
In Vitro and In Vivo Screening of Wild Bitter Melon Leaf for Anti-Inflammatory Activity against Cutibacterium acnes

 ,
, 
Abstract
:1. Introduction
2. Results and Discussion
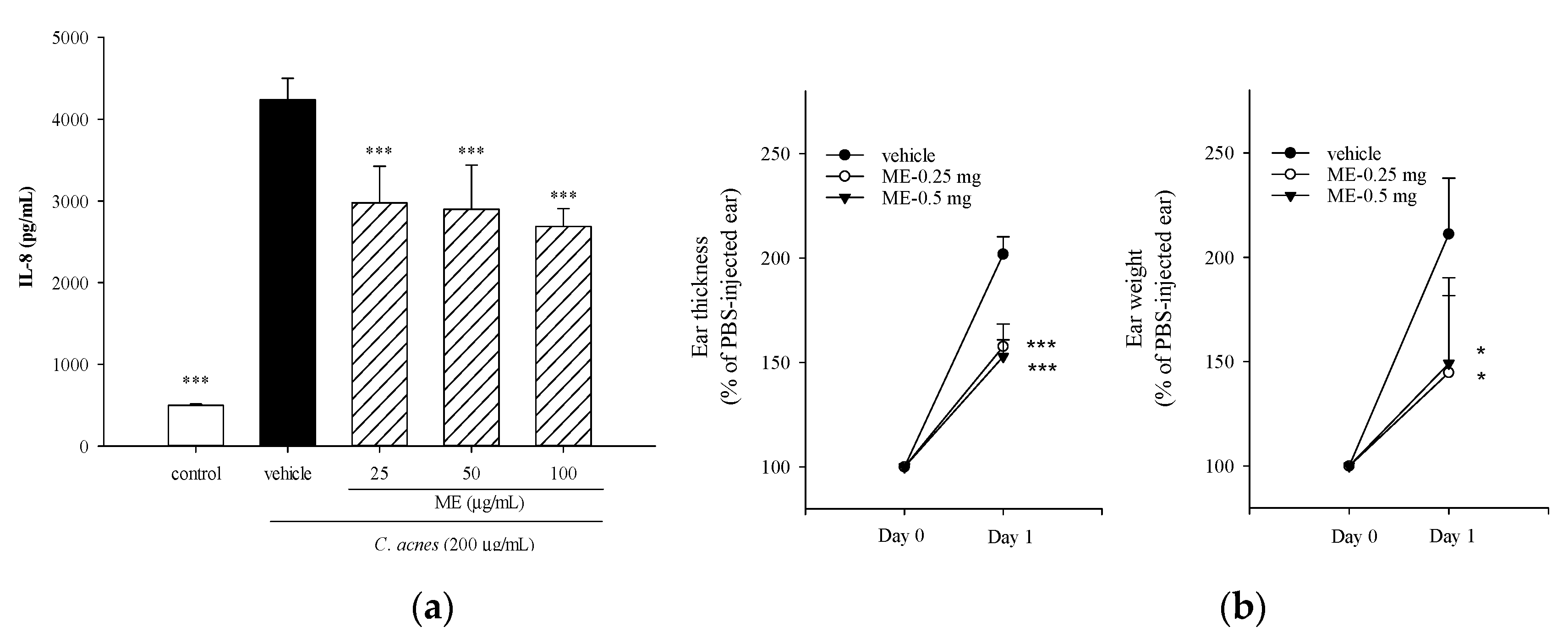
2.1. Effects of ME of WBM Leaf on C. acnes-Induced Cellular IL-8 Production and Mouse Ear Edema
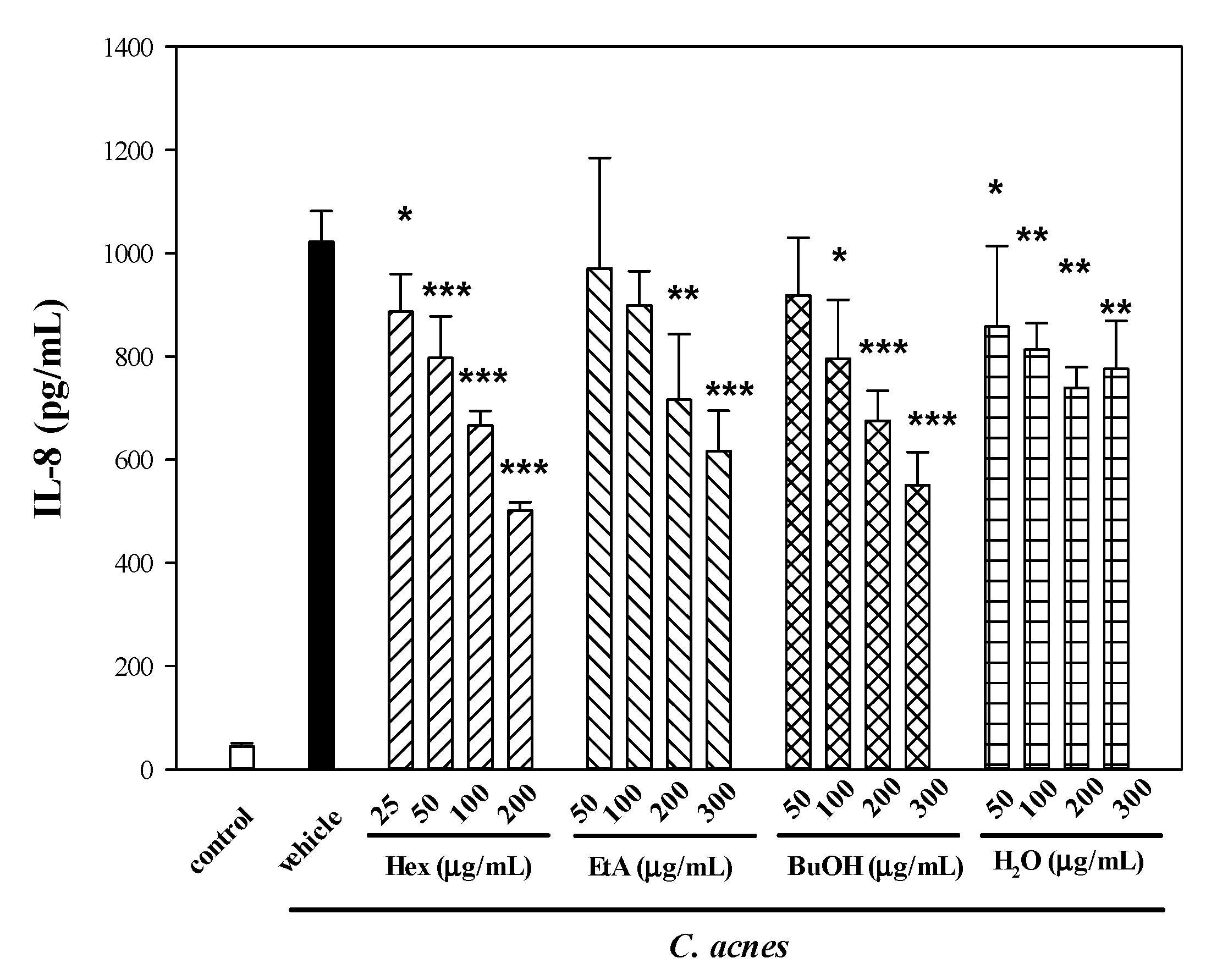
2.2. Effects of Four Partitioned Fractions from ME of WBM Leaf on C. acnes-Induced Cellular IL-8 Production
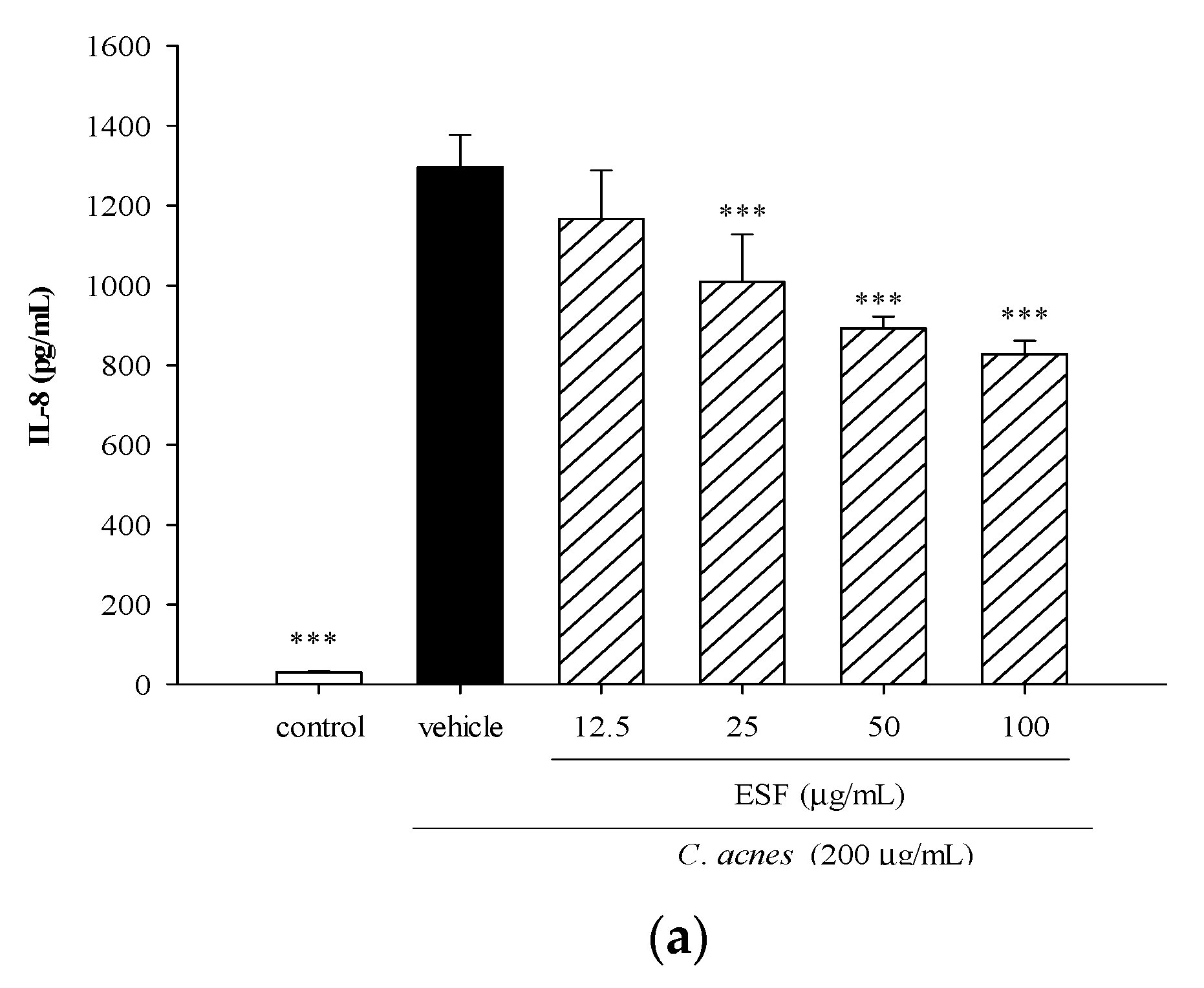
2.3. Effects of Ethanol-Soluble Fraction (ESF) on C. acnes-Induced Cellular IL-8 Production and Mouse Ear Edema
2.4. GC-MS Analysis of ESF
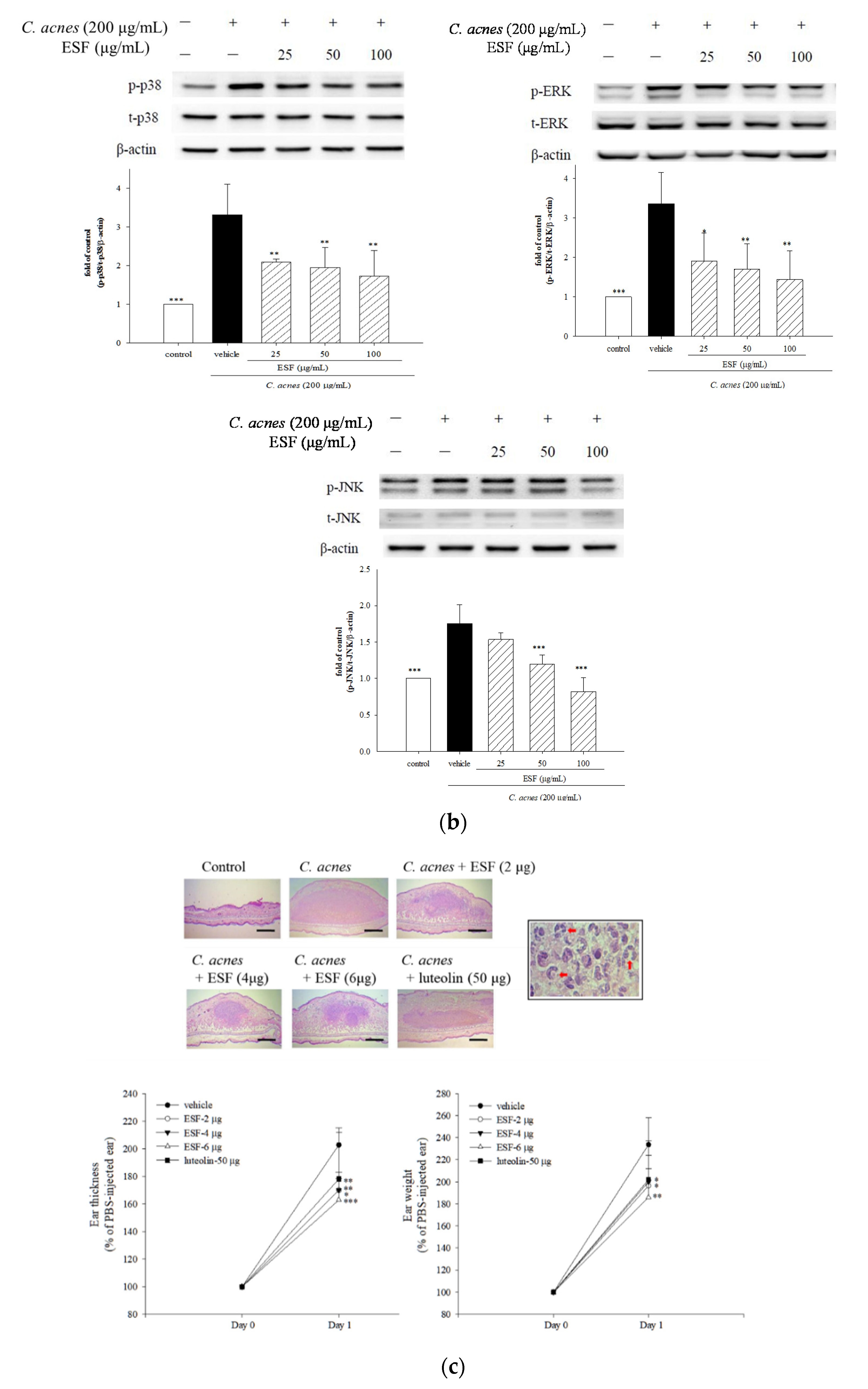
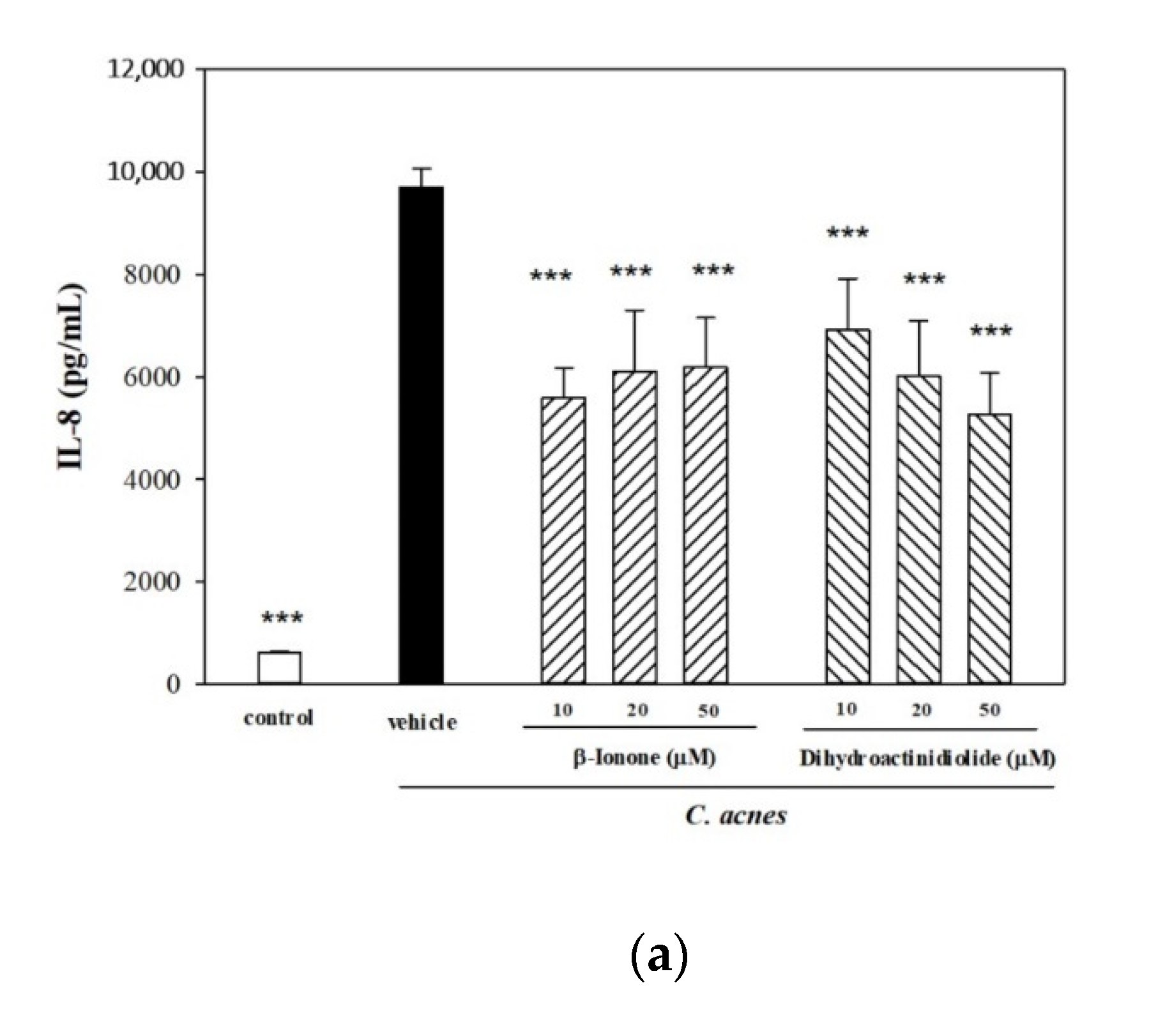
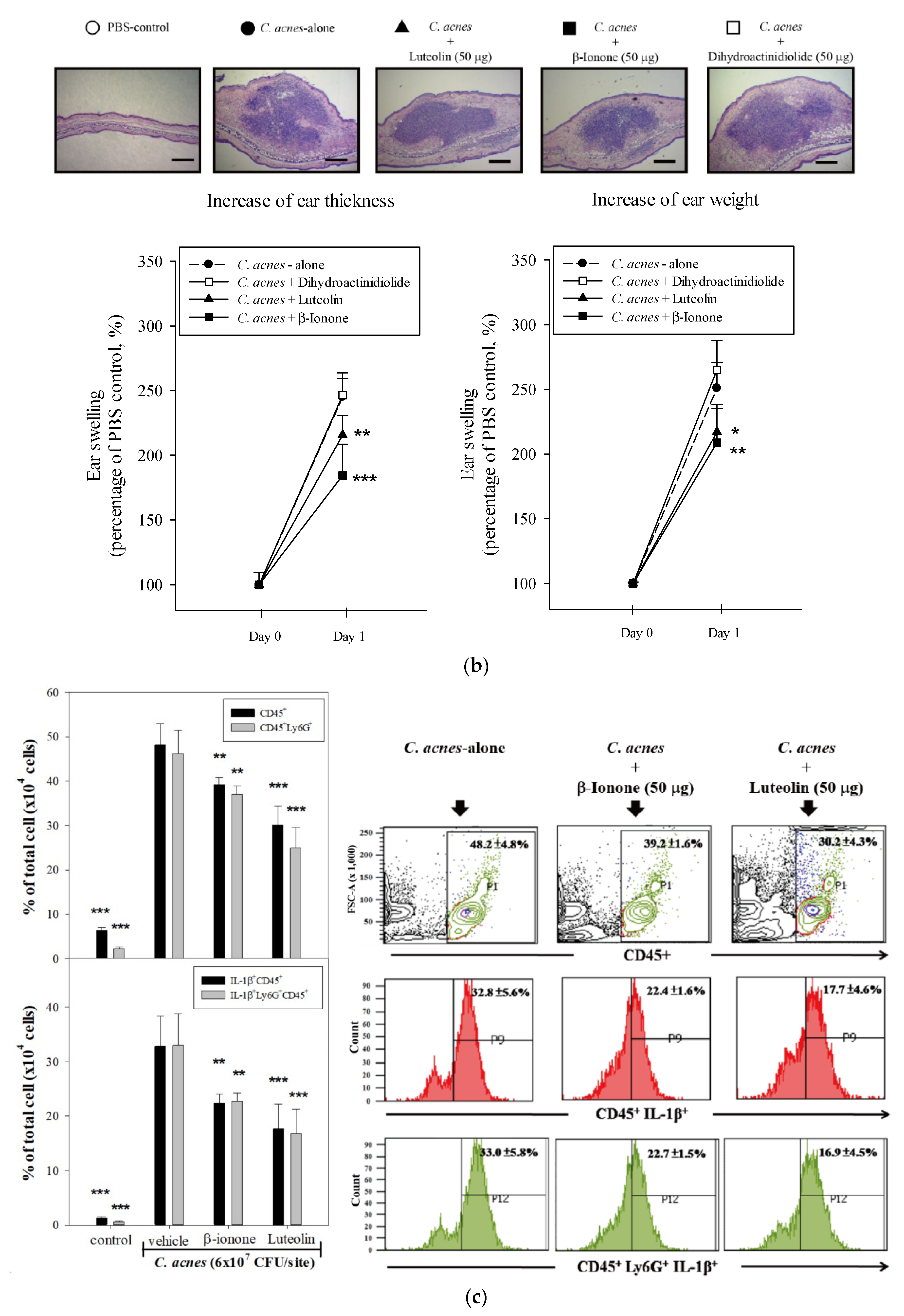
2.5. Effects of β-Ionone and Dihydroactinidiolide on C. acnes-Induced Cellular IL-8 Production and Mouse Ear Edema
2.6. Effects of β-Ionone on Cellular IL-1β Production and Caspase-1 Expression
3. Materials and Methods
3.1. Materials
3.2. Isolation and Determination of Active Compounds from WBM Leaf Extract
3.3. Analysis of ESF by Gas Chromatography–Mass Spectrometry
3.4. Bacterial and Cell Cultures and Growth Conditions
3.5. Measurement of IL-8 Production in Human Monocytic THP-1 Cells
3.6. Protein Lysate Preparation and Western Blot Analysis
3.7. C. acnes-Induced Inflammation in Mouse Ears
3.8. Flow Cytometric Analysis of Single-Cell Suspensions Prepared from Skin
3.9. Statistical Analysis
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Tanghetti, E.A. The role of inflammation in the pathology of acne. J. Clin. Aesthet. Dermatol. 2013, 6, 7–35. [Google Scholar]
- Omer, H.; McDowell, A.; Alexeyev, O.A. Understanding the role of Propionibacterium acnes in acne vulgaris: The critical importance of skin sampling methodologies. Clin. Dermatol. 2017, 35, 118–129. [Google Scholar] [CrossRef] [PubMed]
- Kumar, B.; Pathak, R.; Mary, P.B.; Jha, D.; Sardana, K.; Gautam, H.K. New insights into acne pathogenesis: Exploring the role of acne-associated microbial populations. Dermatol. Sin. 2016, 34, 67–73. [Google Scholar] [CrossRef] [Green Version]
- Vowels, B.R.; Yang, S.; Leyden, J.J. Induction of proinflammatory cytokines by a soluble factor of Propionibacterium acnes: Implications for chronic inflammatory acne. Infect. Immun. 1995, 63, 3158–3165. [Google Scholar] [CrossRef] [Green Version]
- Demina, O.M.; Kartelishev, A.V.; Karpova, E.I.; Olga, I.; Danischuk, O.I. Role of cytokines in the pathogenesis of acne. Int. J. Biomed. 2017, 7, 37–40. [Google Scholar] [CrossRef]
- Dinarello, C.A. Anti-inflammatory agents: Present and future. Cell 2010, 140, 935–950. [Google Scholar] [CrossRef] [Green Version]
- Alam, M.A.; Uddin, R.; Subhan, N.; Rahman, M.M.; Jain, P.; Reza, H.M. Beneficial role of bitter melon supplementation in obesity and related complications in metabolic syndrome. J. Lipids 2015, 2015, 496169. [Google Scholar] [CrossRef]
- Fang, E.F.; Ng, T.B. Bitter gourd (Momordica charantia) is a cornucopia of health: A review of its credited antidiabetic, anti-HIV, and antitumor properties. Curr. Mol. Med. 2011, 11, 417–436. [Google Scholar] [CrossRef]
- Ciou, S.Y.; Hsu, C.C.; Kuo, Y.H.; Chao, C.Y. Effect of wild bitter gourd treatment on inflammatory responses in BALB/c mice with sepsis. Biomedicine (Taipei) 2014, 4, 17. [Google Scholar] [CrossRef]
- Mahwish; Saeed, F.; Arshad, M.S.; Nisa, M.U.; Nadeem, M.T.; Arshad, M.U. Hypoglycemic and hypolipidemic effects of different parts and formulations of bitter gourd (Momordica Charantia). Lipids Health Dis. 2017, 16, 211. [Google Scholar] [CrossRef] [Green Version]
- Kubola, J.; Siriamornpun, S. Phenolic contents and antioxidant activities of bitter gourd (Momordica charantia L.) leaf, stem and fruit fraction extracts in vitro. Food Chem. 2008, 110, 881–890. [Google Scholar] [CrossRef] [PubMed]
- Hsu, C.; Tsai, T.H.; Li, Y.Y.; Wu, W.H.; Huang, C.J.; Tsai, P.J. Wild bitter melon (Momordica charantia Linn. var. abbreviata Ser.) extract and its bioactive components suppress Propionibacterium acnes-induced inflammation. Food Chem. 2012, 135, 976–984. [Google Scholar] [PubMed]
- Tsai, T.H.; Huang, C.J.; Wu, W.H.; Huang, W.C.; Chyuan, J.H.; Tsai, P.J. Antioxidant, cell-protective, and anti-melanogenic activities of leaf extracts from wild bitter melon (Momordica charantia Linn. var. abbreviata Ser.) cultivars. Bot. Stud. 2014, 55, 78–88. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Huang, W.C.; Tsai, T.H.; Huang, C.J.; Li, Y.Y.; Chyuan, J.H.; Chuang, L.T.; Tsai, P.J. Inhibitory effects of wild bitter melon leaf extract on Propionibacterium acnes-induced skin inflammation in mouse and cytokine production in vitro. Food Funct. 2015, 6, 2550–2560. [Google Scholar] [CrossRef] [PubMed]
- Tsai, T.H.; Huang, W.C.; Ying, H.T.; Kuo, Y.H.; Shen, C.C.; Lin, Y.K.; Tsai, P.J. Wild bitter melon leaf extract inhibits Porphyromonas gingivalis-induced inflammation: Identification of active compounds through bioassay-guided isolation. Molecules 2016, 21, 454. [Google Scholar] [CrossRef] [Green Version]
- Lu, Y.L.; Liu, Y.H.; Liang, W.L.; Chyuan, J.H.; Cheng, K.T.; Liang, H.J.; Hou, W.C. Antibacterial and cytotoxic activities of different wild bitter gourd cultivars (Momordica charantia L. var. abbreviata Seringe). Bot. Stud. 2011, 52, 427–434. [Google Scholar]
- Zhu, F.; Du, B.; Xu, B. Anti-inflammatory effects of phytochemicals from fruits, vegetables, and food legumes: A review. Crit. Rev. Food Sci. 2018, 58, 1260–1270. [Google Scholar] [CrossRef]
- Briganti, S.; Picardo, M. Antioxidant activity, lipid peroxidation and skin diseases. What’s new? J. Eur. Acad. Dermatol. Venereol. 2003, 17, 663–669. [Google Scholar] [CrossRef]
- Nand, P.; Drabu, S.; Gupta, K.R. Screening for antioxidant and antibacterial potential of common medicinal plants in the treatment of acne. Int. J. Drug Dev. Res. 2012, 4, 65–71. [Google Scholar]
- Li, W.H.; Zhang, L.; Lyte, P.; Rodriguez, L.; Cavender, D.; Southall, M.D. p38 MAP Kinase inhibition reduces Propionibacterium acnes-induced inflammation in vitro. Dermatol. Ther. (Heidelb) 2015, 5, 53–66. [Google Scholar] [CrossRef] [Green Version]
- Kaminska, B. MAPK signaling pathways as molecular targets for anti-inflammatory therapy--from molecular mechanisms to therapeutic benefits. Biochim. Biophys. Acta 2005, 1754, 253–262. [Google Scholar] [CrossRef] [PubMed]
- Chen, X.; Meng, X.; Foley, N.M.; Shi, X.; Liu, M.; Chai, Y.; Li, Y.; Redmond, H.P.; Wang, J.; Wang, J.H. Activation of the TLR2-mediated downstream signaling pathways NF-κB and MAPK is responsible for B7-H3-augmented inflammatory response during S. pneumoniae infection. J. Neuroimmunol. 2017, 310, 82–90. [Google Scholar] [CrossRef] [PubMed]
- Reddi, K.; Henderson, B.; Meghji, S.; Wilson, M.; Poole, S.; Hopper, C.; Harris, M.; Hodges, S.J. Interleukin 6 production by lipopolysaccharide-stimulated human fibroblasts is potently inhibited by naphthoquinone (vitamin K) compounds. Cytokine 1995, 7, 287–290. [Google Scholar] [CrossRef] [PubMed]
- Ohsaki, Y.; Shirakawa, H.; Miura, A.; Giriwono, P.E.; Sato, S.; Ohashi, A.; Iribe, M.; Goto, T.; Komai, M. Vitamin K suppresses the lipopolysaccharide-induced expression of inflammatory cytokines in cultured macrophage-like cells via the inhibition of the activation of nuclear factor κB through the repression of IKKα/β phosphorylation. J. Nutr. Biochem. 2010, 21, 1120–1126. [Google Scholar] [CrossRef] [PubMed]
- Reitera, E.; Jiang, Q.; Christen, S. Anti-inflammatory properties of α- and γ-tocopherol. Mol. Asp. Med. 2007, 28, 668–691. [Google Scholar] [CrossRef] [Green Version]
- Cárdeno, A.; Aparicio-Soto, M.; Montserrat-d. la Paz, S.; Bermudez, B.; Muriana, F.J.G.; Alarcón-d.-la-Lastra, C. Squalene targets pro- and anti-inflammatory mediators and pathways to modulate over-activation of neutrophils, monocytes and macrophages. J. Funct. Foods 2015, 14, 779–790. [Google Scholar] [CrossRef] [Green Version]
- Gunawardhana, L.; Mobley, S.A.; Sipes, I.G. Modulation of 1,2-dichlorobenzene hepatotoxicity in the Fischer-344 rat by a scavenger of superoxide anions and an inhibitor of Kupffer cells. Toxicol. Appl. Pharmacol. 1993, 119, 205–213. [Google Scholar] [CrossRef]
- Cai, P.; Kaphalia, B.S.; Ansari, G.A.S. Methyl palmitate: Inhibitor of phagocytosis in primary rat Kupffer cells. Toxicology 2005, 210, 197–204. [Google Scholar] [CrossRef]
- Huh, S.; Kim, Y.S.; Jung, E.; Lim, J.; Jung, K.S.; Kim, M.O.; Lee, J.; Park, D. Melanogenesis inhibitory effect of fatty acid alkyl esters isolated from Oxalis triangularis. Biol. Pharm. Bull. 2010, 33, 1242–1245. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bhardwaj, M.; Sali, V.K.; Mani, S.; Vasanthi, H.R. Neophytadiene from Turbinaria ornata suppresses LPS-induced inflammatory response in RAW 264.7 macrophages and Sprague Dawley rats. Inflammation 2020, 43, 937–950. [Google Scholar] [CrossRef]
- Baldermann, S.; Kato, M.; Kurosawa, M.; Kurobayashi, Y.; Fujita, A.; Fleischmann, P.; Watanabe, N. Functional characterization of a carotenoid cleavage dioxygenase 1 and its relation to the carotenoid accumulation and volatile emission during the floral development of Osmanthus fragrans Lour. J. Exp. Bot. 2010, 61, 2967–2977. [Google Scholar] [CrossRef] [Green Version]
- Ribeiro, B.D.; Barreto, D.W.; Coelho, M.A.Z. Technological aspects of β-carotene production. Food Bioprocess. Technol. 2011, 4, 693–701. [Google Scholar] [CrossRef]
- Parker, G.L.; Smith, L.K.; Baxendale, I.R. Development of the industrial synthesis of vitamin A. Tetrahedron 2016, 72, 1645–1652. [Google Scholar] [CrossRef]
- Janakiram, N.B.; Cooma, I.; Mohammed, A.; Steele, V.E.; Rao, C.V. Beta-ionone inhibits colonic aberrant crypt foci formation in rats, suppresses cell growth, and induces retinoid X receptor-alpha in human colon cancer cells. Mol. Cancer Ther. 2008, 7, 181–190. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kang, C.H.; Jayasooriya, R.G.; Choi, Y.H.; Moon, S.K.; Kim, W.J.; Kim, G.Y. β-Ionone attenuates LPS-induced pro-inflammatory mediators such as NO, PGE2 and TNF-α in BV2 microglial cells via suppression of the NF-κB and MAPK pathway. Toxicol. In Vitro 2013, 27, 782–787. [Google Scholar] [CrossRef]
- Das, M.; Prakash, S.; Nayak, C.; Thangavel, N.; Singh, S.K.; Manisankar, P.; Devi, K.P. Dihydroactinidiolide, a natural product against Aβ25–35 induced toxicity in Neuro2A cells: Synthesis, in silico and in vitro studies. Bioorg. Chem. 2018, 81, 340–349. [Google Scholar] [CrossRef] [PubMed]
- Fenini, G.; Contassot, E.; French, L.E. Potential of IL-1, IL-18 and inflammasome inhibition for the treatment of inflammatory skin diseases. Front. Pharmacol. 2017, 8, 278. [Google Scholar] [CrossRef]
- Kistowska, M.; Gehrke, S.; Jankovic, D.; Kerl, K.; Fettelschoss, A.; Feldmeyer, L.; Fenini, G.; Kolios, A.; Navarini, A.; Ganceviciene, R.; et al. IL-1β drives inflammatory responses to Propionibacterium acnes in vitro and in vivo. J. Investig. Dermatol. 2014, 134, 677–685. [Google Scholar] [CrossRef] [Green Version]
- Franchi, L.; Eigenbrod, T.; Muñoz-Planillo, R.; Nuñez, G. The inflammasome: A caspase-1 activation platform regulating immune responses and disease pathogenesis. Nat. Immunol. 2009, 10, 241. [Google Scholar] [CrossRef]
- Sahdo, B.; Särndahl, E.; Elgh, F.; Söderquist, B. Propionibacterium acnes activates caspase-1 in human neutrophils. APMIS 2013, 121, 652–663. [Google Scholar] [CrossRef]
- Li, Z.J.; Choi, D.K.; Sohn, K.C.; Seo, M.S.; Lee, H.E.; Lee, Y.; Seo, Y.J.; Lee, Y.H.; Shi, G.; Zouboulis, C.C.; et al. Propionibacterium acnes activates the NLRP3 inflammasome in human sebocytes. J. Investig. Dermatol. 2014, 134, 2747–2756. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Aubin, G.G.; Baud’huin, M.; Lavigne, J.P.; Régis, B.; François, G.; Didier, L.; Cédric, J.; Dominique, H.; Karim, A.; Stéphane, C. Interaction of Cutibacterium (formerly Propionibacterium) acnes with bone cells: A step toward understanding bone and joint infection development. Sci. Rep. 2017, 7, 42918. [Google Scholar] [CrossRef] [PubMed] [Green Version]
Sample Availability: Samples of the compounds are not available from the authors. |









{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| No. | RT (min) | Identification | EIMS Fragments (70 eV), m/z (Relative Abundance) | Molecular Formula | % of Total |
|---|---|---|---|---|---|
| 1 | 7.8 | trans, trans-2,4-heptadienal | 110 (M+, 25), 81 (100), 68 (14), 53 (20), 41 (20) | C7H10O | 1.3 |
| 2 | 18.25 | β-ionone | 192 (M+, 4), 177 (100), 135 (14), 123 (100), 43 (45) | C13H20O | 1.3 |
| 3 | 19.14 | dihydroactinidiolide | 180 (M+, 24), 137 (42), 111 (100), 67 (23), 43 (35) | C11H16O2 | 2.1 |
| 4 | 19.80 | cis-3-hexenyl benzoate | 105 (100), 82 (83), 77 (44), 67 (56), 51 (10) | C13H16O2 | 1.7 |
| 5 | 24.24 | neophytadiene | 278 (M+, 3), 95 (89), 82 (80), 68 (100), 57 (69) | C20H38 | 1.5 |
| 6 | 24.34 | 6,10,14-trimethyl-2-pentadecanone | 109 (28), 85 (35), 71 (53), 58 (97), 43 (100) | C18H36O | 0.9 |
| 7 | 24.91 | phytol | 123 (63), 95 (88), 82 (100), 68 (72), 57 (66) | C20H40O | 0.6 |
| 8 | 25.26 | palmitoleic acid, methyl ester | 268 (M+, 4), 236 (21), 83 (51), 69 (73), 55 (100) | C17H32O2 | 0.3 |
| 9 | 25.56 | palmitic acid, methyl ester | 270 (M+, 12), 227 (15), 143 (21), 87 (69), 74 (100) | C17H34O2 | 9.9 |
| 10 | 26.58 | palmitic acid, ethyl ester | 284 (M+, 12), 101 (52), 88 (100), 73 (19), 55 (28) | C18H36O2 | 0.2 |
| 11 | 28.05 | linoleic acid, methyl ester | 294 (M+, 18), 95 (72), 81 (100), 67 (100), 55 (64) | C19H34O2 | 0.8 |
| 12 | 28.16 | linolenic acid, methyl ester | 292 (M+, 6), 108 (43), 95 (59), 79 (100), 67 (61) | C19H32O2 | 7 |
| 13 | 28.48 | stearic acid, methyl ester | 298 (M+, 16), 143 (22), 87 (69), 74 (100), 43 (28) | C19H38O2 | 1.5 |
| 14 | 31.15 | eicosanoic acid, methyl ester | 326 (M+, 25), 87 (77), 74 (100), 55 (51), 43 (54) | C21H42O2 | 0.5 |
| 15 | 31.50 | 4,8,12,16-tetramethylheptadecan-4-olide | 126 (14), 99 (100), 83 (24), 69 (25), 43 (30) | C21H40O2 | 0.2 |
| 16 | 31.80 | dodecanoic acid, 2-hexen-l-yl ester | 82 (100), 67 (23), 55 (18), 43 (15) | C18H34O2 | 0.9 |
| 17 | 33.23 | n-pentacosane | 352 (M+, 3), 85 (56), 71 (77), 57 (100), 43 (56) | C25H52 | 0.3 |
| 18 | 33.60 | docosanoic acid, methyl ester | 354 (M+, 30), 143 (24), 87 (76), 74 (100), 43 (37) | C23H46O2 | 0.4 |
| 19 | 35.52 | n-heptacosane | 380 (M+,3), 85 (56), 71 (83), 57 (100), 43 (56) | C27H56 | 0.4 |
| 20 | 35.89 | tetracosanoic acid, methyl ester | 382 (M+, 34), 143 (31), 87 (70), 74 (100), 43 (47) | C25H50O2 | 0.3 |
| 21 | 37.01 | squalene | 410 (M+, 1), 95 (15), 81 (58), 69 (100), 41 (20) | C30H50 | 12.2 |
| 22 | 39.89 | γ-tocopherol | 416 (M+, 90), 191 (18), 151 (100) | C28H48O2 | 2.6 |
| 23 | 41.30 | α-tocopherol | 430(M+, 100), 205 (10), 165 (100) | C29H50O2 | 13.6 |
| 24 | 45.77 | 3-phytylmenadione (vitamin K1) | 450(M+, 100), 225 (58), 198 (44), 186 (52) | C31H46O2 | 15.8 |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Chuang, L.-T.; Shih, Y.-H.; Huang, W.-C.; Lin, L.-C.; Hsu, C.; Chyuan, J.-H.; Tsai, T.-H.; Tsai, P.-J. In Vitro and In Vivo Screening of Wild Bitter Melon Leaf for Anti-Inflammatory Activity against Cutibacterium acnes. Molecules 2020, 25, 4277. https://doi.org/10.3390/molecules25184277
Chuang L-T, Shih Y-H, Huang W-C, Lin L-C, Hsu C, Chyuan J-H, Tsai T-H, Tsai P-J. In Vitro and In Vivo Screening of Wild Bitter Melon Leaf for Anti-Inflammatory Activity against Cutibacterium acnes. Molecules. 2020; 25(18):4277. https://doi.org/10.3390/molecules25184277
Chicago/Turabian StyleChuang, Lu-Te, Ya-Hsin Shih, Wen-Cheng Huang, Lie-Chwen Lin, Chin Hsu, Jong-Ho Chyuan, Tsung-Hsien Tsai, and Po-Jung Tsai. 2020. "In Vitro and In Vivo Screening of Wild Bitter Melon Leaf for Anti-Inflammatory Activity against Cutibacterium acnes" Molecules 25, no. 18: 4277. https://doi.org/10.3390/molecules25184277