
- Wei, P.; Zhang, N.; Wang, Y.; Li, D.; Wang, L.; Sun, X.; Shen, C.; Yang, Y.; Zhou, X.; Du, X. Foxm1 promotes lung adenocarcinoma invasion and metastasis by upregulating snail. Int. J. Biol. Sci. 2015, 11, 186–198. [Google Scholar] [CrossRef] [Green Version]
- Lu, C.; Liao, Z.; Cai, M.; Zhang, G. Microrna-320a downregulation mediates human liver cancer cell proliferation through the wnt/beta-catenin signaling pathway. Oncol. Lett. 2017, 13, 573–578. [Google Scholar] [CrossRef] [Green Version]
- Xie, F.; Yuan, Y.; Xie, L.; Ran, P.; Xiang, X.; Huang, Q.; Qi, G.; Guo, X.; Xiao, C.; Zheng, S. Mirna-320a inhibits tumor proliferation and invasion by targeting c-myc in human hepatocellular carcinoma. Onco Targets Ther. 2017, 10, 885–894. [Google Scholar] [CrossRef] [Green Version]
- Imran, M.; Salehi, B.; Sharifi-Rad, J.; Aslam Gondal, T.; Saeed, F.; Imran, A.; Shahbaz, M.; Tsouh Fokou, P.V.; Umair Arshad, M.; Khan, H.; et al. Kaempferol: A key emphasis to its anticancer potential. Molecules 2019, 24, 2277. [Google Scholar] [CrossRef] [Green Version]
- Kim, T.W.; Lee, S.Y.; Kim, M.; Cheon, C.; Ko, S.G. Kaempferol induces autophagic cell death via ire1-jnk-chop pathway and inhibition of g9a in gastric cancer cells. Cell Death Dis. 2018, 9, 875. [Google Scholar] [CrossRef]
- Wang, X.; Song, Y. Microrna-340 inhibits the growth and invasion of angiosarcoma cells by targeting sirt7. Biomed. Pharmacother. 2018, 103, 1061–1068. [Google Scholar] [CrossRef]
- Li, H.; Tian, Z.; Qu, Y.; Yang, Q.; Guan, H.; Shi, B.; Ji, M.; Hou, P. Sirt7 promotes thyroid tumorigenesis through phosphorylation and activation of akt and p70s6k1 via dbc1/sirt1 axis. Oncogene 2019, 38, 345–359. [Google Scholar] [CrossRef]
- Wang, J.; Liu, Z.; Hu, T.; Han, L.; Yu, S.; Yao, Y.; Ruan, Z.; Tian, T.; Huang, T.; Wang, M.; et al. Nrf2 promotes progression of non-small cell lung cancer through activating autophagy. Cell Cycle 2017, 16, 1053–1062. [Google Scholar] [CrossRef]
- Jiang, W.; Xu, Z.; Yu, L.; Che, J.; Zhang, J.; Yang, J. Microrna-144-3p suppressed tgf-beta1-induced lung cancer cell invasion and adhesion by regulating the src-akt-erk pathway. Cell Biol. Int. 2019, 44, 51–61. [Google Scholar] [CrossRef]
- Zhao, Y.; Xie, Z.; Lin, J.; Liu, P. Mir-144-3p inhibits cell proliferation and induces apoptosis in multiple myeloma by targeting c-met. Am. J. Transl. Res. 2017, 9, 2437–2446. [Google Scholar]
- Elahouel, R.; Blanc, C.; Carpentier, G.; Frechault, S.; Cascone, I.; Destouches, D.; Delbe, J.; Courty, J.; Hamma-Kourbali, Y. Pleiotrophin exerts its migration and invasion effect through the neuropilin-1 pathway. Neoplasia 2015, 17, 613–624. [Google Scholar] [CrossRef] [Green Version]
- Song, L.; Gong, H.; Lin, C.; Wang, C.; Liu, L.; Wu, J.; Li, M.; Li, J. Flotillin-1 promotes tumor necrosis factor-alpha receptor signaling and activation of nf-kappab in esophageal squamous cell carcinoma cells. Gastroenterology 2012, 143, 995–1005.e12. [Google Scholar] [CrossRef]
- Wu, Z.; Owens, C.; Chandra, N.; Popovic, K.; Conaway, M.; Theodorescu, D. Ralbp1 is necessary for metastasis of human cancer cell lines. Neoplasia 2010, 12, 1003–1012. [Google Scholar] [CrossRef] [Green Version]
- Lee, S.; Wurtzel, J.G.; Singhal, S.S.; Awasthi, S.; Goldfinger, L.E. Ralbp1/rlip76 depletion in mice suppresses tumor growth by inhibiting tumor neovascularization. Cancer Res. 2012, 72, 5165–5173. [Google Scholar] [CrossRef] [Green Version]
- Stuckler, D.; Singhal, J.; Singhal, S.S.; Yadav, S.; Awasthi, Y.C.; Awasthi, S. Rlip76 transports vinorelbine and mediates drug resistance in non-small cell lung cancer. Cancer Res. 2005, 65, 991–998. [Google Scholar]
- Liu, N.; Du, C.H. Rlip76 silencing inhibits cell proliferation and invasion in melanoma cell line a375. Eur. Rev. Med. Pharmacol. Sci. 2017, 21, 2054–2060. [Google Scholar]
- Pham, T.N.D.; Stempel, S.; Shields, M.A.; Spaulding, C.; Kumar, K.; Bentrem, D.J.; Matsangou, M.; Munshi, H.G. Quercetin enhances the anti-tumor effects of bet inhibitors by suppressing hnrnpa1. Int. J. Mol. Sci. 2019, 20, 4293. [Google Scholar] [CrossRef] [Green Version]
- Tummala, R.; Lou, W.; Gao, A.C.; Nadiminty, N. Quercetin targets hnrnpa1 to overcome enzalutamide resistance in prostate cancer cells. Mol. Cancer. Ther. 2017, 16, 2770–2779. [Google Scholar] [CrossRef] [Green Version]
- Puca, F.; Tosti, N.; Federico, A.; Kuzay, Y.; Pepe, A.; Morlando, S.; Savarese, T.; D’Alessio, F.; Colamaio, M.; Sarnataro, D.; et al. Hmga1 negatively regulates numb expression at transcriptional and post transcriptional level in glioblastoma stem cells. Cell Cycle 2019, 18, 1446–1457. [Google Scholar] [CrossRef]
- Akaboshi, S.; Watanabe, S.; Hino, Y.; Sekita, Y.; Xi, Y.; Araki, K.; Yamamura, K.; Oshima, M.; Ito, T.; Baba, H.; et al. Hmga1 is induced by wnt/beta-catenin pathway and maintains cell proliferation in gastric cancer. Am. J. Pathol. 2009, 175, 1675–1685. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sun, X.; Xu, C.; Tang, S.C.; Wang, J.; Wang, H.; Wang, P.; Du, N.; Qin, S.; Li, G.; Xu, S.; et al. Let-7c blocks estrogen-activated wnt signaling in induction of self-renewal of breast cancer stem cells. Cancer Gene Ther. 2016, 23, 83–89. [Google Scholar] [CrossRef]
- Nadiminty, N.; Tummala, R.; Liu, C.; Lou, W.; Evans, C.P.; Gao, A.C. Nf-kappab2/p52:C-myc:Hnrnpa1 pathway regulates expression of androgen receptor splice variants and enzalutamide sensitivity in prostate cancer. Mol. Cancer Ther. 2015, 14, 1884–1895. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yamamura, S.; Saini, S.; Majid, S.; Hirata, H.; Ueno, K.; Deng, G.; Dahiya, R. Microrna-34a modulates c-myc transcriptional complexes to suppress malignancy in human prostate cancer cells. PLoS ONE 2012, 7, e29722. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Teng, K.Y.; Barajas, J.M.; Hu, P.; Jacob, S.T.; Ghoshal, K. Role of b cell lymphoma 2 in the regulation of liver fibrosis in mir-122 knockout mice. Biology (Basel) 2020, 9, 157. [Google Scholar] [CrossRef] [PubMed]
- Hu, H.; Xu, Z.; Li, C.; Xu, C.; Lei, Z.; Zhang, H.T.; Zhao, J. Mir-145 and mir-203 represses tgf-beta-induced epithelial-mesenchymal transition and invasion by inhibiting smad3 in non-small cell lung cancer cells. Lung Cancer 2016, 97, 87–94. [Google Scholar] [CrossRef] [PubMed]
- Zhou, Y.; Liang, H.; Liao, Z.; Wang, Y.; Hu, X.; Chen, X.; Xu, L.; Hu, Z. Mir-203 enhances let-7 biogenesis by targeting lin28b to suppress tumor growth in lung cancer. Sci. Rep. 2017, 7, 42680. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Paranjape, A.N.; Balaji, S.A.; Mandal, T.; Krushik, E.V.; Nagaraj, P.; Mukherjee, G.; Rangarajan, A. Bmi1 regulates self-renewal and epithelial to mesenchymal transition in breast cancer cells through nanog. BMC Cancer 2014, 14, 785. [Google Scholar] [CrossRef] [Green Version]
- Rao, P.K.; Missiaglia, E.; Shields, L.; Hyde, G.; Yuan, B.; Shepherd, C.J.; Shipley, J.; Lodish, H.F. Distinct roles for mir-1 and mir-133a in the proliferation and differentiation of rhabdomyosarcoma cells. FASEB J. 2010, 24, 3427–3437. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Taulli, R.; Bersani, F.; Foglizzo, V.; Linari, A.; Vigna, E.; Ladanyi, M.; Tuschl, T.; Ponzetto, C. The muscle-specific microrna mir-206 blocks human rhabdomyosarcoma growth in xenotransplanted mice by promoting myogenic differentiation. J. Clin. Invest. 2009, 119, 2366–2378. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhang, X.Q.; Yao, C.; Bian, W.H.; Chen, X.; Xue, J.X.; Zhu, Z.Y.; Ying, Y.; Xu, Y.L.; Wang, C. Effects of astragaloside iv on treatment of breast cancer cells execute possibly through regulation of nrf2 via pi3k/akt/mtor signaling pathway. Food Sci. Nutr. 2019, 7, 3403–3413. [Google Scholar] [CrossRef] [PubMed]
- Zhu, H.X.; Shi, L.; Zhang, Y.; Zhu, Y.C.; Bai, C.X.; Wang, X.D.; Zhou, J.B. Myocyte enhancer factor 2d provides a cross-talk between chronic inflammation and lung cancer. J. Transl. Med. 2017, 15, 65. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wu, Y.; Wang, Y.; Lin, Y.; Liu, Y.; Wang, Y.; Jia, J.; Singh, P.; Chi, Y.I.; Wang, C.; Dong, C.; et al. Dub3 inhibition suppresses breast cancer invasion and metastasis by promoting snail1 degradation. Nat. Commun. 2017, 8, 14228. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tang, J.; Zhong, G.; Wu, J.; Chen, H.; Jia, Y. Sox2 recruits klf4 to regulate nasopharyngeal carcinoma proliferation via pi3k/akt signaling. Oncogenesis 2018, 7, 61. [Google Scholar] [CrossRef] [PubMed]
- Kfoury, A.; Le Corf, K.; El Sabeh, R.; Journeaux, A.; Badran, B.; Hussein, N.; Lebecque, S.; Manie, S.; Renno, T.; Coste, I. Myd88 in DNA repair and cancer cell resistance to genotoxic drugs. J. Natl. Cancer Inst. 2013, 105, 937–946. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Echizen, K.; Hirose, O.; Maeda, Y.; Oshima, M. Inflammation in gastric cancer: Interplay of the cox-2/prostaglandin e2 and toll-like receptor/myd88 pathways. Cancer Sci. 2016, 107, 391–397. [Google Scholar] [CrossRef] [Green Version]
- Phuah, N.H.; Azmi, M.N.; Awang, K.; Nagoor, N.H. Down-regulation of microrna-210 confers sensitivity towards 1’s-1’-acetoxychavicol acetate (aca) in cervical cancer cells by targeting smad4. Mol. Cells 2017, 40, 291–298. [Google Scholar] [CrossRef] [Green Version]
- Phuah, N.H.; Azmi, M.N.; Awang, K.; Nagoor, N.H. Suppression of microrna-629 enhances sensitivity of cervical cancer cells to 1’s-1’-acetoxychavicol acetate via regulating rsu1. Onco Targets Ther. 2017, 10, 1695–1705. [Google Scholar] [CrossRef] [Green Version]
- Chakrabarti, M.; Ray, S.K. Anti-tumor activities of luteolin and silibinin in glioblastoma cells: Overexpression of mir-7-1-3p augmented luteolin and silibinin to inhibit autophagy and induce apoptosis in glioblastoma in vivo. Apoptosis 2016, 21, 312–328. [Google Scholar] [CrossRef]
- Xue, C.L.; Liu, H.G.; Li, B.Y.; He, S.H.; Yue, Q.F. Physcion 8-o-beta-glucopyranoside exhibits anti-growth and anti-metastatic activities in ovarian cancer by downregulating mir-25. Eur. Rev. Med. Pharmacol. Sci. 2019, 23, 5101–5112. [Google Scholar]
- Jiang, L.; Tao, C.; He, A.; He, X. Overexpression of mir-126 sensitizes osteosarcoma cells to apoptosis induced by epigallocatechin-3-gallate. World J. Surg. Oncol. 2014, 12, 383. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chakrabarti, M.; Ray, S.K. Direct transfection of mir-137 mimics is more effective than DNA demethylation of mir-137 promoter to augment anti-tumor mechanisms of delphinidin in human glioblastoma u87mg and ln18 cells. Gene 2015, 573, 141–152. [Google Scholar] [CrossRef] [PubMed]
- Chakrabarti, M.; Banik, N.L.; Ray, S.K. Mir-138 overexpression is more powerful than htert knockdown to potentiate apigenin for apoptosis in neuroblastoma in vitro and in vivo. Exp. Cell Res. 2013, 319, 1575–1585. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhu, X.; Li, H.; Long, L.; Hui, L.; Chen, H.; Wang, X.; Shen, H.; Xu, W. Mir-126 enhances the sensitivity of non-small cell lung cancer cells to anticancer agents by targeting vascular endothelial growth factor A. Acta Biochim. Biophys. Sin. (Shanghai) 2012, 44, 519–526. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhang, W.; Zhou, J.; Zhu, X.; Yuan, H. Mir-126 reverses drug resistance to trail through inhibiting the expression of c-flip in cervical cancer. Gene 2017, 627, 420–427. [Google Scholar] [CrossRef]
- Lechman, E.R.; Gentner, B.; Ng, S.W.; Schoof, E.M.; van Galen, P.; Kennedy, J.A.; Nucera, S.; Ciceri, F.; Kaufmann, K.B.; Takayama, N.; et al. Mir-126 regulates distinct self-renewal outcomes in normal and malignant hematopoietic stem cells. Cancer Cell 2016, 29, 214–228. [Google Scholar] [CrossRef] [Green Version]
- Zhang, H.; Zuo, Z.; Lu, X.; Wang, L.; Wang, H.; Zhu, Z. Mir-25 regulates apoptosis by targeting bim in human ovarian cancer. Oncol. Rep. 2012, 27, 594–598. [Google Scholar]
- Wang, F.; Mayca Pozo, F.; Tian, D.; Geng, X.; Yao, X.; Zhang, Y.; Tang, J. Shikonin inhibits cancer through p21 upregulation and apoptosis induction. Front. Pharmacol. 2020, 11, 861. [Google Scholar] [CrossRef]
- Kim, H.J.; Hwang, K.E.; Park, D.S.; Oh, S.H.; Jun, H.Y.; Yoon, K.H.; Jeong, E.T.; Kim, H.R.; Kim, Y.S. Shikonin-induced necroptosis is enhanced by the inhibition of autophagy in non-small cell lung cancer cells. J. Transl. Med. 2017, 15, 123. [Google Scholar] [CrossRef]
- Wiench, B.; Eichhorn, T.; Paulsen, M.; Efferth, T. Shikonin directly targets mitochondria and causes mitochondrial dysfunction in cancer cells. Evid. Based Complement. Alternat. Med. 2012, 2012, 726025. [Google Scholar] [CrossRef] [Green Version]
- Chen, J.C.; Hsieh, Y.Y.; Lo, H.L.; Li, A.; Chou, C.J.; Yang, P.M. In vitro and in silico mechanistic insights into mir-21-5p-mediated topoisomerase drug resistance in human colorectal cancer cells. Biomolecules 2019, 9, 467. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ni, W.; Luo, L.; Zuo, P.; Li, R.; Xu, X.; Wen, F.; Hu, D. Mir-374a inhibitor enhances etoposide-induced cytotoxicity against glioma cells through upregulation of foxo1. Oncol. Res. 2019, 27, 703–712. [Google Scholar] [CrossRef] [PubMed]
- Kollinerova, S.; Dostal, Z.; Modriansky, M. Microrna hsa-mir-29b potentiates etoposide toxicity in hela cells via down-regulation of mcl-1. Toxicol. In Vitro 2017, 40, 289–296. [Google Scholar] [CrossRef] [PubMed]
- Wang, H.; Zhao, F.; Cai, S.; Pu, Y. Mir-193a regulates chemoresistance of human osteosarcoma cells via repression of irs2. J. Bone Oncol. 2019, 17, 100241. [Google Scholar] [CrossRef]
- Rebucci, M.; Sermeus, A.; Leonard, E.; Delaive, E.; Dieu, M.; Fransolet, M.; Arnould, T.; Michiels, C. Mirna-196b inhibits cell proliferation and induces apoptosis in hepg2 cells by targeting igf2bp1. Mol. Cancer 2015, 14, 79. [Google Scholar] [CrossRef] [Green Version]
- Tao, L.; Wu, Y.Q.; Zhang, S.P. Mir-21-5p enhances the progression and paclitaxel resistance in drug-resistant breast cancer cell lines by targeting pdcd4. Neoplasma 2019, 66, 746–755. [Google Scholar] [CrossRef]
- Feng, L.; Shen, F.; Zhou, J.; Li, Y.; Jiang, R.; Chen, Y. Hypoxia-induced up-regulation of mir-27a promotes paclitaxel resistance in ovarian cancer. Biosci. Rep. 2020, 40, BSR20192457. [Google Scholar] [CrossRef] [Green Version]
- Li, Z.; Hu, S.; Wang, J.; Cai, J.; Xiao, L.; Yu, L.; Wang, Z. Mir-27a modulates mdr1/p-glycoprotein expression by targeting hipk2 in human ovarian cancer cells. Gynecol. Oncol. 2010, 119, 125–130. [Google Scholar] [CrossRef]
- Zhou, M.; Liu, Z.; Zhao, Y.; Ding, Y.; Liu, H.; Xi, Y.; Xiong, W.; Li, G.; Lu, J.; Fodstad, O.; et al. Microrna-125b confers the resistance of breast cancer cells to paclitaxel through suppression of pro-apoptotic bcl-2 antagonist killer 1 (bak1) expression. J. Biol. Chem. 2010, 285, 21496–21507. [Google Scholar] [CrossRef] [Green Version]
- Zhao, K.; Li, X.; Chen, X.; Zhu, Q.; Yin, F.; Ruan, Q.; Xia, J.; Niu, Z. Inhibition of mir-140-3p or mir-155-5p by antagomir treatment sensitize chordoma cells to chemotherapy drug treatment by increasing pten expression. Eur. J. Pharmacol. 2019, 854, 298–306. [Google Scholar] [CrossRef]
- Filipska, M.; Skrzypski, M.; Czetyrbok, K.; Stokowy, T.; Stasilojc, G.; Supernat, A.; Jassem, J.; Zaczek, A.J.; Bigda, J. Mir-192 and mir-662 enhance chemoresistance and invasiveness of squamous cell lung carcinoma. Lung Cancer 2018, 118, 111–118. [Google Scholar] [CrossRef] [PubMed]
- Ren, L.L.; Yan, T.T.; Shen, C.Q.; Tang, J.Y.; Kong, X.; Wang, Y.C.; Chen, J.; Liu, Q.; He, J.; Zhong, M.; et al. The distinct role of strand-specific mir-514b-3p and mir-514b-5p in colorectal cancer metastasis. Cell Death Dis. 2018, 9, 687. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Geng, W.; Song, H.; Zhao, Q.; Dong, K.; Pu, Q.; Gao, H.; Lv, Y. Mir-520h stimulates drug resistance to paclitaxel by targeting the otud3-pten axis in breast cancer. Biomed. Res. Int. 2020, 2020, 9512793. [Google Scholar] [CrossRef] [PubMed]
- Hou, X.; Niu, Z.; Liu, L.; Guo, Q.; Li, H.; Yang, X.; Zhang, X. Mir-1207-5p regulates the sensitivity of triple-negative breast cancer cells to taxol treatment via the suppression of lzts1 expression. Oncol. Lett. 2019, 17, 990–998. [Google Scholar] [CrossRef] [PubMed]
- Sun, H.; Zhou, X.; Bao, Y.; Xiong, G.; Cui, Y.; Zhou, H. Involvement of mir-4262 in paclitaxel resistance through the regulation of pten in non-small cell lung cancer. Open Biol. 2019, 9, 180227. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Deng, L.M.; Tan, T.; Zhang, T.Y.; Xiao, X.F.; Gu, H. Mir1 reverses multidrug resistance in gastric cancer cells via downregulation of sorcin through promoting the accumulation of intracellular drugs and apoptosis of cells. Int. J. Oncol. 2019, 55, 451–461. [Google Scholar]
- Hong, T.; Ding, J.; Li, W. Mir-7 reverses breast cancer resistance to chemotherapy by targeting mrp1 and bcl2. Onco Targets Ther. 2019, 12, 11097–11105. [Google Scholar] [CrossRef] [Green Version]
- Huang, L.; Hu, C.; Chao, H.; Wang, R.; Lu, H.; Li, H.; Chen, H. Mir-29c regulates resistance to paclitaxel in nasopharyngeal cancer by targeting itgb1. Exp. Cell Res. 2019, 378, 1–10. [Google Scholar] [CrossRef]
- Yang, G.; Fu, Y.; Lu, X.; Wang, M.; Dong, H.; Li, Q. Mir34a regulates the chemosensitivity of retinoblastoma cells via modulation of magea/p53 signaling. Int. J. Oncol. 2019, 54, 177–187. [Google Scholar]
- Wang, P.; Li, Z.; Liu, H.; Zhou, D.; Fu, A.; Zhang, E. Microrna-126 increases chemosensitivity in drug-resistant gastric cancer cells by targeting ezh2. Biochem. Biophys. Res. Commun. 2016, 479, 91–96. [Google Scholar] [CrossRef]
- Chen, M.; Li, D.; Gong, N.; Wu, H.; Su, C.; Xie, C.; Xiang, H.; Lin, C.; Li, X. Mir-133b down-regulates abcc1 and enhances the sensitivity of crc to anti-tumor drugs. Oncotarget 2017, 8, 52983–52994. [Google Scholar] [CrossRef] [Green Version]
- Guan, X.; Guan, Y. Mir-145-5p attenuates paclitaxel resistance and suppresses the progression in drug-resistant breast cancer cell lines. Neoplasma 2020, 67, 972–981. [Google Scholar] [CrossRef]
- Juang, V.; Chang, C.H.; Wang, C.S.; Wang, H.E.; Lo, Y.L. Ph-responsive peg-shedding and targeting peptide-modified nanoparticles for dual-delivery of irinotecan and microrna to enhance tumor-specific therapy. Small 2019, 15, e1903296. [Google Scholar] [CrossRef]
- Zhu, W.; Xu, H.; Zhu, D.; Zhi, H.; Wang, T.; Wang, J.; Jiang, B.; Shu, Y.; Liu, P. Mir-200bc/429 cluster modulates multidrug resistance of human cancer cell lines by targeting bcl2 and xiap. Cancer Chemother. Pharmacol. 2012, 69, 723–731. [Google Scholar] [CrossRef] [PubMed]
- Liu, X.; Heng, C.; Li, Y.; Yu, L. Mir-302a sensitizes leukemia cells to etoposide by targeting rad52. Oncotarget 2017, 8, 73884–73891. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wang, C.; Su, K.; Zhang, Y.; Zhang, W.; Chu, D.; Zhao, Q.; Guo, R. Microrna-365 targets multiple oncogenes to inhibit proliferation, invasion, and self-renewal of aggressive endometrial cancer cells. Cancer Manag. Res. 2018, 10, 5171–5185. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jiang, J.; Xie, C.; Liu, Y.; Shi, Q.; Chen, Y. Up-regulation of mir-383-5p suppresses proliferation and enhances chemosensitivity in ovarian cancer cells by targeting trim27. Biomed. Pharmacother. 2019, 109, 595–601. [Google Scholar] [CrossRef]
- Chen, S.; Wu, J.; Jiao, K.; Wu, Q.; Ma, J.; Chen, D.; Kang, J.; Zhao, G.; Shi, Y.; Fan, D.; et al. Microrna-495-3p inhibits multidrug resistance by modulating autophagy through grp78/mtor axis in gastric cancer. Cell Death Dis. 2018, 9, 1070. [Google Scholar] [CrossRef]
- Lyu, H.; Wang, S.; Huang, J.; Wang, B.; He, Z.; Liu, B. Survivin-targeting mir-542-3p overcomes her3 signaling-induced chemoresistance and enhances the antitumor activity of paclitaxel against her2-overexpressing breast cancer. Cancer Lett. 2018, 420, 97–108. [Google Scholar] [CrossRef]
- Abdelfattah, N.; Rajamanickam, S.; Panneerdoss, S.; Timilsina, S.; Yadav, P.; Onyeagucha, B.C.; Garcia, M.; Vadlamudi, R.; Chen, Y.; Brenner, A.; et al. Mir-584-5p potentiates vincristine and radiation response by inducing spindle defects and DNA damage in medulloblastoma. Nat. Commun. 2018, 9, 4541. [Google Scholar] [CrossRef] [Green Version]
- Xue, J.; Chi, Y.; Chen, Y.; Huang, S.; Ye, X.; Niu, J.; Wang, W.; Pfeffer, L.M.; Shao, Z.M.; Wu, Z.H.; et al. Mirna-621 sensitizes breast cancer to chemotherapy by suppressing fbxo11 and enhancing p53 activity. Oncogene 2016, 35, 448–458. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Huang, H.; Tang, J.; Zhang, L.; Bu, Y.; Zhang, X. Mir-874 regulates multiple-drug resistance in gastric cancer by targeting atg16l1. Int. J. Oncol. 2018, 53, 2769–2779. [Google Scholar] [CrossRef] [Green Version]
- Jia, M.; Wei, Z.; Liu, P.; Zhao, X. Silencing of abcg2 by microrna-3163 inhibits multidrug resistance in retinoblastoma cancer stem cells. J. Korean Med. Sci. 2016, 31, 836–842. [Google Scholar] [CrossRef]
- Kannathasan, T.; Kuo, W.W.; Chen, M.C.; Viswanadha, V.P.; Shen, C.Y.; Tu, C.C.; Yeh, Y.L.; Bharath, M.; Shibu, M.A.; Huang, C.Y. Chemoresistance-associated silencing of mir-4454 promotes colorectal cancer aggression through the gnl3l and nf-kappab pathway. Cancers (Basel) 2020, 12, 1231. [Google Scholar] [CrossRef]
- Liu, M.; Gong, C.; Xu, R.; Chen, Y.; Wang, X. Microrna-5195-3p enhances the chemosensitivity of triple-negative breast cancer to paclitaxel by downregulating eif4a2. Cell Mol. Biol. Lett. 2019, 24, 47. [Google Scholar] [CrossRef] [PubMed]
- Nogueira, A.; Fernandes, M.; Catarino, R.; Medeiros, R. Rad52 functions in homologous recombination and its importance on genomic integrity maintenance and cancer therapy. Cancers (Basel) 2019, 11, 1622. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Westover, D.; Li, F. New trends for overcoming abcg2/bcrp-mediated resistance to cancer therapies. J. Exp. Clin. Cancer Res. 2015, 34, 159. [Google Scholar] [CrossRef] [Green Version]
- Olszewski, U.; Liedauer, R.; Ausch, C.; Thalhammer, T.; Hamilton, G. Overexpression of cyp3a4 in a colo 205 colon cancer stem cell model in vitro. Cancers 2011, 3, 1467–1479. [Google Scholar] [CrossRef] [PubMed]
- He, Y.; Liu, X. The tumor-suppressor gene lzts1 suppresses hepatocellular carcinoma proliferation by impairing pi3k/akt pathway. Biomed. Pharmacother. 2015, 76, 141–146. [Google Scholar] [CrossRef] [PubMed]
- Aoudjit, F.; Vuori, K. Integrin signaling inhibits paclitaxel-induced apoptosis in breast cancer cells. Oncogene 2001, 20, 4995–5004. [Google Scholar] [CrossRef] [Green Version]
- Ma, L.; Yao, N.; Chen, P.; Zhuang, Z. Trim27 promotes the development of esophagus cancer via regulating pten/akt signaling pathway. Cancer Cell Int. 2019, 19, 283. [Google Scholar] [CrossRef] [PubMed]
- Gonzalez, M.E.; Moore, H.M.; Li, X.; Toy, K.A.; Huang, W.; Sabel, M.S.; Kidwell, K.M.; Kleer, C.G. Ezh2 expands breast stem cells through activation of notch1 signaling. Proc. Natl. Acad. Sci. USA 2014, 111, 3098–3103. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Muhammad, N.; Bhattacharya, S.; Steele, R.; Phillips, N.; Ray, R.B. Involvement of c-fos in the promotion of cancer stem-like cell properties in head and neck squamous cell carcinoma. Clin. Cancer Res. 2017, 23, 3120–3128. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gao, H.; Teng, C.; Huang, W.; Peng, J.; Wang, C. Sox2 promotes the epithelial to mesenchymal transition of esophageal squamous cells by modulating slug expression through the activation of stat3/hif-alpha signaling. Int. J. Mol. Sci. 2015, 16, 21643–21657. [Google Scholar] [CrossRef] [Green Version]
- Yamagishi, N.; Nakao, R.; Kondo, R.; Nishitsuji, M.; Saito, Y.; Kuga, T.; Hatayama, T.; Nakayama, Y. Increased expression of sorcin is associated with multidrug resistance in leukemia cells via up-regulation of mdr1 expression through camp response element-binding protein. Biochem. Biophys. Res. Commun. 2014, 448, 430–436. [Google Scholar] [CrossRef]
- Xu, P.; Jiang, Y.F.; Wang, J.H. Shrna-mediated silencing of sorcin increases drug chemosensitivity in myeloma km3/ddp and u266/adm cell lines. Int. J. Clin. Exp. Pathol. 2015, 8, 2300–2310. [Google Scholar]
- Nam, J.S.; Sharma, A.R.; Nguyen, L.T.; Chakraborty, C.; Sharma, G.; Lee, S.S. Application of bioactive quercetin in oncotherapy: From nutrition to nanomedicine. Molecules 2016, 21, 108. [Google Scholar] [CrossRef] [Green Version]
- Alles, J.; Fehlmann, T.; Fischer, U.; Backes, C.; Galata, V.; Minet, M.; Hart, M.; Abu-Halima, M.; Grasser, F.A.; Lenhof, H.P.; et al. An estimate of the total number of true human mirnas. Nucleic Acids Res. 2019, 47, 3353–3364. [Google Scholar] [CrossRef] [Green Version]
- Matarlo, J.S.; Krumpe, L.R.H.; Heinz, W.F.; Oh, D.; Shenoy, S.R.; Thomas, C.L.; Goncharova, E.I.; Lockett, S.J.; O’Keefe, B.R. The natural product butylcycloheptyl prodiginine binds pre-mir-21, inhibits dicer-mediated processing of pre-mir-21, and blocks cellular proliferation. Cell Chem. Biol. 2019, 26, 1133–1142.e4. [Google Scholar] [CrossRef]
- Li, Z.; Rana, T.M. Therapeutic targeting of micrornas: Current status and future challenges. Nat. Rev. Drug Discov. 2014, 13, 622–638. [Google Scholar] [CrossRef]
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
Figure 1.
Effects of nitrogen-containing compounds and organosulfur/phytosterol compounds on the expression level of oncogenic miRNAs (red) and tumor-suppressive miRNAs (blue). Arrows indicate the upregulation (↑) and downregulation (↓) of miRNA levels and consequential effects on cancer. The role of miRNAs in cancer therapy with phytochemicals is described in Section 2 and Section 3.
Figure 1.
Effects of nitrogen-containing compounds and organosulfur/phytosterol compounds on the expression level of oncogenic miRNAs (red) and tumor-suppressive miRNAs (blue). Arrows indicate the upregulation (↑) and downregulation (↓) of miRNA levels and consequential effects on cancer. The role of miRNAs in cancer therapy with phytochemicals is described in Section 2 and Section 3.

Figure 2.
Effects of phenolic compounds on the expression level of oncogenic miRNAs (red) and tumor-suppressive miRNAs (blue). Arrows indicate the upregulation (↑) and downregulation (↓) of miRNA levels and consequential effects on cancer. The role of miRNAs in cancer therapy with phytochemicals is described in Section 2 and Section 3.
Figure 2.
Effects of phenolic compounds on the expression level of oncogenic miRNAs (red) and tumor-suppressive miRNAs (blue). Arrows indicate the upregulation (↑) and downregulation (↓) of miRNA levels and consequential effects on cancer. The role of miRNAs in cancer therapy with phytochemicals is described in Section 2 and Section 3.

Figure 3.
Effects of terpenoid compounds on the expression level of oncogenic miRNAs (red) and tumor-suppressive miRNAs (blue). Arrows indicate the upregulation (↑) and downregulation (↓) of miRNA levels and consequential effects on cancer. The role of miRNAs in cancer therapy with phytochemicals is described in Section 2 and Section 3.
Figure 3.
Effects of terpenoid compounds on the expression level of oncogenic miRNAs (red) and tumor-suppressive miRNAs (blue). Arrows indicate the upregulation (↑) and downregulation (↓) of miRNA levels and consequential effects on cancer. The role of miRNAs in cancer therapy with phytochemicals is described in Section 2 and Section 3.

Figure 4.
MiRNAs regulating the effectiveness of phytochemicals in cancer. MiRNAs can either sensitize (blue) or desensitize (red) cancer cells to various phytochemicals. It is described in Section 4.
Figure 4.
MiRNAs regulating the effectiveness of phytochemicals in cancer. MiRNAs can either sensitize (blue) or desensitize (red) cancer cells to various phytochemicals. It is described in Section 4.

Figure 5.
MiRNAs regulating the efficacy of plant-derived anti-cancer agents in cancer. Resistance-promoting and -suppressing miRNAs are indicated in red and blue colors, respectively. It is described in Section 5.
Figure 5.
MiRNAs regulating the efficacy of plant-derived anti-cancer agents in cancer. Resistance-promoting and -suppressing miRNAs are indicated in red and blue colors, respectively. It is described in Section 5.


{kind=link}
{kind=link}
{kind=link}
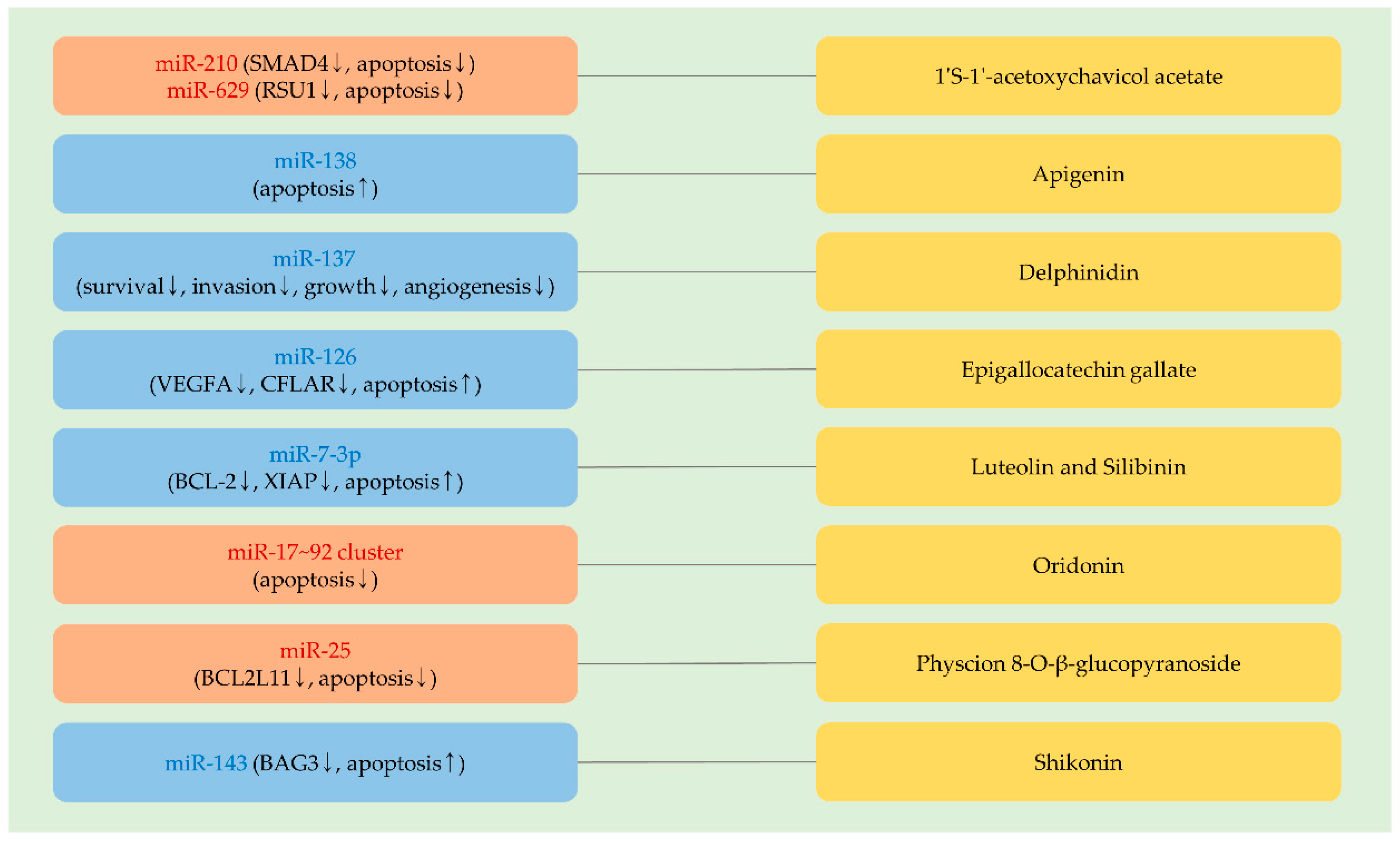{kind=link}
{kind=link}
Table 1.
The list of phytochemicals and derivatives currently tested in preclinical studies and clinical trials.
Table 1.
The list of phytochemicals and derivatives currently tested in preclinical studies and clinical trials.
| Phytochemical | Source of Phytochemical (NCT Number, Condition/Disease, Recruitment Status if Applicable) | Ref. |
|---|---|---|
| Nitrogen-containing compounds (alkaloids and amino acids) | ||
| 5-Aminolevulinic acid * | A precursor of tetrapyrroles in higher plants (NCT02144077, basal cell carcinoma, active) | [16] |
| Berberine * | An isoquinoline alkaloid in plants, such as barberry and goldenseal (NCT03281096, colorectal adenomas, recruiting) | [17] |
| Coptisine | A protoberberine alkaloid in Rhizoma coptidis | [18] |
| Evodiamine | An indoloquinazoline alkaloid in Evodia rutaecarpa | [19] |
| Indole-3-carbinol | An indole alkaloid in vegetables such as cauliflower | [20] |
| Matrine | A quinolizidine alkaloid in the root of Sophora flavescens | [21] |
| Neferine | A bisbenzylisoquinline alkaloid in the seed embryo of Nelumbo nucifera | [22] |
| Nitidine chloride | A benzophenanthridine alkaloid in the roots of Zanthoxylum nitidum | [23] |
| Piperlongumine | An alkaloid from Piper longum Linn | [24] |
| Sanguinarine | A benzophenanthridine alkaloid in Papaveraceae plants | [25] |
| Sinomenine | An isoquinoline alkaloid in the dry roots and stems of Sinomenium acutum | [26] |
| Sophocarpine | A tetracyclic quinolizidine alkaloid in Sophora alopecuroides Linn | [27] |
| Swainsonine | An indolizidine alkaloid in Swainsona canescens | [28] |
| Organosulfur and phytosterol compounds | ||
| Allicin | An organosulfur compound from garlic (Allium sativum) | [29] |
| Sulforaphane * | An isothiocyanate abundant in cruciferous vegetables such as broccoli sprouts (NCT03182959, head and neck cancer, active) | [30] |
| β-sitosterol-d-glucoside | A phytosterol from sweet potato | [31] |
| Phenolic compounds | ||
| 1′S-1′-acetoxychavicol acetate | A phenylpropanoid from Alpinia conchigera | [32] |
| Apigenin | A flavone from fruits (e.g., oranges) and vegetables (e.g., onions) | [33] |
| Astragalin | A flavonoid broadly present in food such as lotus leaves | [34] |
| Baicalein(aglycone of baicalin) | A flavonoid from Scutellaria radix | [35] |
| Baicalin | A flavonoid from the root of Scutellaria baicalensis Georgi | [36] |
| Brazilein | A polyphenolic compound from Caesalpinia sappan | [37] |
| Chrysin | A flavone from several plants, including Oroxylum indicum | [38] |
| Curcumin * | A polyphenolic curcuminoid in Curcuma longa (turmeric plant) (NCT02064673, prostate cancer, recruiting) | [39] |
| Delphinidin | A flavonoid in fruits and vegetables such as tomatoes | [40] |
| Epigallocatechin gallate * | A polyphenol in green tea (NCT02891538, colorectal cancer, recruiting) | [41] |
| Formononetin | An isoflavone in the root of Astragalus membranaceus | [42] |
| Galangin | A flavonoid in Alpinia officinarum | [43] |
| Gambogic acid | A xanthonoid from Garcinia hanburyi trees | [44] |
| Genistein * | An isoflavone and phytoestrogen primarily in Soybeans (NCT00118040, bladder cancer, active) | [45] |
| Gossypol (AT-101) * | A polyphenol from cotton roots and seeds (NCT01633541, advanced laryngeal cancer, active) | [46] |
| Hydroxygenkwanin | A flavonoid from Daphne genkwa | [47] |
| Honokiol | A polyphenol in the genus Magnolia | [48] |
| Icariin | A flavonoid in Epimedium (Horny Goat Weed and Yin Yang Huo) | [49] |
| Isoliquiritigenin | A chalcone from Glycyrrhizae radix | [50] |
| Isorhapontigenin | A stilbene from Gnetum cleistostachyum | [51] |
| Kaempferol | A flavonoid in various plants such as Brussels sprouts | [52] |
| Licochalcone A | A chalcone from Glycyrrhiza uralensis | [53] |
| Luteolin | A flavonoid in several vegetables such as cabbage | [54] |
| Physcion 8-O-β-glucopyranoside | An anthraquinone in Rumex japonicus Houtt | [55] |
| Procyanidin | A polyphenol in dietary fruits such as grapes | [56] |
| Puerarin | An isoflavone in the root of Pueraria (Radix puerariae) | [57] |
| Quercetin * | A flavonoid in fruits and vegetables, such as onions and broccoli (NCT01912820, prostate cancer, active) | [58] |
| Resveratrol | A stilbenoid in grapes and red wine, etc | [59] |
| Shikonin | A naphthoquinone from Lithospermum erythrorhizon | [60] |
| Silibinin | A polyphenolic flavonoid in milk thistle (Silybum marianum) | [61,62,63] |
| Silymarin | A mixture of flavonolignans from Silybum marianum L. Gaertn. | [64] |
| Trans-3,5,4′-trimethoxystilbene | A derivative of resveratrol | [65] |
| Terpenoid compounds | ||
| Ailanthone | A quassinoid from Ailanthus altissima | [66] |
| Andrographolide * | A diterpene lactone from Andrographis paniculate (NCT04196075, advanced esophageal cancer, recruiting) | [67] |
| Artemisinin | A sesquiterpene lactone from Artemisia annua | [68] |
| Artesunate * | A derivative of artemisinin (NCT02633098, colorectal cancer, recruiting) | [68] |
| Astragaloside IV | A pentacyclic triterpenoid from Astragali radix | [69] |
| Brucein D | A triterpene quassinoid in Brucea javanica fruit | [70] |
| Cannabidiol * | A terpenophenolic compound from Cannabis sativa (NCT04428203, prostate cancer, recruiting) | [71] |
| Celastrol | A triterpene in Tripterygium wilfordii | [72] |
| Cucurbitacin D | A derivative of cucurbitacin that is a tetracyclic triterpene from the Cucurbitaceae family | [73] |
| Curcumol | A sesquiterpenoid from Rhizoma Curcumae | [74] |
| Ginsenoside Rd and Rh2 | Triterpene saponins in Panax genus | [75,76] |
| Lycopene * | A carotenoid from fruits such as tomatoes (NCT03167268, colorectal cancer, recruiting) | [77] |
| Maytenin and 22-β-hydroxymaytenin | Quinone-methide triterpenes in Maytenus ilicifolia | [78] |
| Oleanolic acid | A pentacyclic triterpene in herbs and vegetables | [79] |
| Oridonin | A diterpenoid in genus Isodon plants | [80] |
| Pristimerin | A triterpenoid from the Celastraceae and Hippocrateaceae families | [81] |
| Toosendanin | A triterpenoid from Melia toosendan Sieb et Zucc | [82] |
| Triptolide * | A diterpene triepoxide in Tripterygium wilfordii Hook F (NCT03129139, advanced solid tumors, recruiting) | [83] |
| Tubeimoside-1 | A triterpenoid saponin from Bolbostemma paniculatum | [84] |
| Ursolic acid * | A pentacyclic triterpene in plants such as apples (NCT04403568, prostate cancer, not yet recruiting) | [85] |
| α-pinene | A monoterpene in pine needles | [86] |
* indicates phytochemical compounds currently in clinical trials on cancer. The national clinical trial (NCT) number, condition/disease, and recruitment status registered in ClinicalTrials.gov are referred.
Table 2.
Anti-cancer compounds derived from plants that are currently used in clinical practice.
| Anti-Cancer Agent | Source | Primary Anti-Cancer Action/Application * | Ref. |
|---|---|---|---|
| Etoposide | A derivative of podophyllotoxin, a non-alkaloid lignan that is isolated from Podophyllum peltatum | Topoisomerase II inhibition/Approved for small cell lung cancer and testicular cancer | [10] |
| Irinotecan | A derivative of camptothecin that is a monoterpene indole alkaloid from Camptotheca acuminata | Topoisomerase I inhibition/Approved for colorectal cancer | [9] |
| Paclitaxel | A terpenoid isolated from the Pacific yew tree | Stabilization of microtubule polymer/Approved for AIDS-related Kaposi sarcoma, breast cancer, non-small cell lung cancer, and ovarian cancer | [11] |
| Vincristine | A vinca alkaloid from Catharanthus roseus | An inhibition of microtubule polymerization/Approved for acute leukemia. Also used to treat Hodgkin lymphoma, neuroblastoma, non-Hodgkin lymphoma, rhabdomyosarcoma, and Wilms tumor | [10] |
* information on the drug application is from the National Cancer Institute.
Table 3.
Oncogenic miRNAs downregulated by phytochemicals in cancer.
| miRNA | Phytochemical (A Type of Cancer) | Effective in Vitro Concentration of Phytochemical/Treatment Time | Effective in Vivo Dose of Phytochemical in Mouse Models of Cancers (A Route of Administration) | Ref. |
|---|---|---|---|---|
| miR-17–92 cluster | Ginsenoside Rd (breast cancer) | ≥50 μM/72 h | 50 mg/kg (intraperitoneal) | [75] |
| Maytenin and 22-β-hydroxymaytenin (head and neck squamous cell carcinomas) | 1.5–1.6 μM/24 h * (maytenin), 1.9–2.5 μM/24 h * (22-β-hydroxymaytenin) | 2 mg/kg (maytenin, intraperitoneal) | [78] | |
| Nitidine chloride (chronic myeloid leukemia) | ≥4 μM/72 h | - | [91] | |
| Oridonin (myelogenous leukemia) | ≥5 μM/24 h | 10–15 mg/kg (intraperitoneal) | [80] | |
| Procyanidin (lung cancer) | ≥10 μM/24 h | 56–112 mg/kg (oral) | [56] | |
| Resveratrol (breast cancer) | ≥6.25 μM/48 h | 25–100 mg/kg (intraperitoneal) | [92] | |
| Swainsonine (glioblastoma) | ≥20 μM/12 h | - | [28] | |
| Triptolide (hepatocellular carcinoma) | ≥50 nM/48 h | 0.2 mg/kg (intraperitoneal) | [83] | |
| miR-21 | Celastrol (colorectal cancer) | 3.2 μM/72 h * | - | [72] |
| Curcumin (osteosarcoma) | ≥2.5 μM/72 h | - | [39] | |
| Formononetin (bladder cancer) | ≥50 μM/24 h | - | [42] | |
| Galangin (cholangiocarcinoma) | ≥50 μM/24 h | - | [43] | |
| Gambogic acid (colorectal cancer) | ≥1 μM/48 h | - | [44] | |
| Honokiol (osteosarcoma) | ≥1 μM/24 h | - | [48] | |
| Puerarin (hepatocellular carcinoma) | ≥50 μM/48 h | 40 mg/kg (intravenous) | [57] | |
| Silibinin (breast cancer) | 200 μM/48 h * | - | [61] | |
| Sophocarpine (head and neck cancer) | 1–1.5 μM/48 h * | 5 mg/kg (intravenous) | [27] | |
| Sulforaphane (colorectal cancer) | ≥5 μM/72 h | - | [30] | |
| Sulforaphane (glioblastoma) | ≥5 μM/24 h | - | [93] | |
| miR-23 | Baicalin (colorectal cancer) | 165.5 μM/24 h * | 50–100 mg/kg (intraperitoneal) | [36] |
| Sinomenine (prostate cancer) | ≥0.25 mM/24 h | - | [26] | |
| Sulforaphane (breast cancer) | ≥2.5 μM/24 h | - | [94] | |
| miR-32 | Isoliquiritigenin (nasopharyngeal cancer) | ≥12.5 μM/48 h | 25–100 mg/kg (oral) | [50] |
| miR-95 | Brucein D (hepatocellular carcinoma) | ≥0.5 μM/72 h | 1.5 mg/kg (intraperitoneal) | [70] |
| miR-99a–125b cluster | Berberine (multiple myeloma) | 75 μM/48 h ** | - | [17] |
| miR-103-3p | Physcion 8-O-β-glucopyranoside (gastric cancer) | ≥10 μM/96 h | 30–50 mg/kg (intraperitoneal) | [55] |
| miR-106b–25 cluster | Epigallocatechin gallate (breast cancer) | ≥10 μM/24 h | 100 mg/kg (oral) | [95] |
| Matrine (gastric cancer) | ≥10 μM/24 h | - | [96] | |
| Triptolide (hepatocellular carcinoma) | ≥50 nM/48 h | 0.2 mg/kg (intraperitoneal) | [83] | |
| miR-155 | Silibinin (breast cancer) | 200 μM/48 h * | - | [61] |
| miR-183-96-182 cluster | Curcumin (breast cancer) | 20 μM/24 h * | - | [97] |
| Matrine (papillary thyroid cancer) | ≥1 mM/48 h | - | [21] | |
| Sanguinarine (gastric cancer) | 150–200 μM/48 h * | 5–10 mg/kg (intraperitoneal) | [25] | |
| miR-196b | Resveratrol (acute lymphoblastic leukemia) | ≥25 μM/72 h | - | [98] |
| miR-217 | Baicalin (colorectal cancer) | ≥10 μM/24 h | - | [99] |
| miR-221 | Resveratrol (melanoma) | 50 μM/24 h ** | 30 mg/kg (intraperitoneal) | [59] |
| α-pinene (hepatocellular carcinoma) | ≥16 μM/24 h | - | [86] | |
| miR-222 | Resveratrol (thyroid cancer) | ≥0.5 μM/72 h | - | [100] |
| miR-223 | Genistein (pancreatic cancer) | 50–60 μM/72 h * | - | [101] |
| miR-301-3p | Isoliquiritigenin (melanoma) | ≥10 μM/24 h | 20 mg/kg (intraperitoneal) | [102] |
| Luteolin (pancreatic cancer) | 23.3 μM/96 h * | - | [103] | |
| miR-324-5p | Sinomenine (breast cancer) | ≥0.25 mM/24 h | - | [104] |
| miR-374a | Isoliquiritigenin (breast cancer) | ≥6.25 μM/72 h | - | [105] |
| Neferine (breast cancer) | ≥4 μM/24 h | - | [22] | |
| miR-382 | Sulforaphane (breast cancer) | ≥2.5 μM/24 h | - | [94] |
| miR-429 | Berberine (colorectal cancer) | ≥4 μM/72 h | - | [90] |
| Evodiamine (colorectal cancer) | ≥3 μM/72 h | - | ||
| miR-625-3p | Icariin (thyroid cancer) | ≥20 μM/24 h | - | [49] |
| miR-1290 | Resveratrol (acute lymphoblastic leukemia) | ≥25 μM/72 h | - | [98] |
| miR-4295 | Ginsenoside Rh2 (prostate cancer) | ≥16.6 μM/96 h | - | [76] |
| miR-7641 | Curcumin (bladder cancer) | ≥10 μM/48 h | - | [106] |
* the half maximal inhibitory concentration (IC50) at an indicated treatment time. ** tests only at an indicated concentration.
Table 4.
Tumor-suppressive miRNAs modulated by phytochemicals.
| miRNA | Phytochemical (A Type of Cancer) | Effective in Vitro Concentration of Phytochemical/Treatment Time | Effective in Vivo Dose of Phytochemical in Mouse Models of Cancers (A Route of Administration) | Ref. |
|---|---|---|---|---|
| Let-7c | Quercetin (pancreatic cancer) | 50 μM/48 h *** | - | [58] |
| Let-7f | Lycopene (prostate cancer) | ≥10 μM/48 h | - | [77] |
| miR-9 | Sulforaphane (lung cancer) | ≥2.5 μM/120 h | - | [129] |
| miR-10a | β-sitosterol-d-glucoside (breast cancer) | 30.82–49.76 μM/24 h ** | 60 mg–120 mg/kg (intragastric) | [31] |
| miR-15a | Gossypol (pituitary cancer) | ≥5 μM/96 h | - | [130] |
| miR-16 | Sanguinarine (hepatocellular carcinoma) | ≥0.5 μM/48 h | 10 mg/kg (intraperitoneal) | [131] |
| miR-22 | Matrine (colorectal cancer) | ≥0.25 mM/72 h | - | [43] |
| miR-27a-3p * | Maytenin (head and neck cancer) | 1.5–1.6 μM/24 h ** | 2 mg/kg (intraperitoneal) | [78] |
| miR-29-3p * | Sanguinarine (gastric cancer) | 150–200 μM/48 h ** | 5–10 mg/kg (intraperitoneal) | [25] |
| miR-34 | 5-Aminolevulinic acid (melanoma) | 1 mM/4 h + ultrasound | 200 mg/kg (intraperitoneal) + ultrasound | [132] |
| Artemisinin and Artesunate (breast cancer) | 300 μM/48 h *** (artemisinin), ≥5 μM/48 h (artesunate) | - | [20] | |
| Cucurbitacin D (cervical cancer) | ≥0.05 μM/72 h | 1 mg/kg (intra-tumoral) | [73] | |
| Curcumin (breast cancer) | ≥20 μM/48 h | - | [133] | |
| Genistein (head and neck cancer) | ≥20 μM/24 h | 25 mg–50 mg/kg (oral) | [45] | |
| Indole-3-carbinol (breast cancer) | ≥200 μM/48 h | - | [20] | |
| Luteolin (lung cancer) | 40 μM/72 h ** | 50 mg–200 mg/kg (intragastric) | [54] | |
| Resveratrol (breast cancer) | ≥12.5 μM/72 h | - | [134] | |
| miR-101 | Apigenin (hepatocellular carcinoma) | 10 μM/48 h *** | - | [33] |
| miR-122 | Coptisine (hepatocellular carcinoma) | ≥3.1 μM/24 h | 150 mg/kg (oral) | [18] |
| Oleanolic acid (lung cancer) | ≥65 μM/48 h | 120 mg/kg (oral) | [79] | |
| Resveratrol (breast cancer) | ≥100 μM/24 h | - | [135] | |
| miR-124 | Physcion 8-O-β-glucopyranoside (melanoma) | ≥10 μM/24 h | - | [136] |
| miR-125b | Astragalin (hepatocellular carcinoma) | ≥11 μM/48 h | 10–20 mg/kg (oral) | [34] |
| miR-126 | Tubeimoside-1 (lung cancer) | ≥10 μM/48 h | - | [84] |
| miR-132 | Chrysin and Curcumin (breast cancer) | ≥20 μM/48 h (chrysin), ≥10 μM/48 h (curcumin) | - | [38] |
| miR-133a | Artesunate (rhabdomyosarcoma) | ≥5 μM/24 h | 25 mg/kg (intraperitoneal) | [137] |
| Brazilein (vestibular schwannoma) | ≥10 μM/96 h | - | [138] | |
| miR-134 | Astragaloside IV (colorectal cancer) | ≥6.4 μM/48 h | - | [139] |
| miR-137 | Isorhapontigenin (bladder cancer) | 10 μM/24 h *** | 150 mg/kg (intraperitoneal) | [51] |
| Isorhapontigenin (urothelial cancer) | ≥5 μM/24 h | - | [140] | |
| miR-139-5p | Resveratrol (osteosarcoma) | ≥5 μM/48 h | - | [141] |
| miR-143 | 5-Aminolevulinic acid (cervical cancer) | ≥0.25 μM + photodynamic therapy/24 h | - | [142] |
| Cucurbitacin D (cervical cancer) | ≥0.05 μM/72 h | 1 mg/kg (intra-tumoral) | [73] | |
| miR-144-3p | Licochalcone A (lung cancer) | ≥10 μM/48 h | - | [143] |
| miR-145 | Cucurbitacin D (cervical cancer) | ≥0.05 μM/72 h | 1 mg/kg (intra-tumoral) | [73] |
| Isorhapontigenin (glioblastoma) | ≥10 μM/24 h | - | [144] | |
| miR-148a | Ailanthone (breast cancer) | ≥5 μM/48 h | - | [145] |
| miR-149-5p | Ursolic acid (breast cancer) | 20 μM/48 h *** | 10 mg/kg (intraperitoneal) | [85] |
| Ursolic acid (lung cancer) | ≥5 μM/48 h | Treated cells with ursolic acid at 20 μM for 72 h before implanting in mice | [146] | |
| miR-181-3p | Curcumol (breast cancer) | ≥254 μM/48 h | 20 mg/kg of curcumol + 2.5 mg/kg of doxorubicin (intraperitoneal) | [74] |
| miR-193-3p | Triptolide (nephroblastoma) | ≥10 nM/72 h | - | [147] |
| miR-200 | Quercetin (pancreatic cancer) | 50 μM/72 h *** | - | [148] |
| Resveratrol (colorectal cancer) | ≥25 μM/48 h | - | [149] | |
| Toosendanin (gastric cancer) | ≥0.5 μM/48 h | 0.2 mg/kg (intraperitoneal) | [82] | |
| miR-203 | Silymarin (lung cancer) | ≥10 μM/24 h | - | [64] |
| miR-204-3p | Delphinidin (colorectal cancer) | ≥25 μM/24 h | Treated cells with delphinidin at 100 μM for 24 h before implanting in mice | [40] |
| miR-206 | Artesunate (rhabdomyosarcoma) | ≥5 μM/24 h | 25 mg/kg (intraperitoneal) | [137] |
| miR-218 | Andrographolide (oral cancer) | ≥6.25 μM/24 h | 10 mg/kg (oral) | [67] |
| miR-296-3p | Epigallocatechin gallate (nasopharyngeal cancer) | ≥20 μM/24 h | - | [41] |
| miR-320a | Hydroxygenkwanin (hepatocellular carcinoma) | ≥10 μM/72 h | 1 mg/kg (intraperitoneal) | [47] |
| miR-340 | Kaempferol (lung cancer) | ≥20 μM | - | [52] |
| miR-345 | Trans-3,5,4′-trimethoxystilbene (lung cancer) | ≥0.5 μM/72 h | 30 mg/kg (oral) | [65] |
| miR-383-5p | Allicin (gastric cancer) | 62 μM/48 h *** | - | [29] |
| miR-384 | Luteolin (colorectal cancer) | ≥10 μM/48 h | 100 mg/kg (intragastric) | [150] |
| miR-424 | Resveratrol (breast cancer) | ≥12.5 μM/72 h | - | [134] |
| miR-449a | Ailanthone (acute myeloid leukemia) | ≥0.5 μM/48 h | - | [66] |
| miR-485 | Epigallocatechin gallate (lung cancer) | ≥10 μM/24 h | 20 mg/kg (intraperitoneal) | [151] |
| miR-498 | Trans-3,5,4′-trimethoxystilbene (lung cancer) | ≥0.5 μM/72 h | 30 mg/kg (oral) | [65] |
| miR-503 | Resveratrol (breast cancer) | ≥12.5 μM/72 h | - | [134] |
| miR-520b | Apigenin (hepatocellular carcinoma) | ≥10 μM/48 h | 50 mg/kg (intraperitoneal) | [152] |
| miR-542-5p | Pristimerin (breast cancer) | ≥1 μM/24 h | - | [81] |
| miR-663 | Resveratrol (breast cancer) | 100 μM/24 h *** | - | [153] |
| miR-744 | Resveratrol (breast cancer) | 100 μM/24 h *** | - | [153] |
| miR-1972 | Cannabidiol (neuroblastoma) | ≥5 μM/24 h | - | [71] |
| miR-3127-5p | Baicalein (hepatocellular carcinoma) | ≥20 μM/24 h | - | [35] |
| miR-6809-5p | Luteolin (hepatocellular carcinoma) | ≥10 μM/120 h | 50 mg/kg (intraperitoneal) | [154] |
* indicates tumor-suppressive miRNAs that are downregulated by phytochemicals. ** the half maximal inhibitory concentration (IC50) at an indicated treatment time. *** tests only at an indicated concentration.
Table 5.
MiRNAs controlling anti-cancer activity of phytochemicals.
| miRNA | Phytochemical (A Type of Cancer) | In Vitro Finding | In Vivo Experiment Condition | Ref. |
|---|---|---|---|---|
| miR-7-3p | Luteolin and Silibinin (glioblastoma) | Treatment with luteolin (20 μM/24 h) or silibinin (50 μM/24 h) induces apoptosis and miR-7-3p expression | 10 mg/kg (luteolin, oral) or 200 mg/kg (silibinin, oral) with and without miR-7-3p (50 μg, intravenous) | [279] |
| miR-25 | Physcion 8-O-β-glucopyranoside (ovarian cancer) | Overexpression of miR-25 reduces the cytotoxicity of physcion 8-O-β-glucopyranoside (10 μM) | - | [280] |
| miR-17–92 cluster | Oridonin (myelogenous leukemia) | Inhibition of miR-17 or miR-20a enhances the cytotoxicity of oridonin (2.5 and 5 μM for 72 h) | 10–15 mg/kg (oridonin, intraperitoneal) | [80] |
| miR-126 | Epigallocatechin gallate (osteosarcoma) | Overexpression of miR-126 potentiates the cytotoxicity of epigallocatechin gallate (100 μM for 48 h) | - | [281] |
| miR-137 | Delphinidin (glioblastoma) | Overexpression of miR-137 enhances apoptosis induced by delphinidin (50 μM for 24 h) | - | [282] |
| miR-138 | Apigenin (neuroblastoma) | Ectopic expression of miR-138 enhances apoptosis induced by apigenin (100 μM for 24 h) | 10 μg/mouse (apigenin, intra-tumoral) + hTERT shRNA plasmid or miR-138 mimic | [283] |
| miR-143 | Shikonin (glioblastoma) | Overexpression of miR-143 enhances apoptosis induced by shikonin (2 μM for 24 h) | 2 mg/kg (shikonin, intraperitoneal) in mice bearing miR-143-overexpressing cells | [60] |
| miR-210 | 1′S-1′-acetoxychavicol acetate (cervical cancer) | Knockdown of miR-210 potentiates the cytotoxicity of 1′S-1′-acetoxychavicol acetate (5–20 μM for 48 h) | - | [277] |
| miR-629 | 1′S-1′-acetoxychavicol acetate (cervical cancer) | Knockdown of miR-629 potentiates the cytotoxicity of 1′S-1′-acetoxychavicol acetate (5–20 μM for 48 h) | - | [278] |
Table 6.
MiRNAs involving in the regulation of efficacy of plant-derived anti-cancer agents.
| miRNAs | Phytochemicals (A Type of Cancer) | In Vitro Finding | In Vivo Experiment Condition | Ref. |
|---|---|---|---|---|
| Cancer resistance-promoting miRNAs | ||||
| miR-21 | Etoposide (colorectal cancer) | 3.97 μM/72 h * (miR-control-overexpressing cells), 12.7 μM/72 h * (miR-21-overexpressing cells) | - | [291] |
| Paclitaxel (breast cancer) | ≈ 6 μM/48 h * (anti-miR-control-treated cells), ≈ 3 μM/48 h * (anti-miR-21-treated cells) | 1 mg/kg (paclitaxel, intravenous) + anti-miR-21 (intra-tumoral) | [296] | |
| miR-27a-3p | Paclitaxel (ovarian cancer) | Silencing of miR-27a-3p increases the cytotoxicity of paclitaxel (4, 8, and 12 μM for 7 days) | 5 mg/kg (paclitaxel) in mice bearing miR-27a-3p-overexpressing cells | [297] |
| Paclitaxel (ovarian cancer) | Silencing of miR-27a-3p increases the cytotoxicity of paclitaxel (0.04–23 μM for 48 h) | - | [298] | |
| miR-125b | Paclitaxel (breast cancer) | Overexpression of miR-125b reduces the cytotoxicity of paclitaxel (4, 8, and 16 nM for 48 h) | - | [299] |
| miR-140-3p | Paclitaxel (chordoma) | Silencing of miR-140-3p or miR-155-5p increases the cytotoxicity of paclitaxel (10 μM for 24–72 h) | - | [300] |
| miR-155-5p | ||||
| miR-192 | Etoposide (lung cancer) | Knockdown of miR-192 sensitizes cells to etoposide (≤100 μM for 24 h) | - | [301] |
| miR-374a | Etoposide (glioblastoma) | Knockdown of miR-374a sensitizes cells to etoposide (0.5–8 μM for 48 h) | - | [292] |
| miR-514b-5p | Irinotecan (colorectal cancer) | Overexpression of miR-514b-5p reduces the cytotoxicity of irinotecan (25, 50, and 100 μM for 48 h) | 40 mg/kg (irinotecan) in mice bearing miR-514b-5p-overexpressing cells | [302] |
| miR-520h | Paclitaxel (breast cancer) | Overexpression of miR-520h reduces the cytotoxicity of paclitaxel (1, 5, and 10 nM for 24 h) | - | [303] |
| miR-662 | Etoposide (lung cancer) | Knockdown of miR-662 sensitizes cells to etoposide (≤100 μM for 24 h) | - | [301] |
| miR-1207-5p | Paclitaxel (breast cancer) | Knockdown of miR-1207-5p sensitizes cells to paclitaxel (10 nM for 5 days) | - | [304] |
| miR-4262 | Paclitaxel (lung cancer) | Overexpression of miR-4262 reduces the cytotoxicity of paclitaxel (4 and 8 μM) | Paclitaxel + anti-miR-4262 (intravenous) | [305] |
| Cancer resistance-suppressing miRNAs | ||||
| miR-1 | Vincristine (gastric cancer) | Knockdown of miR-1 reduces the cytotoxicity of vincristine (10 and 20 μM) | - | [306] |
| miR-7-5p | Paclitaxel (breast cancer) | 25–35 μM/72 h * (miR-control-overexpressing cells), 5–15 μM/72 h * (miR-7-5p-overexpressing cells) | - | [307] |
| miR-29-3p | Etoposide (cervical cancer) | Overexpression of miR-29-3p enhances the cytotoxicity of etoposide (60 μM for 48 h) | - | [293] |
| Paclitaxel (nasopharyngeal cancer) | 6.4–7.5 nM/96 h * (miR-control-overexpressing cells), 0.7–0.8 nM/96 h * (miR-29-3p-overexpressing cells) | 5 mg/kg (paclitaxel, intraperitoneal) in mice bearing miR-29-3p inhibitor treated cells | [308] | |
| miR-34 | Etoposide (retinoblastoma) | Overexpression of miR-34 enhances the cytotoxicity of etoposide (35 μM for 48 h) | - | [309] |
| Vincristine (retinoblastoma) | Overexpression of miR-34 enhances the cytotoxicity of vincristine (130 nM for 48 h) | - | [309] | |
| miR-126 | Vincristine (gastric cancer) | Overexpression of miR-126 enhances the cytotoxicity of vincristine (1.2, 6, and 12 μM for 48 h) | - | [310] |
| miR-133b | Vincristine (colorectal cancer) | 420 μM/24 h * (miR-control-overexpressing cells), 120 μM/24 h * (miR-133b-overexpressing cells) | - | [311] |
| miR-145-5p | Paclitaxel (breast cancer) | ≈ 6 μM/24 h * (miR-control-overexpressing cells), ≤3 μM/24 h * (miR-145-5p-overexpressing cells) | miR-145-5p mimic (intra-tumoral) in mice bearing paclitaxel-resistant cells | [312] |
| miR-193-3p | Etoposide (osteosarcoma) | Overexpression of miR-193-3p enhances the cytotoxicity of etoposide at IC50/72 h | - | [294] |
| miR-196-5p | Etoposide (hepatocellular carcinoma) | Overexpression of miR-196-5p increases apoptosis induced by etoposide (50 μM for 16 h) | - | [295] |
| miR-200 | Irinotecan (colorectal cancer) | Delivery of miR-200 using nanoparticles increases apoptosis after treatment with irinotecan (incorporated in liposomes) for 48 h | 100 mg/kg (irinotecan, intravenous) + 1.25 mg/kg (miR-200, intravenous) | [313] |
| Vincristine (gastric cancer) | Overexpression of miR-200 increases apoptosis induced by vincristine (4 μM for 48 h) | - | [314] | |
| miR-302 | Etoposide (leukemia) | Overexpression of miR-302 enhances the cytotoxicity of etoposide (20, 50, 100, and 200 μM) | 20 mg/kg (etoposide, intraperitoneal) in mice bearing miR-302-overexpressing cells | [315] |
| miR-365 | Paclitaxel (endometrial cancer) | Overexpression of miR-365 enhances the cytotoxicity of paclitaxel (200 and 300 nM for 24 h) | - | [316] |
| miR-383-5p | Paclitaxel (ovarian cancer) | Overexpression of miR-383-5p enhances the cytotoxicity of paclitaxel (≤5 μM) | Subcutaneous injection of miR-383-5p-overexpressing cells | [317] |
| miR-429 | Vincristine (gastric cancer) | Overexpression of miR-429 increases apoptosis induced by vincristine (4 μM for 48 h) | - | [314] |
| miR-495-3p | Vincristine (gastric cancer) | Overexpression of miR-495-3p enhances the cytotoxicity of vincristine (≤1.65 μM for 24 h) | Subcutaneous injection of miR-495-3p-overexpressing cells | [318] |
| miR-542-3p | Paclitaxel (breast cancer) | Overexpression of miR-542-3p increases apoptosis induced by paclitaxel (3 nM for 24 h) | 6 mg/kg (paclitaxel, intraperitoneal) + 5μg (miR-542-3p, intra-tumoral) | [319] |
| miR-584-5p | Vincristine (medulloblastoma) | Overexpression of miR-584-5p enhances the cytotoxicity of vincristine (≤1 μM for 72 h) | 0.5 mg/kg (vincristine, intraperitoneal) in mice bearing miR-584-5p-overexpressing cells | [320] |
| miR-621 | Paclitaxel (breast cancer) | Overexpression of miR-621 promotes apoptosis induced by paclitaxel (20 μM for 24 h) | 15 mg/kg (paclitaxel, intraperitoneal) in mice bearing miR-621-overexpressing cells | [321] |
| miR-627 | Irinotecan (colorectal cancer) | Overexpression of miR-627 enhances the cytotoxicity of irinotecan (5, 10, and 20 μM for 48 h) | 50 mg/kg (irinotecan, intraperitoneal) + 0.4 μg (calcitriol, intraperitoneal) | [15] |
| miR-874 | Vincristine (gastric cancer) | ≈ 4 μM/48 h * (anti-miR-control treated cells), ≈ 2 μM /48 h * (anti-miR-874 treated cells) | - | [322] |
| miR-3163 | Etoposide (retinoblastoma) | 0.57 μM/72 h * (miR-control-overexpressing cells), 0.39 μM/72 h * (miR-3163-overexpressing cells) | - | [323] |
| Vincristine (retinoblastoma) | 1.27 μM/72 h * (miR-control-overexpressing cells), 0.77 μM/72 h * (miR-3163-overexpressing cells) | - | [323] | |
| miR-4454 | Irinotecan (colorectal cancer) | Knockdown of miR-4454 reduces apoptosis induced by irinotecan (10 μM for 48 h) | Subcutaneous injection of miR-4454 expressing cells | [324] |
| miR-5195-3p | Paclitaxel (breast cancer) | Overexpression of miR-5195-3p enhances the cytotoxicity of paclitaxel (2.5–10 μM) | - | [325] |
* the half maximal inhibitory concentration (IC50) at indicated treatment time.
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
MDPI and ACS Style
Son, S.W.; Lee, H.Y.; Moeng, S.; Kuh, H.J.; Choi, S.Y.; Park, J.K. Participation of MicroRNAs in the Treatment of Cancer with Phytochemicals. Molecules 2020, 25, 4701. https://doi.org/10.3390/molecules25204701
AMA Style
Son SW, Lee HY, Moeng S, Kuh HJ, Choi SY, Park JK. Participation of MicroRNAs in the Treatment of Cancer with Phytochemicals. Molecules. 2020; 25(20):4701. https://doi.org/10.3390/molecules25204701
Chicago/Turabian StyleSon, Seung Wan, Han Yeoung Lee, Sokviseth Moeng, Hyo Jeong Kuh, Soo Young Choi, and Jong Kook Park. 2020. "Participation of MicroRNAs in the Treatment of Cancer with Phytochemicals" Molecules 25, no. 20: 4701. https://doi.org/10.3390/molecules25204701