Treatment with Subcritical Water-Hydrolyzed Citrus Pectin Ameliorated Cyclophosphamide-Induced Immunosuppression and Modulated Gut Microbiota Composition in ICR Mice

Abstract
:1. Introduction
2. Results
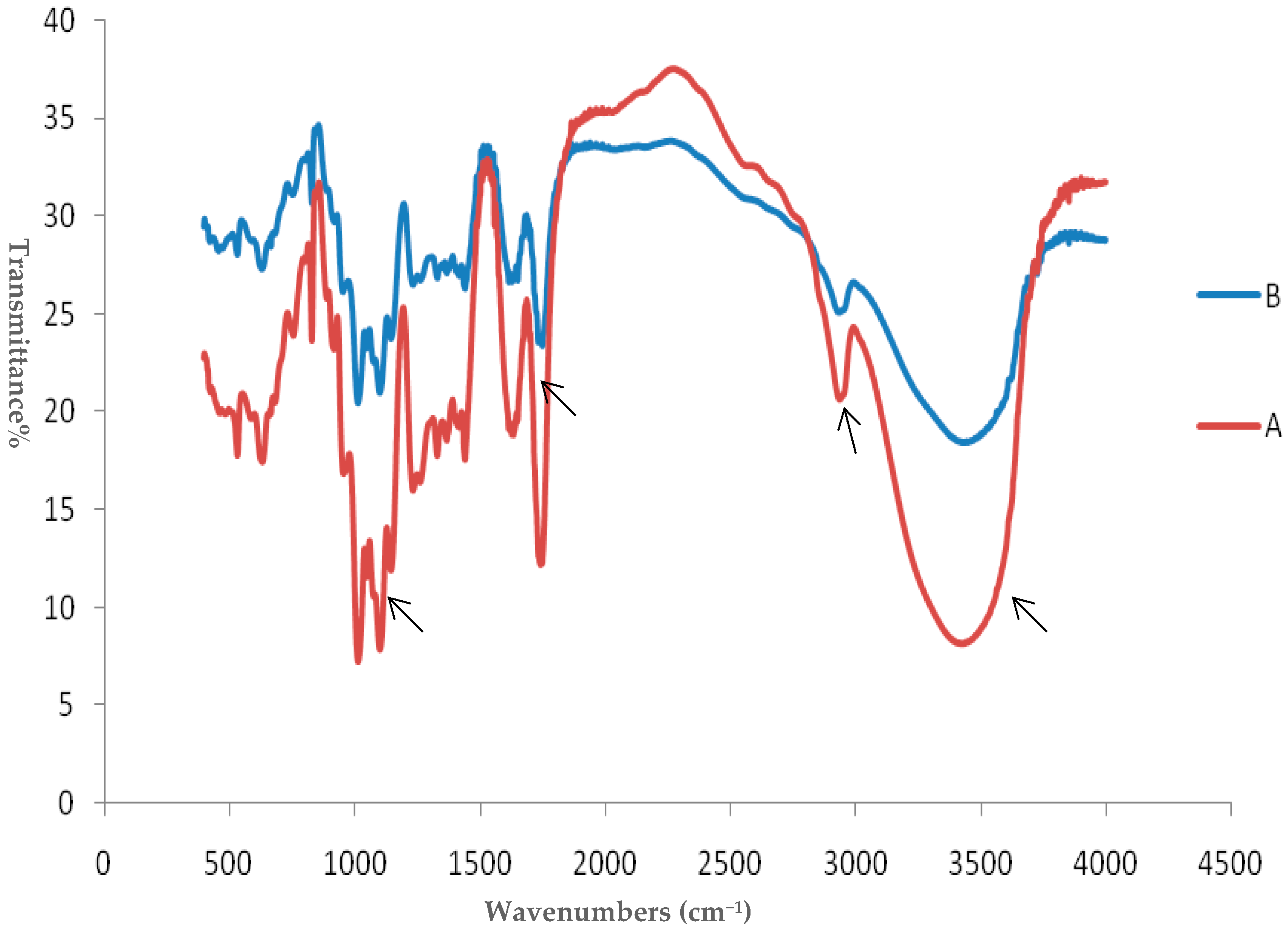
2.1. Preliminary Characterization of SCP
2.2. Effects of SCP on Body Weight and Immune Organ Indices
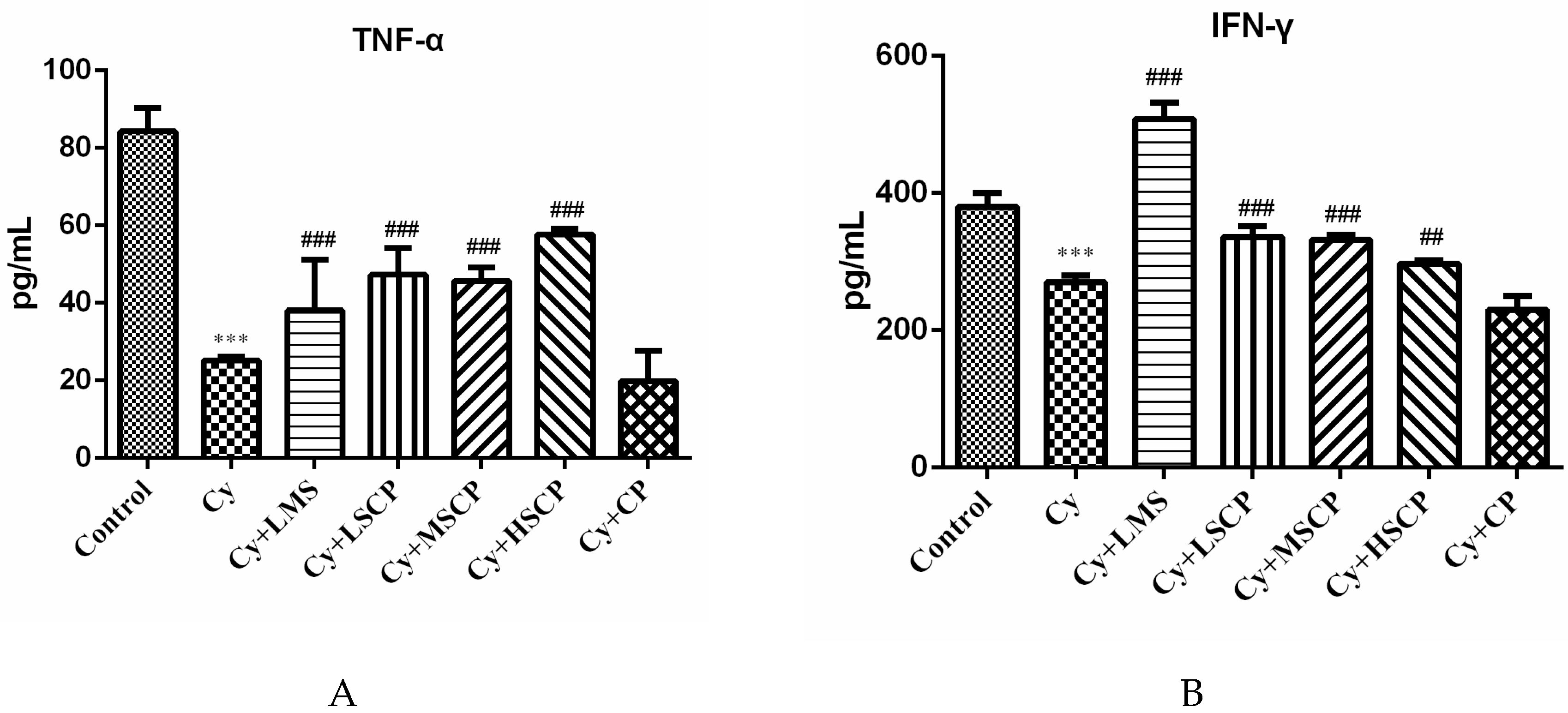
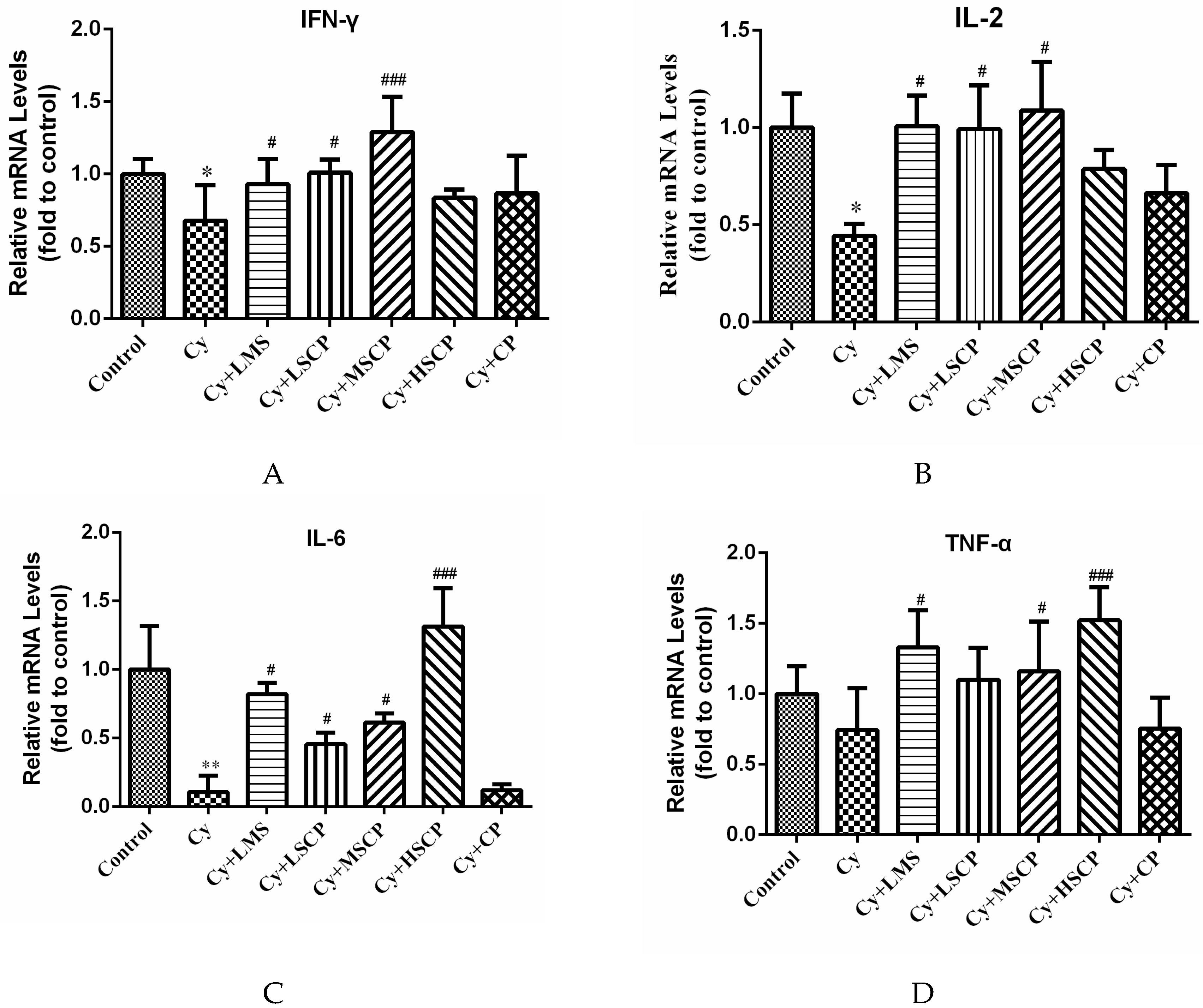
2.3. Effects of SCP on the Expression of IFN-γ, IL-2, IL-6, and TNF-α in Mice
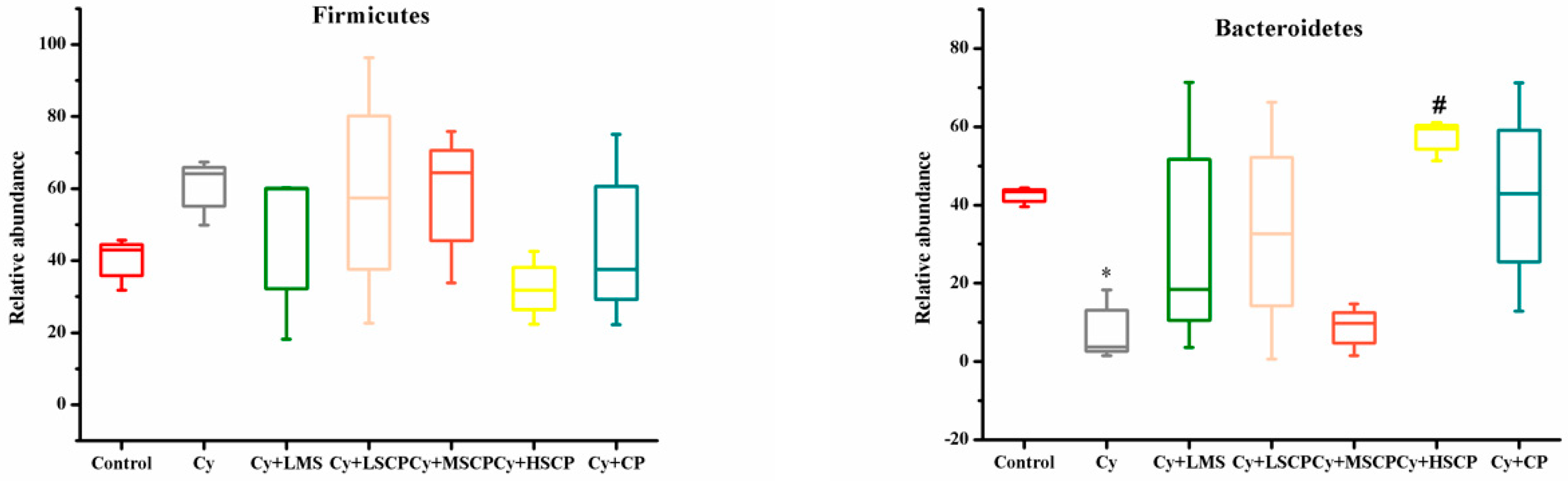
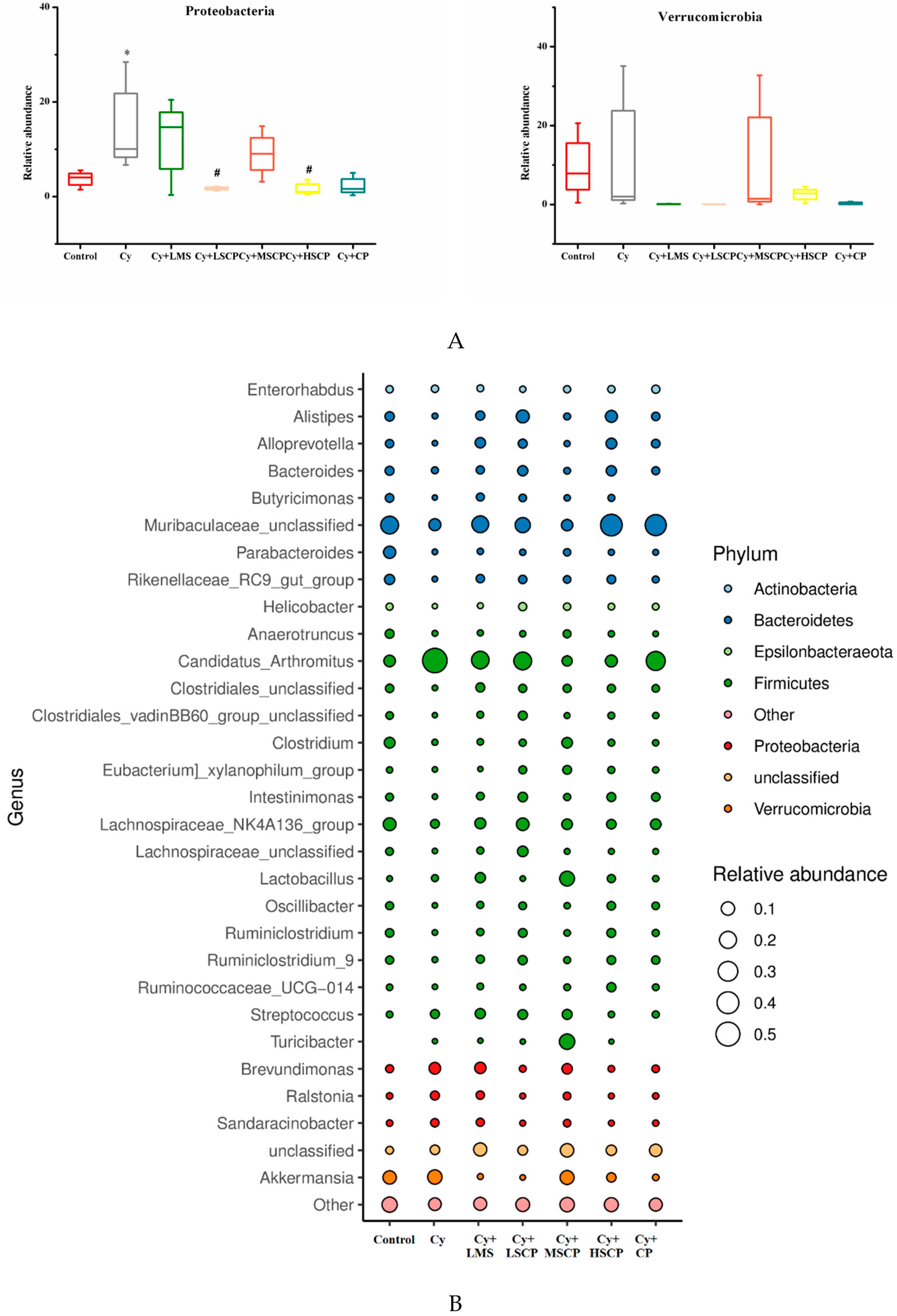
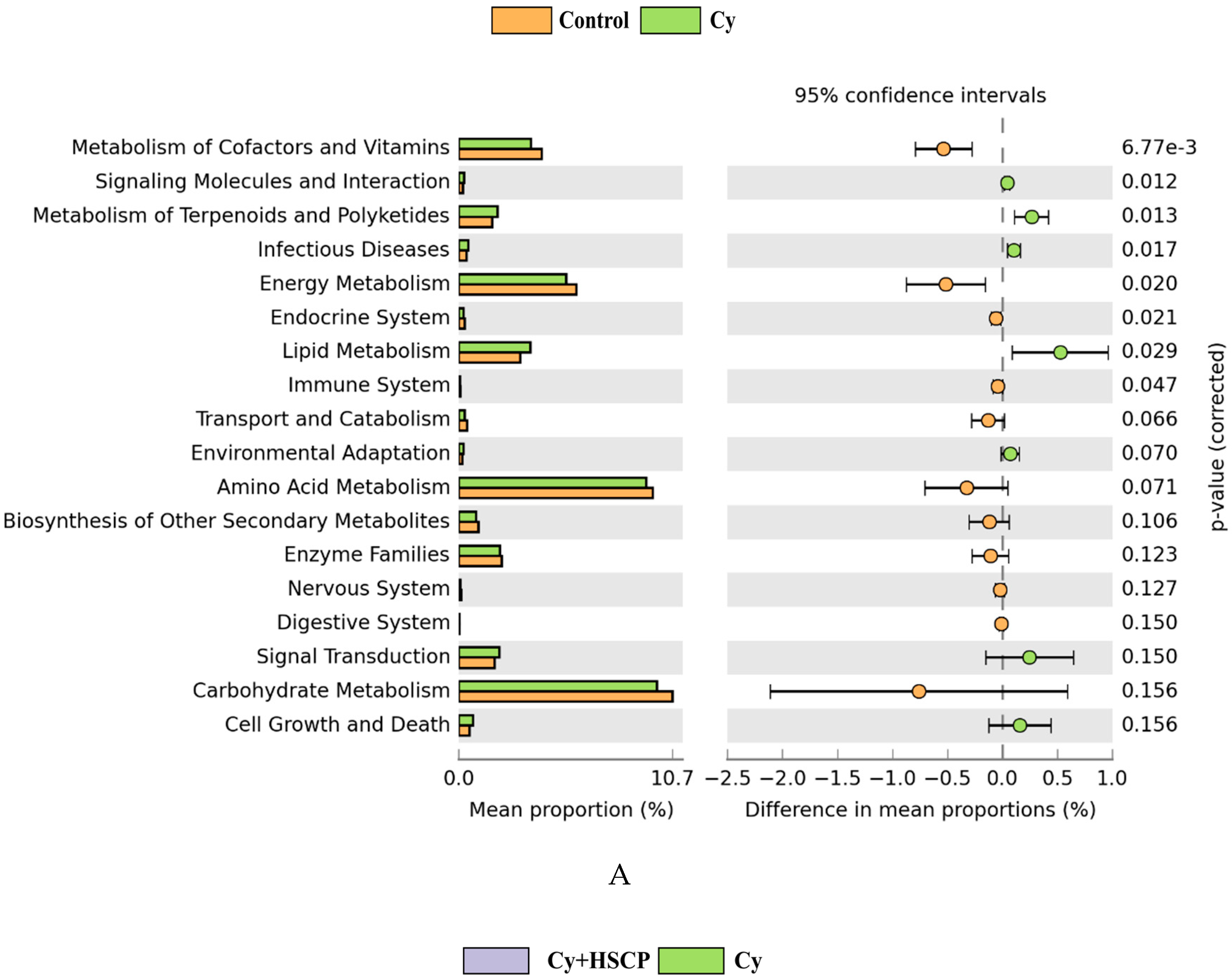
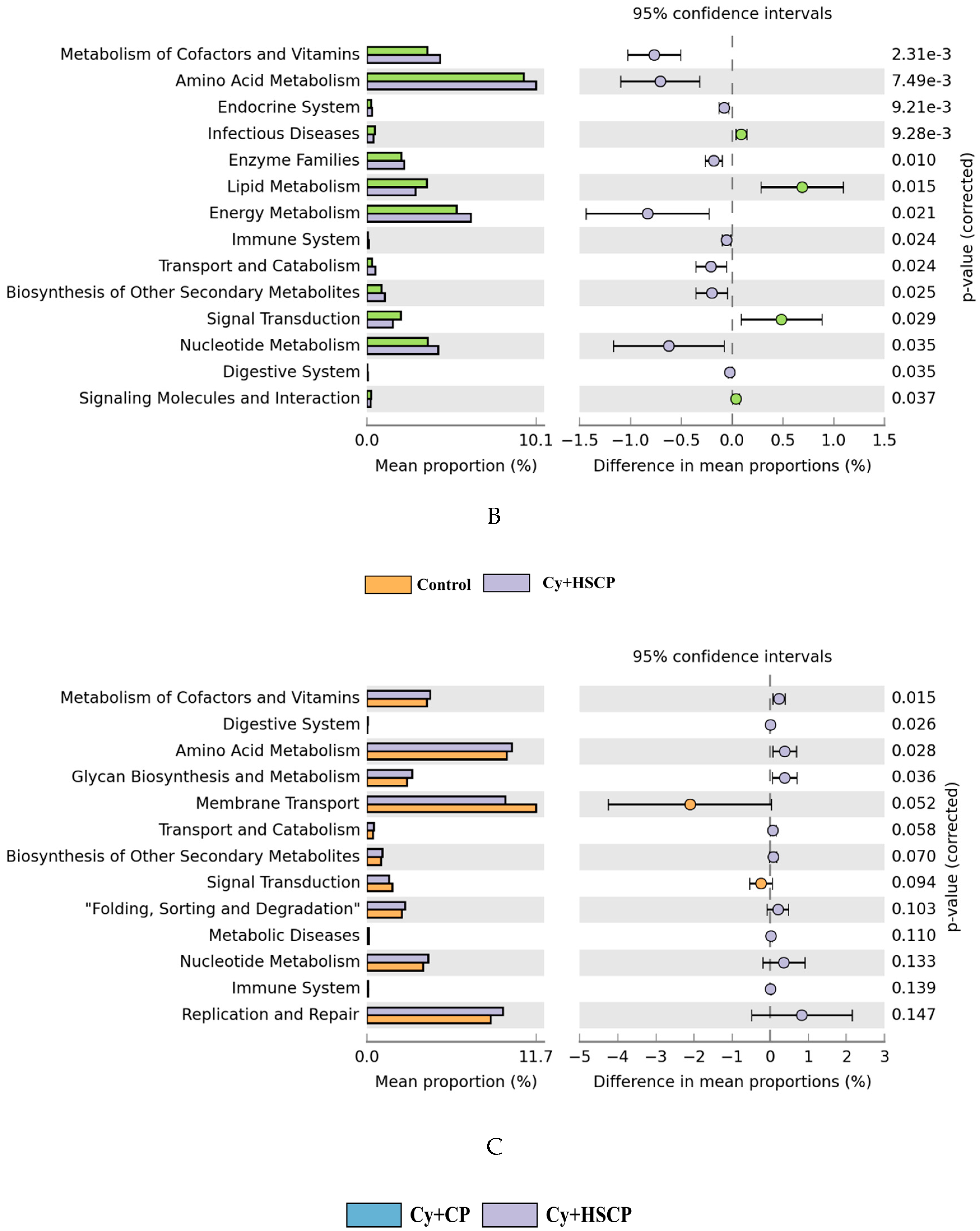
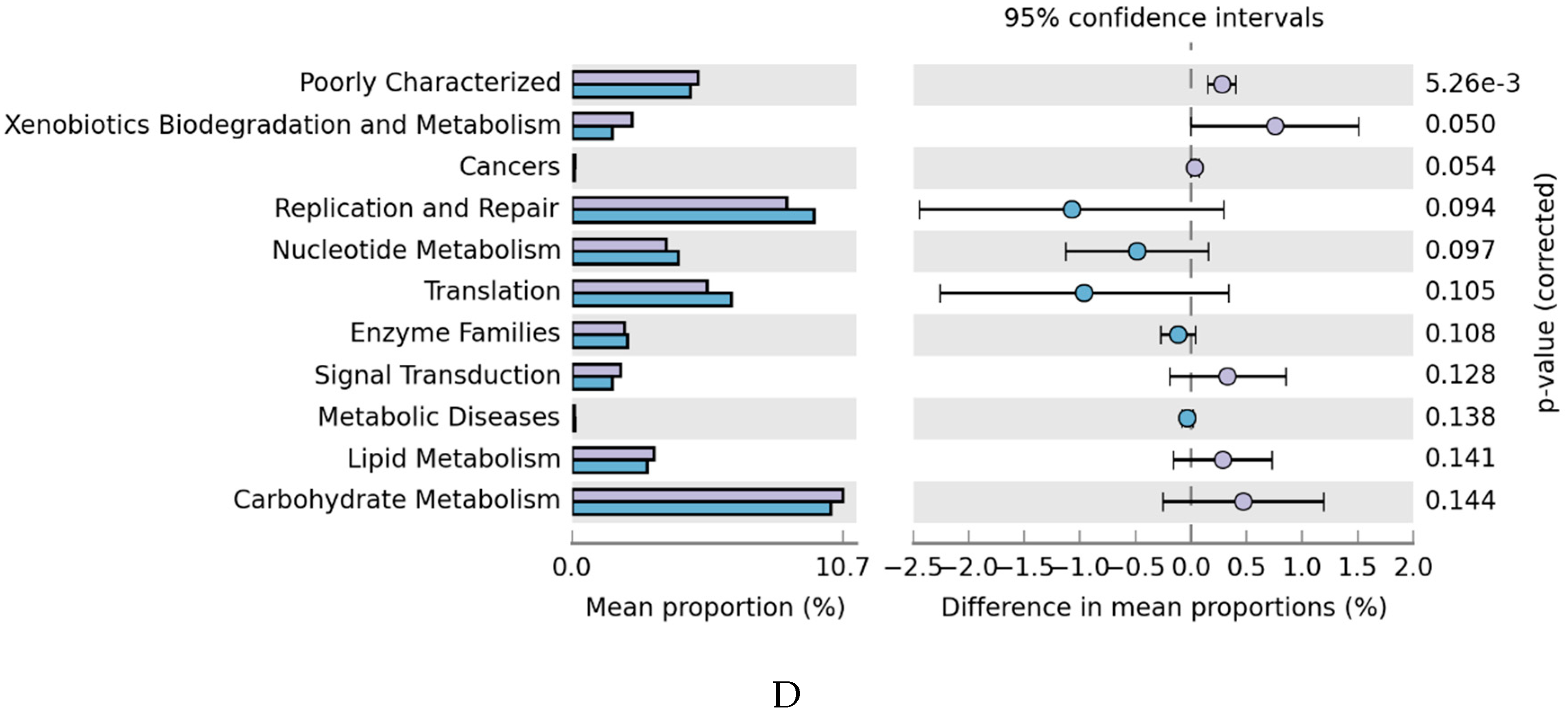
2.4. Effects of SCP on the Gut Microbiota
3. Discussion
4. Materials and Methods
4.1. Preparation of Subcritical Water-Hydrolyzed Citrus Pectin
4.2. The Properties of Subcritical Water-Hydrolyzed Citrus Pectin
4.3. Animals and Experimental Design
4.4. Analysis of Immune Organ Indices
4.5. Quantification of Cytokines by ELISA
4.6. Quantitative Real-Time PCR
4.7. Fecal DNA Extraction and Pyrosequencing
4.8. Statistical Analysis
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Conflicts of Interest
References
- Zhu, G.; Jiang, Y.; Yao, Y.; Wu, N.; Luo, J.; Hu, M.; Tu, Y.; Xu, M. Ovotransferrin ameliorates the dysbiosis of immunomodulatory function and intestinal microbiota induced by cyclophosphamide. Food Funct. 2019, 10, 1109–1122. [Google Scholar] [CrossRef] [PubMed]
- Horvat, T.Z.; Adel, N.G.; Dang, T.O.; Momtaz, P.; Postow, M.A.; Callahan, M.K.; Carvajal, R.D.; Dickson, M.A.; D’Angelo, S.P.; Woo, K.M.; et al. Immune-related adverse events, need for systemic immunosuppression, and effects on survival and time to treatment failure in patients with melanoma treated with ipilimumab at memorial sloankettering cancer center. J. Clin.Oncol. 2015, 33, 3193–3198. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Xu, X.; Zhang, X. Effects of cyclophosphamide on immune system and gut microbiota in mice. Microbiol. Res. 2015, 171, 97–106. [Google Scholar] [CrossRef] [PubMed]
- Iida, N.; Dzutsev, A.; Stewart, C.A.; Smith, L.; Bouladoux, N.; Weingarten, R.A.; Molina, D.A.; Salcedo, R.; Back, T.; Cramer, S.; et al. Commensal bacteria control cancer response to therapy by modulating the tumor microenvironment. Science. 2013, 342, 967–970. [Google Scholar] [CrossRef]
- Ding, Y.; Yan, Y.; Chen, D.; Ran, L.; Mi, J.; Lu, L.; Jing, B.; Li, X.; Zeng, X.; Cao, Y. Modulating effects of polysaccharides from the fruits of Lyciumbarbarum on the immune response and gut microbiota in cyclophosphamide-treated mice. Food Funct. 2019, 10, 3671–3683. [Google Scholar] [CrossRef]
- LIU, W.; YAN, R.; ZHANG, L. Dendrobiumsonia polysaccharide regulates immunity and restores the dysbiosis of the gut microbiota of the cyclophosphamide-induced immunosuppressed mice. Chin. J. Nat. Medicines. 2019, 17, 600–607. [Google Scholar] [CrossRef]
- Chu, Q.; Zhang, Y.; Chen, W.; Jia, R.; Yu, X.; Wang, Y.; Li, Y.; Liu, Y.; Ye, X.; Yu, L.; et al. ApiosamericanaMedik flowers polysaccharide (AFP) alleviate Cyclophosphamide-induced immunosuppression in ICR mice. Int. J. Biol.Macromol. 2020, 144, 829–836. [Google Scholar] [CrossRef]
- Cao, J.; Tang, D.; Wang, Y.; Li, X.; Hong, L.; Sun, C. Characteristics and immune-enhancing activity of pectic polysaccharides from sweet cherry (Prunus avium). Food Chem. 2018, 254, 47–54. [Google Scholar] [CrossRef]
- Koh, A.; De Vadder, F.; Kovatcheva-Datchary, P.; Backhed, F. From dietary fiber to host physiology: Short-chain fatty acids as key bacterial metabolites. Cell 2016, 165, 1332–1345. [Google Scholar] [CrossRef] [Green Version]
- Maxwell, E.G.; Belshaw, N.J.; Waldron, K.W.; Morris, V.J. Pectin-an emerging new bioactive food polysaccharide. Trends Food Sci. Tech. 2012, 24, 64–73. [Google Scholar] [CrossRef]
- Wang, W.; Chen, W.; Zou, M.; Lv, R.; Wang, D.; Hou, F.; Feng, H.; Ma, X.; Zhong, J.; Ding, T.; et al. Applications of power ultrasound in oriented modification and degradation of pectin: A review. J. Food Eng. 2018, 234, 98–107. [Google Scholar] [CrossRef]
- Ma, X.; Wang, D.; Chen, W.; Ismail, B.B.; Wang, W.; Lv, R.; Ding, T.; Ye, X.; Liu, D. Effects of ultrasound pretreatment on the enzymolysis of pectin: Kinetic study, structural characteristics and anti-cancer activity of the hydrolysates. Food Hydrocolloid. 2018, 79, 90–99. [Google Scholar] [CrossRef]
- Chen, X.; Qi, Y.; Zhu, C.; Wang, Q. Effect of ultrasound on the properties and antioxidant activity of hawthorn pectin. Int. J. Biol. Macromol. 2019, 131, 273–281. [Google Scholar] [CrossRef] [PubMed]
- Muñoz-Almagro, N.; Montilla, A.; Moreno, F.J.; Villamiel, M. Modification of citrus and apple pectin by power ultrasound: Effects of acid and enzymatic treatment. Ultrason.Sonochem. 2017, 38, 807–819. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Klinchongkon, K.; Khuwijitjaru, P.; Adachi, S. Properties of subcritical water-hydrolyzed passion fruit (Passifloraedulis) pectin. Food Hydrocolloid. 2018, 74, 72–77. [Google Scholar] [CrossRef]
- Klinchongkon, K.; Khuwijitjaru, P.; Adachi, S. Degradation kinetics of passion fruit pectin in subcritical water. Biosci. Biotechnol. Biochem. 2017, 81, 712–717. [Google Scholar] [CrossRef] [Green Version]
- Chen, D.; Chen, G.; Ding, Y.; Wan, P.; Peng, Y.; Chen, C.; Ye, H.; Zeng, X.; Ran, L. Polysaccharides from the flowers of tea (Camellia sinensis L.) modulate gut health and ameliorate cyclophosphamide-induced immunosuppression. J. Funct. Foods 2019, 61, 103470. [Google Scholar] [CrossRef]
- Ma, X.; Wang, W.; Wang, D.; Ding, T.; Ye, X.; Liu, D. Degradation kinetics and structural characteristics of pectin under simultaneous sonochemical-enzymatic functions. Carbohyd. Polym. 2016, 154, 176–185. [Google Scholar] [CrossRef] [Green Version]
- Coimbra, M.A.; Barros, A.; Barros, M.; Rutledge, D.N.; Delgadillo, I. Multivariate analysis of uronic acid and neutral sugars in whole pectic samples by FT-IR spectroscopy. Carbohyd. Polym. 1998, 37, 241–248. [Google Scholar] [CrossRef]
- Gautam, M.; Saha, S.; Bani, S.; Kaul, A.; Mishra, S.; Patil, D.; Satti, N.K.; Suri, K.A.; Gairola, S.; Suresh, K.; et al. Immunomodulatory activity of Asparagus racemosus on systemic Th1/Th2 immunity: Implications for immunoadjuvant potential. J. Ethnopharmacol. 2009, 121, 241–247. [Google Scholar] [CrossRef]
- Agarwal, R.; Diwanay, S.; Patki, P.; Patwardhan, B. Studies on immunomodulatory activity of Withaniasomnifera (Ashwagandha) extracts in experimental immune inflammation. J. Ethnopharmacol. 1999, 67, 27–35. [Google Scholar] [CrossRef]
- Meng, F.; Xu, P.; Wang, X.; Huang, Y.; Wu, L.; Chen, Y.; Teng, L.; Wang, D. Investigation on the immunomodulatory activities of Sarcodonimbricatus extracts in a cyclophosphamide (CTX)-induced immunosuppressanted mouse model. Saudi Pharm. J. 2017, 25, 460–463. [Google Scholar] [CrossRef] [PubMed]
- Sherif, I.O. The effect of natural antioxidants in cyclophosphamide-induced hepatotoxicity: Role of Nrf2/HO-1 pathway. Int. ImmunoPharmacol. 2018, 61, 29–36. [Google Scholar] [CrossRef]
- Zhu, Z.Y.; Meng, M.; Sun, H.; Li, Y.; Ren, Y.Y.; Zhang, Y. Immunostimulatory activity of glycopeptides from Paecilomycessinensis under normal and cyclophosphamide induced immunosuppressive conditions in mice models. Food Funct. 2016, 7, 3566–3576. [Google Scholar] [CrossRef] [PubMed]
- Wang, Y.; Qi, Q.; Li, A.; Yang, M.; Huang, W.; Xu, H.; Zhao, Z.; Li, S. Immuno-enhancement effects of YifeiTongluo Granules on cyclophosphamide-induced immunosuppression in Balb/c mice. J. Ethnopharmacol. 2016, 194, 72–82. [Google Scholar] [CrossRef] [PubMed]
- Schoenaker, M.H.D.; Henriet, S.S.; Zonderland, J.; van Deuren, M.; Pan-Hammarstrom, Q.; Posthumus-van Sluijs, S.J.; Pico-Knijnenburg, I.; Weemaes, C.M.R.; IJspeert, H. Immunodeficiency in bloom’s syndrome. J. Clin. Immunol. 2018, 38, 35–44. [Google Scholar] [CrossRef] [Green Version]
- He, L.X.; Zhang, Z.F.; Sun, B.; Chen, Q.H.; Liu, R.; Ren, J.W.; Wang, J.B.; Li, Y. Sea cucumber (Codonopsispilosula) oligopeptides: Immunomodulatory effects based on stimulating Th cells, cytokine secretion and antibody production. Food Funct. 2016, 7, 1208–1216. [Google Scholar] [CrossRef]
- Payling, L.; Fraser, K.; Loveday, S.M.; Sims, I.; Roy, N.; McNabb, W. The effects of carbohydrate structure on the composition and functionality of the human gut microbiota. Trends Food Sci. Tech. 2020, 97, 233–248. [Google Scholar] [CrossRef]
- Chen, D.; Zheng, C.; Yang, J.; Li, J.; Su, J.; Xie, Y.; Lai, G. Immunomodulatory activities of a fungal protein extracted from hericiumerinaceus through regulating the gut microbiota. Front Immunol. 2017, 8, 666. [Google Scholar]
- Han, X.Y.; Andrade, R.A. Brevundimonasdiminuta infections and its resistance to fluoroquinolones. J. Antimicrob. Chemother. 2005, 55, 853–859. [Google Scholar] [CrossRef] [Green Version]
- Reeves, A.E.; Koenigsknecht, M.J.; Bergin, I.L.; Young, V.B. Suppression of Clostridium difficile in the gastrointestinal tracts of germfree mice inoculated with a murine isolate from the family Lachnospiraceae. Infect Immun. 2012, 80, 3786–3794. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tang, C.; Sun, J.; Zhou, B.; Jin, C.; Liu, J.; Kan, J.; Qian, C.; Zhang, N. Effects of polysaccharides from purple sweet potatoes on immune response and gut microbiota composition in normal and cyclophosphamide treated mice. Food Funct. 2018, 9, 937–950. [Google Scholar] [CrossRef] [PubMed]
- Makki, K.; Deehan, E.C.; Walter, J.; Backhed, F. The impact of dietary fiber on gut microbiota in host health and disease. Cell Host Microbe. 2018, 23, 705–715. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Klinchongkon, K.; Khuwijitjaru, P.; Adachi, S.; Bindereif, B.; Karbstein, H.P.; van der Schaaf, U.S. Emulsifying properties of conjugates formed between whey protein isolate and subcritical-water hydrolyzed pectin. Food Hydrocolloid. 2019, 91, 174–181. [Google Scholar] [CrossRef]
- Lu, J.; Li, J.; Jin, R.; Li, S.; Yi, J.; Huang, J. Extraction and characterization of pectin from PremnamicrophyllaTurcz leaves. Int. J. Biol. MacroMol. 2019, 131, 323–328. [Google Scholar] [CrossRef]
- Ge, X.; Chen, S.; Liu, M.; Liang, T.; Liu, C. Evodiamine attenuates pdgf-bb-induced migration of rat vascular smooth muscle cells through activating ppar gamma. Int. J. Mol. Sci. 2015, 16, 28180–28193. [Google Scholar] [CrossRef] [Green Version]
- Zhang, C.; Wu, W.; Li, X.; Xin, X.; Liu, D. Daily supplementation with fresh angelica keiskei juice alleviates high-fat diet-induced obesity in mice by modulating gut microbiota composition. Mol. Nutr. Food Res. 2019, 63, 1900248. [Google Scholar] [CrossRef]
Sample Availability: Samples of the compounds subcritical water-hydrolyzed citrus pectin are available from the authors. |










{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Groups | Body Weight (g) | Body Weight Gain after Treatment with Cy (g) | Thymus Index (mg/g) | Spleen Index (mg/g) |
|---|---|---|---|---|
| Control | 33.38±1.32 | 5.24 ± 1.57 | 1.47 ± 0.38 | 2.72 ± 0.67 |
| Cy | 31.62 ± 1.94 | 3.61 ± 0.54 * | 1.39 ± 0.01 | 2.31 ± 0.50 |
| Cy + LMS | 32.46 ± 1.16 | 4.62 ± 0.72 # | 1.26 ± 0.26 | 2.70 ± 0.24 |
| Cy + LSCP | 31.76 ± 2.93 | 4.22 ± 1.34 # | 1.31 ± 0.24 | 3.06 ± 0.26 # |
| Cy + MSCP | 30.02 ± 2.29 * | 3.75 ± 0.90 | 1.40 ± 0.41 | 2.65 ± 0.29 |
| Cy + HSCP | 30.72 ± 2.63 * | 4.14 ± 0.83 | 1.18 ± 0.24 | 2.41 ± 0.33 |
| Cy + CP | 29.50 ± 1.83 *** | 3.02 ± 1.16 * | 1.20 ± 0.25 | 2.46 ± 0.34 |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Chen, J.; Zhang, C.; Xia, Q.; Liu, D.; Tan, X.; Li, Y.; Cao, Y. Treatment with Subcritical Water-Hydrolyzed Citrus Pectin Ameliorated Cyclophosphamide-Induced Immunosuppression and Modulated Gut Microbiota Composition in ICR Mice. Molecules 2020, 25, 1302. https://doi.org/10.3390/molecules25061302
Chen J, Zhang C, Xia Q, Liu D, Tan X, Li Y, Cao Y. Treatment with Subcritical Water-Hydrolyzed Citrus Pectin Ameliorated Cyclophosphamide-Induced Immunosuppression and Modulated Gut Microbiota Composition in ICR Mice. Molecules. 2020; 25(6):1302. https://doi.org/10.3390/molecules25061302
Chicago/Turabian StyleChen, Jianbing, Chengcheng Zhang, Qile Xia, Daqun Liu, Xinghe Tan, Yingdi Li, and Yan Cao. 2020. "Treatment with Subcritical Water-Hydrolyzed Citrus Pectin Ameliorated Cyclophosphamide-Induced Immunosuppression and Modulated Gut Microbiota Composition in ICR Mice" Molecules 25, no. 6: 1302. https://doi.org/10.3390/molecules25061302