
Evaluating the Potential of Ursolic Acid as Bioproduct for Cutaneous and Visceral Leishmaniasis

 , , ,
, , ,
Abstract
:
1. Introduction
2. Results
2.1. Assessment of Topical Formulations and Quantification of UA
2.2. In Vitro Activity against Leishmania spp. and Cytotoxicity against J774 Macrophages
2.3. In Vivo Activity against L. infantum
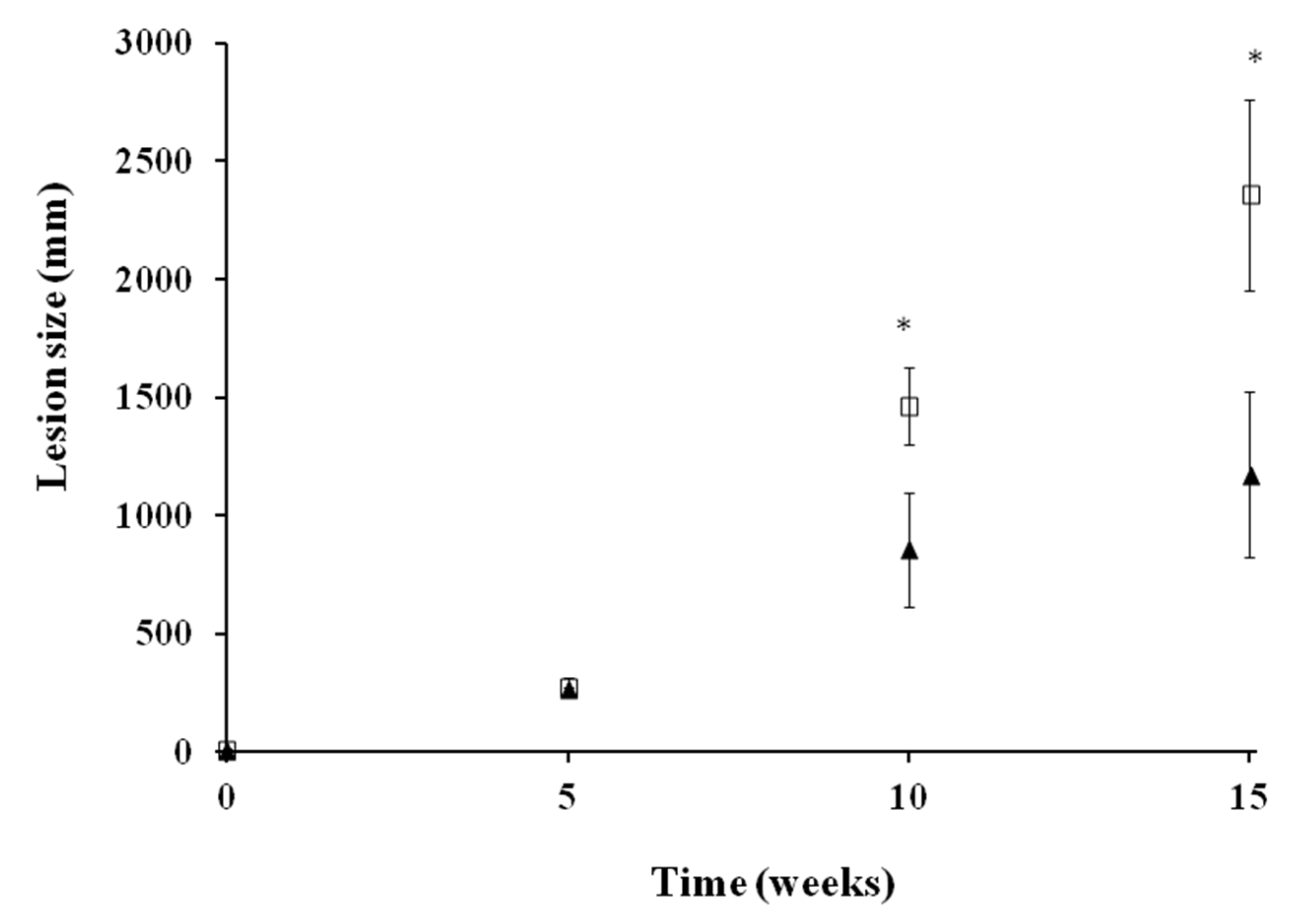
2.3.1. Efficacy in Acute Infection Model of VL in BALB/c
2.3.2. Efficacy in Chronic infection Hamster Model of VL
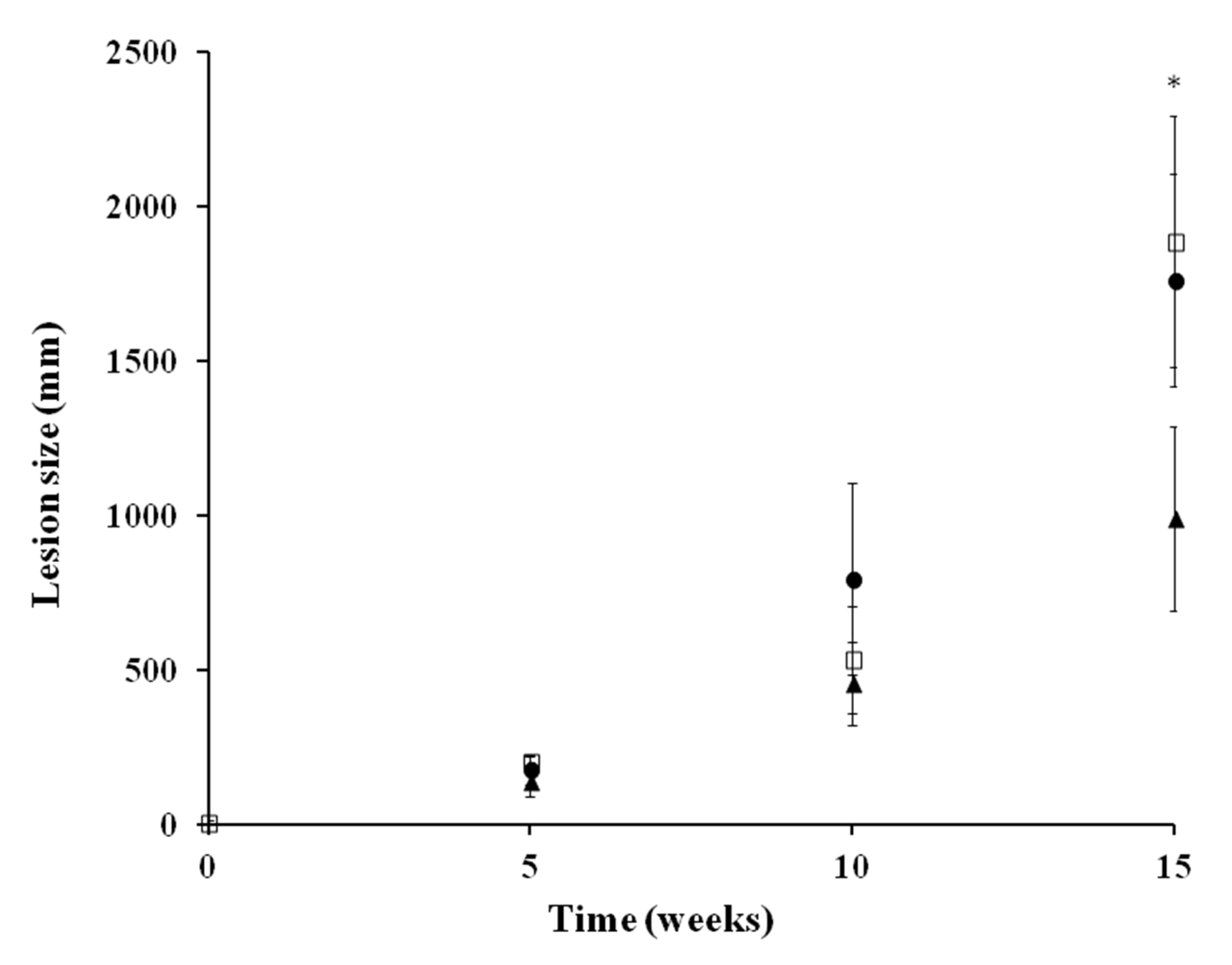
2.3.3. In Vivo Activity against L. amazonensis in an Experiment Model of CL
2.4. Production of Cytokines
3. Discussion
4. Materials and Methods
4.1. Chemical Reagents
4.2. Preparation of UA Topical Formulations
4.3. Quantification of UA in the Topical Formulations
4.4. Parasites and Culture Procedure
4.4.1. In Vitro Promastigote Susceptibility Assay
4.4.2. In Vitro Intracellular Amastigote Susceptibility Assay
4.4.3. Cytotoxicity Assay on Macrophages
4.5. In Vivo Studies
4.5.1. Acute Infection Model of VL in BALB/c
4.5.2. Chronic Infection Model of VL in Golden Hamster
4.5.3. Chronic Infection Model of CL in Golden Hamster
4.6. Effect of UA in the Cytokines Production in Macrophages Infected with Leishmania In Vitro
4.6.1. Splenocytes Culture and Evaluation of Cytokines Production
4.6.2. Analysis of Cytokines
4.7. Statistics
4.7.1. Statistical and Data Analysis for In Vitro Assays
4.7.2. Statistical and Data Analysis for in Vivo Assays
5. Conclusions
Author Contributions
Funding
Conflicts of Interest
References
- Alvar, J.; Vélez, I.D.; Bern, C.; Herrero, M.; Desjeux, P.; Cano, J.; Jannin, J.; den Boer, M.; WHO Leishmaniasis Control Team. Leishmaniasis worldwide and global estimates of its incidence. PLoS ONE 2012, 7, e35671. [Google Scholar] [CrossRef] [PubMed]
- WHO. Leishmaniasis. 2015. Available online: http://www.who.int/leishmaniasis/en/ (accessed on 15 May 2018).
- Yarbuh, A.L.; Premoli-de-Percoco, G.; Valera, M. Localized cutaneous leishmaniasis using polymerase chain reaction: A venezuelan family report. Parasitol. Dia 1997, 21, 3–4. [Google Scholar]
- Grimaldi, G., Jr.; Tesh, R.B. Leishmaniases of the New World: Current concepts and implications for future research. Clin. Microbiol. Rev. 1993, 6, 230–250. [Google Scholar] [CrossRef] [PubMed]
- Alvar, J.; Croft, S.; Olliaro, P. Chemotherapy in the treatment and control of leishmaniasis. Adv. Parasitol. 2006, 61, 223–274. [Google Scholar]
- Freitas-Junior, L.H.; Chatelainb, E.; Kima, H.A.; Siqueira-Neto, J. Visceral leishmaniasis treatment: What do we have, what do we need and how to deliver it? Int. J. Parasitol. Drugs Drug Resis. 2012, 2, 11–19. [Google Scholar] [CrossRef] [Green Version]
- Frézard, F.; Demicheli, C.; Ribeiro, R.R. Pentavalent antimonials: New perspectives for old drugs. Molecules 2009, 14, 2317–2336. [Google Scholar] [CrossRef] [Green Version]
- Roy, P.; Das, S.; Auddy, R.G.; Mukherjee, A. Biological targeting and drug delivery in control of Leishmaniasis. J. Cell Anim. Biol. 2012, 6, 73–87. [Google Scholar]
- Sundar, S.; Olliaro, P.L. Miltefosine in the treatment of leishmaniasis: Clinical evidence for informed clinical risk management. Ther. Clin. Risk Manag. 2007, 3, 733–740. [Google Scholar]
- Rocha, L.G.; Almeida, J.R.; Macêdo, R.O.; Barbosa-Filho, J.M. A review of natural products with antileishmanial activity. Phytomedicine 2005, 12, 514–535. [Google Scholar] [CrossRef]
- Jucá, M.M.; Cysne Filho, F.M.S.; de Almeida, J.C.; Mesquita, D.D.S.; Barriga, J.R.M.; Dias, K.C.F.; Barbosa, T.M.; Vasconcelos, L.C.; Leal, L.K.A.M.; Ribeiro, J.E.; et al. Flavonoids: Biological activities and therapeutic potential. Nat. Prod. Res. 2018, 16, 1–14. [Google Scholar] [CrossRef]
- Nazemiyeh, H.; Bahadori, F.; Delazar, A.; Ay, M.; Topcu, G.; Kolak, U.; Nahar, L.; Auzie, A.A.; Sarker, S.D. Tricetin 4′-O-α-L-rhamnopyranoside: A new flavonoid from the aerial parts of Erica arborea. Chem. Nat. Compd. 2008, 44, 174–177. [Google Scholar] [CrossRef]
- Kowalczyk, E.; Krzesiński, P.; Kura, M.; Szmigiel, B.; Błaszczyk, J. Anthocyanins in medicine. Pol. J. Pharmacol. 2003, 55, 699–702. [Google Scholar] [PubMed]
- Abebe, W. Herbal medication: Potential for adverse interactions with analgesic drugs. J. Clin. Pharm. Ther. 2002, 27, 391–401. [Google Scholar] [CrossRef] [PubMed]
- Bhattacharya, S.; Biswas, M.; Haldar, P.K. The triterpenoid fraction from Trichosanthes dioica root exhibits in vitro antileishmanial effect against Leishmania donovani promastigotes. Pharmacog. Res. 2013, 5, 109–112. [Google Scholar] [CrossRef] [Green Version]
- Szakiel, A.; Paczkowski, C.; Pensec, F.; Bertsch, C. Fruit cuticular waxes as a source of biologically active triterpenoids. Phytochem. Rev. 2012, 11, 263–284. [Google Scholar] [CrossRef] [Green Version]
- Checker, R.; Sandur, S.K.; Sharma, D.; Patwardhan, R.S.; Jayakumar, S.; Kohli, V.; Sethi, G.; Aggarwal, B.B.; Sainis, K.B. Potent anti-inflammatory activity of ursolic acid, a triterpenoid antioxidant, is mediated through suppression of NF-kappaB, AP-1 and NF-AT. PLoS ONE 2012, 7, e31318. [Google Scholar] [CrossRef] [Green Version]
- Torres-Santos, E.C.; Lopes, D.; Oliveira, R.R.; Carauta, J.P.; Falcao, C.A.; Kaplan, M.A.; Rossi-Bergmann, B. Antileishmanial activity of isolated triterpenoids from Pourouma guianensis. Phytomedicine 2004, 11, 114–120. [Google Scholar] [CrossRef]
- Innocente, A.M.; Silva, G.N.; Cruz, L.N.; Moraes, M.S.; Nakabashi, M.; Sonnet, P.; Gosmann, G.; Garcia, C.R.; Gnoatto, S.C. Synthesis and antiplasmodial activity of betulinic acid and ursolic acid analogues. Molecules 2012, 17, 12003–12014. [Google Scholar] [CrossRef] [Green Version]
- Yamamoto, E.S.; Campos, B.L.; Laurenti, M.D.; Lago, J.H.; Grecco Sdos, S.; Corbett, C.E.; Passero, L.F. Treatment with triterpenic fraction purified from Baccharis uncinella leaves inhibits Leishmania (Leishmania) amazonensis spreading and improves Th1 immune response in infected mice. Parasitol. Res. 2014, 113, 333–339. [Google Scholar] [CrossRef]
- da Silva Ferreira, D.; Esperandim, V.R.; Toldo, M.P.; Kuehn, C.C.; do Prado Junior, J.C.; Cunha, W.R.; e Silva, M.L.; de Albuquerque, S. In vivo activity of ursolic and oleanolic acids during the acute phase of Trypanosoma cruzi infection. Exp. Parasitol. 2013, 134, 455–459. [Google Scholar] [CrossRef]
- Liu, J. Oleanolic acid and ursolic acid: Research perspectives. J. Ethnopharmacol. 2005, 100, 92–94. [Google Scholar] [CrossRef] [PubMed]
- Martin-Cordero, C.; Reyes, M.; Ayuso, M.J.; Toro, M.V. Cytotoxic triterpenoids from Erica andevalensis. Z. Nat. C 2001, 56, 45–48. [Google Scholar] [CrossRef]
- Wang, X.H.; Zhou, S.Y.; Qian, Z.Z.; Zhang, H.L.; Qiu, L.H.; Song, Z.; Zhao, J.; Wang, P.; Hao, X.S.; Wang, H.Q. Evaluation of toxicity and single-dose pharmacokinetics of intravenous ursolic acid liposomes in healthy adult volunteers and patients with advanced solid tumors. Expert Opin. Drug Metab. Toxicol. 2013, 9, 117–125. [Google Scholar] [CrossRef] [PubMed]
- Zhu, Z.; Qian, Z.; Yan, Z.; Zhao, C.; Wang, H.; Ying, G. A phase I pharmacokinetic study of ursolic acid nanoliposomes in healthy volunteers and patients with advanced solid tumors. Int. J. Nanomed. 2013, 8, 129–136. [Google Scholar]
- do Nascimento, P.G.; Lemos, T.L.; Bizerra, A.M.; Arriaga, A.M.; Ferreira, D.A.; Santiago, G.M.; Braz-Filho, R.; Costa, J.G. Antibacterial and antioxidant activities of ursolic acid and derivatives. Molecules 2013, 19, 1317–1327. [Google Scholar] [CrossRef]
- Alqahtani, A.; Hamid, K.; Kam, A.; Wong, K.H.; Abdelhak, Z.; Razmovski-Naumovski, V.; Chan, K.; Li, K.M.; Groundwater, P.W.; Li, G.Q. The pentacyclic triterpenoids in herbal medicines and their pharmacological activities in diabetes and diabetic complications. Curr. Med. Chem. 2013, 20, 908–931. [Google Scholar]
- Kashiwada, Y.; Nagao, T.; Hashimoto, A.; Ikeshiro, Y.; Okabe, H.; Cosentino, L.M.; Cosentino, L.M.; Lee, K.H. Anti-AIDS agents 38. Anti-HIV activity of 3-O-acyl ursolic acid derivatives. J. Nat. Prod. 2000, 63, 1619–1622. [Google Scholar] [CrossRef]
- Horiuchi, K.; Shiota, S.; Hatano, T.; Yoshida, T.; Kuroda, T.; Tsuchiya, T. Antimicrobial activity of oleanolic acid from Salvia officinalis and related compounds on vancomycin-resistant enterococci (VRE). Biol. Pharm. Bull. 2007, 30, 1147–1149. [Google Scholar] [CrossRef] [Green Version]
- Gnoatto, S.C.; Dalla Vechia, L.; Lencina, C.L.; Dassonville-Klimpt, A.; Da Nascimento, S.; Mossalayi, D.; Guillon, J.; Gosmann, G.; Sonnet, P. Synthesis and preliminary evaluation of new ursolic and oleanolic acids derivatives as antileishmanial agents. J. Enzyme Inhib. Med. Chem. 2008, 23, 604–610. [Google Scholar] [CrossRef] [Green Version]
- Kwon, T.H.; Lee, B.; Chung, S.H.; Kim, D.H.; Lee, Y.S. Synthesis and NO Production inhibitory activities of ursolic acid and oleanolic acid derivatives. Bull. Korean Chem. Soc. 2009, 30, 119–123. [Google Scholar] [CrossRef]
- Zerin, T.; Lee, M.; Jang, W.S.; Nam, K.W.; Song, H.Y. Anti-inflammatory potential of ursolic acid in Mycobacterium tuberculosis-sensitized and concanavalin A-stimulated cells. Mol. Med. Rep. 2016, 13, 2736–2744. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lopez-Garcia, S.; Castaneda-Sanchez, J.I.; Jimenez-Arellanes, A.; Dominguez-Lopez, L.; Castro-Mussot, M.E.; Hernandez-Sanchez, J.; Luna-Herrera, J. Macrophage Activation by Ursolic and Oleanolic Acids during Mycobacterial Infection. Molecules 2015, 20, 14348–14364. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Choi, W.H.; Lee, I.A. The Mechanism of Action of Ursolic Acid as a Potential Anti-Toxoplasmosis Agent, and Its Immunomodulatory Effects. Pathogens 2019, 8, 61. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Passero, L.F.; Bonfim-Melo, A.; Corbett, C.E.; Laurenti, M.D.; Toyama, M.H.; de Toyama, D.O.; Romoff, P.; Fávero, O.A.; dos Grecco, S.S.; Zalewsky, C.A.; et al. Anti-leishmanial effects of purified compounds from aerial parts of Baccharis uncinella C. DC. (Asteraceae). Parasitol. Res. 2011, 108, 529–536. [Google Scholar] [CrossRef] [PubMed]
- Jesus, J.A.; Fragoso, T.N.; Yamamoto, E.S.; Laurenti, M.D.; Silva, M.S.; Ferreira, A.F.; Lago, J.H.; Santos-Gomes, G.; Passero, L.F. Therapeutic effect of ursolic acid in experimental visceral leishmaniasis. Int. J. Parasitol. Drugs Drug Resist. 2017, 7, 1–11. [Google Scholar] [CrossRef] [PubMed]
- Yamamoto, E.S.; Campos, B.L.; Jesus, J.A.; Laurenti, M.D.; Ribeiro, S.P.; Kallas, E.G.; Rafael-Fernandes, M.; Santos-Gomes, G.; Silva, M.S.; Sessa, D.P.; et al. The Effect of Ursolic Acid on Leishmania (Leishmania) amazonensis is Related to Programed Cell Death and Presents Therapeutic Potential in Experimental Cutaneous Leishmaniasis. PLoS ONE 2015, 10, e0144946. [Google Scholar] [CrossRef] [Green Version]
- Tan, N.; Kaloga, M.; Radtke, O.A.; Kiderlen, A.F.; Oksuz, S.; Ulubelen, A.; Kolodziej, H. Abietane diterpenoids and triterpenoic acids from Salvia cilicica and their antileishmanial activities. Phytochemistry 2002, 61, 881–884. [Google Scholar] [CrossRef]
- Moulisha, B.; Kumar, G.A.; Kanti, H.P. Anti-leishmanial and anti-cancer activities of a pentacyclic triterpenoid isolated from the leaves of Terminalia arjuna Combretaceae. Trop. J. Pharm. Res. 2010, 9, 135–140. [Google Scholar] [CrossRef]
- Odonne, G.; Herbette, G.; Eparvier, V.; Bourdy, G.; Rojas, R.; Sauvain, M.; Stien, D. Antileishmanial sesquiterpene lactones from Pseudelephantopus spicatus, a traditional remedy from the Chayahuita Amerindians (Peru). Part III. J. Ethnopharmacol. 2011, 137, 875–879. [Google Scholar] [CrossRef]
- Peixoto, J.A.; Andrade, E.S.M.L.; Crotti, A.E.; Cassio Sola Veneziani, R.; Gimenez, V.M.; Januario, A.H.; Groppo, M.; Magalhães, L.G.; Dos Santos, F.F.; Albuquerque, S.; et al. Antileishmanial activity of the hydroalcoholic extract of Miconia langsdorffii, isolated compounds, and semi-synthetic derivatives. Molecules 2011, 16, 1825–1833. [Google Scholar] [CrossRef] [Green Version]
- You, H.J.; Choi, C.Y.; Kim, J.Y.; Park, S.J.; Hahm, K.S.; Jeong, H.G. Ursolic acid enhances nitric oxide and tumor necrosis factor-alpha production via nuclear factor-kappa B activation in the resting macrophages. FEBS Lett. 2001, 509, 156–160. [Google Scholar] [CrossRef] [Green Version]
- Lafuse, W.P.; Story, R.; Mahylis, J.; Gupta, G.; Varikuti, S.; Steinkamp, H.; Oghumu, S.; Satoskar, A.R. Leishmania donovani infection induces anemia in hamsters by differentially altering erythropoiesis in bone marrow and spleen. PLoS ONE 2013, 8, e59509. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ference, J.D.; Last, A.R. Choosing topical corticosteroids. Am. Fam. Physician 2009, 79, 135–140. [Google Scholar] [PubMed]
- Wong, T.W.; Chiu, H.C.; Chang, C.H.; Lin, L.J.; Liu, C.C.; Chen, J.S. Silicone cream occlusive dressing--a novel noninvasive regimen in the treatment of keloid. Dermatology 1996, 192, 329–333. [Google Scholar] [CrossRef] [PubMed]
- Jesus, J.A.; Lago, J.H.; Laurenti, M.D.; Yamamoto, E.S.; Passero, L.F. Antimicrobial activity of oleanolic and ursolic acids: An update. Evid. Based Complement. Alternat. Med. 2015, 2015, 620472. [Google Scholar] [CrossRef]
- Ho, J.L.; Reed, S.G.; Wick, E.A.; Giordano, M. Granulocyte-macrophage and macrophage colony-stimulating factors activate intramacrophage killing of Leishmania mexicana amazonensis. J. Infect. Dis. 1990, 162, 224–230. [Google Scholar]
- Almeida, R.P.; Brito, J.; Machado, P.L.; de Jesus, A.R.; Schriefer, A.; Guimarães, L.H.; Carvalho, E.M. Successful treatment of refractory cutaneous leishmaniasis with GM-CSF and antimonials. Am. J. Trop. Med. Hyg. 2005, 73, 79–81. [Google Scholar] [CrossRef]
- Miltefosine and GM-CSF in Cutaneous Leishmaniasis. Available online: https://clinicaltrials.gov/ct2/show/NCT03023111 (accessed on 15 April 2019).
- Bilbao-Ramos, P.; Galiana-Rosello, C.; Dea-Ayuela, M.A.; Gonzalez-Alvarez, M.; Vega, C.; Rolon, M.; Pérez-Serrano, J.; Bolás-Fernández, F.; González-Rosende, M.E. Nuclease activity and ultrastructural effects of new sulfonamides with anti-leishmanial and trypanocidal activities. Parasitol. Int. 2012, 61, 604–613. [Google Scholar] [CrossRef]
- Bilbao-Ramos, P.; Sifontes-Rodriguez, S.; Dea-Ayuela, M.A.; Bolas-Fernandez, F. A fluorometric method for evaluation of pharmacological activity against intracellular Leishmania amastigotes. J. Microbiol. Methods 2012, 89, 8–11. [Google Scholar] [CrossRef]
- Galiana-Rosello, C.; Bilbao-Ramos, P.; Dea-Ayuela, M.A.; Rolon, M.; Vega, C.; Bolas-Fernandez, F.; García-España, E.; Alfonso, J.; Coronel, C.; González-Rosende, M.E. In vitro and in vivo antileishmanial and trypanocidal studies of new N-benzene- and N-naphthalenesulfonamide derivatives. J. Med. Chem. 2013, 56, 8984–8998. [Google Scholar] [CrossRef]
- Dea-Ayuela, M.A.; Castillo, E.; Gonzalez-Alvarez, M.; Vega, C.; Rolon, M.; Bolas-Fernandez, F.; Borrás, J.; González-Rosende, M.E. In vivo and in vitro anti-leishmanial activities of 4-nitro-N-pyrimidin- and N-pyrazin-2-ylbenzenesulfonamides, and N2-(4-nitrophenyl)-N1-propylglycinamide. Bioorg. Med. Chem. 2009, 17, 7449–7456. [Google Scholar] [CrossRef] [PubMed]
- Titus, R.G.; Marchand, M.; Boon, T.; Louis, J.A. A limiting dilution assay for quantifying Leishmania major in tissues of infected mice. Parasite Immunol. 1985, 7, 545–555. [Google Scholar] [CrossRef] [PubMed]
- Dea-Ayuela, M.A.; Rama-Iniguez, S.; Alunda, J.M.; Bolas-Fernandez, F. Setting new immunobiological parameters in the hamster model of visceral leishmaniasis for in vivo testing of antileishmanial compounds. Vet. Res. Commun. 2007, 31, 703–717. [Google Scholar] [CrossRef] [PubMed]
- Morais-Teixeira, E.D.; Carvalho, A.S.; Costa, J.C.; Duarte, S.L.; Mendonça, J.S.; Boechat, N.; Rabello, A. In vitro and in vivo activity of meglumine antimoniate produced at Farmanguinhos-Fiocruz, Brazil, against Leishmania (Leishmania) amazonensis, L (L.) chagasi and L (Viannia) braziliensis. Mem. Inst. Oswaldo Cruz 2008, 103, 358–362. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Perez-Cantero, A.; Serrano, D.R.; Navarro-Rodriguez, P.; Schatzlein, A.G.; Uchegbu, I.F.; Torrado, J.J.; Capilla, J. Increased Efficacy of Oral Fixed-Dose Combination of Amphotericin B and AHCC((R)) Natural Adjuvant against Aspergillosis. Pharmaceutics 2019, 11, 456. [Google Scholar] [CrossRef] [PubMed] [Green Version]
Sample Availability: Samples of the compounds are available from the authors. |



{kind=link}
{kind=link}
{kind=link}
| Compound | IC50 (μg/mL) a | CC50 (μg/mL) c | |||||||
|---|---|---|---|---|---|---|---|---|---|
| Leishmania brazilensis | SI b | Leishmania guyanensis | SI b | Leishmania amazonensis | SI b | Leishmania infantum | SI b | J774 macrophages | |
| UA | 17.0 ± 0.92 | 3.2 | 28.0 ± 0.09 | 2 | 14.1 ± 0.14 | 4 | 20.9 ± 1.40 | 2.7 | 55.8 ± 3.98 |
| Miltefosine | 7.16 ± 0.22 | 7.9 | 6.85 ± 0.35 | 8.2 | 12.44 ± 0.49 | 4.6 | 7.19 ± 0.60 | 7.9 | 57.1 ± 4.6 |
| Compound | IC50 (μg/mL) a | |||
|---|---|---|---|---|
| Leishmania amazonensis | SI b | Leishmania infantum | SI b | |
| UA | 2.24 ± 0.16 | 24.9 | 6.7 ± 0.6 | 8.3 |
| Miltefosine | 20.09 ± 1.47 | 2.8 | 23.7 ± 1.78 | 2.5 |
| Group | Parasite Burden (Amastigotes/mg Organ (× 103) | |
|---|---|---|
| Spleen | Liver | |
| Treated group (with UA) | 0.18 ± 0.02 * | 0.06 ± 0.01 * |
| Untreated group (control) | 111.84 ± 2.5 | 27.96 ± 7.8 |
| Group | Parasite Burden (Amastigotes/mg Organ (×106) | |
|---|---|---|
| Spleen | Liver | |
| Treated group (with UA) | 9.75 ± 4.73 | 4.0 ± 2.88+* |
| Untreated group (control) | 16.78 ± 5.30 | 19.04 ± 7.60 |
| Cytokines (pg/mL) | Splenocytes Mice BALBc | ||
|---|---|---|---|
| Untreated Group | Ursolic Acid (2.24 µg/mL) | Ursolic Acid (6.7 µg/mL) | |
| GM-CSF | 11.6 ± (3) | 60.0 ± (10) * | 100.7 ± (20) * |
| IFN-γ | 0 | 44.4 ± (4.2) * | 223.4 ± (87.7) ** |
| IL-4 | 3.5 ± (0.9) | 9.3 ± (1.4) * | 6.4 ± (1.1) |
| IL-6 | 42.8 ± (5.6) | 103.1 ± (11) * | 50.2 ± (7.9) |
| IL-9 | 153.1 ± (20.5) | 216.3 ± (9.6) | 232.0 ± (25.5) * |
| IL-10 | 72.2 ± (17.6) | 234.1 ± (36.2) * | 135.8 ± (33.3) * |
| RANTES | 16.2 ± (0.9) | 1.6 ± (0.3) ** | 0 ** |
| Cytokines (pg/mL) | L. infantum/macrophages | L. amazonensis/macrophages | ||
|---|---|---|---|---|
| Untreated Mean (ESM) | Ursolic Acid Mean (ESM) | Untreated Mean (ESM) | Ursolic Acid Mean (ESM) | |
| GM-CSF | 23.2 ± (4.4) | 67.3 ± (2.8) * | 49.8 ± (6.9) | 44.8 ± (0.9) |
| IFN-γ | 90.2 ± (125.9) | 86.9 ± (9.8) | 139.2 ± (20.2) | 43.5 ± (26.8) * |
| IL-1b | 23.4 ± (8.5) | 24.5 ± (4.9) | 42.3 ± (5.8) | 20 ± (1.3) * |
| IL-2 | 0 | 3.9 ± (1.1) * | 3 ± (0.4) | 1.4 ± (0.4) * |
| IL-6 | 53.4 ± (11.4) | 97.8 ± (7.2) * | 71 ± (9.5) | 66.8 ± (8.5) |
| TNF-α | 0 | 22.72 ± (3.9)* | 0 | 40.22 ± (6.9)* |
| IL-10 | 146.4 ± (10.8) | 258.9 ± (14.6) * | 197.1 ± (7.9) | 184.1 ± (37.6) |
| MCP-1 | 33 ± (4.0) | 50.3 ± (16.4) | 147.9 ± (19.5) | 37.1 ± (2.2) * |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Bilbao-Ramos, P.; Serrano, D.R.; Ruiz Saldaña, H.K.; Torrado, J.J.; Bolás-Fernández, F.; Dea-Ayuela, M.A. Evaluating the Potential of Ursolic Acid as Bioproduct for Cutaneous and Visceral Leishmaniasis. Molecules 2020, 25, 1394. https://doi.org/10.3390/molecules25061394
Bilbao-Ramos P, Serrano DR, Ruiz Saldaña HK, Torrado JJ, Bolás-Fernández F, Dea-Ayuela MA. Evaluating the Potential of Ursolic Acid as Bioproduct for Cutaneous and Visceral Leishmaniasis. Molecules. 2020; 25(6):1394. https://doi.org/10.3390/molecules25061394
Chicago/Turabian StyleBilbao-Ramos, Pablo, Dolores R. Serrano, Helga Karina Ruiz Saldaña, Juan J. Torrado, Francisco Bolás-Fernández, and María Auxiliadora Dea-Ayuela. 2020. "Evaluating the Potential of Ursolic Acid as Bioproduct for Cutaneous and Visceral Leishmaniasis" Molecules 25, no. 6: 1394. https://doi.org/10.3390/molecules25061394