Marine Microorganism-Derived Macrolactins Inhibit Inflammatory Mediator Effects in LPS-Induced Macrophage and Microglial Cells by Regulating BACH1 and HO-1/Nrf2 Signals through Inhibition of TLR4 Activation


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Result
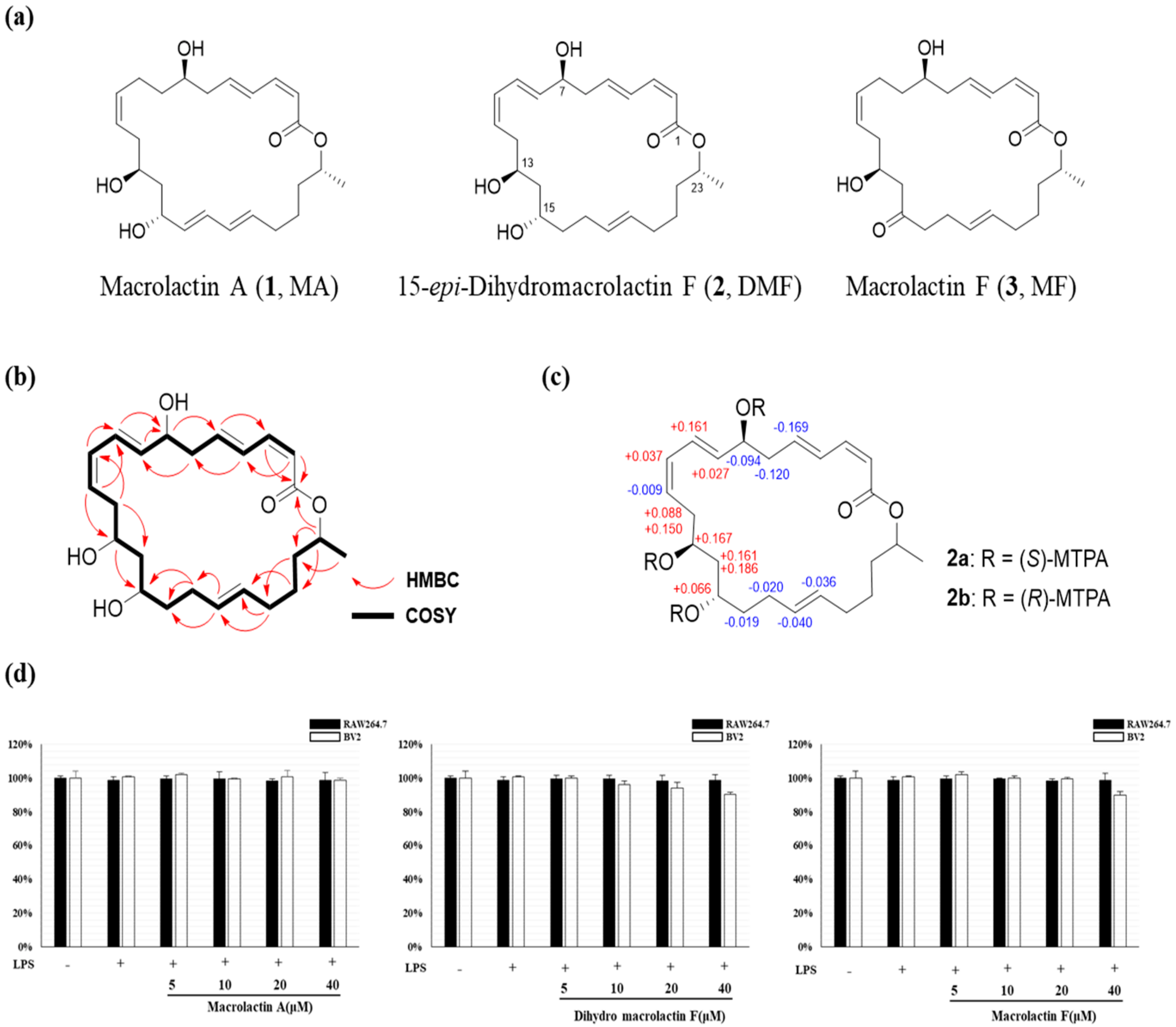
2.1. Structure Elucidation of DMF, MA and MF
2.2. Effect of MA, DMF and MF on Cell Viability of RAW264.7 and BV2 Cells
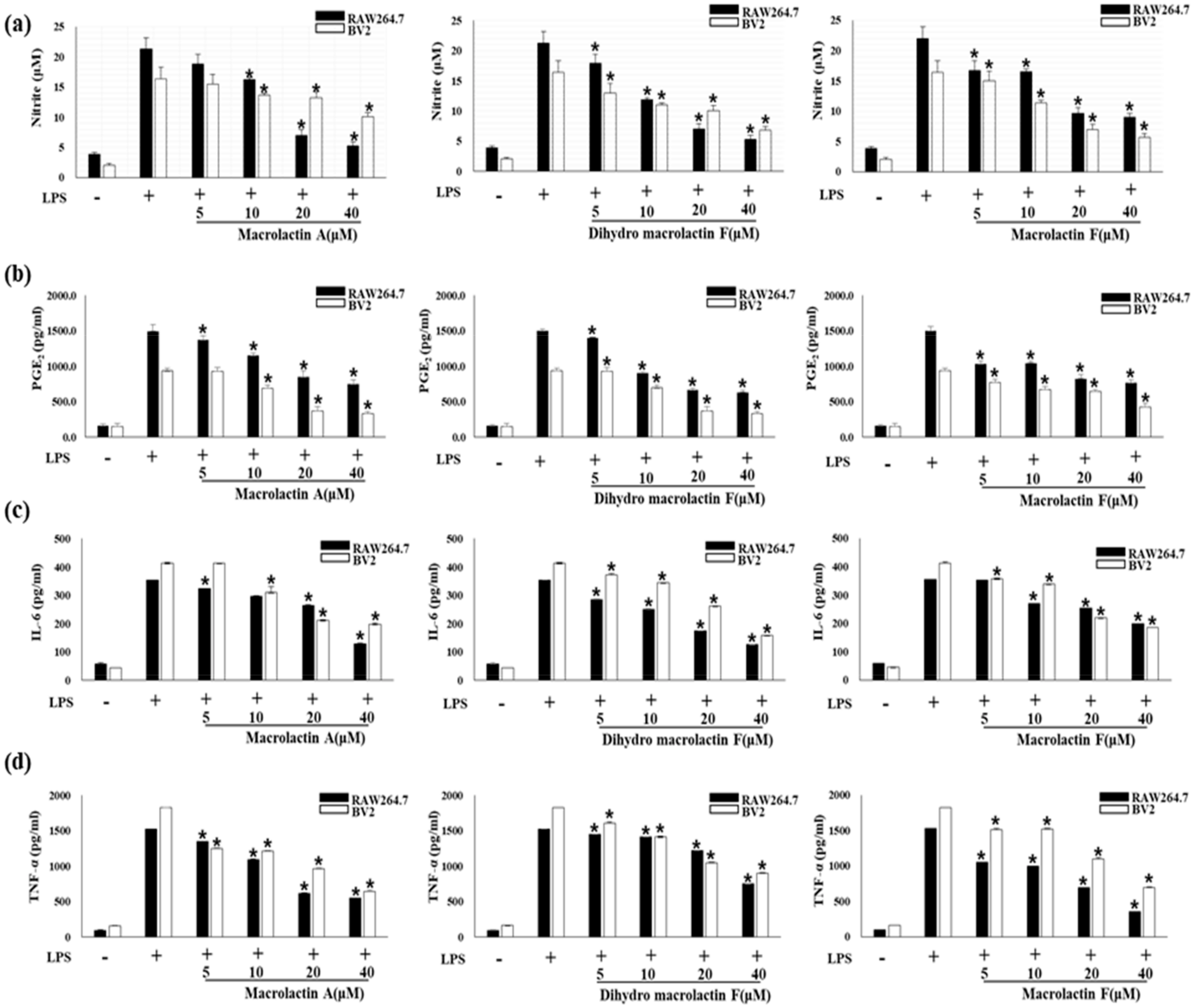
2.3. Effect of MA, DMF and MF on NO Production and Pro-Inflammatory Cytokines in LPS-Stimulated RAW264.7 and BV2 Cells
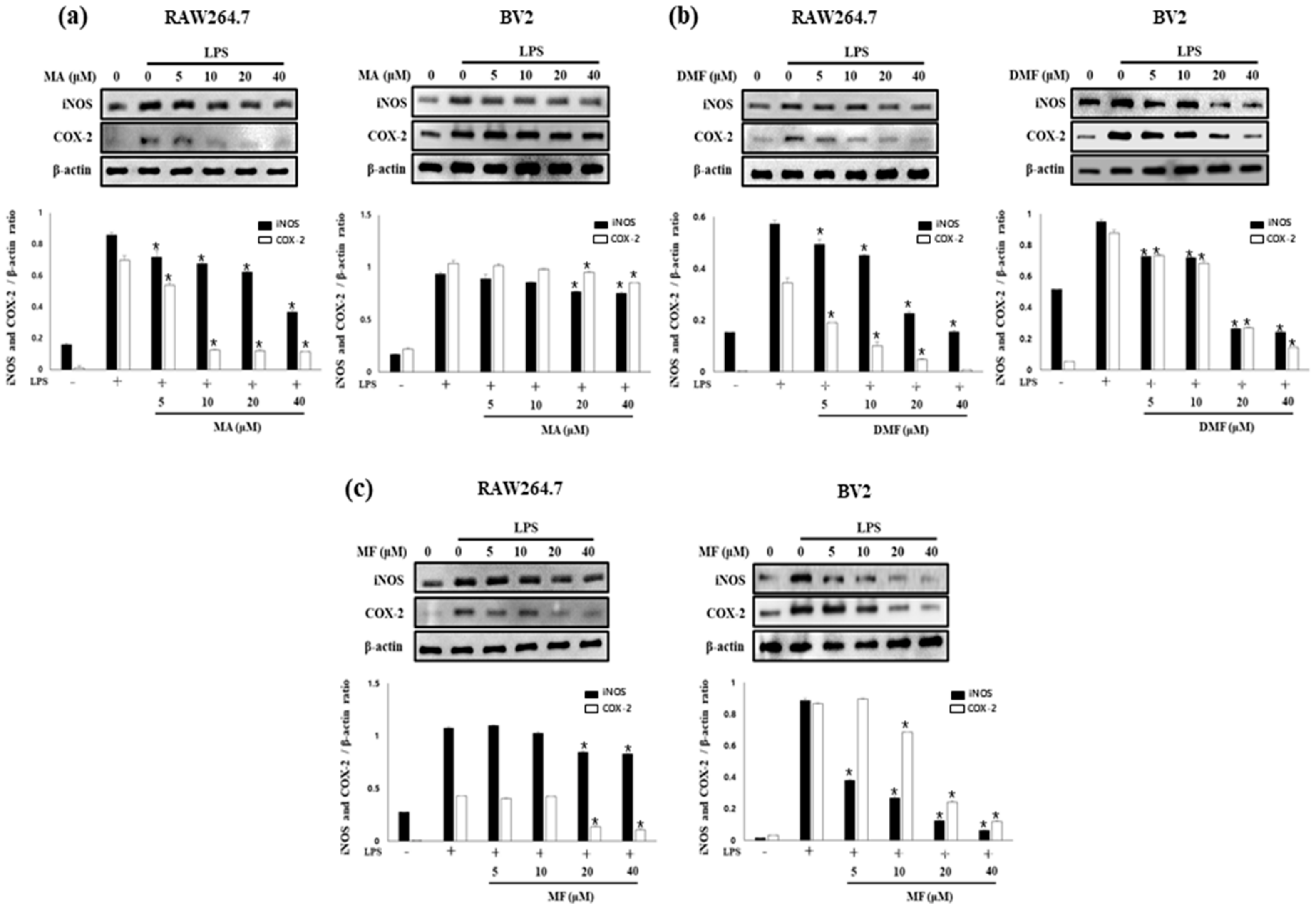
2.4. Effect of MA, DMF and MF on the Expression of iNOS and COX-2 in LPS-Stimulated RAW264.7 and BV2 Cells
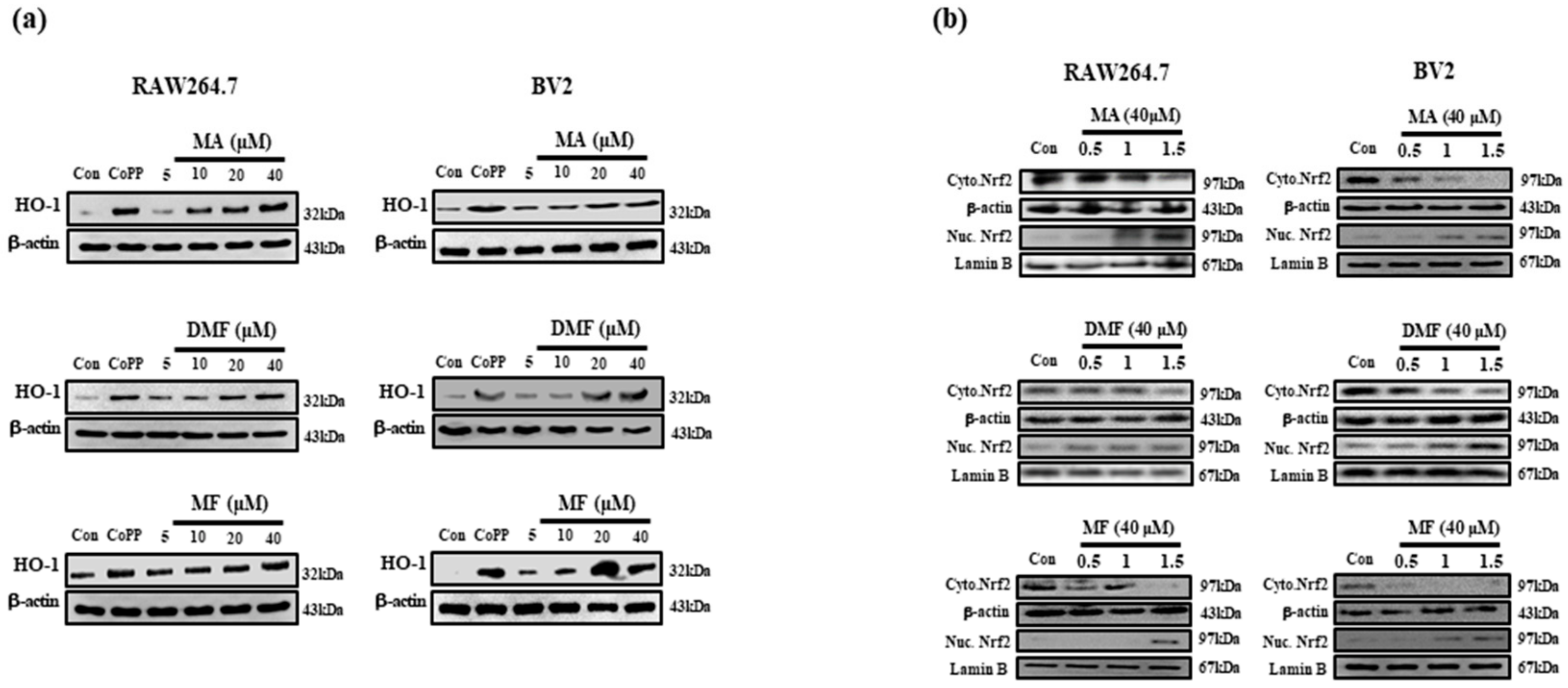
2.5. MA, DMF and MF Promote Upregulation of HO-1 Protein Expression and Nucleus Translocation of Nrf2 Protein and Activated Nrf2 Pathway.
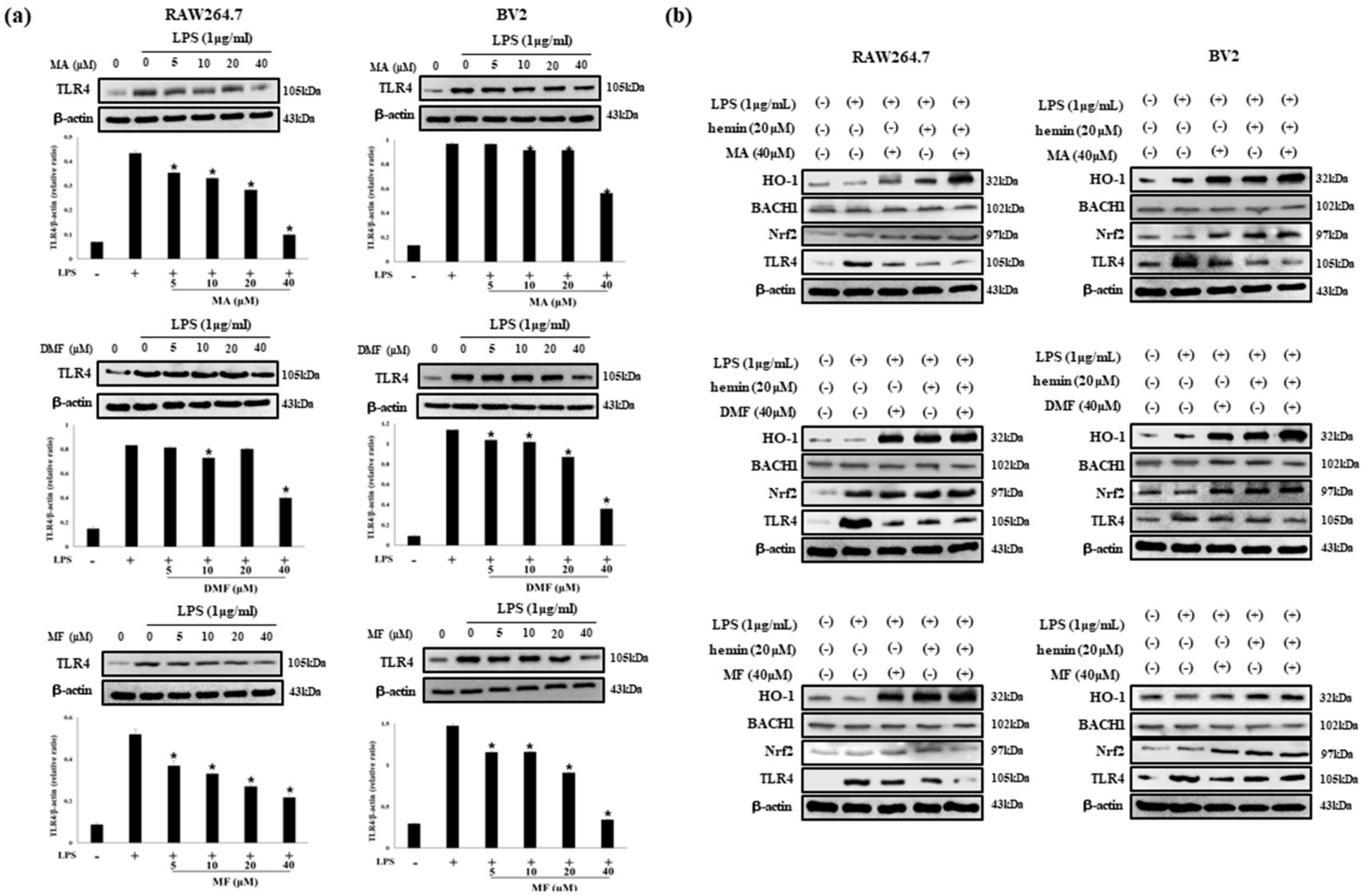
2.6. MA, DMF and MF Inhibit TLR4 Expression by LPS and Regulate BACH1 and HO-1/Nrf2 Signals.
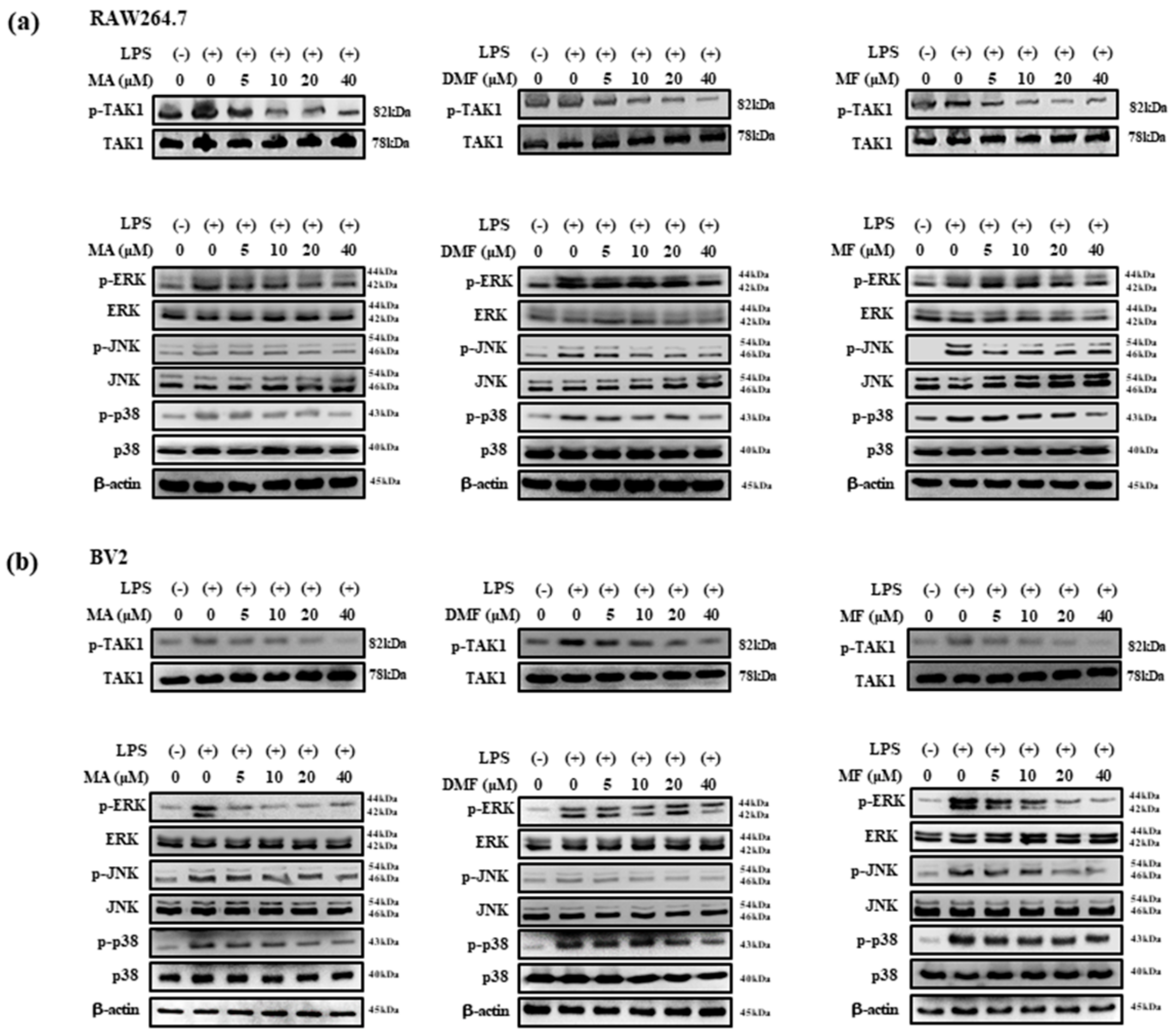
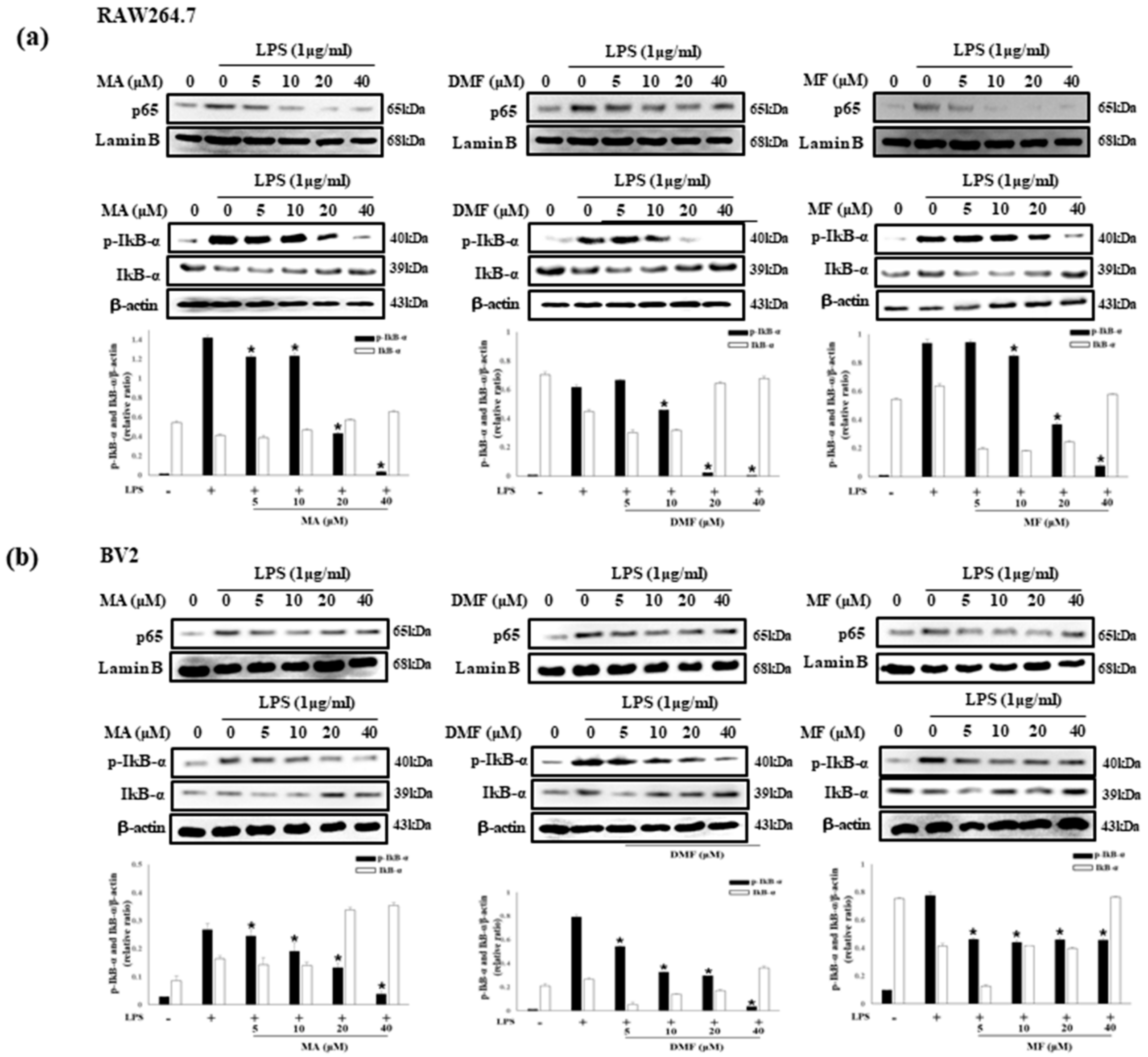
2.7. Effect of MA, DMF and MF on LPS-Induced Phosphorylation of TAK1, MAPK and NF-κB
3. Discussion
4. Summary and Conclusion
5. Materials and Methods
5.1. General Experimental Procedures
5.2. Isolation of Bacterial Strain and Taxonomic Identification
5.3. Culture of Bacterial Strain
5.4. Extraction and Isolation
5.5. Preparation of (R/S)-tris-MTPA Esters of Compound 2
5.6. Materials and Reagents
5.7. Cell Culture
5.8. Cell Viability Assay
5.9. NO Production and Assays
5.10. Cytokine Analysis
5.11. Western Blot Analysis
5.12. Cytosolic and Nuclear Protein Extraction
5.13. Statistical Analysis
Supplementary Materials
Author Contributions
Funding
Conflicts of Interest
References
- Peng, S. Salidroside regulates inflammatory response in RAW 264.7 macrophages via TLR4/TAK1 and ameliorates inflammation in alcohol binge drinking-induced liver injury. Molecules 2016, 21, 1490. [Google Scholar]
- Jayson, M.I. Vascular damage, fibrosis, and chronic inflammation in mechanical back pain problems. Semin. Arthritis Rheum. 1989, 18, 73–76. [Google Scholar] [CrossRef]
- Linde, A.; Blecha, F.; Mosier, D.; Melgarejo, T. Innate immunity and inflammation—New frontiers in comparative cardiovascular pathology. Cardiovasc. Res. 2007, 73, 26–36. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chayanut, K.; Sasimol, U.; Thanyaluck, P.; Peraphan, P.; Prachya, K. Cyanidin-3–O-glucoside protects PC12 cells against neuronal apoptosis mediated by LPS-stimulated BV2 microglial activation. Neurotox. Res. 2019, 1–15. [Google Scholar]
- Hsieh, Y.H.; Deng, J.S.; Pan, H.P.; Liao, J.C.; Huang, S.S.; Huang, G.J. Sclareol ameliorate lipopolysaccharide-induced acute lung injury through inhibition of MAPK and induction of HO–1 signaling. Int. Immunopharmacol. 2016, 44, 16–25. [Google Scholar] [CrossRef]
- Liu, B.; Hong, J.-S. Role of Microglia in Inflammation-Mediated Neurodegenerative Diseases: Mechanisms and Strategies for Therapeutic Intervention. J. Pharmacol. Exp. Ther. 2003, 304, 1–7. [Google Scholar] [CrossRef] [Green Version]
- Lin, W.C.; Deng, J.S.; Huang, S.S.; Wu, S.H.; Chen, C.C.; Lin, W.R.; Lin, H.Y.; Huang, G.J. Anti-Inflammatory Activity of Sanghuangporus sanghuang Mycelium. Int. J. Mol. Sci. 2017, 18, 347. [Google Scholar] [CrossRef] [Green Version]
- Billack, B. Macrophage Activation: Role of Toll-like Receptors, Nitric Oxide, and Nuclear Factor kappa B. Am. J. Pharm. Educ. 2006, 70, 102. [Google Scholar] [CrossRef]
- Maines, M.D. Heme oxygenase: Function, multiplicity, regulatory mechanisms, and clinical applications. FASEB J. 1988, 2, 2557–2568. [Google Scholar] [CrossRef] [Green Version]
- Tenhunen, R.; Marver, H.S.; Schmid, R. The enzymatic conversion of heme to bilirubin by microsomal heme oxygenase. Proc. Natl. Acad. Sci. USA 1968, 61, 748–755. [Google Scholar] [CrossRef] [Green Version]
- Gozzelino, R.; Jeney, V.; Soares, M.P. Mechanisms of Cell Protection by Heme Oxygenase-1. Annu. Rev. Pharmacol. Toxicol. 2010, 50, 323–354. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ryter, S.W.; Alam, J.; Choi, A.M.K. Heme Oxygenase-1/Carbon Monoxide: From Basic Science to Therapeutic Applications. Physiol. Rev. 2006, 86, 583–650. [Google Scholar] [CrossRef] [PubMed]
- Paine, A.; Eiz-Vesper, B.; Blasczyk, R.; Immenschuh, S. Signaling to heme oxygenase-1 and its anti-inflammatory therapeutic potential. Biochem. Pharmacol. 2010, 80, 1895–1903. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Miyazaki, T.; Kirino, Y.; Takeno, M.; Samukawa, S.; Hama, M.; Tanaka, M.; Yamaji, S.; Ueda, A.; Tomita, N.; Fujita, H.; et al. Expression of heme oxygenase-1 in human leukemic cells and its regulation by transcriptional repressor Bach1. Cancer Sci. 2010, 101, 1409–1416. [Google Scholar] [CrossRef] [PubMed]
- Rateb, M.E.; Ebel, R. Secondary metabolites of fungi from marine habitats. Nat. Prod. Rep. 2011, 28, 290. [Google Scholar] [CrossRef]
- Blunt, J.W.; Copp, B.R.; Keyzers, R.A.; Munro, M.H.G.; Prinsep, M.R. Marine natural products. Nat. Prod. Rep. 2014, 31, 160–258. [Google Scholar] [CrossRef] [Green Version]
- Look, S.A.; Fenical, W.; Jacobs, R.S.; Clardy, J. The pseudopterosins: Anti-inflammatory and analgesic natural products from the sea whip Pseudopterogorgia elisabethae. Proc. Natl. Acad. Sci. USA 1986, 83, 6238–6240. [Google Scholar] [CrossRef] [Green Version]
- Newman, D.J.; Cragg, G.M. Marine Natural Products and Related Compounds in Clinical and Advanced Preclinical Trials. J. Nat. Prod. 2004, 67, 1216–1238. [Google Scholar] [CrossRef]
- Mitchell, S.S.; Nicholson, B.; Teisan, S.; Lam, K.S.; Potts, B.C.M. Aureoverticillactam, a Novel 22-Atom Macrocyclic Lactam from the Marine ActinomyceteStreptomyces aureoverticillatus. J. Nat. Prod. 2004, 67, 1400–1402. [Google Scholar] [CrossRef]
- Nicolaou, K.C.; Nold, A.L.; Milburn, R.R.; Schindler, C.S. Total synthesis of marinomycins A–C. Angew. Chem. Int. Ed. 2006, 45, 6527–6532. [Google Scholar] [CrossRef]
- Kwon, H.C.; Kauffman, C.A.; Jensen, P.R.; Fenical, W. Marinomycins A–D, antitumor-antibiotics of a new structure class from a marine actinomycete of the recently discovered genus “Marinispora”. J. Am. Chem. Soc. 2006, 128, 1622–1632. [Google Scholar] [CrossRef] [PubMed]
- Sitachitta, N.; Gerwick, W.H. Grenadadiene and Grenadamide, Cyclopropyl-Containing Fatty Acid Metabolites from the Marine CyanobacteriumLyngbya majuscula. J. Nat. Prod. 1998, 61, 681–684. [Google Scholar] [CrossRef] [PubMed]
- Singh, I.P.; Milligan, E.; Gerwick, K.; Tanikolide, W.H. A toxic and antifungal lactone from the marine cyanobacterium Lyngbya majuscula. J. Nat. Prod. 1999, 62, 1333–1335. [Google Scholar] [CrossRef] [PubMed]
- Gutirrez, M. Structural and synthetic investigations of tanikolide dimer, a SIRT2 selective inhibitor, and tanikolide seco-acid from the madagascar marine cyanobacterium Lyngbya majuscula. J. Org. Chem. 2009, 74, 5267–5275. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kim, G.J.; Li, X.; Kim, S.-H.; Yang, I.; Hahn, D.; Chin, J.; Nam, S.-J.; Nam, J.-W.; Nam, D.H.; Oh, D.-C.; et al. Seongsanamides A–D: Antiallergic Bicyclic Peptides from Bacillus safensis KCTC 12796BP. Org. Lett. 2018, 20, 7539–7543. [Google Scholar] [CrossRef] [PubMed]
- Li, D.; Carr, G.; Zhang, Y.; Williams, D.E.; Amlani, A.; Bottriell, H.; Mui, A.L.; Andersen, R.J. Turnagainolides A and B, cyclic depsipeptides produced in culture by a Bacillus sp.: Isolation, structure elucidation, and synthesis. J. Nat. Prod. 2011, 74, 1093–1099. [Google Scholar] [CrossRef]
- Gustafson, K.; Roman, M.; Fenical, W. The macrolactins, a novel class of antiviral and cytotoxic macrolides from a deep-sea marine bacterium. J. Am. Chem. Soc. 1989, 111, 7519–7524. [Google Scholar] [CrossRef]
- Zheng, C.J.; Lee, S.K.; Lee, C.H.; Kim, W.G. Macrolactins O-R, glycosylated 24-membered lactones from Bacillus sp. AH159-1. J. Nat. Prod. 2007, 70, 1632–1635. [Google Scholar] [CrossRef]
- Xue, C.; Tian, L.; Xu, M.; Deng, Z.; Lin, W. A New 24-membered Lactone and a New Polyene δ-Lactone from the Marine Bacterium Bacillus marinus. J. Antibiot. 2008, 61, 668–674. [Google Scholar] [CrossRef] [Green Version]
- Mondol, M. New antimicrobial compounds from a marine-derived Bacillus sp. J. Antibiot. 2013, 66, 89–95. [Google Scholar] [CrossRef]
- Bae, S.H.; Kwon, M.J.; Park, J.B.; Kim, D.; Kim, N.-H.; Kang, J.-S.; Kim, C.-G.; Oh, E.; Bae, S.K. Metabolic Drug-Drug Interaction Potential of Macrolactin A and 7-O-Succinyl Macrolactin A Assessed by Evaluating Cytochrome P450 Inhibition and Induction and UDP-Glucuronosyltransferase Inhibition In Vitro. Antimicrob. Agents Chemother. 2014, 58, 5036–5046. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Regmi, S.C.; Park, S.Y.; Kim, S.J.; Banskota, S.; Shah, S.; Kim, N.-H.; Kim, J.-A. The Anti-Tumor Activity of Succinyl Macrolactin A Is Mediated through the β-Catenin Destruction Complex via the Suppression of Tankyrase and PI3K/Akt. PLoS ONE 2015, 10, e0141753. [Google Scholar] [CrossRef] [PubMed]
- Kang, Y.; Regmi, S.C.; Kim, M.Y.; Banskota, S.; Gautam, J.; Kim, D.H.; Kim, J.A. Anti-angiogenic activity of macrolactin A and its succinyl derivative is mediated through inhibition of class I PI3K activity and its signaling. Arch. Pharm. Res. 2015, 38, 249–260. [Google Scholar] [CrossRef] [PubMed]
- Park, S.; Regmi, S.C.; Park, S.-Y.; Lee, E.K.; Chang, J.-H.; Ku, S.K.; Kim, N.-H.; Kim, J.-A. Protective effect of 7-O-succinyl macrolactin A against intestinal inflammation is mediated through PI3-kinase/Akt/mTOR and NF-κB signaling pathways. Eur. J. Pharmacol. 2014, 735, 184–192. [Google Scholar] [CrossRef] [PubMed]
- Li, L.; Sapkota, M.; Gao, M.; Choi, H.; Soh, Y. Macrolactin F inhibits RANKL-mediated osteoclastogenesis by suppressing Akt, MAPK and NFATc1 pathways and promotes osteoblastogenesis through a BMP-2/smad/Akt/Runx2 signaling pathway. Eur. J. Pharmacol. 2017, 815, 202–209. [Google Scholar] [CrossRef]
- Rychnovsky, S.D.; Skalitzky, D.J.; Pathirana, C.; Jensen, P.R.; Fenical, W. Stereochemistry of the macrolactins. J. Am. Chem. Soc. 1992, 114, 671–677. [Google Scholar] [CrossRef]
- Alam, J.; Stewart, D.; Touchard, C.; Boinapally, S.; Choi, A.M.K.; Cook, J.L. Nrf2, a Cap’n’Collar Transcription Factor, Regulates Induction of the Heme Oxygenase-1 Gene. J. Boil. Chem. 1999, 274, 26071–26078. [Google Scholar] [CrossRef] [Green Version]
- Ci, X. Schisantherin a exhibits anti-inflammatory properties by down-regulating NF-κB and MAPK signaling pathways in lipopolysaccharide-treated RAW264.7 cells. Inflammation 2010, 33, 126–136. [Google Scholar] [CrossRef]
- Kawai, T.; Akira, S. Signaling to NF-κB by Toll-like receptors. Trends Mol. Med. 2007, 13, 460–469. [Google Scholar] [CrossRef]
- Smith, J.A.; Das, A.; Ray, S.K.; Banik, N.L. Role of pro-inflammatory cytokines released from microglia in neurodegenerative diseases. Brain Res. Bull. 2012, 87, 10–20. [Google Scholar] [CrossRef]
- Michlewska, S.M.C.F.; Rossi, A. Macrophage phagocytosis of apoptotic neutrophils is critically regulated by the opposing actions of pro-inflammatory and anti-inflammatory agents: Key role for TNF-alpha. FASEB. J. 2009, 23, 844–854. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Abraham, N.G.; Lavrovsky, Y.; Schwartzman, M.L.; Stoltz, R.A.; Levere, R.D.; Gerritsen, M.E.; Shibahara, S.; Kappas, A. Transfection of the human heme oxygenase gene into rabbit coronary microvessel endothelial cells: Protective effect against heme and hemoglobin toxicity. Proc. Natl. Acad. Sci. USA 1995, 92, 6798–6802. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Beutler, B.; Rietschel, E.T. Innate immune sensing and its roots: The story of endotoxin. Nat. Rev. Immunol. 2003, 3, 169–176. [Google Scholar] [CrossRef]
- Hichor, M.; Sundaram, V.K.; Eid, S.A.; Abdel-Rassoul, R.; Petit, P.X.; Borderie, D.; Bastin, J.; Eid, A.A.; Manuel, M.; Grenier, J.; et al. Liver X Receptor exerts a protective effect against the oxidative stress in the peripheral nerve. Sci. Rep. 2018, 8, 2524. [Google Scholar] [CrossRef] [Green Version]
- Cho, H.-Y.; Reddy, S.P.; Kleeberger, S.R. Nrf2 Defends the Lung from Oxidative Stress. Antioxid. Redox Signal. 2006, 8, 76–87. [Google Scholar] [CrossRef]
- Ogawa, K.; Sun, J.; Taketani, S.; Nakajima, O.; Nishitani, C.; Sassa, S.; Hayashi, N.; Yamamoto, M.; Shibahara, S.; Fujita, H.; et al. Heme mediates derepression of Maf recognition element through direct binding to transcription repressor Bach1. EMBO J. 2001, 20, 2835–2843. [Google Scholar] [CrossRef] [Green Version]
- Yu, R.; Chen, C.; Mo, Y.-Y.; Hebbar, V.; Owuor, E.D.; Tan, T.-H.; Kong, A.-N.T. Activation of Mitogen-activated Protein Kinase Pathways Induces Antioxidant Response Element-mediated Gene Expression via a Nrf2-dependent Mechanism. J. Boil. Chem. 2000, 275, 39907–39913. [Google Scholar] [CrossRef] [Green Version]
- Liu, C.; Tang, X.; Zhang, W.; Li, G.; Chen, Y.; Guo, A.; Hu, C. 6-Bromoindirubin-3-Oxime Suppresses LPS-Induced Inflammation via Inhibition of the TLR4/NF-κB and TLR4/MAPK Signaling Pathways. Inflammation 2019, 42, 2192–2204. [Google Scholar] [CrossRef]
Sample Availability: Samples of the compounds are not available from the authors. |







© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Kim, E.-N.; Gao, M.; Choi, H.; Jeong, G.-S. Marine Microorganism-Derived Macrolactins Inhibit Inflammatory Mediator Effects in LPS-Induced Macrophage and Microglial Cells by Regulating BACH1 and HO-1/Nrf2 Signals through Inhibition of TLR4 Activation. Molecules 2020, 25, 656. https://doi.org/10.3390/molecules25030656
Kim E-N, Gao M, Choi H, Jeong G-S. Marine Microorganism-Derived Macrolactins Inhibit Inflammatory Mediator Effects in LPS-Induced Macrophage and Microglial Cells by Regulating BACH1 and HO-1/Nrf2 Signals through Inhibition of TLR4 Activation. Molecules. 2020; 25(3):656. https://doi.org/10.3390/molecules25030656
Chicago/Turabian StyleKim, Eun-Nam, Ming Gao, Hyukjae Choi, and Gil-Saeng Jeong. 2020. "Marine Microorganism-Derived Macrolactins Inhibit Inflammatory Mediator Effects in LPS-Induced Macrophage and Microglial Cells by Regulating BACH1 and HO-1/Nrf2 Signals through Inhibition of TLR4 Activation" Molecules 25, no. 3: 656. https://doi.org/10.3390/molecules25030656