
Ginseng Extract Ameliorates the Negative Physiological Effects of Heat Stress by Supporting Heat Shock Response and Improving Intestinal Barrier Integrity: Evidence from Studies with Heat-Stressed Caco-2 Cells, C. elegans and Growing Broilers

 , , and
, , and
Abstract
:
1. Introduction
2. Results
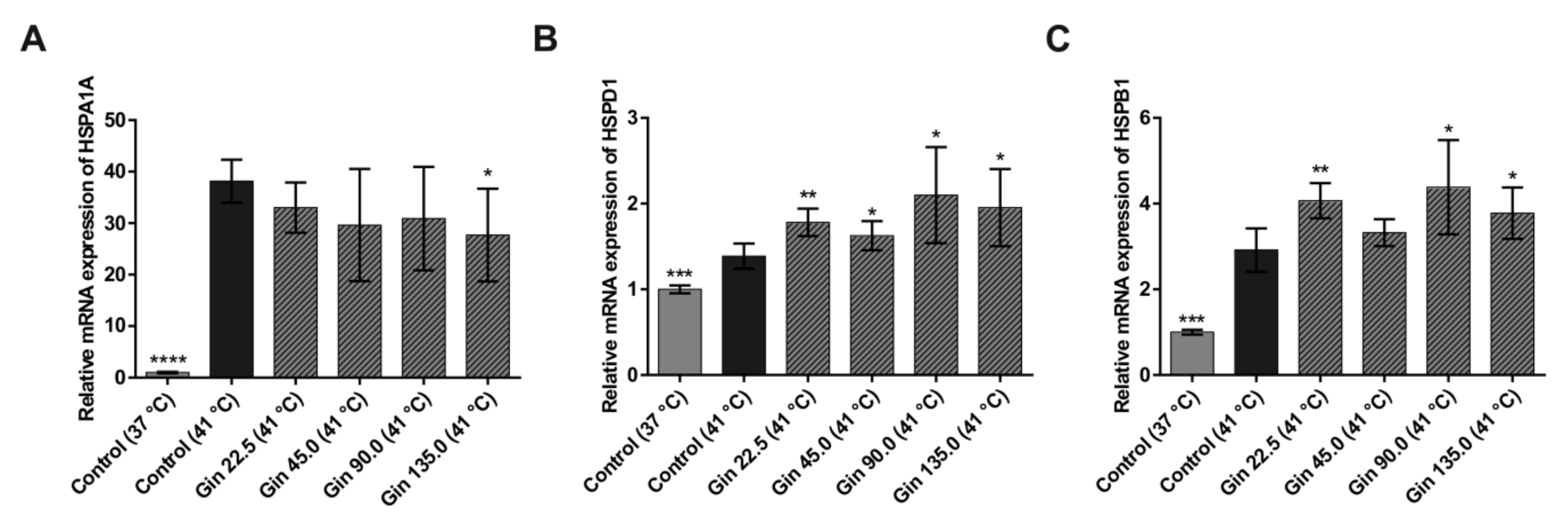
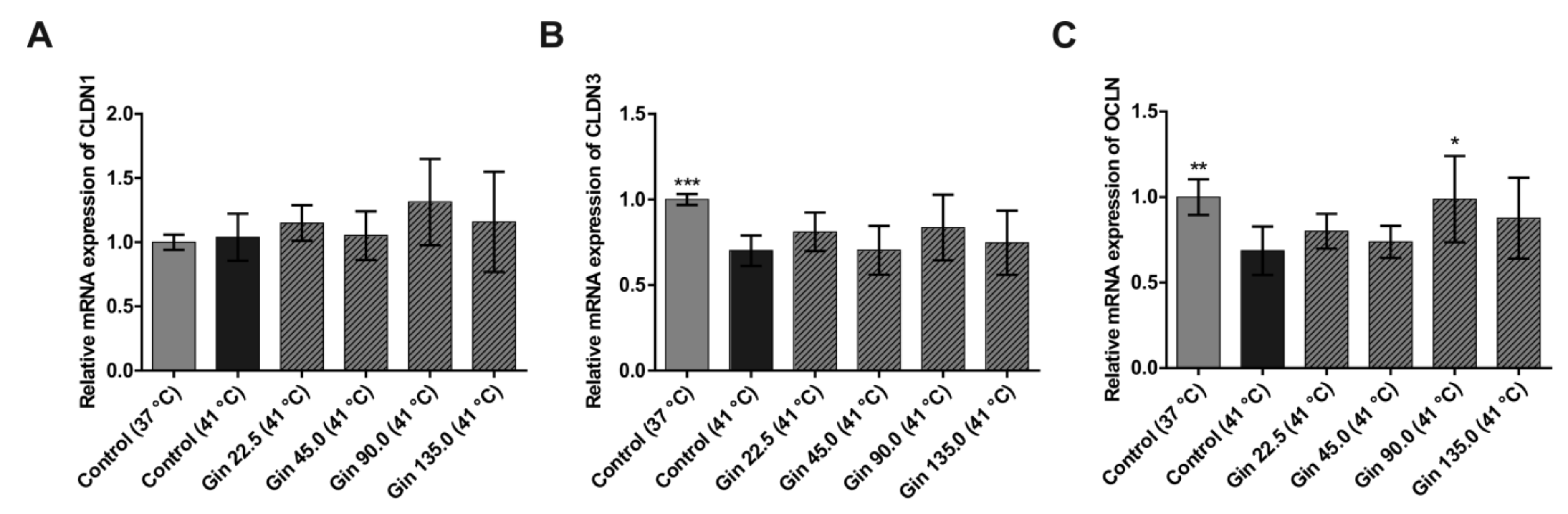
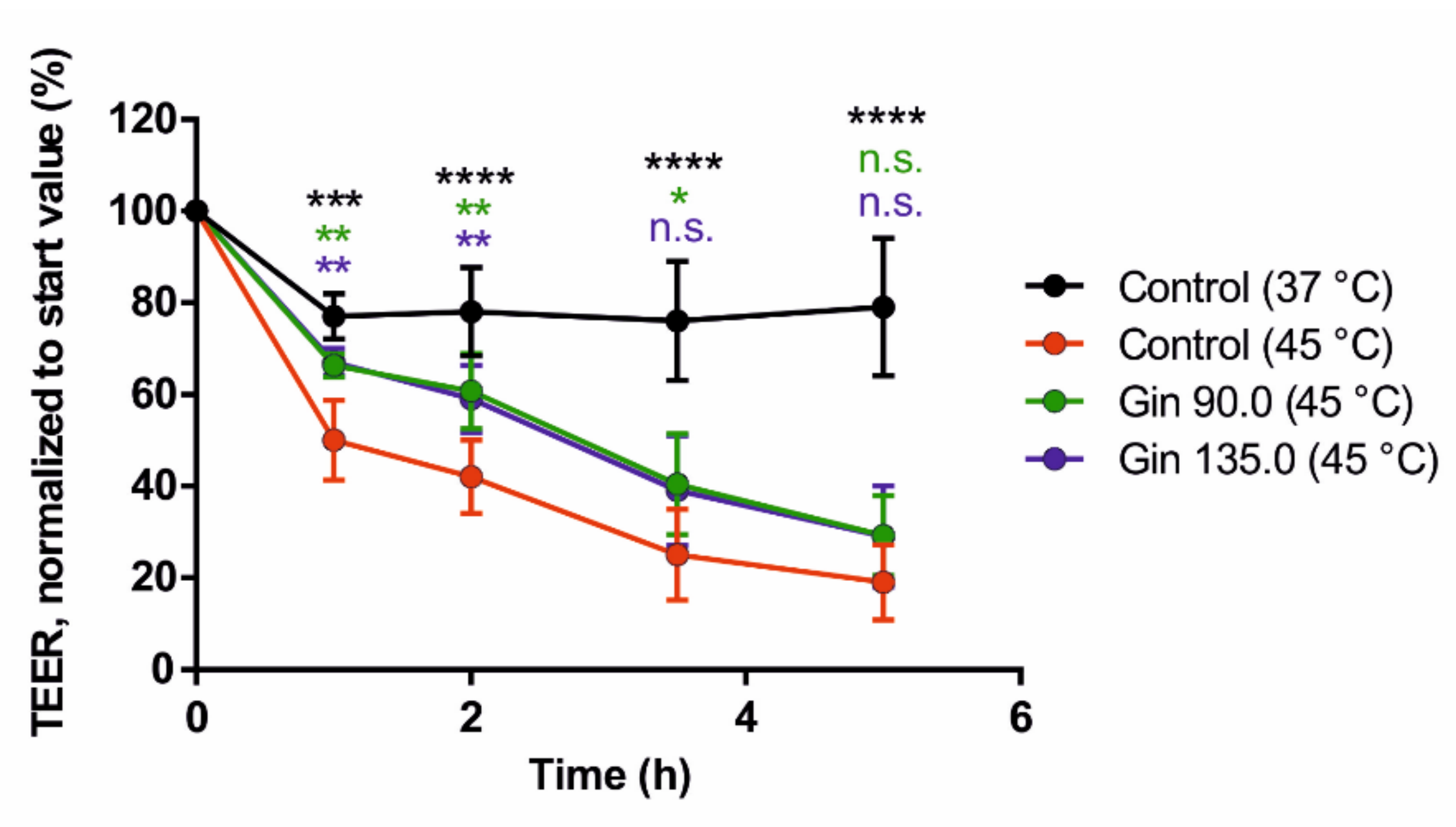
2.1. Study With Caco-2 Cells
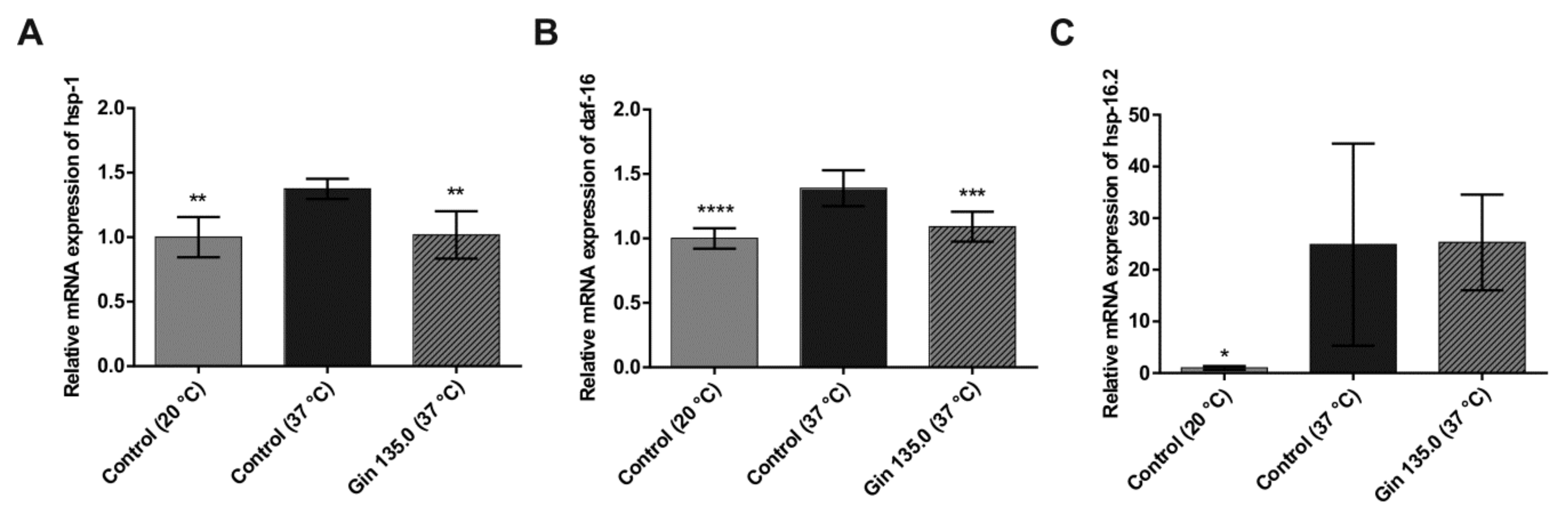
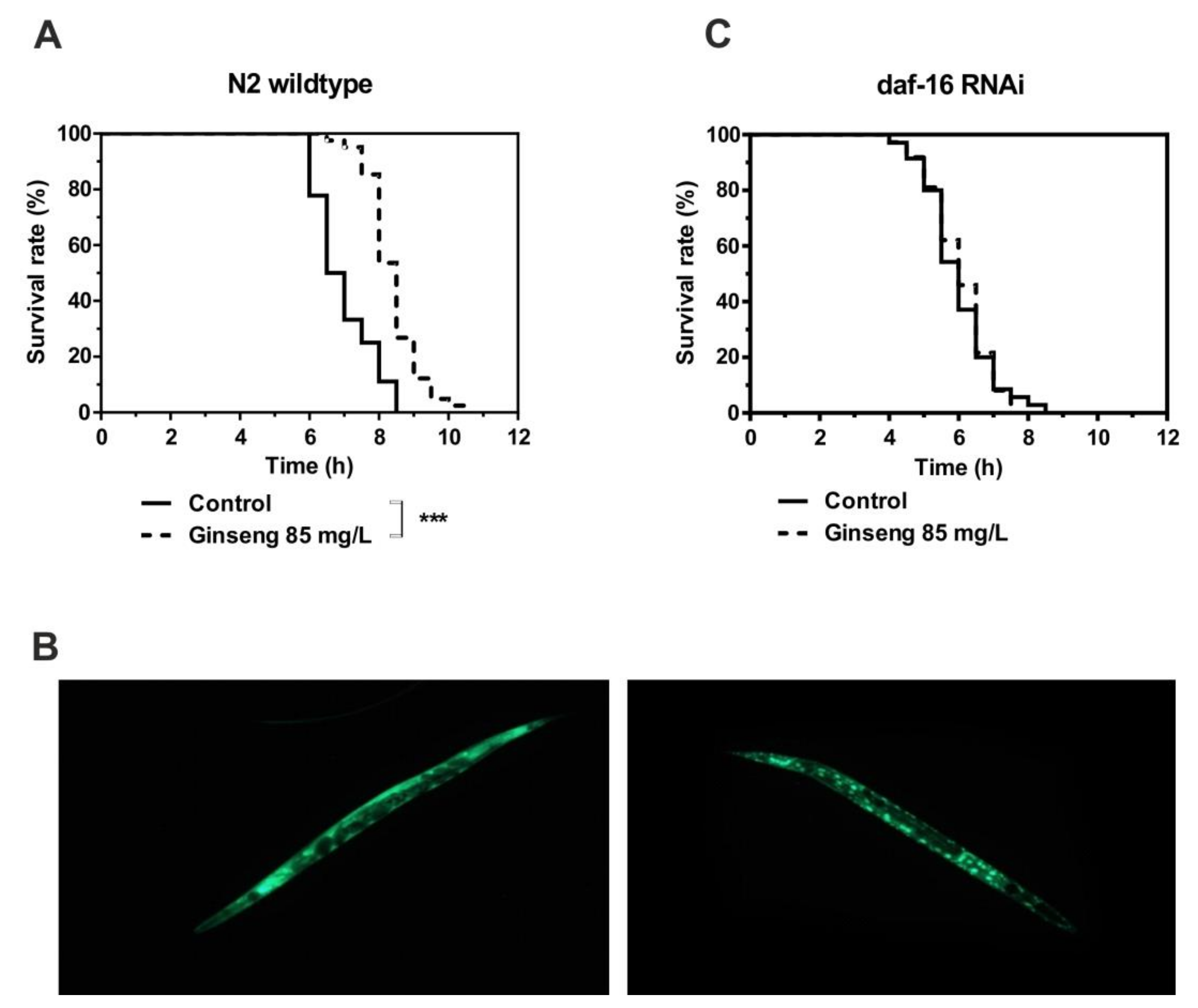
2.2. Study With C. elegans
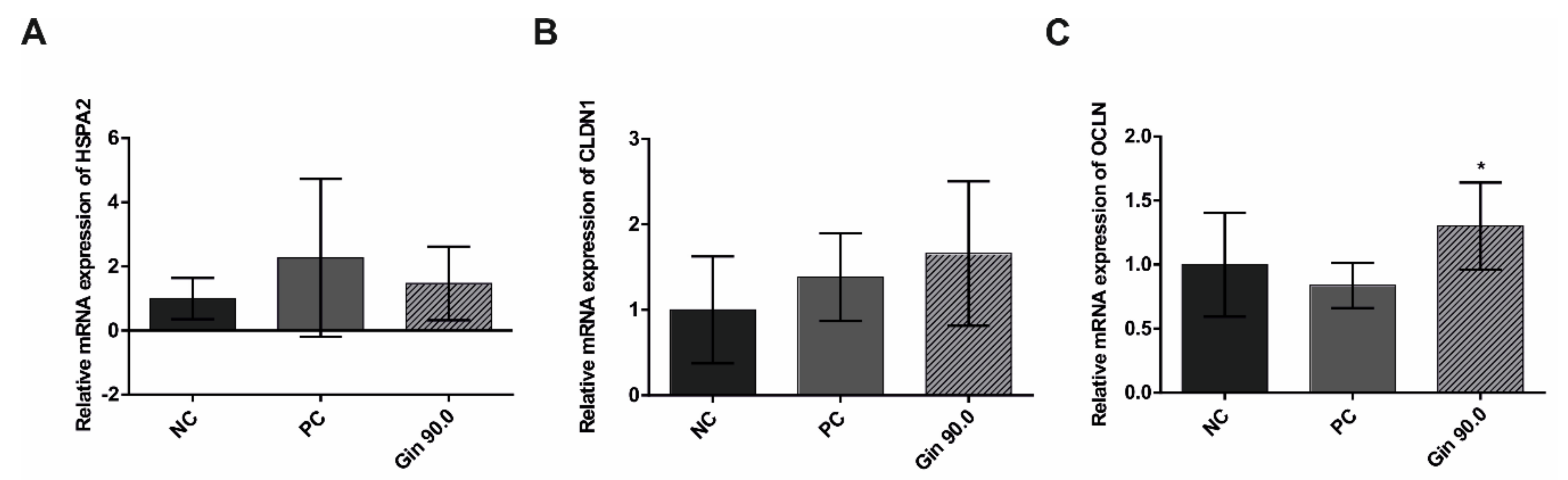
2.3. Study With Growing Broilers
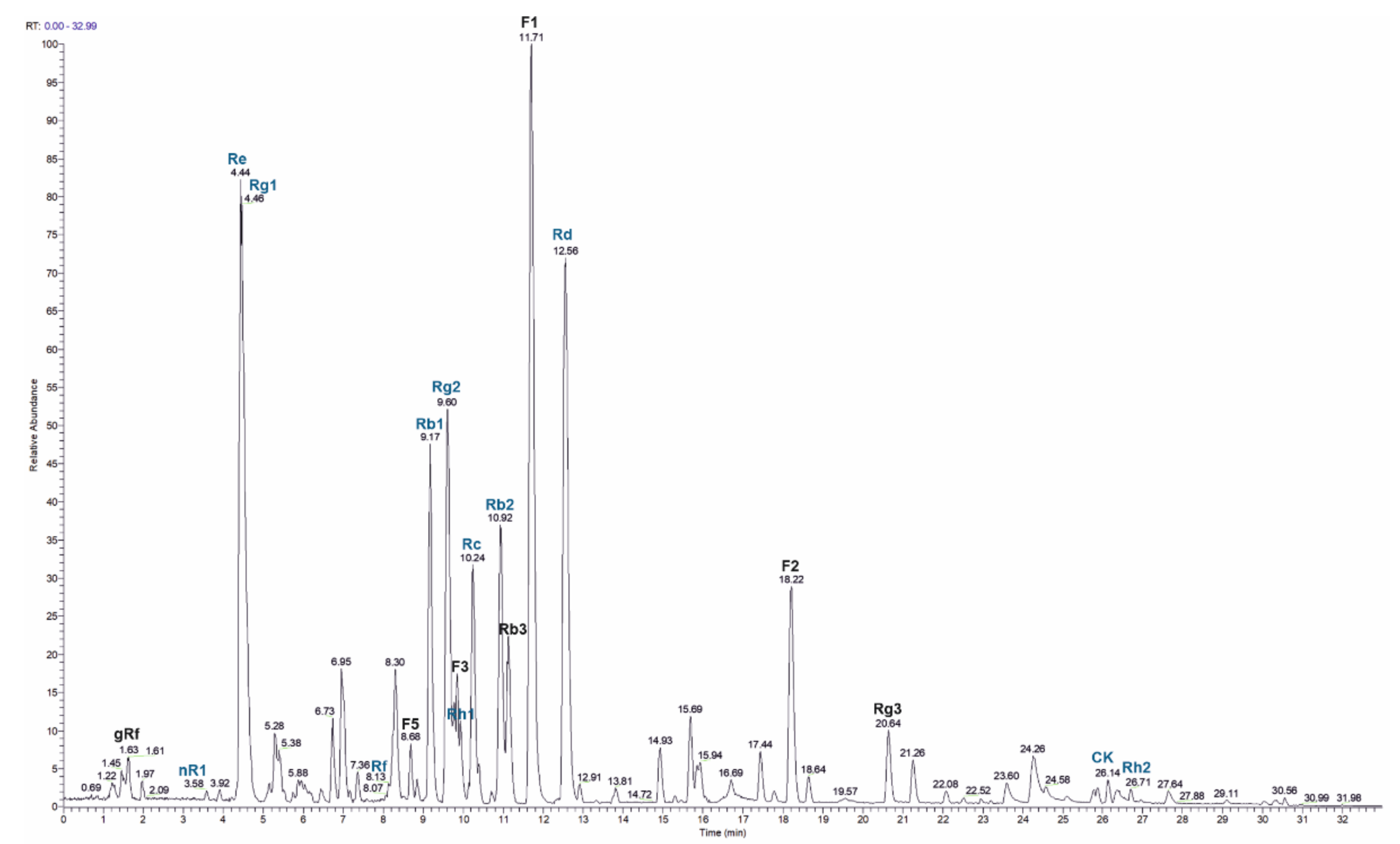
2.4. HPLC Analysis of Ginseng Extract
3. Discussion
4. Materials and Methods
4.1. Study With Caco-2 Cells
4.1.1. Caco-2 Cell Culture and Differentiation
4.1.2. Induction of Heat Stress
4.1.3. Detection of HSPA1A, HSPD1, HSPB1, CLDN1, CLDN3 and OCLN mRNA Expression by Quantitative Real-Time PCR
4.1.4. Validation of Intestinal Barrier Integrity by TEER Measurements
4.2. Study With C. elegans
4.2.1. C. elegans Maintenance
4.2.2. Treatment of Nematodes with Ginseng Extract
4.2.3. mRNA Expression of hsp-16.2, hsp-1 and daf-16 by Quantitative Real-Time PCR
4.2.4. Determination of Survival Under Heat Stress
4.2.5. Nuclear daf-16 Translocation
4.2.6. Daf-16 RNAi
4.3. Study With Growing Broilers
4.3.1. Animals, Diets and Experimental Design
4.3.2. Detection of HSP2A, CLDN1 and OCLN mRNA Expression by Quantitative Real-Time PCR
4.4. HPLC Analysis of Ginseng Extract
4.5. Calculations and Statistics
5. Conclusions
Author Contributions
Funding
Conflicts of Interest
References
- Gummow, B. Challenges posed by new and re-emerging infectious diseases in livestock production, wildlife and humans. Livest. Sci. 2010, 130, 41–46. [Google Scholar] [CrossRef]
- Hellberg, R.S.; Chu, E. Effects of climate change on the persistence and dispersal of foodborne bacterial pathogens in the outdoor environment: A review. Crit. Rev. Microbiol. 2016, 42, 548–572. [Google Scholar] [CrossRef] [PubMed]
- Selye, H. Forty years of stress research: Principal remaining problems and misconceptions. Can. Med. Assoc. J. 1976, 115, 53–56. [Google Scholar] [PubMed]
- Settar, P.; Yalcin, S.; Turkmut, L.; Ozkan, S.; Cahanar, A. Season by genotype interaction related to broiler growth rate and heat tolerance. Poult. Sci. J. 1999, 78, 1353–1358. [Google Scholar] [CrossRef] [PubMed]
- Ebeid, T.A.; Suzuki, T.; Sugiyama, T. High ambient temperature influences eggshell quality and calbindin-D28k localization of eggshell gland and all intestinal segments of laying hens. Poult. Sci. J. 2012, 91, 2282–2287. [Google Scholar] [CrossRef]
- Quinteiro-Filho, W.M.; Ribeiro, A.; Ferraz-de-Paula, V.; Pinheiro, M.L.; Sakai, M.; Sa, L.R.; Ferreira, A.J.; Palermo-Neto, J. Heat stress impairs performance parameters, induces intestinal injury, and decreases macrophage activity in broiler chickens. Poult. Sci. J. 2010, 89, 1905–1914. [Google Scholar] [CrossRef]
- Wang, R.H.; Liang, R.R.; Lin, H.; Zhu, L.X.; Zhang, Y.M.; Mao, Y.W.; Dong, P.C.; Niu, L.B.; Zhang, M.H.; Luo, X. Effect of acute heat stress and slaughter processing on poultry meat quality and postmortem carbohydrate metabolism. Poult. Sci. J. 2017, 96, 738–746. [Google Scholar] [CrossRef]
- Gupta, A.; Chauhan, N.R.; Chowdhury, D.; Singh, A.; Meena, R.C.; Chakrabarti, A.; Singh, S.B. Heat stress modulated gastrointestinal barrier dysfunction: Role of tight junctions and heat shock proteins. Scand. J. Gastroentero. 2017, 52, 1315–1319. [Google Scholar] [CrossRef]
- Koch, F.; Thom, U.; Albrecht, E.; Weikard, R.; Nolte, W.; Kuhla, B.; Kuehn, C. Heat stress directly impairs gut integrity and recruits distinct immune cell populations into the bovine intestine. PNAS 2019, 116, 10333–10338. [Google Scholar] [CrossRef] [Green Version]
- Song, J.; Xiao, K.; Ke, Y.L.; Jiao, L.F.; Hu, C.H.; Diao, Q.Y.; Shi, B.; Zou, X.T. Effect of a probiotic mixture on intestinal microflora, morphology, and barrier integrity of broilers subjected to heat stress. Poult. Sci. J. 2014, 93, 581–588. [Google Scholar] [CrossRef]
- Wieten, L.; Broere, F.; van der Zee, R.; Koerkamp, E.K.; Wagenaar, J.; van Eden, W. Cell stress induced HSP are targets of regulatory T cells: A role for HSP inducing compounds as anti-inflammatory immuno-modulators? FEBS Lett. 2007, 581, 3716–3722. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gu, X.H.; Hao, Y.; Wang, X.L. Overexpression of heat shock protein 70 and its relationship to intestine under acute heat stress in broilers: 2. Intestinal oxidative stress. Poult. Sci. J. 2012, 91, 790–799. [Google Scholar] [CrossRef] [PubMed]
- Eklund, M.; Bauer, E.; Wamatu, J.; Mosenthin, R. Potential nutritional and physiological functions of betaine in livestock. Nutr. Res. Rev. 2005, 18, 31–48. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Liu, W.; Yuan, Y.; Sun, C.; Balasubramanian, B.; Zhao, Z.; An, L. Effects of Dietary Betaine on Growth Performance, Digestive Function, Carcass Traits, and Meat Quality in Indigenous Yellow-Feathered Broilers under Long-Term Heat Stress. Animals 2019, 9, 506. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Park, S.O.; Kim, W.K. Effects of betaine on biological functions in meat-type ducks exposed to heat stress. Poult. Sci. J. 2017, 96, 1212–1218. [Google Scholar] [CrossRef] [PubMed]
- Sayed, M.A.; Downing, J. The effects of water replacement by oral rehydration fluids with or without betaine supplementation on performance, acid-base balance, and water retention of heat-stressed broiler chickens. Poult. Sci. J. 2011, 90, 157–167. [Google Scholar] [CrossRef]
- Kitts, D.; Hu, C. Efficacy and safety of ginseng. Public Health Nutr. 2000, 3, 473–485. [Google Scholar] [CrossRef] [Green Version]
- Kim, J.S.; Kim, K.W.; Choi, K.J. Screening of active components of red ginseng saponin. J. Ginseng Res. 1996, 20, 173–178. [Google Scholar]
- Kim, Y.H.; Park, K.H.; Rho, H.M. Transcriptional activation of the Cu,Zn-superoxide dismutase gene through the AP2 site by ginsenoside Rb2 extracted from a medicinal plant, Panax ginseng. J. Biol. Chem. 1996, 271, 24539–24543. [Google Scholar] [CrossRef] [Green Version]
- Seong, M.A.; Woo, J.K.; Kang, J.H.; Jang, Y.S.; Choi, S.; Lee, T.H.; Jung, K.H.; Kang, D.K.; Hurh, B.S.; Kim, D.E.; et al. Oral administration of fermented wild ginseng ameliorates DSS-induced acute colitis by inhibiting NF-kappaB signaling and protects intestinal epithelial barrier. BMB Rep. 2015, 48, 419–425. [Google Scholar] [CrossRef] [Green Version]
- Kim, K.J.; Yoon, K.Y.; Hong, H.D.; Lee, B.Y. Role of the Red Ginseng in Defense against the Environmental Heat Stress in Sprague Dawley Rats. Molecules 2015, 20, 20240–20253. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hartwig, K.; Heidler, T.; Moch, J.; Daniel, H.; Wenzel, U. Feeding a ROS-generator to Caenorhabditis elegans leads to increased expression of small heat shock protein HSP-16.2 and hormesis. Genes Nutr. 2009, 4, 59–67. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Artursson, P. Epithelial transport of drugs in cell culture. I: A model for studying the passive diffusion of drugs over intestinal absorptive (Caco-2) cells. J. Pharm. Sci. 1990, 79, 476–482. [Google Scholar] [CrossRef] [PubMed]
- Agostini, P.; Auer, B.; van Hofstraeten, W.; de Lange, L. The effect of four betaine products on replacing Methionine and Choline supplementation on growth performance in broiler chickens from 10 to 28 days of age. Available online: https://www.agrana.com/fileadmin/inhalte/agrana_group/2017/images/Starch/AGR_PB_Schothorst_A4.pdf. (accessed on 13 February 2020).
- Srinongkote, S.; Auer, B.; Gavrau, A. Comparing the Effect of Different Methyl Group Donors on the Carcass Quality of Broilers in Challenged Conditions and the Effect on the Litter Quality, from 0 to 38 Days of Age. Available online: https://www.agrana.com/fileadmin/inhalte/agrana_group/2017/images/Starch/AGR_PB_Trial_Thailand_A4_EN_FG.pdf. (accessed on 13 February 2020).
- Lee, J.W.; Ji, S.H.; Lee, Y.S.; Choi, D.J.; Choi, B.R.; Kim, G.S.; Baek, N.I.; Lee, D.Y. Mass Spectrometry Based Profiling and Imaging of Various Ginsenosides from Panax ginseng Roots at Different Ages. Int J. Mol. Sci. 2017, 18, 1114. [Google Scholar] [CrossRef] [Green Version]
- Lara, L.J.; Rostagno, M.H. Impact of Heat Stress on Poultry Production. Animals 2013, 3, 356–369. [Google Scholar] [CrossRef]
- Leung, K.W.; Wong, A.S. Pharmacology of ginsenosides: A literature review. Chin. Med. 2010, 5, 20. [Google Scholar] [CrossRef] [Green Version]
- Kang, H.K.; Park, S.B.; Kim, C.H. Effect of Dietary Supplementation of Red Ginseng By-product on Laying Performance, Blood Biochemistry, Serum Immunoglobulin and Microbial Population in Laying Hens. Asian Australas J. Anim. Sci. 2016, 29, 1464–1469. [Google Scholar] [CrossRef] [Green Version]
- Yang, C.M.; Han, Q.J.; Wang, K.L.; Xu, Y.L.; Lan, J.H.; Cao, G.T. Astragalus and Ginseng Polysaccharides Improve Developmental, Intestinal Morphological, and Immune Functional Characters of Weaned Piglets. Front. Physiol. 2019, 10, 418. [Google Scholar] [CrossRef] [Green Version]
- Murphy, C.T.; McCarroll, S.A.; Bargmann, C.I.; Fraser, A.; Kamath, R.S.; Ahringer, J.; Li, H.; Kenyon, C. Genes that act downstream of DAF-16 to influence the lifespan of Caenorhabditis elegans. Nature 2003, 424, 277–283. [Google Scholar] [CrossRef]
- Tissenbaum, H.A. DAF-16: FOXO in the Context of C. elegans. Curr. Top. Dev. Biol. 2018, 127, 1–21. [Google Scholar]
- Tóth, M.E.; Gombos, I.; Sántha, M. Heat shock proteins and their role in human diseases. Acta Biol. Szeged. 2015, 59, 121–141. [Google Scholar]
- Chen, M.Z.; Moily, N.S.; Bridgford, J.L.; Wood, R.J.; Radwan, M.; Smith, T.A.; Song, Z.; Tang, B.Z.; Tilley, L.; Xu, X.; et al. A thiol probe for measuring unfolded protein load and proteostasis in cells. Nat. Commun. 2017, 8, 474. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Song, J.H.; Kim, K.J.; Choi, S.Y.; Koh, E.J.; Park, J.; Lee, B.Y. Korean ginseng extract ameliorates abnormal immune response through the regulation of inflammatory constituents in Sprague Dawley rat subjected to environmental heat stress. J. Ginseng Res. 2019, 43, 252–260. [Google Scholar] [CrossRef] [PubMed]
- Yeo, M.; Kim, D.K.; Cho, S.W.; Hong, H.D. Ginseng, the root of Panax ginseng C.A. Meyer, protects ethanol-induced gastric damages in rat through the induction of cytoprotective heat-shock protein 27. Dig. Dis. Sci. 2008, 53, 606–613. [Google Scholar] [CrossRef]
- Awad, W.A.; Hess, C.; Hess, M. Enteric Pathogens and Their Toxin-Induced Disruption of the Intestinal Barrier through Alteration of Tight Junctions in Chickens. Toxins 2017, 9, 60. [Google Scholar] [CrossRef] [Green Version]
- Kim, C.Y.; Kim, K.H. Curcumin prevents leptin-induced tight junction dysfunction in intestinal Caco-2 BBe cells. J. Nutr. Biochem. 2014, 25, 26–35. [Google Scholar] [CrossRef]
- Dun, Y.; Liu, M.; Chen, J.; Peng, D.; Zhao, H.; Zhou, Z.; Wang, T.; Liu, C.; Guo, Y.; Zhang, C.; et al. Regulatory effects of saponins from Panax japonicus on colonic epithelial tight junctions in aging rats. J. Ginseng Res. 2018, 42, 50–56. [Google Scholar] [CrossRef]
- He, L.X.; Zhang, Z.F.; Zhao, J.; Li, L.; Xu, T.; Bin, S.; Ren, J.W.; Liu, R.; Chen, Q.H.; Wang, J.B.; et al. Ginseng oligopeptides protect against irradiation-induced immune dysfunction and intestinal injury. Sci. Rep. 2018, 8, 13916. [Google Scholar] [CrossRef] [Green Version]
- Tsapara, A.; Matter, K.; Balda, M.S. The heat-shock protein Apg-2 binds to the tight junction protein ZO-1 and regulates transcriptional activity of ZONAB. Mol. Cell. Biol. 2006, 17, 1322–1330. [Google Scholar] [CrossRef] [Green Version]
- Suzuki, T.; Yoshinaga, N.; Tanabe, S. Interleukin-6 (IL-6) regulates claudin-2 expression and tight junction permeability in intestinal epithelium. J. Biol. Chem. 2011, 286, 31263–31271. [Google Scholar] [CrossRef] [Green Version]
- Kang, O.J.; Kim, J.S. Comparison of Ginsenoside Contents in Different Parts of Korean Ginseng (Panax ginseng C.A. Meyer). Prev. Nutr. Food Sci. 2016, 21, 389–392. [Google Scholar] [CrossRef] [Green Version]
- Kim, J.H.; Yi, Y.S.; Kim, M.Y.; Cho, J.Y. Role of ginsenosides, the main active components of Panax ginseng, in inflammatory responses and diseases. J. Ginseng Res. 2017, 41, 435–443. [Google Scholar] [CrossRef] [Green Version]
- Lee, I.A.; Hyam, S.R.; Jang, S.E.; Han, M.J.; Kim, D.H. Ginsenoside Re ameliorates inflammation by inhibiting the binding of lipopolysaccharide to TLR4 on macrophages. J. Agric. Food Chem. 2012, 60, 9595–9602. [Google Scholar] [CrossRef] [PubMed]
- Vandesompele, J.; De Preter, K.; Pattyn, F.; Poppe, B.; Van Roy, N.; De Paepe, A.; Speleman, F. Accurate normalization of real-time quantitative RT-PCR data by geometric averaging of multiple internal control genes. Genome Biol. 2002, 3. [Google Scholar] [CrossRef] [Green Version]
- Haselgrubler, R.; Stadlbauer, V.; Stubl, F.; Schwarzinger, B.; Rudzionyte, I.; Himmelsbach, M.; Iken, M.; Weghuber, J. Insulin Mimetic Properties of Extracts Prepared from Bellis perennis. Molecules 2018, 23, 2605. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Muller, U.; Stubl, F.; Schwarzinger, B.; Sandner, G.; Iken, M.; Himmelsbach, M.; Schwarzinger, C.; Ollinger, N.; Stadlbauer, V.; Hoglinger, O.; et al. In Vitro and In Vivo Inhibition of Intestinal Glucose Transport by Guava (Psidium Guajava) Extracts. Mol. Nutr. Food. Res. 2018, 62, e1701012. [Google Scholar] [CrossRef] [Green Version]
- Mi, Z.; Fu, J.; Xiong, Y.; Tang, H. SUMOylation of RIG-I positively regulates the type I interferon signaling. Protein cell 2010, 1, 275–283. [Google Scholar] [CrossRef] [Green Version]
- Yamaguchi, H.; Kojima, T.; Ito, T.; Kimura, Y.; Imamura, M.; Son, S.; Koizumi, J.; Murata, M.; Nagayama, M.; Nobuoka, T.; et al. Transcriptional control of tight junction proteins via a protein kinase C signal pathway in human telomerase reverse transcriptase-transfected human pancreatic duct epithelial cells. Am. J. Pathol. 2010, 177, 698–712. [Google Scholar] [CrossRef]
- Bein, A.; Lubetzky, R.; Mandel, D.; Schwartz, B. TIMP-1 inhibition of occludin degradation in Caco-2 intestinal cells: A potential protective role in necrotizing enterocolitis. Pediatr. Res. 2015, 77, 649–655. [Google Scholar] [CrossRef] [Green Version]
- Yang, F.; Lei, X.; Rodriguez-Palacios, A.; Tang, C.; Yue, H. Selection of reference genes for quantitative real-time PCR analysis in chicken embryo fibroblasts infected with avian leukosis virus subgroup J. BMC Res. Notes 2013, 6, 402. [Google Scholar] [CrossRef] [Green Version]
- Stiernagle, T. Maintenance of C. elegans. In WormBook: The Online Review of C. elegans Biology; Caenorhabditis Genetics Center, University of Minnesota: Minneapolis, MN, USA, 2006. [Google Scholar] [CrossRef] [Green Version]
- Gill, M.S.; Olsen, A.; Sampayo, J.N.; Lithgow, G.J. An automated high-throughput assay for survival of the nematode Caenorhabditis elegans. Free Radic. Biol. Med. 2003, 35, 558–565. [Google Scholar] [CrossRef]
- Lehner, B.; Tischler, J.; Fraser, A.G. RNAi screens in Caenorhabditis elegans in a 96-well liquid format and their application to the systematic identification of genetic interactions. Nat. Protoc. 2006, 1, 1617–1620. [Google Scholar] [CrossRef] [PubMed]
- Timmons, L.; Court, D.L.; Fire, A. Ingestion of bacterially expressed dsRNAs can produce specific and potent genetic interference in Caenorhabditis elegans. Gene 2001, 263, 103–112. [Google Scholar] [CrossRef]
- Vantress, C. Cobb 500: Broiler Performance and Nutrition Supplement. Available online: https://cobbstorage.blob.core.windows.net/guides/5a171aa0-6994-11e8-9f14-bdc382f8d47e (accessed on 13 February 2020).
- Haselgrubler, R.; Lanzerstorfer, P.; Rohrl, C.; Stubl, F.; Schurr, J.; Schwarzinger, B.; Schwarzinger, C.; Brameshuber, M.; Wieser, S.; Winkler, S.M.; et al. Hypolipidemic effects of herbal extracts by reduction of adipocyte differentiation, intracellular neutral lipid content, lipolysis, fatty acid exchange and lipid droplet motility. Sci. Rep. 2019, 9, 10492. [Google Scholar] [CrossRef] [PubMed]
- Konig, A.; Schwarzinger, B.; Stadlbauer, V.; Lanzerstorfer, P.; Iken, M.; Schwarzinger, C.; Kolb, P.; Schwarzinger, S.; Morwald, K.; Brunner, S.; et al. Guava (Psidium guajava) Fruit Extract Prepared by Supercritical CO2 Extraction Inhibits Intestinal Glucose Resorption in a Double-Blind, Randomized Clinical Study. Nutrients 2019, 11, 1512. [Google Scholar] [CrossRef] [Green Version]
- Livak, K.J.; Schmittgen, T.D. Analysis of relative gene expression data using real-time quantitative PCR and the 2(-Delta Delta C(T)) Method. Methods 2001, 25, 402–408. [Google Scholar] [CrossRef]
Sample Availability: Samples of the compounds are not available from the authors. |








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Items | Negative Control | Positive Control ActiBeet® | Ginseng Extract | p-Value |
|---|---|---|---|---|
| Feed intake (g), 1–21 d | 866 ± 28.3 | 856 ± 27.5 | 856 ± 36.8 | 0.907 |
| Body weight (g), 21 d | 686 ± 35.7 | 690 ± 46.3 | 718 ± 17.7 | 0.519 |
| Body weight gain (g), 1–21 d | 643 ± 36.7 | 656 ± 33.4 | 676 ± 18.3 | 0.454 |
| Feed conversion (g/g), 1–21 d | 1.350 ± 0.055 A | 1.307 ± 0.027 AB | 1.267 ± 0.021 B | 0.086 |
| Feed intake (g), 22–42 d | 1879 ± 39.6 | 1905 ± 42.5 | 1879 ± 100.2 | 0.869 |
| Body weight (g), 42 d | 1966 ± 74.9 | 2034 ± 48.0 | 2062 ± 58.1 | 0.221 |
| Body weight gain (g), 22–42 d | 1280 ± 64.8 | 1344 ± 94.3 | 1345 ± 63.6 | 0.527 |
| Feed conversion (g/g), 22–42 d | 1.470 ± 0.080 | 1.421 ± 0.068 | 1.398 ± 0.017 | 0.395 |
| Feed intake (g), 1–42 d | 2745 ± 61.7 | 2761 ± 16.1 | 2735 ± 79.6 | 0.867 |
| Body weight gain (g), 1–42 d | 1923 ± 74.9 | 1991 ± 47.0 | 2020 ± 58.0 | 0.214 |
| Feed conversion (g/g), 1–42 d | 1.429 ± 0.038 a | 1.387 ± 0.026 ab | 1.354 ± 0.006 b | 0.040 |
| Compound | m/z Observed | tR/min | c/mg.g−1 | Standard Purity |
|---|---|---|---|---|
| 20-O-glucoginsenoside gRf 1 | 1007.5455 | 1.63 | ||
| Notoginsenoside nR1 | 977.5354 | 3.7 | 1.1 | ≥98% |
| Ginsenoside Re | 991.5505 | 4.35 | 143.4 | ≥98% |
| Ginsenoside Rg1 | 845.4929 | 4.44 | 35.2 | ≥98% |
| Ginsenoside Rf | 845.4925 | 8.06 | 1.4 | ≥98% |
| Ginsenoside F5 1 | 815.4811 | 9.15 | ||
| Ginsenoside Rb1 | 1153.6035 | 9.46 | 15.5 | ≥98% |
| Ginsenoside Rg2 | 829.4959 | 9.56 | 13.5 | ≥97% |
| Ginsenoside F3 1 | 815.4818 | 9.59 | ||
| Ginsenoside Rh1 | 683.4388 | 9.73 | 6.1 | ≥98% |
| Ginsenoside Rc | 1123.5927 | 10.12 | 40.2 | ≥98% |
| Ginsenoside Rb2 | 1123.5924 | 10.89 | 46.1 | ≥97% |
| Ginsenoside Rb3 1 | 1123.5922 | 11.09 | ||
| Ginsenoside F1 1 | 683.4383 | 11.68 | ||
| Ginsenoside Rd | 991.5494 | 12.46 | 75.6 | ≥98% |
| Ginsenoside F2 1 | 829.4966 | 18.19 | ||
| Ginsenoside Rg3 1 | 829.4968 | 20.61 | ||
| Ginsenoside CK | 667.4445 | 25.81 | 1.1 | |
| Ginsenoside Rh2 | 667.4445 | 26.72 | 1.9 |
| Genes | Forward Primer Sequence (5’–3’) | Reverse Primer Sequence (5’–3’) | Accession No. |
|---|---|---|---|
| Human | |||
| ACTB [49] | GCGGGAAATCGTGCGTGACATT | GATGGAGTTGAAGGTAGTTTCGTG | NM_001101 |
| B2M | TGAAGCTGACAGCATTCG | CAGACACATAGCAATTCAGG | NM_004048 |
| CLDN1 [50] | AACGCGGGGCTGCAGCTGTTG | GATGTTGTCGCCGGCATA | NM_021101 |
| CLDN3 | CACGCGAGAAGAAGTACA | TCTGTCCCTTAGACGTAGT | NM_001306 |
| GAPDH | TGGTATCGTGGAAGGACTCA | CAGTGAGCTTCCCGTTCAG | NM_002046 |
| HPRT1 | GACCCCACGAAGTGTTGGAT | ACTGGCGATGTCAATAGGACTC | NM_000194 |
| HSPA1A | GTGGAGGAGTTCAAGAGAA | GGTGATGGACGTGTAGAA | NM_005345 |
| HSPB1 | CTGGATGTCAACCACTTCGC | TATTTCCGCGTGAAGCACC | NM_001540 |
| HSPD1 | GAAATTGCCAATGCTCACCG | CTTGACTGCCACAACCTGAA | NM_002156 |
| OCLN [51] | GGACTCTACGTGGATCAGTATTTG | AATAATCATGAACCCCAGTACAATG | NM_002538 |
| RPL5 | TGGGCCAGAATGTTGCAGAT | AGGGACATTTTGGGACGGTT | NM_000969 |
| RPLP0 | TCTACAACCCTGAAGTGC | AAGGTGTAATCCGTCTCC | NM_001002 |
| C. elegans | |||
| act-1 | TGTTCCCATCCATTGTC | GCTCATTGTAGAAGGTGTG | NM_073418 |
| ama-1 | CTCCGTCGTTGACTGTAT | ATACCCATTCCTCGTCTTC | NM_068122 |
| pmp-3 | ATACGAAGCCACGGATAG | CTGTGTCAATGTCGTGAAG | NM_001269679 |
| hsp-16.2 | GAGAGATATGGCTCTGATGG | TCTCCTTGGATTGATAGCG | NM_071106 |
| hsp-1 | GCACGGAAAGGTAGAAATC | CGAACTTGCGTCCAATAAG | NM_070667 |
| daf-16 | GAATGGATGGTCCAGAATG | GATTCCTTCCTGGCTTTG | NM_001026423 |
| G. gallus | |||
| ACTB | ATGAAGCCCAGAGCAAAAGA | GGGGTGTTGAAGGTCTCAAA | NM_205518 |
| PGK1 | GGATAAGGTGGATGTGAAGG | AGAACTTGTCAGGCATGG | NM_204985 |
| SDHA [52] | CAGGGATGTAGTGTCTCGT | GGGAATAGGCTCCTTAGTG | NM_001277398 |
| YWHAZ | AGAGTCGTCTCAAGTATCG | CAACCTCAGCCAAGTAAC | NM_001031343 |
| CLDN1 | GTCATGGTATGGCAACAG, | GGTGGGTAGGATGTTTCA | NM_001013611 |
| HSPA2 | GGCTGGAGAGAAGAATGT | GTGCTTACGCTTGAACTC | NM_001006685 |
| OCLN | GCAGATGTCCAGCGGTTAC | GGTCCCAGTAGATGTTGGC | NM_205128 |
| Ingredients | Experimental Period | |||
|---|---|---|---|---|
| 1–21 d | 22–42 d | |||
| Maize | 327.00 | 18.25% | 329.66 | 18.37% |
| Wheat | 314.60 | 17.56% | 299.96 | 16.71% |
| Soybean meal (44% crude protein) | 250.90 | 14.00% | 265.00 | 14.77% |
| Soybean oil | 50.80 | 2.84% | 50.08 | 2.79% |
| Negative control premix or phytogenic premixes filled with a 1:1 mixture of wheat semolina and wheat bran to 5 g | 5.00 | 0.28% | 5.00 | 0.28% |
| Limestone | 14.80 | 0.83% | 14.60 | 0.81% |
| Monocalcium phosphate | 14.00 | 0.78% | 13.80 | 0.77% |
| Vitamin and mineral premix 1 | 12.00 | 0.67% | 12.00 | 0.67% |
| Titanium dioxide | 5.00 | 0.28% | 5.00 | 0.28% |
| l-Lysine HCl | 1.30 | 0.07% | 2.30 | 0.13% |
| DL-Methionine | 2.60 | 0.15% | 2.00 | 0.11% |
| Threonine | 2.00 | 0.11% | 0.60 | 0.03% |
| Nutrient levels | ||||
| MEN 2 | 12.48 | 0.70% | 12.56 | 0.70% |
| Crude protein | 220.8 | 12.32% | 200.80 | 11.19% |
| Lysine | 12.70 | 0.71% | 12.00 | 0.67% |
| Methionine | 5.90 | 0.33% | 5.00 | 0.28% |
| Methionine/cysteine | 9.70 | 0.54% | 8.50 | 0.47% |
| Tryptophane | 2.50 | 0.14% | 2.30 | 0.13% |
| Threonine | 8.60 | 0.48% | 8.10 | 0.45% |
| Crude fibre | 24.50 | 1.37% | 24.00 | 1.34% |
| Crude fat | 73.60 | 4.11% | 73.90 | 4.12% |
| Starch | 360.60 | 20.12% | 390.80 | 21.78% |
| Sugars | 42.90 | 2.39% | 39.50 | 2.20% |
| Calcium | 9.00 | 0.50% | 8.80 | 0.49% |
| Total phosphorus | 7.00 | 0.39% | 6.70 | 0.37% |
| Sodium | 1.60 | 0.09% | 1.70 | 0.09% |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Sandner, G.; Mueller, A.S.; Zhou, X.; Stadlbauer, V.; Schwarzinger, B.; Schwarzinger, C.; Wenzel, U.; Maenner, K.; van der Klis, J.D.; Hirtenlehner, S.; et al. Ginseng Extract Ameliorates the Negative Physiological Effects of Heat Stress by Supporting Heat Shock Response and Improving Intestinal Barrier Integrity: Evidence from Studies with Heat-Stressed Caco-2 Cells, C. elegans and Growing Broilers. Molecules 2020, 25, 835. https://doi.org/10.3390/molecules25040835
Sandner G, Mueller AS, Zhou X, Stadlbauer V, Schwarzinger B, Schwarzinger C, Wenzel U, Maenner K, van der Klis JD, Hirtenlehner S, et al. Ginseng Extract Ameliorates the Negative Physiological Effects of Heat Stress by Supporting Heat Shock Response and Improving Intestinal Barrier Integrity: Evidence from Studies with Heat-Stressed Caco-2 Cells, C. elegans and Growing Broilers. Molecules. 2020; 25(4):835. https://doi.org/10.3390/molecules25040835
Chicago/Turabian StyleSandner, Georg, Andreas S. Mueller, Xiaodan Zhou, Verena Stadlbauer, Bettina Schwarzinger, Clemens Schwarzinger, Uwe Wenzel, Klaus Maenner, Jan Dirk van der Klis, Stefan Hirtenlehner, and et al. 2020. "Ginseng Extract Ameliorates the Negative Physiological Effects of Heat Stress by Supporting Heat Shock Response and Improving Intestinal Barrier Integrity: Evidence from Studies with Heat-Stressed Caco-2 Cells, C. elegans and Growing Broilers" Molecules 25, no. 4: 835. https://doi.org/10.3390/molecules25040835