Salvianolic Acid A Ameliorates Early-Stage Atherosclerosis Development by Inhibiting NLRP3 Inflammasome Activation in Zucker Diabetic Fatty Rats

 ,
,
Abstract
:1. Introduction
2. Results
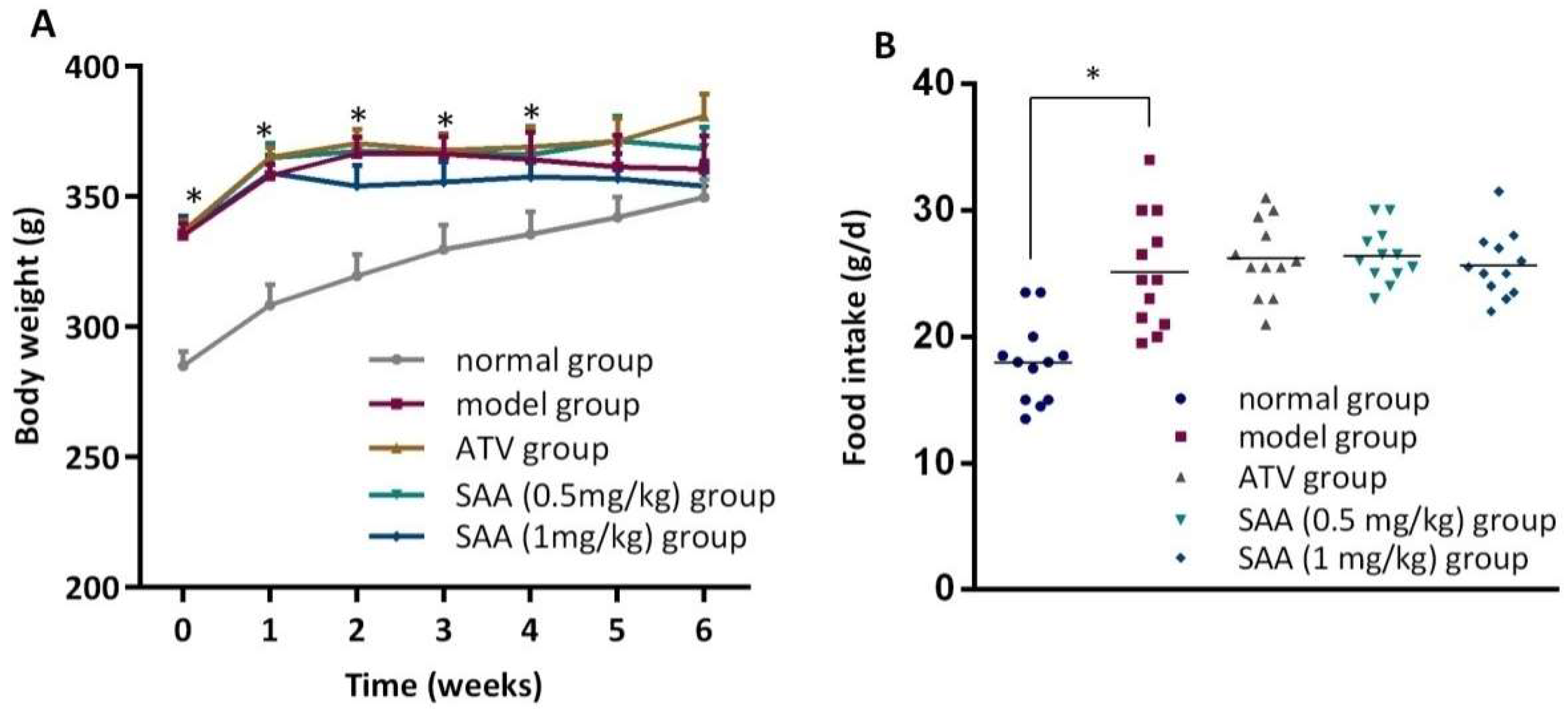
2.1. Effects of SAA on the Body Weight and Food Intake in T2DM ZDF Rats with AS
2.2. Effects of SAA on Blood Glucose and HbA1c Levels in T2DM ZDF Rats with AS
2.3. Effects of SAA on Blood Lipid Levels in T2DM ZDF Rats with AS
2.4. SAA Ameliorated Histopathological Changes in the Aortic Tissues of T2DM ZDF Rats with AS
2.5. Effects of SAA on Serum Hs-CRP in T2DM ZDF Rats with AS
2.6. Effects of SAA on the Expression of NLRP3 and Caspase-1 in the Aorta Tissue of T2DM ZDF Rats with AS
2.7. Effects of SAA on the Expression Levels of NF-κB and IL-1β in the Aortic Tissue of T2DM ZDF Rats with AS
3. Materials and Methods
3.1. Experimental Animals and Chemical Reagents
3.2. Establishing a ZDF Rat Model of T2DM with AS
3.3. Measurement of Blood Biochemistry
3.4. Histological Evaluation of the Aortas
3.5. RNA Extraction and Quantitative Real-Time PCR (qPCR)
3.6. Western Blotting Analysis
3.7. Statistical Analysis
4. Discussion
Author Contributions
Funding
Conflicts of Interest
References
- Chaudhury, A.; Duvoor, C.; Dendi, R.; Sena, V.; Kraleti, S.; Chada, A.; Ravilla, R.; Marco, A.; Shekhawat, N.S.; Montales, M.T. Clinical review of antidiabetic drugs: Implications for type 2 diabetes mellitus management. Front. Endocrinol. 2017, 8, 6. [Google Scholar] [CrossRef] [Green Version]
- The Emerging Risk Factors Collaboration. Diabetes mellitus, fasting blood glucose concentration, and risk of vascular disease: A collaborative meta-analysis of 102 prospective studies. Lancet 2010, 375, 2215–2222. [Google Scholar] [CrossRef] [Green Version]
- Low Wang, C.C.; Hess, C.N.; Hiatt, W.R.; Goldfine, A.B. Clinical update: Cardiovascular disease in diabetes mellitus: Atherosclerotic cardiovascular disease and heart failure in type 2 diabetes mellitus–mechanisms, management, and clinical considerations. Circ. J. 2016, 133, 2459–2502. [Google Scholar] [CrossRef]
- Rehman, K.; Akash, M.S.H. Mechanism of generation of oxidative stress and pathophysiology of type 2 diabetes mellitus: How are they interlinked? J. Cell. Biochem. 2017, 118, 3577–3585. [Google Scholar] [CrossRef]
- Wang, D.; Zhang, Y.; Shen, C. Research update on the association between SFRP5, an anti-inflammatory adipokine, with obesity, type 2 diabetes mellitus and coronary heart disease. J. Cell. Mol. Med. 2020. [Google Scholar] [CrossRef] [PubMed]
- La Sala, L.; Prattichizzo, F.; Ceriello, A. The link between diabetes and atherosclerosis. Eur. J. Prev. Cardiol. 2019, 26, 15–24. [Google Scholar] [CrossRef] [PubMed]
- Doodnauth, S.A.; Grinstein, S.; Maxson, M.E. Constitutive and stimulated macropinocytosis in macrophages: Roles in immunity and in the pathogenesis of atherosclerosis. Philos. Trans. R. Soc. Lond. B Biol. Sci. 2019, 374, 20180147. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Soeki, T.; Sata, M. Inflammatory biomarkers and atherosclerosis. Int. Heart J. 2016, 15–346. [Google Scholar] [CrossRef] [Green Version]
- Ho, J.H.-C.; Hong, C.-Y. Salvianolic acids: Small compounds with multiple mechanisms for cardiovascular protection. J. Biomed. Sci. Eng. 2011, 18, 30. [Google Scholar] [CrossRef] [Green Version]
- Zhou, A.-M.; Xiang, Y.-J.; Liu, E.-Q.; Cai, C.-H.; Wu, Y.-H.; Yang, L.-B.; Zeng, C.-L. Salvianolic acid a inhibits platelet activation and aggregation in patients with type 2 diabetes mellitus. BMC Cardiovasc. Disord. 2020, 20, 1–10. [Google Scholar] [CrossRef]
- Hou, B.; Qiang, G.; Zhao, Y.; Yang, X.; Chen, X.; Yan, Y.; Wang, X.; Liu, C.; Zhang, L.; Du, G. Salvianolic acid A protects against diabetic nephropathy through ameliorating glomerular endothelial dysfunction via inhibiting AGE-RAGE signaling. Cell. Physiol. Biochem. 2017, 44, 2378–2394. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wu, P.; Yan, Y.; Ma, L.-L.; Hou, B.-Y.; He, Y.-Y.; Zhang, L.; Niu, Z.-R.; Song, J.-K.; Pang, X.-C.; Yang, X.-Y. Effects of the Nrf2 protein modulator salvianolic acid A alone or combined with metformin on diabetes-associated macrovascular and renal injury. J. Biol. Chem. 2016, 291, 22288–22301. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wang, K.; Yang, Q.; Ma, Q.; Wang, B.; Wan, Z.; Chen, M.; Wu, L. Protective effects of salvianolic acid a against dextran sodium sulfate-induced acute colitis in rats. Nutrients 2018, 10, 791. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhang, H.-F.; Wang, Y.-L.; Gao, C.; Gu, Y.-T.; Huang, J.; Wang, J.-H.; Wang, J.-H.; Zhang, Z. Salvianolic acid A attenuates kidney injury and inflammation by inhibiting NF-κB and p38 MAPK signaling pathways in 5/6 nephrectomized rats. Acta Pharmacol. Sin. 2018, 39, 1855–1864. [Google Scholar] [CrossRef] [Green Version]
- Mao, K.; Shu, W.; Qiu, Q.; Gu, Q.; Wu, X. Salvianolic acid A protects retinal pigment epithelium from OX-LDL-induced inflammation in an age-related macular degeneration model. Discov. Med. 2017, 23, 129–148. [Google Scholar]
- Zeng, X.; Chen, X.; Qin, H.; Han, Y.; Chen, X.; Han, Z.; Zhao, W. Preventive effects of a natural anti-inflammatory agent salvianolic acid A on acute kidney injury in mice. Food Chem. Toxicol. 2020, 135, 110901. [Google Scholar] [CrossRef]
- Kaneto, H.; Katakami, N.; Matsuhisa, M.; Matsuoka, T.-A. Role of reactive oxygen species in the progression of type 2 diabetes and atherosclerosis. Mediators Inflamm. 2010, 2010. [Google Scholar] [CrossRef] [Green Version]
- Qiang, G.; Yang, X.; Shi, L.; Zhang, H.; Chen, B.; Zhao, Y.; Zu, M.; Zhou, D.; Guo, J.; Yang, H. Antidiabetic effect of salvianolic acid A on diabetic animal models via AMPK activation and mitochondrial regulation. Cell. Physiol. Biochem. 2015, 36, 395–408. [Google Scholar] [CrossRef]
- Lü, L.; Zhang, D.; Sun, B.; Hu, Y.; Yan, M.; Liu, K.; Li, X.; Ren, L. Apocynum leaf extract inhibits the progress of atherosclerosis in rats via the AMPK/mTOR pathway. Pharmazie 2017, 72, 41–48. [Google Scholar]
- Cordero-Herrera, I.; Martín, M.Á.; Escrivá, F.; Álvarez, C.; Goya, L.; Ramos, S. Cocoa-rich diet ameliorates hepatic insulin resistance by modulating insulin signaling and glucose homeostasis in Zucker diabetic fatty rats. J. Nutr. Biochem. 2015, 26, 704–712. [Google Scholar] [CrossRef]
- Almofti, M.R.; Huang, Z.; Yang, P.; Rui, Y.; Yang, P. Proteomic analysis of rat aorta during atherosclerosis induced by high cholesterol diet and injection of vitamin D3. Clin. Exp. Pharmacol. Physiol. 2006, 33, 305–309. [Google Scholar] [CrossRef]
- Currie, C.J.; Peters, J.R.; Tynan, A.; Evans, M.; Heine, R.J.; Bracco, O.L.; Zagar, T.; Poole, C.D. Survival as a function of HbA1c in people with type 2 diabetes: A retrospective cohort study. Lancet 2010, 375, 481–489. [Google Scholar] [CrossRef]
- Taskinen, M.-R.; BorÚn, J. New insights into the pathophysiology of dyslipidemia in type 2 diabetes. Atherosclerosis 2015, 239, 483–495. [Google Scholar] [CrossRef] [PubMed]
- Quispe, R.; Manalac, R.J.; Faridi, K.F.; Blaha, M.J.; Toth, P.P.; Kulkarni, K.R.; Nasir, K.; Virani, S.S.; Banach, M.; Blumenthal, R.S. Relationship of the triglyceride to high-density lipoprotein cholesterol (TG/HDL-C) ratio to the remainder of the lipid profile: The Very Large Database of Lipids-4 (VLDL-4) study. Atherosclerosis 2015, 242, 243–250. [Google Scholar] [CrossRef] [PubMed]
- Orekhov, A.N. LDL and foam cell formation as the basis of atherogenesis. Curr. Opin. Lipidol. 2018, 29, 279–284. [Google Scholar] [CrossRef] [PubMed]
- Plotkin, J.D.; Elias, M.G.; Dellinger, A.L.; Kepley, C.L. NF-κB inhibitors that prevent foam cell formation and atherosclerotic plaque accumulation. Nanomed. Nanotechnol. Biol. Med. 2017, 13, 2037–2048. [Google Scholar] [CrossRef]
- Chistiakov, D.A.; Orekhov, A.N.; Bobryshev, Y.V. LOX-1-mediated effects on vascular cells in atherosclerosis. Cell. Physiol. Biochem. 2016, 38, 1851–1859. [Google Scholar] [CrossRef]
- Qiang, G.; Yang, X.; Xuan, Q.; Shi, L.; Zhang, H.; Chen, B.; Li, X.; Zu, M.; Zhou, D.; Guo, J. Salvianolic acid A prevents the pathological progression of hepatic fibrosis in high-fat diet-fed and streptozotocin-induced diabetic rats. Am. J. Chin. Med. 2014, 42, 1183–1198. [Google Scholar] [CrossRef]
- Libby, P.; Ridker, P.M. Inflammation and atherosclerosis: Role of C-reactive protein in risk assessment. Am. J. Med. 2004, 116, 9–16. [Google Scholar] [CrossRef]
- Li, Y.; Zhong, X.; Cheng, G.; Zhao, C.; Zhang, L.; Hong, Y.; Wan, Q.; He, R.; Wang, Z. Hs-CRP and all-cause, cardiovascular, and cancer mortality risk: A meta-analysis. Atherosclerosis 2017, 259, 75–82. [Google Scholar] [CrossRef]
- Jin, Y.; Fu, J. Novel Insights Into the NLRP 3 Inflammasome in Atherosclerosis. J. Am. Heart. Assoc. 2019, 8, e012219. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jo, E.-K.; Kim, J.K.; Shin, D.-M.; Sasakawa, C. Molecular mechanisms regulating NLRP3 inflammasome activation. Cell. Mol. Immunol. 2016, 13, 148–159. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Orlowski, G.M.; Colbert, J.D.; Sharma, S.; Bogyo, M.; Robertson, S.A.; Rock, K.L. Multiple cathepsins promote pro–IL-1β synthesis and NLRP3-mediated IL-1β activation. J. Immunol. 2015, 195, 1685–1697. [Google Scholar] [CrossRef] [PubMed]
- Patel, S. Inflammasomes, the cardinal pathology mediators are activated by pathogens, allergens and mutagens: A critical review with focus on NLRP3. Biomed. Pharmacother. 2017, 92, 819–825. [Google Scholar] [CrossRef] [PubMed]
- Shao, B.-Z.; Xu, Z.-Q.; Han, B.-Z.; Su, D.-F.; Liu, C. NLRP3 inflammasome and its inhibitors: A review. Front. Pharmacol. 2015, 6, 262. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ranson, N.; Veldhuis, M.; Mitchell, B.; Fanning, S.; Cook, A.L.; Kunde, D.; Eri, R. NLRP3-Dependent and-Independent Processing of Interleukin (IL)-1β in Active Ulcerative Colitis. Int. J. Mol. Sci. 2019, 20, 57. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jiang, Y.; Du, H.; Liu, X.; Fu, X.; Li, X.; Cao, Q. Artemisinin alleviates atherosclerotic lesion by reducing macrophage inflammation via regulation of AMPK/NF-κB/NLRP3 inflammasomes pathway. J. Drug Target. 2020, 28, 70–79. [Google Scholar] [CrossRef]
- Zhuang, T.; Liu, J.; Chen, X.; Zhang, L.; Pi, J.; Sun, H.; Li, L.; Bauer, R.; Wang, H.; Yu, Z. Endothelial Foxp1 suppresses atherosclerosis via modulation of Nlrp3 inflammasome activation. Circ. Res. 2019, 125, 590–605. [Google Scholar] [CrossRef]
- Ding, C.; Zhao, Y.; Shi, X.; Zhang, N.; Zu, G.; Li, Z.; Zhou, J.; Gao, D.; Lv, L.; Tian, X. New insights into salvianolic acid A action: Regulation of the TXNIP/NLRP3 and TXNIP/ChREBP pathways ameliorates HFD-induced NAFLD in rats. Sci. Rep. 2016, 6, 28734. [Google Scholar] [CrossRef]
Sample Availability: Not available. |









{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Index | Normal Group | Model Group | ATV Group | SAA Group (0.5 mg/kg) | SAA Group (1 mg/kg) |
|---|---|---|---|---|---|
| HOMA-IR | 4.01 ± 0.42 | 13.18 ± 1.39 * | 9.43 ± 0.66 | 11.72 ± 1.16 | 12.00 ± 0.58 |
| HOMA-IS (×10−3) | 11.87 ± 1.55 | 3.49 ± 0.31 * | 4.51 ± 0.43 | 3.55 ± 0.39 | 3.72 ± 0.19 |
| Atherosclerosis Index | 0.71 ± 0.02 | 6.28 ± 0.32 * | 3.91 ± 0.42 # | 4.30 ± 0.53 # | 4.14 ± 0.61 # |
| Group | Time | |||
|---|---|---|---|---|
| 0 Week | 2 Weeks | 4 Weeks | 6 Weeks | |
| Normal group | 84.61 ± 3.18 | 83.52 ± 3.05 | 82.40 ± 3.03 | 84.12 ± 3.76 |
| Model group | 118.01 ± 5.66 | 198.33 ± 4.33 * | 207.16 ± 3.86 * | 198.15 ± 11.28 * |
| ATV Group | 110.52 ± 6.63 | 172.58 ± 6.02 # | 184.36 ± 5.78 # | 176.33 ± 6.80 |
| SAA Group (0.5 mg/kg) | 116.58 ± 4.87 | 193.86 ± 6.06 | 189.03 ± 5.26 # | 151.55 ± 6.79 # |
| SAA Group (1 mg/kg) | 109.37 ± 5.83 | 199.33 ± 5.90 | 179.51 ± 3.08 # | 152.95 ± 8.53 # |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Ma, Q.; Yang, Q.; Chen, J.; Yu, C.; Zhang, L.; Zhou, W.; Chen, M. Salvianolic Acid A Ameliorates Early-Stage Atherosclerosis Development by Inhibiting NLRP3 Inflammasome Activation in Zucker Diabetic Fatty Rats. Molecules 2020, 25, 1089. https://doi.org/10.3390/molecules25051089
Ma Q, Yang Q, Chen J, Yu C, Zhang L, Zhou W, Chen M. Salvianolic Acid A Ameliorates Early-Stage Atherosclerosis Development by Inhibiting NLRP3 Inflammasome Activation in Zucker Diabetic Fatty Rats. Molecules. 2020; 25(5):1089. https://doi.org/10.3390/molecules25051089
Chicago/Turabian StyleMa, Quanxin, Qinqin Yang, Jiaojiao Chen, Chen Yu, Lizong Zhang, Weimin Zhou, and Minli Chen. 2020. "Salvianolic Acid A Ameliorates Early-Stage Atherosclerosis Development by Inhibiting NLRP3 Inflammasome Activation in Zucker Diabetic Fatty Rats" Molecules 25, no. 5: 1089. https://doi.org/10.3390/molecules25051089