The War against Tuberculosis: A Review of Natural Compounds and Their Derivatives

 and
and
Abstract
:1. Introduction
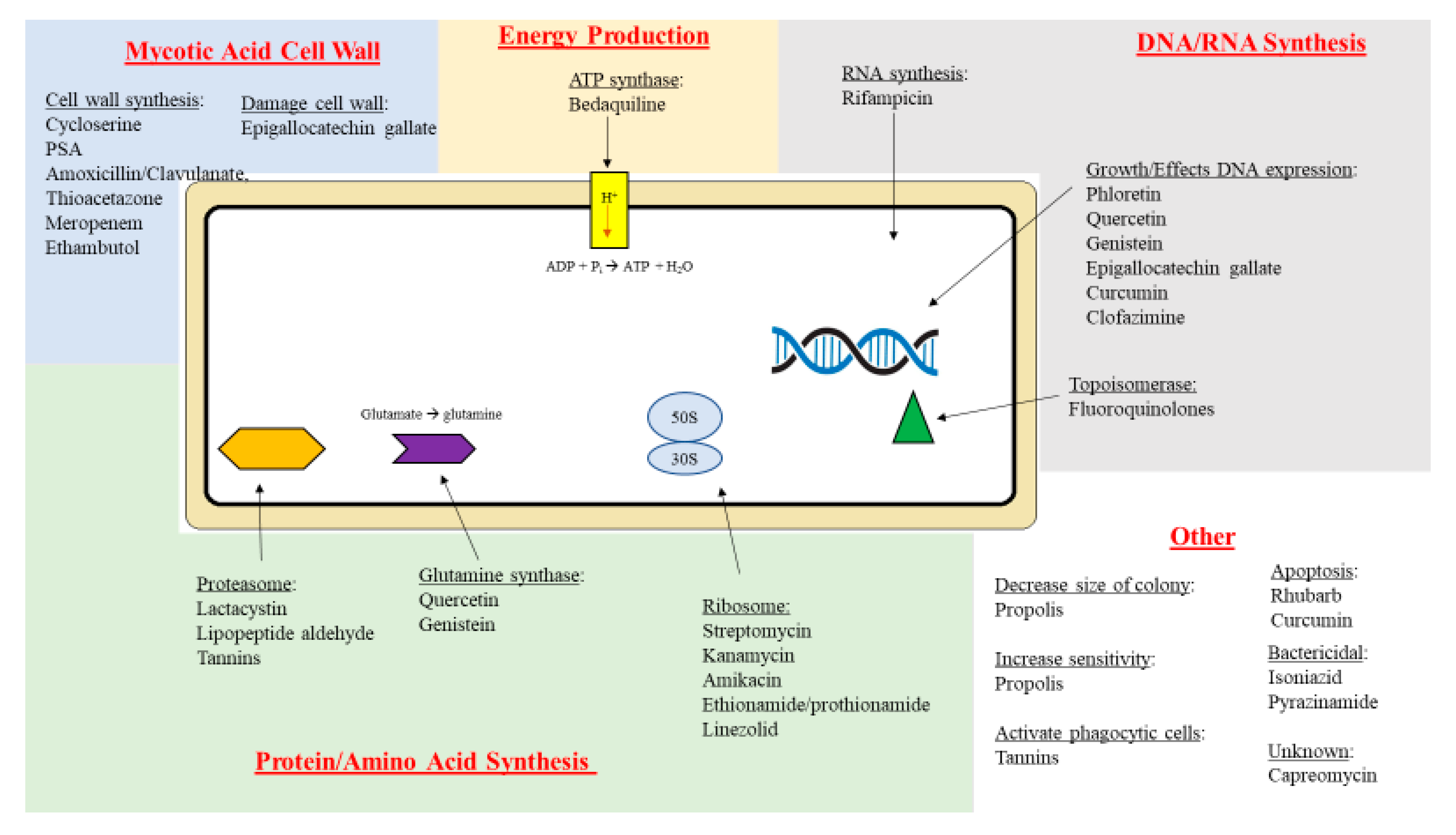
2. Therapeutic Targets
3. Treatment
3.1. Newly Diagnosed Patients
3.2. Multidrug-Resistant (MDR) and Extensively Drug-Resistant (XDR) Tuberculosis
3.3. Important Factors in M. tuberculosis Infections and Limitations of Treatment
4. Natural Compounds
4.1. Curcumin
4.2. Phloretin
4.3. Quercetin
4.4. Tannins
4.5. Tea Polyphenols
4.6. Resveratrol
4.7. Propolis
4.8. Lactacystin
4.9. Fellutamide B
5. Conclusions and Future Directions
Author Contributions
Funding
Conflicts of Interest
References
- Holmes, K.K.; Bertozzi, S.; Bloom, B.R.; Jha, P.; Gelband, H.; DeMaria, L.M.; Horton, S. Major Infectious Diseases: Key Messages From Disease Control Priorities, 3rd ed.; The International Bank for Reconstruction and Development/The World Bank: Washington, DC, USA, 2017; Volume 3. [Google Scholar]
- Magge, J.G.; Ward, A.C. Mycobacterium. Bergey’s Man. Syst. Archaea Bact. 2015, 1–84. [Google Scholar] [CrossRef]
- Alderwick, L.; Harrison, J.; Lloyd, G.S.; Birch, H.L. The Mycobacterial Cell Wall - Peptidoglycan and Aradinogalactan. Cold Spring Harb. Perspect. Med. 2015, 5, 8. [Google Scholar] [CrossRef] [Green Version]
- Chiaradia, L.; Lefebvre, C.; Parra, J.; Marcoux, J.; Burlet-Schiltz, O.; Etienne, G.; Tropis, M.; Daffé, M. Dissecting the mycobacterial ell envelope and defining the composition of the native mycomembrane. Sci. Rep. 2017, 7, 1–12. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pai, M.; Schito, M. Tuberculosis Diagnostics in 2015: Landscape, Priorities, Needs, and Prospects. J. Infect. Dis. 2015, 211, S21–S28. [Google Scholar] [CrossRef]
- Delogu, G.; Sali, M.; Fadda, G. The Biology of Mycobacterium Tuberculosis Infection. Mediterr. J. Hematol. Infect. Dis. 2013, 5, e2013070. [Google Scholar] [CrossRef] [Green Version]
- Turner, R.D.; Bothamley, G.H. Cough and the Transmission of Tuberculosis. J. Infect. Dis. 2015, 211, 1367–1372. [Google Scholar] [CrossRef] [Green Version]
- Churchyard, G.; Kim, P.; Shah, N.S.; Rustomjee, R.; Gandhi, N.; Mathema, B.; Dowdy, D.; Kasmar, A.; Cardenas, V. What We Know About Tuberculosis Transmission: An Overview. J. Infect. Dis. 2017, 216, S629–S635. [Google Scholar] [CrossRef] [Green Version]
- Christianakis, S.; Banerjee, J.; Kim, J.S.; Jones, B.E. Tuberculosis in Immunocompromised Hosts. In Challenging Cases in Pulmonology; Springer Science Los Angeles: New York, NY, USA, 2012. [Google Scholar]
- Guirado, E.; Schlesinger, L.S. Modeling the Mycobacterium tuberculosis Granuloma – the Critical Battlefield in Host Immunity and Disease. Front. Immunol. 2013, 4, 98. [Google Scholar] [CrossRef] [Green Version]
- Orme, I.M.; Basaraba, R.J. The formation of the granuloma in tuberculosis infection. Semin. Immunol. 2014, 26, 601–609. [Google Scholar] [CrossRef]
- Al-Humadi, H.; Al-Saigh, R.; Al-Humadi, A. Addressing the Challenges of Tuberculosis: A Brief Historical Account. Front. Pharmacol. 2017, 8, 689. [Google Scholar] [CrossRef] [Green Version]
- Getahun, H.; Matteelli, A.; Chaisson, R.E.; Raviglione, M. Latent Mycobacterium tuberculosis Infection. New Engl. J. Med. 2015, 372, 2127–2135. [Google Scholar] [CrossRef] [Green Version]
- Drain, P.K.; Bajema, K.L.; Dowdy, D.; Dheda, K.; Naidoo, K.; Schumacher, S.G.; Ma, S.; Meermeier, E.; Lewinsohn, D.M.; Sherman, D.R. Incipient and Subclinical Tuberculosis: A Clinical Review of Early Stages and Progression of Infection. Clin. Microbiol. Rev. 2018, 31, e00021-18. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pai, M.; Behr, M.A.; Dowdy, D.; Dheda, K.; Divangahi, M.; Boehme, C.C.; Ginsberg, A.; Swaminathan, S.; Spigelman, M.; Getahun, H.; et al. Tuberculosis. Nat. Rev. Dis. Prim. 2016, 2, 16076. [Google Scholar] [CrossRef]
- Parsons, L.M.; Somoskövi, Á.; Gutierrez, C.; Lee, E.; Paramasivan, C.N.; Abimiku, A.L.; Spector, S.; Roscigno, G.; Nkengasong, J. Laboratory Diagnosis of Tuberculosis in Resource-Poor Countries: Challenges and Opportunities. Clin. Microbiol. Rev. 2011, 24, 314–350. [Google Scholar] [CrossRef] [Green Version]
- Lewinsohn, D.M.; Leonard, M.K.; LoBue, P.A.; Cohn, D.L.; Daley, C.L.; Desmond, E.; Keane, J.; Lewinsohn, D.A.; Loeffler, A.M.; Mazurek, G.H.; et al. Official American Thoracic Society/Infectious Diseases Society of America/Centers for Disease Control and Prevention Clinical Practice Guidelines: Diagnosis of Tuberculosis in Adults and Children. Clin. Infect. Dis. 2017, 64, 111–115. [Google Scholar] [CrossRef]
- Azadi, D.; Motallebirad, T.; Ghaffari, K.; Shojaei, H. Mycobacteriosis and Tuberculosis: Laboratory Diagnosis. Open Microbiol. J. 2018, 12, 41–58. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Forbes, B.A.; Hall, G.S.; Miller, M.B.; Novak, S.M.; Rowlinson, M.-C.; Salfinger, M.; Somoskövi, A.; Warshauer, D.M.; Wilson, M.L. Practice Guidelines for Clinical Microbiology Laboratories: Mycobacteria. Clin. Microbiol. Rev. 2018, 31, 00038-17. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hendon-Dunn, C.L.; Doris, K.S.; Thomas, S.R.; Allnutt, J.C.; Marriott, A.A.N.; Hatch, K.A.; Watson, R.J.; Bottley, G.; Marsh, P.D.; Taylor, S.C.; et al. A Flow Cytometry Method for Rapidly Assessing Mycobacterium tuberculosis Responses to Antibiotics with Different Modes of Action. Antimicrob. Agents Chemother. 2016, 60, 3869–3883. [Google Scholar] [CrossRef] [Green Version]
- Lawn, S.D.; Zumla, A. Tuberculosis. Lancet 2011, 378, 57–72. [Google Scholar] [CrossRef]
- Global Tuberculosis Report 2019. Available online: https://www.who.int/tb/global-report-2019 (accessed on 26 May 2019).
- Tiberi, S.; Du Plessis, N.; Walzl, G.; Vjecha, M.J.; Rao, M.; Ntoumi, F.; Mfinanga, S.; Kapata, N.; Mwaba, P.; McHugh, T.D.; et al. Tuberculosis: Progress and advances in development of new drugs, treatment regimens, and host-directed therapies. Lancet Infect. Dis. 2018, 18, e183-198. [Google Scholar] [CrossRef]
- Efremenko, Y.V.; Arjanova, O.V.; Prihoda, N.D.; Yurchenko, L.V.; I Sokolenko, N.; Mospan, I.V.; Pylypchuk, V.S.; Rowe, J.; Jirathitikal, V.; Bourinbaiar, A.S.; et al. Clinical validation of sublingual formulations of Immunoxel (Dzherelo) as an adjuvant immunotherapy in treatment of TB patients. Immunotherapy 2012, 4, 273–282. [Google Scholar] [CrossRef] [PubMed]
- Ahmad, S.; Mokaddas, E. Recent advances in the diagnosis and treatment of multidrug-resistant tuberculosis. Respir. Med. 2009, 103, 1777–1790. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Van Boeckel, T.P.; Pires, J.; Silvester, R.; Zhao, C.; Song, J.; Criscuolo, N.G.; Gilbert, M.; Bonhoeffer, S.; Laxminarayan, R. Global trends in antimicrobial resistance in animals in low- and middle-income countries. Science 2019, 365, eaaw1944. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Global tuberculosis report 2015 20th edition. Available online: https://apps.who.int/iris/handle/10665/191102 (accessed on 6 December 2015).
- Fitzwater, S.P.; Sechler, G.A.; Jave, O.; Coronel, J.; Mendoza, A.; Gilman, R.H.; Friedland, J.S.; Moore, D.A.J. Second-line anti-tuberculosis drug concentrations for susceptibility testing in the MODS assay. Eur. Respir. J. 2012, 41, 1163–1171. [Google Scholar] [CrossRef] [Green Version]
- Alene, K.A.; Yi, H.; Viney, K.; McBryde, E.; Yang, K.; Bai, L.; Gray, D.; Clements, A.; Xu, Z. Treatment outcomes of patients with multidrug-resistant and extensively drug resistant tuberculosis in Hunan Province, China. BMC Infect. Dis. 2017, 17, 573. [Google Scholar] [CrossRef] [Green Version]
- Companion Handbook to the WHO Guidelines for the Programmatic Management of Drug-Resistant Tuberculosis Treatment strategies for MDR-TB and XDR-TB. Available online: https://www.ncbi.nlm.nih.gov/books/NBK247431/ (accessed on 6 December 2019).
- Salomon, C.E.; Schmidt, L.E. Natural products as leads for tuberculosis drug development. Curr. Top. Med. Chem. 2012, 12, 735–765. [Google Scholar] [CrossRef]
- Mbaveng, A.T.; Sandjo, L.P.; Tankeo, S.B.; Ndifor, A.R.; Pantaleon, A.; Ngadjui, B.T.; Kuete, V. Antibacterial activity of nineteen selected natural products against multi-drug resistant Gram-negative phenotypes. SpringerPlus 2015, 4, 823. [Google Scholar] [CrossRef] [Green Version]
- Bhat, Z.S.; Rather, M.A.; Maqbool, M.; Ahmad, Z. Drug targets exploited in Mycobacterium tuberculosis: Pitfalls and promises on the horizon. Biomed. Pharmacother. 2018, 103, 1733–1747. [Google Scholar] [CrossRef]
- Shetty, A.; Dick, T. Mycobacterial Cell Wall Synthesis Inhibitors Cause Lethal ATP Burst. Front. Microbiol. 2018, 9, 9. [Google Scholar] [CrossRef] [Green Version]
- Reiche, M.A.; Warner, D.F.; Mizrahi, V. Targeting DNA Replication and Repair for the Development of Novel Therapeutics against Tuberculosis. Front. Mol. Biosci. 2017, 4, 75. [Google Scholar] [CrossRef] [Green Version]
- Scharf, N.; Molodtsov, V.; Kontos, A.; Murakami, K.S.; Garcia, G.A. Novel Chemical Scaffolds for Inhibition of Rifamycin-Resistant RNA Polymerase Discovered from High-Throughput Screening. SLAS Discov. Adv. Life Sci. R&D 2016, 22, 287–297. [Google Scholar] [CrossRef] [Green Version]
- Minato, Y.; Thiede, J.M.; Kordus, S.L.; McKlveen, E.J.; Turman, B.J.; Baughn, A.D. Mycobacterium tuberculosis Folate Metabolism and the Mechanistic Basis forpara-Aminosalicylic Acid Susceptibility and Resistance. Antimicrob. Agents Chemother. 2015, 59, 5097–5106. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Huisjes, R.; Card, D.J. Laboratory Assessment of Vitamin Status. Methods for assessment of panthothenic acid (Vitamin B5); Elsevier Inc.: London, UK, 2018. [Google Scholar]
- Laurenzi, M.; Ginsberg, A.; Spigelman, M. Challenges associated with current and future TB treatment. Infect. Disord. - Drug Targets 2007, 7, 105–119. [Google Scholar] [CrossRef] [Green Version]
- Schito, M.; Maeurer, M.; Kim, P.; Hanna, D.; Zumla, A. Translating the Tuberculosis Research Agenda: Much Accomplished, but Much More to Be Done. Clin. Infect. Dis. 2015, 61, S95–S101. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Guillemont, J.; Meyer, C.; Poncelet, A.; Bourdrez, X.; Andries, K. Diarylquinolines, synthesis pathways and quantitative structure–activity relationship studies leading to the discovery of TMC207. Futur. Med. Chem. 2011, 3, 1345–1360. [Google Scholar] [CrossRef]
- Sacksteder, K.A.; Protopopova, M.; Barry, C.E.; Andries, K.; Nacy, C.A. Discovery and development of SQ109: A new antitubercular drug with a novel mechanism of action. Futur. Microbiol. 2012, 7, 823–837. [Google Scholar] [CrossRef] [Green Version]
- Tong, H.; Tai, Y.; Ye, C.; Wu, H.; Zhang, L.-H.; Gao, J.-H.; Yan, Z.-P.; Huang, Z.-Y.; Tang, C.-W. Carbohydrate antigen 125 and carcinoembryonic antigen in the differentiation of tuberculous peritonitis and peritonitis carcinomatosa. Oncotarget 2017, 8, 78068–78075. [Google Scholar] [CrossRef] [Green Version]
- Matteelli, A.; Carvalho, A.C.; E Dooley, K.; Kritski, A.L. TMC207: The first compound of a new class of potent anti-tuberculosis drugs. Futur. Microbiol. 2010, 5, 849–858. [Google Scholar] [CrossRef] [Green Version]
- Koul, A.; Dendouga, N.; Vergauwen, K.; Molenberghs, B.; Vranckx, L.; Willebrords, R.; Ristic, Z.; Lill, H.; Dorange, I.; Guillemont, J.; et al. Diarylquinolines target subunit c of mycobacterial ATP synthase. Nat. Methods 2007, 3, 323–324. [Google Scholar] [CrossRef]
- Andries, K. A Diarylquinoline Drug Active on the ATP Synthase of Mycobacterium tuberculosis. Science 2005, 307, 223–227. [Google Scholar] [CrossRef]
- Pérez-Camacho, I.; Rivero-Juárez, A.; Kindelán, J.M.; Rivero, A. Present-day treatment of tuberculosis and latent tuberculosis infection. Enfermedades Infecciosas y Microbiología Clínica 2011, 29, 41–46. [Google Scholar] [CrossRef]
- Treatment for TB Disease / Treatment/ TB CDC. Available online: https://www.cdc.gov/tb/topic/treatment/tbdisease.htm (accessed on 7 June 2019).
- Horsburgh, C.R.; Barry, C.E.; Lange, C. Treatment of Tuberculosis. N. Engl. J. Med. 2015, 373, 2149–2160. [Google Scholar] [CrossRef] [PubMed]
- Valencia, S.; León, M.; Losada, I.; Sequera, V.-G.; Quevedo, M.F.; García-Basteiro, A.L. How do we measure adherence to anti-tuberculosis treatment? Expert Rev. Anti-infective Ther. 2016, 15, 157–165. [Google Scholar] [CrossRef] [PubMed]
- Sotgiu, G.; Centis, R.; D’Ambrosio, L.; Migliori, G.B. Tuberculosis Treatment and Drug Regimens. Cold Spring Harb. Perspect. Med. 2015, 5, a017822. [Google Scholar] [CrossRef] [Green Version]
- Mitnick, C.; Horsburgh, C.R. Encouraging news for multidrug-resistant tuberculosis treatment. Am. J. Respir. Crit. Care Med. 2010, 182, 1337–1338. [Google Scholar] [CrossRef]
- Hartman-Adams, H.; Clark, K.; Juckett, G. Update on latent tuberculosis infection. Am. Fam. Physician 2014, 89, 889–896. [Google Scholar]
- Rée, H. Treatment of tuberculosis: Guidelines for national programmes (2nd edition). Trans. R. Soc. Trop. Med. Hyg. 1999, 93, 72. [Google Scholar] [CrossRef]
- Out, A.A.; Umoh, V.A. Is the directly observed therapy short course (DOTS) an effective strategy for tuberculosis control in a developing country? Asian Pac. J. Trop. Dis. 2013, 3, 227–231. [Google Scholar] [CrossRef]
- Seung, K.J.; Keshavjee, S.; Rich, M.L. Multidrug-Resistant Tuberculosis and Extensively Drug-Resistant Tuberculosis. Cold Spring Harb. Perspect. Med. 2015, 5, a017863. [Google Scholar] [CrossRef] [Green Version]
- Mkopi, A.; Range, N.; Lwilla, F.; Egwaga, S.; Schulze, A.; Geubbels, E.; Van Leth, F. Adherence to Tuberculosis Therapy among Patients Receiving Home-Based Directly Observed Treatment: Evidence from the United Republic of Tanzania. PLoS ONE 2012, 7, e51828. [Google Scholar] [CrossRef] [Green Version]
- Holzman, S.B.; Zenilman, A.; Shah, M. Advancing Patient-Centered Care in Tuberculosis Management: A Mixed-Methods Appraisal of Video Directly Observed Therapy. Open Forum Infect. Dis. 2018, 5, ofy046. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sotgiu, G.; Nardell, E.; Yedilbayev, A.; D’Ambrosio, L.; Centis, R.; Tadolini, M.; Boom, M.V.D.; Ehsani, S.; Sotgiu, G.; Dara, M. Reducing tuberculosis transmission: A consensus document from the World Health Organization Regional Office for Europe. Eur. Respir. J. 2019, 53, 1900391. [Google Scholar] [CrossRef]
- Padmapriyadarsini, C.; Saravanan, N.; Bethunaickan, R.; Tripathy, S. Malnutrition: Modulator of Immune Responses in Tuberculosis. Front. Immunol. 2017, 8, 1316. [Google Scholar] [CrossRef] [Green Version]
- Systematic screening for active tuberculosis: Principles and recommendations. Available online: https://www.who.int/tb/tbscreening/en/ (accessed on 16 December 2019).
- Cazabon, D.; Alsdurf, H.; Satyanarayana, S.; Nathavitharana, R.; Subbaraman, R.; Daftary, A.; Pai, M. Quality of tuberculosis care in high burden countries: The urgent need to address gaps in the care cascade. Int. J. Infect. Dis. 2016, 56, 111–116. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mauch, V.; Bonsu, F.; Gyapong, M.; Awini, E.; Suarez, P.; Marcelino, B.; E Melgen, R.; Lonnroth, K.; Nhung, N.V.; Hoa, N.B.; et al. Free tuberculosis diagnosis and treatment are not enough: Patient cost evidence from three continents. Int. J. Tuberc. Lung Dis. 2013, 17, 381–387. [Google Scholar] [CrossRef]
- Jeffries, C.; LoBue, P.; Chorba, T.; Metchock, B.; Kashef, I. Role of the Health Department in Tuberculosis Prevention and Control—Legal and Public Health Considerations. Microbiol. Spectr. 2017, 5, 261–282. [Google Scholar] [CrossRef]
- Goswami, N.D.; Mase, S.; Griffith, D.; Bhavaraju, R.; Lardizabal, A.; Lauzardo, M.; Chen, L.; Wilson, J.; Chappelle, C.; A Haley, C. Tuberculosis in the United States: Medical Consultation Services Provided by 5 Tuberculosis Regional Training and Medical Consultation Centers, 2013-2017. Open Forum Infect. Dis. 2019, 6, ofz167. [Google Scholar] [CrossRef] [Green Version]
- Balaban, V.; Marks, S.M.; Etkind, S.C.; Katz, L.J.; Higashi, J.; Flood, J.; Cronin, A.; Ho, C.S.; Khan, A.; Chorba, T. Tuberculosis Elimination Efforts in the United States in the Era of Insurance Expansion and the Affordable Care Act. Public Heal. Rep. 2015, 130, 349–354. [Google Scholar] [CrossRef] [Green Version]
- Taylor, Z.; Nolan, C.M.; Blumberg, H.M. Controlling tuberculosis in the United States. Recommendations from the American Thoracic Society, CDC, and the Infectious Diseases Society of America. MMWR. Recomm. Rep. 2005, 54. [Google Scholar]
- Truven Health Analytics, Inc, Ann Arbor, MI. Available online: http://www.micromedexsolutions.com (accessed on 16 January 2020).
- Li, W.-B.; Zhang, Y.-Q.; Xing, J.; Ma, Z.-Y.; Qu, Y.-H.; Li, X.-X. Factors associated with primary transmission of multidrug-resistant tuberculosis compared with healthy controls in Henan Province, China. Infect. Dis. Poverty 2015, 4, 14. [Google Scholar] [CrossRef] [Green Version]
- Gebreweld, F.H.; Kifle, M.M.; Gebremicheal, F.E.; Simel, L.L.; Gezae, M.M.; Ghebreyesus, S.S.; Mengsteab, Y.T.; Wahd, N.G. Factors influencing adherence to tuberculosis treatment in Asmara, Eritrea: A qualitative study. J. Heal. Popul. Nutr. 2018, 37, 1. [Google Scholar] [CrossRef] [PubMed]
- Jnawali, H.N.; Ryoo, S. First-and second-line drugs and drug resistance. Tuberc. Curr. Issues Diagn. Manag. 2013, 163–180. [Google Scholar]
- Alsultan, A.; Peloquin, C. Therapeutic Drug Monitoring in the Treatment of Tuberculosis: An Update. Drugs 2014, 74, 839–854. [Google Scholar] [CrossRef]
- Nahid, P.; Dorman, S.E.; Alipanah, N.; Barry, P.M.; Brozek, J.L.; Cattamanchi, A.; Chaisson, L.H.; Chaisson, R.E.; Daley, C.L.; Grzemska, M.; et al. Official American Thoracic Society/Centers for Disease Control and Prevention/Infectious Diseases Society of America Clinical Practice Guidelines: Treatment of Drug-Susceptible Tuberculosis. Clin. Infect. Dis. 2016, 63, e147–e195. [Google Scholar] [CrossRef] [PubMed]
- Holzman, S.B.; Atre, S.; Sahasrabudhe, T.; Ambike, S.; Jagtap, D.; Sayyad, Y.; Kakrani, A.L.; Gupta, A.; Mave, V.; Shah, M.; et al. Use of Smartphone-Based Video Directly Observed Therapy (vDOT) in Tuberculosis Care: Single-Arm, Prospective Feasibility Study. JMIR Form. Res. 2019, 3, e13411. [Google Scholar] [CrossRef]
- Dashti, Y.; Grkovic, T.; Quinn, R.J. Predicting natural product value, an exploration of anti-TB drug space. Nat. Prod. Rep. 2014, 31, 990–998. [Google Scholar] [CrossRef]
- Burman, W.J.; Gallicano, K.; Peloquin, C.; Burman, W.J. Comparative Pharmacokinetics and Pharmacodynamics of the Rifamycin Antibacterials. Clin. Pharmacokinet. 2001, 40, 327–341. [Google Scholar] [CrossRef]
- Yuan, D.; Yang, X.; Guo, J.-C. A great honor and a huge challenge for China: You-you TU getting the Nobel Prize in Physiology or Medicine. J. Zhejiang Univ. Sci. B 2016, 17, 405–408. [Google Scholar] [CrossRef] [Green Version]
- Meshnick, S.R. Artemisinin: Mechanisms of action, resistance and toxicity. Int. J. Parasitol. 2002, 32, 1655–1660. [Google Scholar] [CrossRef]
- Shandilya, A.; Chacko, S.; Jayaram, B.; Ghosh, I. A plausible mechanism for the antimalarial activity of artemisinin: A computational approach. Sci. Rep. 2013, 3, 2513. [Google Scholar] [CrossRef] [Green Version]
- Hou, X.-M.; Wang, C.-Y.; Gerwick, W.H.; Shao, C.-L. Marine natural products as potential anti-tubercular agents. Eur. J. Med. Chem. 2019, 165, 273–292. [Google Scholar] [CrossRef] [PubMed]
- Han, X.; Chen, C.; Yan, Q.; Jia, L.; Taj, A.; Ma, Y. Action of Dicumarol on Glucosamine-1-Phosphate Acetyltransferase of GlmU and Mycobacterium tuberculosis. Front. Microbiol. 2019, 10, 1799. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Shang, A.; Cao, S.-Y.; Xu, X.-Y.; Gan, R.-Y.; Tang, G.-Y.; Corke, H.; Mavumengwana, V.; Li, H.-B. Bioactive Compounds and Biological Functions of Garlic (Allium sativum L.). Foods 2019, 8, 246. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Batbold, U.; O Butov, D.; A Kutsyna, G.; Damdinpurev, N.; A Grinishina, E.; Mijiddorj, O.; E Kovolev, M.; Baasanjav, K.; Butova, T.S.; Sandagdorj, M.; et al. Double-blind, placebo-controlled, 1:1 randomized Phase III clinical trial of Immunoxel honey lozenges as an adjunct immunotherapy in 269 patients with pulmonary tuberculosis. Immunotherapy 2017, 9, 13–24. [Google Scholar] [CrossRef]
- Bai, X.; Oberley--Deegan, R.E.; Ovrutsky, A.R.; Kinney, W.H.; Weaver, M.; Zhang, G.; Honda, J.R.; Chan, E.D. Curcumin enhances human macrophage control of Mycobacterium tuberculosis infection. Respirology 2016, 21, 951–957. [Google Scholar] [CrossRef]
- Tsuchiya, S.; Yamabe, M.; Yamaguchi, Y.; Kobayashi, Y.; Konno, T.; Tada, K. Establishment and characterization of a human acute monocytic leukemia cell line (THP-1). Int. J. Cancer 1980, 26, 171–176. [Google Scholar] [CrossRef]
- Tousif, S.; Singh, D.K.; Mukherjee, S.; Ahmad, S.; Arya, R.; Nanda, R.; Ranganathan, A.; Bhattacharyya, M.; Van Kaer, L.; Kar, S.K.; et al. Nanoparticle-Formulated Curcumin Prevents Posttherapeutic Disease Reactivation and Reinfection with Mycobacterium tuberculosis following Isoniazid Therapy. Front. Immunol. 2017, 8, 8. [Google Scholar] [CrossRef] [Green Version]
- Jeon, D.; Jeong, M.-C.; Jnawali, H.N.; Kwak, C.; Ryoo, S.; Jung, I.D.; Kim, Y. Phloretin Exerts Anti-Tuberculosis Activity and Suppresses Lung Inflammation. Molecules 2017, 22, 183. [Google Scholar] [CrossRef] [Green Version]
- Németh, E.; Nagy, Z.; Pécsváradi, A. Chloroplast Glutamine Synthetase, the Key Regulatory of Nitrogen Metabolism in Wheat, Performs Its Role by Fine Regulation of Enzyme Activity via Negative Cooperativity of Its Subunits. Front. Plant. Sci. 2018, 9, 191. [Google Scholar] [CrossRef] [Green Version]
- Safwat, N.A.; Kashef, M.T.; Aziz, R.K.; Amer, K.F.; Ramadan, M.A. Quercetin 3-O-glucoside recovered from the wild Egyptian Sahara plant, Euphorbia paralias L., inhibits glutamine synthetase and has antimycobacterial activity. Tuberculosis 2018, 108, 106–113. [Google Scholar] [CrossRef]
- Shukla, H.; Kumar, V.; Singh, A.K.; Rastogi, S.; Khan, S.R.; Siddiqi, M.I.; Krishnan, M.Y.; Akhtar, S. Isocitrate lyase of Mycobacterium tuberculosis is inhibited by quercetin through binding at N-terminus. Int. J. Boil. Macromol. 2015, 78, 137–141. [Google Scholar] [CrossRef] [PubMed]
- Dey, D.; Ray, R.; Hazra, B. Antimicrobial activity of pomegranate fruit constituents against drug-resistant Mycobacterium tuberculosis and? -lactamase producing Klebsiella pneumoniae. Pharm. Boil. 2015, 53, 1–7. [Google Scholar] [CrossRef] [Green Version]
- Hickey, A.J.; Gounder, L.; Moosa, Y.; Drain, P.K. A systematic review of hepatic tuberculosis with considerations in human immunodeficiency virus co-infection. BMC Infect. Dis. 2015, 15, 209. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Singh, B.; Singh, G.; Trajkovic, V.; Sharma, P. Intracellular expression of Mycobacterium tuberculosis-specific 10-kDa antigen down-regulates macrophage B7.1 expression and nitric oxide release. Clin. Exp. Immunol. 2003, 134, 70–77. [Google Scholar] [CrossRef]
- Khlifi, D.; Hamdi, M.; El Hayouni, A.; Cazaux, S.; Souchard, J.P.; Couderc, F.; Bouajila, J. Global Chemical Composition and Antioxidant and Anti-Tuberculosis Activities of Various Extracts of Globularia alypum L. (Globulariaceae) Leaves. Molecules 2011, 16, 10592–10603. [Google Scholar] [CrossRef] [Green Version]
- Kedare, S.B.; Singh, R.P. Genesis and development of DPPH method of antioxidant assay. J. Food Sci. Technol. 2011, 48, 412–422. [Google Scholar] [CrossRef] [Green Version]
- Re, R.; Pellegrini, N.; Proteggente, A.; Pannala, A.; Yang, M.; Rice-Evans, C. Antioxidant activity applying an improved ABTS radical cation decolorization assay. Free. Radic. Boil. Med. 1999, 26, 1231–1237. [Google Scholar] [CrossRef]
- Watson, R.R.; Preedy, V.R. Bioactive Food as Dietary Interventions for Diabetes; Spondias pinnata (L.f.) Kurz. (Anacardiaceae): Profiles and Applications to Diabetes; Academic Press: Cambridge, MA, USA, 2019; Volume 2. [Google Scholar]
- Herzog, S.A.; Leikauf, G.; Jakse, H.; Siebenhofer, A.; Haeusler, M.; Berghold, A. Prevalence of anemia in pregnant women in Styria, Austria—A retrospective analysis of mother-child examinations 2006–2014. PLoS ONE 2019, 14, e0219703. [Google Scholar] [CrossRef]
- Soh, A.Z.; Pan, A.; Chee, C.B.E.; Wang, Y.-T.; Yuan, J.-M.; Koh, W.-P. Tea Drinking and Its Association with Active Tuberculosis Incidence among Middle-Aged and Elderly Adults: The Singapore Chinese Health Study. Nutrients 2017, 9, 544. [Google Scholar] [CrossRef] [Green Version]
- Sun, T.; Qin, B.; Gao, M.; Yin, Y.-L.; Wang, C.; Zang, S.; Li, X.; Zhang, C.; Xin, Y.; Jiang, T. Effects of epigallocatechin gallate on the cell-wall structure of Mycobacterial smegmatis mc 2 155. Nat. Prod. Res. 2014, 29, 1–3. [Google Scholar] [CrossRef]
- Levy, C.W.; Roujeinikova, A.; Sedelnikova, S.; Baker, P.; Stuitje, A.R.; Slabas, A.R.; Rice, D.W.; Rafferty, J. Molecular basis of triclosan activity. Nature 1999, 398, 383–384. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Slayden, R.; Lee, R.E.; Barry, C.E. Isoniazid affects multiple components of the type II fatty acid synthase system of Mycobacterium tuberculosis. Mol. Microbiol. 2000, 38, 514–525. [Google Scholar] [CrossRef] [Green Version]
- Scheller, S.; Dworniczak, S.; Waldemar-Klimmek, K.; Rajca, M.; Tomczyk, A.; Shani, J. Synergism between ethanolic extract of propolis (EEP) and anti-tuberculosis drugs on growth of mycobacteria. Zeitschrift für Naturforschung C 1999, 54, 549–553. [Google Scholar] [CrossRef] [PubMed]
- Ōmura, S.; Crump, A. Lactacystin: First-in-class proteasome inhibitor still excelling and an exemplar for future antibiotic research. J. Antibiot. 2019, 72, 189–201. [Google Scholar] [CrossRef] [Green Version]
- Lin, G.; Li, N.; Chidawanyika, T.; Nathan, C.; Li, H. Fellutamide B is a potent inhibitor of the Mycobacterium tuberculosis proteasome. Arch. Biochem. Biophys. 2010, 501, 214–220. [Google Scholar] [CrossRef] [Green Version]
- Li, A.-N.; Li, S.; Zhang, Y.-J.; Xu, X.-R.; Chen, Y.-M.; Li, H.-B. Resources and Biological Activities of Natural Polyphenols. Nutrients 2014, 6, 6020–6047. [Google Scholar] [CrossRef] [PubMed]
- Akbik, D.; Ghadiri, M.; Chrzanowski, W.; Rohanizadeh, R. Curcumin as a wound healing agent. Life Sci. 2014, 116, 1–7. [Google Scholar] [CrossRef]
- Tousif, S.; Singh, D.K.; Ahmad, S.; Moodley, P.; Bhattacharyya, M.; Van Kaer, L.; Das, G. Isoniazid induces apoptosis of activated CD4+ T cells: Implications for post-therapy tuberculosis reactivation and reinfection. J. Boil. Chem. 2014, 289, 30190–30195. [Google Scholar] [CrossRef] [Green Version]
- Jaisinghani, R.N. Antibacterial properties of quercetin. Microbiol. Res. 2017, 8. [Google Scholar] [CrossRef] [Green Version]
- Zu Bentrup, K.H.; Miczak, A.; Swenson, D.L.; Russell, D.G. Characterization of Activity and Experssion of Isocitrate Lyase in Mycobacterium avium and Mycobacterium tuberculosis. J. Bacteriol. 1999, 181, 7161–7167. [Google Scholar]
- Chung, K.-T.; Wong, T.Y.; Wei, C.-I.; Huang, Y.-W.; Lin, Y. Tannins and Human Health: A Review. Crit. Rev. Food Sci. Nutr. 1998, 38, 421–464. [Google Scholar] [CrossRef] [PubMed]
- Ahmad, N.; Mukhtar, H. Green tea polyphenols and cancer: Biologic mechanisms and practical implications. Nutr. Rev. 1999, 57, 78–83. [Google Scholar] [CrossRef] [PubMed]
- Higdon, J.V.; Frei, B. Tea catechins and polyphenols: Health effects, metabolism, and antioxidant functions. Crit. Rev. Food Sci. Nutr. 2003, 43, 89–143. [Google Scholar] [CrossRef] [PubMed]
- Khan, A.; Ali, N.H.; Santercole, V.; Paglietti, B.; Rubino, S.; Kazmi, S.U.; Farooqui, A. Camellia sinensisMediated Enhancement of Humoral Immunity to Particulate and Non-particulate Antigens. Phytother. Res. 2015, 30, 41–48. [Google Scholar] [CrossRef]
- Li, Y.; Jiang, X.; Hao, J.; Zhang, Y.; Huang, R. Tea polyphenols: Application in the control of oral microorganism infectious diseases. Arch. Oral Boil. 2019, 102, 74–82. [Google Scholar] [CrossRef]
- Khokhar, S.; Magnusdottir, S.G.M. Total Phenol, Catechin, and Caffeine Contents of Teas Commonly Consumed in the United Kingdom. J. Agric. Food Chem. 2002, 50, 565–570. [Google Scholar] [CrossRef]
- Sharma, S.K.; Kumar, G.; Kapoor, M.; Surolia, A. Combined effect of epigallocatechin gallate and triclosan on enoyl-ACP reductase of Mycobacterium tuberculosis. Biochem. Biophys. Res. Commun. 2008, 368, 12–17. [Google Scholar] [CrossRef]
- Seto, S.; Tsujimura, K.; Koide, Y. Coronin-1a inhibits autophagosome formation around Mycobacterium tuberculosis-containing phagosomes and assists mycobacterial survival in macrophages. Cell. Microbiol. 2012, 14, 710–727. [Google Scholar] [CrossRef]
- Anand, P.K.; Kaul, D.; Sharma, M. Green tea polyphenol inhibits Mycobacterium tuberculosis survival within human macrophages. Int. J. Biochem. Cell Boil. 2006, 38, 600–609. [Google Scholar] [CrossRef]
- Weatherly, L.M.; Gosse, J. Triclosan exposure, transformation, and human health effects. J. Toxicol. Environ. Heal. Part. B 2017, 20, 447–469. [Google Scholar] [CrossRef]
- Saw, W.-G.; Wu, M.L.; Ragunathan, P.; Biuković, G.; Lau, A.M.; Shin, J.; Harikishore, A.; Cheung, C.Y.; Hards, K.; Sarathy, J.P.; et al. Disrupting coupling within mycobacterial F-ATP synthases subunit E causes dysregulated energy production and cell wall biosynthesis. Sci. Rep. 2019, 9, 16759. [Google Scholar] [CrossRef] [PubMed]
- Smolarz, H.D.; Swatko-Ossor, M.; Ginalska, G.; Medyńska, E. Antimycobacterial Effect of Extract and Its Components from Rheum rhaponticum. J. AOAC Int. 2013, 96, 155–160. [Google Scholar] [CrossRef] [PubMed]
- Sun, D.; Hurdle, J.G.; Lee, R.; Lee, R.E.; Cushman, M.; Pezzuto, J.M. Evaluation of flavonoid and resveratrol chemical libraries reveals abyssinone II as a promising antibacterial lead. ChemMedChem 2012, 7, 1541–1545. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Teixeira, É.W.; Message, D.; Meira, R.M. Methacrylate: An alternative fixing agent for identifying the botanical origin of propolis. Appl. Plant Sci. 2019, 7, e11309. [Google Scholar] [CrossRef] [Green Version]
- Zabaiou, N.; Fouache, A.; Trousson, A.; Baron, S.; Zellagui, A.; Lahouel, M.; Lobaccaro, J.-M.A. Biological properties of propolis extracts: Something new from an ancient product. Chem. Phys. Lipids 2017, 207, 214–222. [Google Scholar] [CrossRef]
- Szliszka, E.; Zydowicz, G.; Janoszka, B.; Dobosz, C.; Kowalczyk-Ziomek, G.; Krol, W. Ethanolic extract of Brazilian green propolis sensitizes prostate cancer cells to TRAIL-induced apoptosis. Int. J. Oncol. 2011, 38, 941–953. [Google Scholar] [CrossRef] [Green Version]
- Latham, M.P.; Sekhar, A.; E Kay, L. Understanding the mechanism of proteasome 20S core particle gating. Proc. Natl. Acad. Sci. USA 2014, 111, 5532–5537. [Google Scholar] [CrossRef] [Green Version]
- Becker, S.H.; Jastrab, J.B.; Dhabaria, A.; Chaton, C.T.; Rush, J.S.; Korotkov, K.V.; Ueberheide, B.; Darwin, K.H. The Mycobacterium tuberculosis Pup-proteasome system regulates nitrate metabolism through an essential protein quality control pathway. Proc. Natl. Acad. Sci. USA 2019, 116, 3202–3210. [Google Scholar] [CrossRef] [Green Version]
- Lin, G.; Li, N.; De Carvalho, L.P.S.; Deng, H.; Tao, H.; Vogt, G.; Wu, K.; Schneider, J.; Chidawanyika, T.; Warren, J.D.; et al. Inhibitors selective for mycobacterial versus human proteasomes. Nature 2009, 461, 621–626. [Google Scholar] [CrossRef]
- Hines, J.; Groll, M.; Fahnestock, M.; Crews, C.M. Proteasome Inhibition by Fellutamide B Induces Nerve Growth Factor Synthesis. Chem. Boil. 2008, 15, 501–512. [Google Scholar] [CrossRef] [Green Version]
- Scazzocchio, F.; D’Auria, F.; Alessandrini, D.; Pantanella, F. Multifactorial aspects of antimicrobial activity of propolis. Microbiol. Res. 2006, 161, 327–333. [Google Scholar] [CrossRef] [PubMed]
- Liu, Q.; Han, F.; Pan, L.; Jia, H.; Li, Q.; Zhang, Z. Inflammation responses in patients with pulmonary tuberculosis in an intensive care unit. Exp. Ther. Med. 2018, 15, 2719–2726. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Goodman, S.N.; Fanelli, D.; Ioannidis, J.P.A. What does research reproductivity mean? Sci. Integr. 2016, 8, 341ps312. [Google Scholar]
- Wallach, J.D.; Boyack, K.W.; Ioannidis, J. Reproducible research practices, transparency, and open access data in the biomedical literature, 2015–2017. PLoS Boil. 2018, 16, e2006930. [Google Scholar] [CrossRef]
- Tu, Y. The discovery of artemisinin (qinghaosu) and gifts from Chinese medicine. Nat. Med. 2011, 17, 1217–1220. [Google Scholar] [CrossRef]
- Zhang, A.; Sun, H.; Wang, X. Recent advances in natural products from plants for treatment of liver diseases. Eur. J. Med. Chem. 2013, 63, 570–577. [Google Scholar] [CrossRef]
- DiMasi, J.A.; Grabowski, H.G.; Hansen, R.W. Innovation in the pharmaceutical industry: New estimates of R&D costs. J. Heal. Econ. 2016, 47, 20–33. [Google Scholar] [CrossRef] [Green Version]
- Wang, R.; Zhao, S.; Wang, Z.; Koffas, M.A.G. Recent advances in modular co-culture engineering for synthesis of natural products. Curr. Opin. Biotechnol. 2020, 62, 65–71. [Google Scholar] [CrossRef] [PubMed]
- Mdluli, K.; Kaneko, T.; Upton, A. The Tuberculosis Drug Discovery and Development Pipeline and Emerging Drug Targets. Cold Spring Harb. Perspect. Med. 2015, 5, a021154. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kellogg, J.J.; Paine, M.; McCune, J.S.; Oberlies, N.H.; Cech, N.B. Selection and characterization of botanical natural products for research studies: A NaPDI center recommended approach. Nat. Prod. Rep. 2019, 36, 1196–1221. [Google Scholar] [CrossRef] [PubMed] [Green Version]


{kind=link}
| Anti-TB Therapy | ||||
|---|---|---|---|---|
| First-Line Defense (Treatment for TB) | Brand | Generic | Mechanism of Action | Year of Approval |
| Nydrazid | Isoniazid * | Bactericidal agent; inhibits the enoyl-acyl carrier protein reductase InhA upon Kat activation (important for virulence); related to mycolic acid synthesis | 1952 | |
| Rifadin, Rimactane | Rifampicin * | Inhibits bacterial RNA synthesis | 1971 | |
| Myambutol | Ethambutol * | Inhibits arabinosyl transferase in bacteriostatic manner | 1961 | |
| Pyrazinamide | Pyrazinamide * | Interferes with mRNA binding Bacteriostatic/bactericidal | 1972 | |
| Streptomycin | Streptomycin sulfate *, Streptomycin nitrate | Inhibits bacterial protein synthesis | 1998 | |
| Second-Line Defense (Treatment for resistant TB) | Group A (Fluoroquinolones): | |||
| Levaquin, Quixin, Iquix | Levofloxacin | Interferes with topoisomerase IV and DNA gyrase (DNA replication) | 1996 | |
| Avelox, Vigamox, Moxeza | Moxifloxacin | Inhibits topoisomerases II and IV (DNA replication) | 1999 | |
| Tequin, Zymar, Zymaxid | Gatifloxacin | Inhibits DNA topoisomerases II and IV as well as DNA gyrase (DNA replication) | 1999 | |
| Group B (Injectable Antibiotics): | ||||
| Amikin | Amikacin | Aminoglycoside active against susceptible Gram-negative pathogens and Gram-positive bacteria | 1976 | |
| Capastat sulfate | Capreomycin * | Cyclic polypeptide antimicrobial | 1968 | |
| Kanamycin A | Kanamycin | Binds to the bacterial 30S ribosomal subunit-inhibits protein synthesis) | 1981 | |
| Group C (Oral drugs): | ||||
| Trecator/Prothionamide | Ethionamide */Prothionamide | Inhibits mycolic acid synthesis | 1965 | |
| Seromycin | Cycloserine * | Inhibits cell wall synthesis | 1964 | |
| Paser | para-Aminosalicylic acid | Inhibits folic acid synthesis, inhibits cell wall synthesis | 1994 | |
| Lamprene | Clofazimine | Inhibits mycobacterium growth and binds to mycobacterial DNA | 1986 | |
| Third-Line Defense (Treatment for MDR TB) | Sirturo | Bedaquiline * | Inhibits mycobacterial ATP synthase | 2012 |
| Zyvox | Linezolid | Inhibits bacterial reproduction of aerobic Gram-positive bacteria, some Gram-negative bacteria, and anaerobic bacteria; inhibits protein synthesis by binding to bacterial 23s ribosomal RNA of the 50s subunit | 2000 | |
| Natural Compound | Source | Chemical Constituent | Activity | References |
|---|---|---|---|---|
| Curcuma longa | Turmeric | Curcumin | Inhibits activation of NF-κB, and caspase-3; enhances T cell- mediated immunity and prevents post therapy susceptibility to reinfection/reactivation of Mtb. | [84,85,86] |
 | ||||
| Malus domestica | Apples (peels), apple leaves | Phloretin | Inhibits release of TNF-α, IL-1β and IL-12 and reduces phosphorylation of JNK, ERK and p38 MAPK. | [87] |
 | ||||
| Euphoria paralias | Whole plant extract | Quercetin | Inhibits glutamine synthetase and isocitrate lyase (important in nitrogen and TCA cycle, respectively). | [88,89,90,91,92] |
| Punica granatum | Pomegranate juice and peel | Quercetin | ||
 | ||||
| Glycine max | Soybeans | Genistein | Impairs B7-1 expression and nitric oxide release in MTSA-transfected macrophages. | [93] |
 | ||||
| Globularia alypum L. | Perennial flowering plant | Tannins | Activates phagocytic cells, proteasome inhibitor. | [94,95,96,97,98] |
| Camellia sinensis (L.) Kuntze | Green and black tea | Epigallocatechin gallate | Downregulates TACO gene transcription, inflicts structural damage to Mycobacterial cell wall, by inhibiting InhA. | [99,100,101,102] |
 | ||||
| Rheum rhaponticum | Rhubarb | Resveratrol extracts | Increases cell apoptosis. | [103,104] |
 | ||||
| Honeybee hives | Ethanolic extract of propolis | Decreases the size and number of Mtb colonies and increases sensitivity of mycobacteria to antibiotic therapy. | [103] | |
| Streptomyces lactacystinaeus | Microorganism | Lactacystin | Inhibits proteasome function. | [104] |
 | ||||
| Penicillium fellutanum | Marine fungus | Lipopeptide aldehyde (Fellutamide B) | Inhibits Mtb proteasome and human proteasome β5 function. | [105] |
 | ||||
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Maiolini, M.; Gause, S.; Taylor, J.; Steakin, T.; Shipp, G.; Lamichhane, P.; Deshmukh, B.; Shinde, V.; Bishayee, A.; Deshmukh, R.R. The War against Tuberculosis: A Review of Natural Compounds and Their Derivatives. Molecules 2020, 25, 3011. https://doi.org/10.3390/molecules25133011
Maiolini M, Gause S, Taylor J, Steakin T, Shipp G, Lamichhane P, Deshmukh B, Shinde V, Bishayee A, Deshmukh RR. The War against Tuberculosis: A Review of Natural Compounds and Their Derivatives. Molecules. 2020; 25(13):3011. https://doi.org/10.3390/molecules25133011
Chicago/Turabian StyleMaiolini, Morgan, Stacey Gause, Jerika Taylor, Tara Steakin, Ginger Shipp, Purushottam Lamichhane, Bhushan Deshmukh, Vaibhav Shinde, Anupam Bishayee, and Rahul R. Deshmukh. 2020. "The War against Tuberculosis: A Review of Natural Compounds and Their Derivatives" Molecules 25, no. 13: 3011. https://doi.org/10.3390/molecules25133011