
Quercetin Exerts Age-Dependent Beneficial Effects on Blood Pressure and Vascular Function, But Is Inefficient in Preventing Myocardial Ischemia-Reperfusion Injury in Zucker Diabetic Fatty Rats

 , , , , ,
, , , , , 
Abstract
:1. Introduction
2. Results
2.1. Effect of QCT on Biometric Parameters and Biochemical Characteristics of Rats
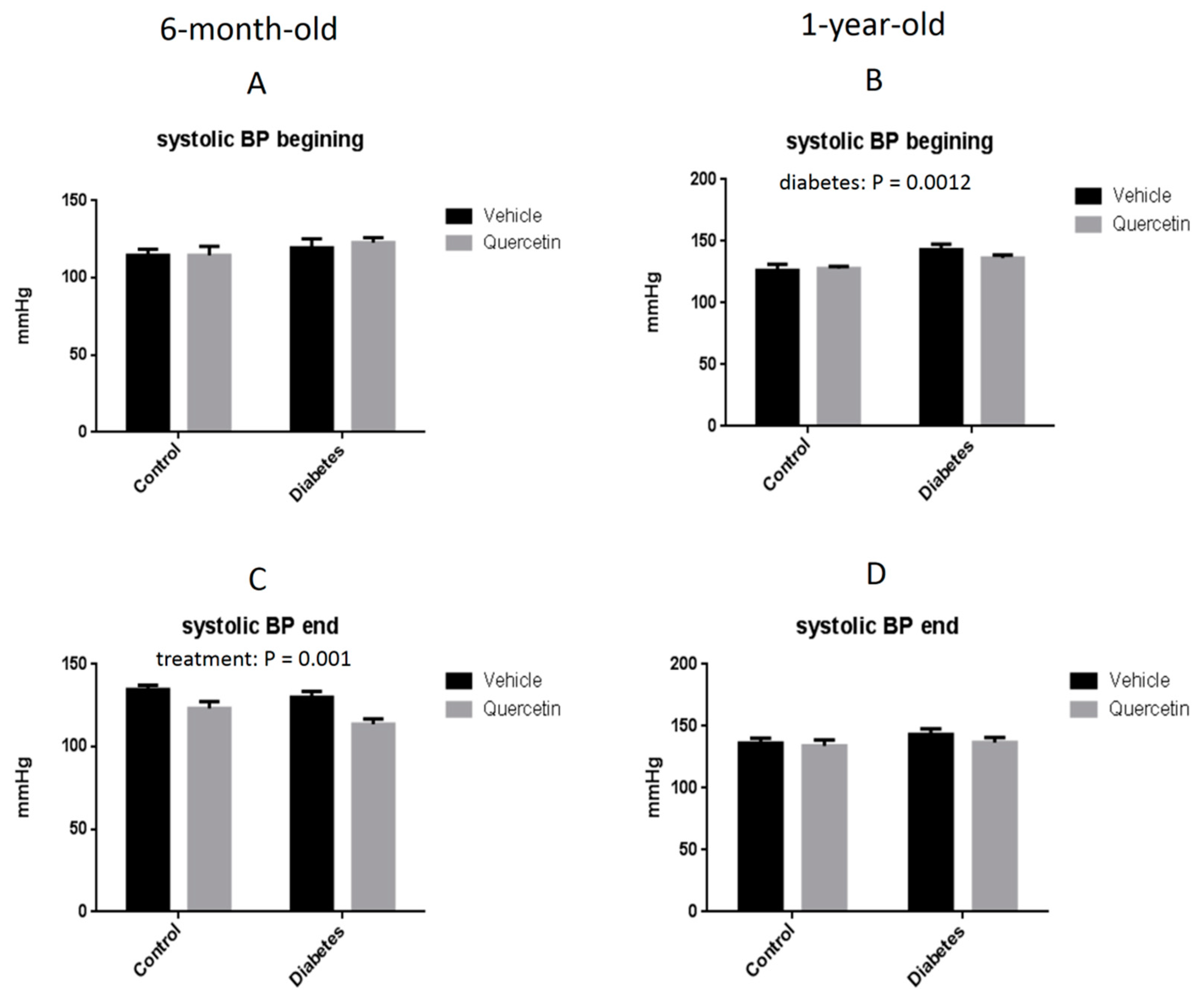
2.2. Effect of QCT on Blood Pressure
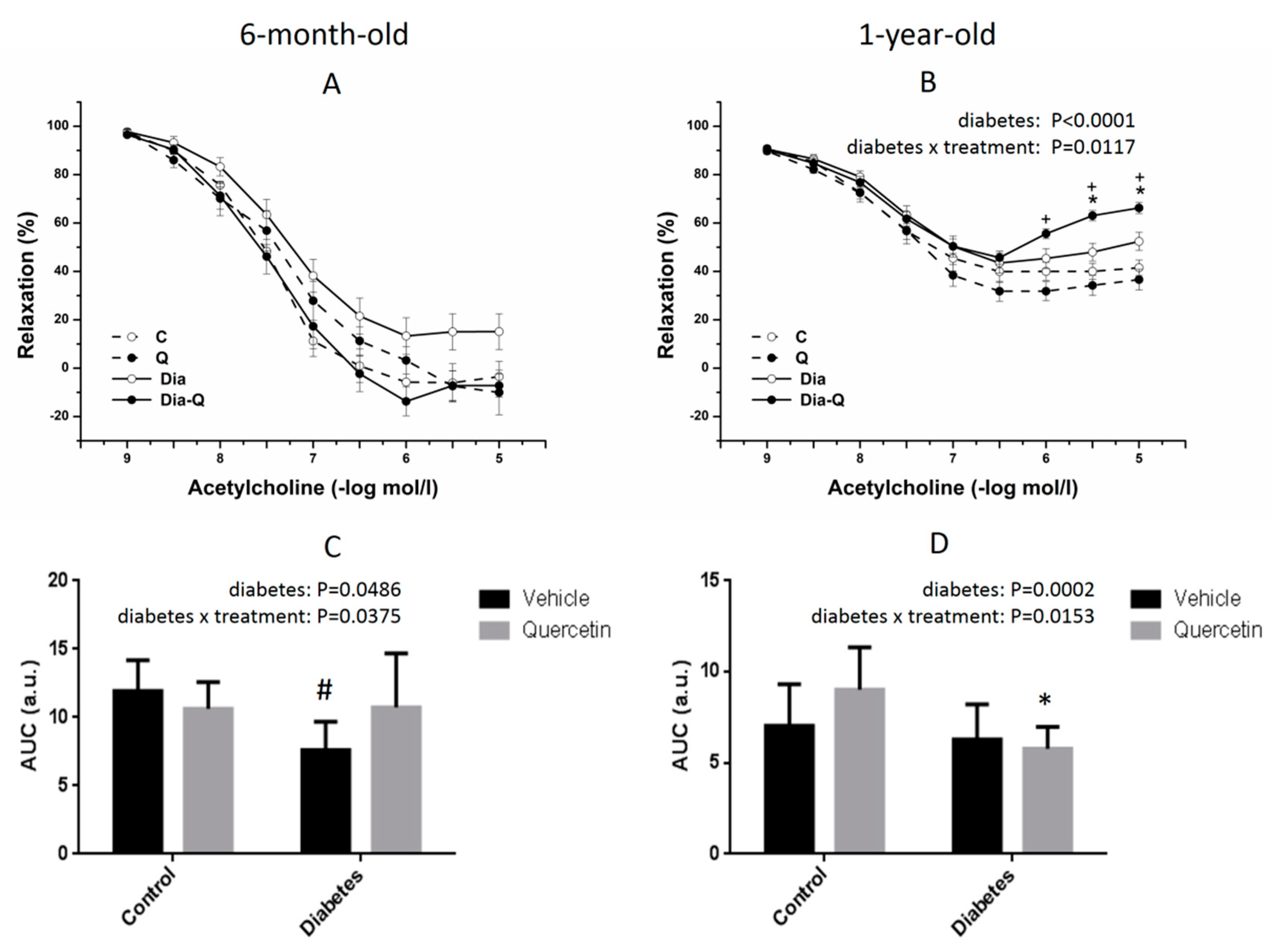
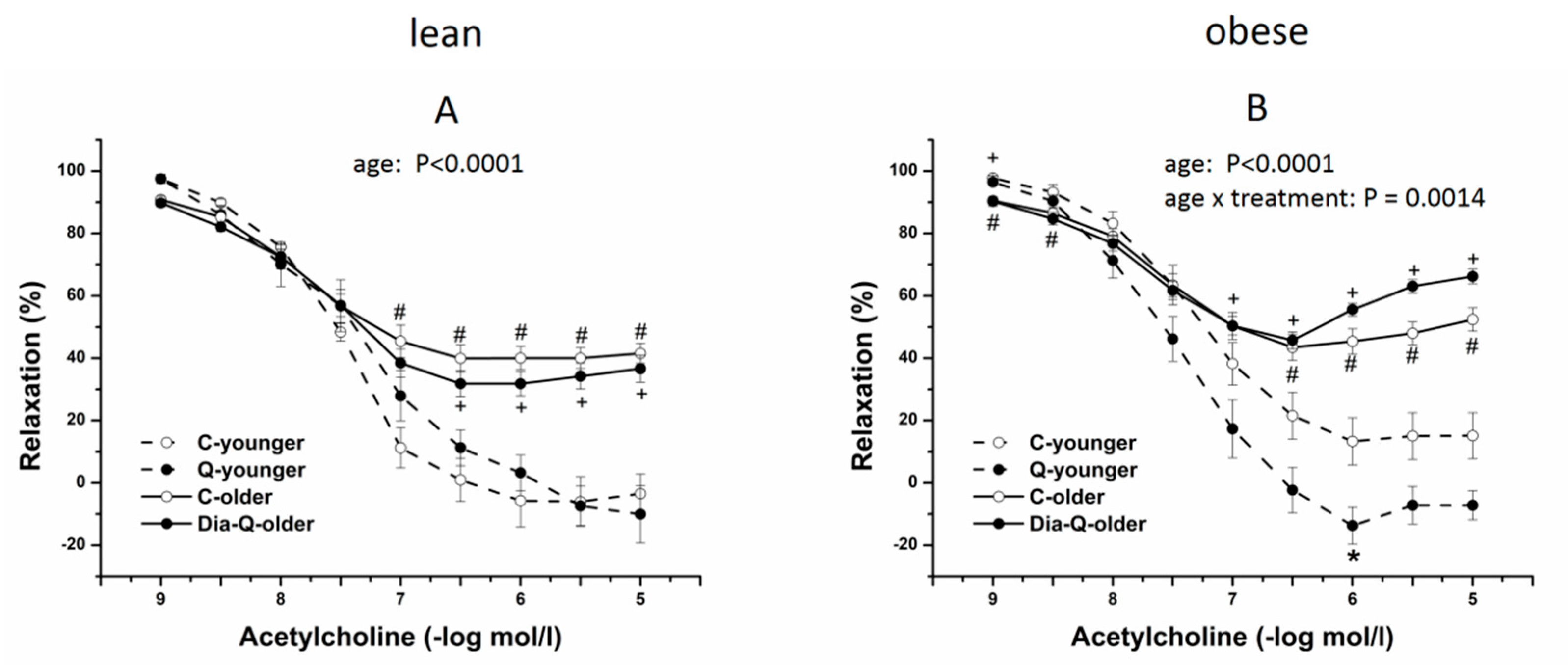
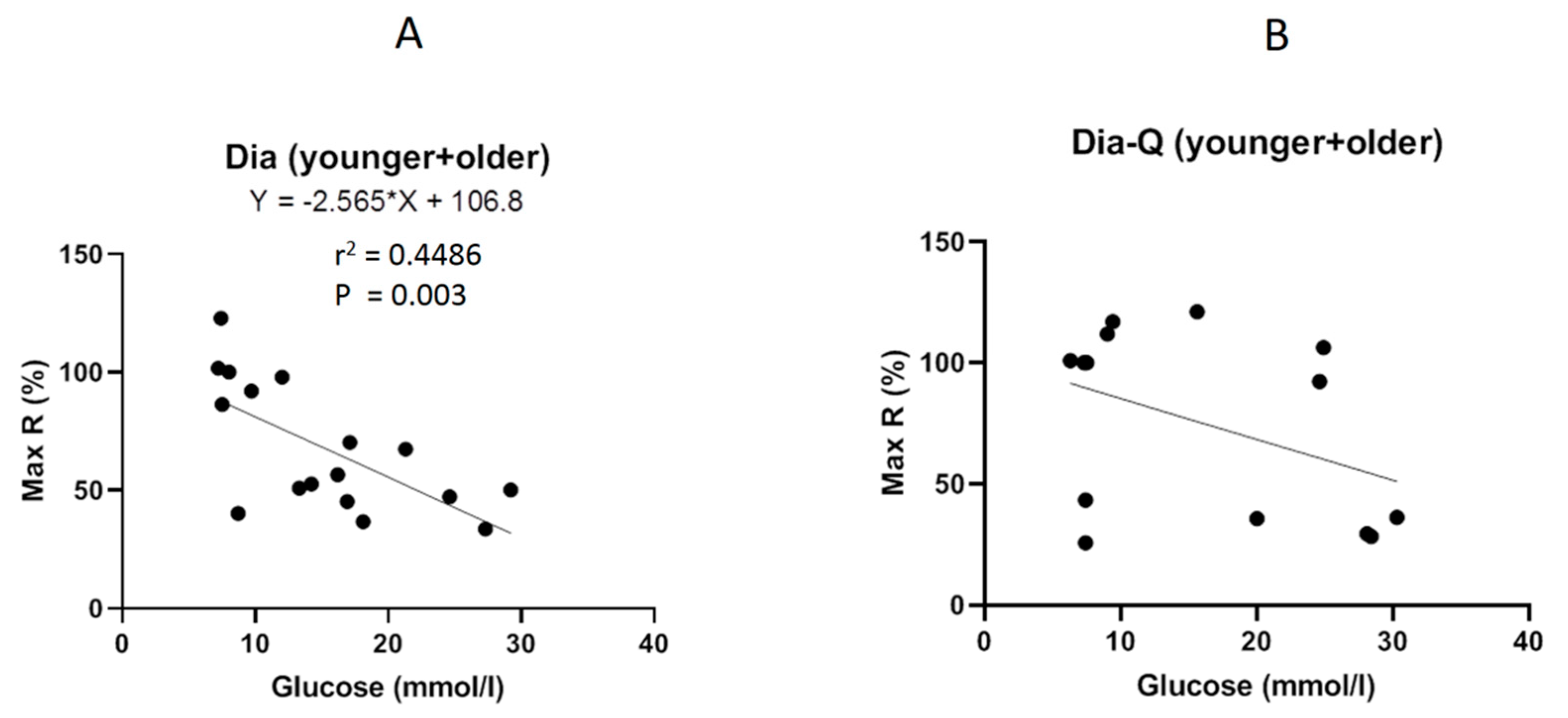
2.3. Effect of QCT on Vascular Reactivity of Isolated Thoracic Aortas
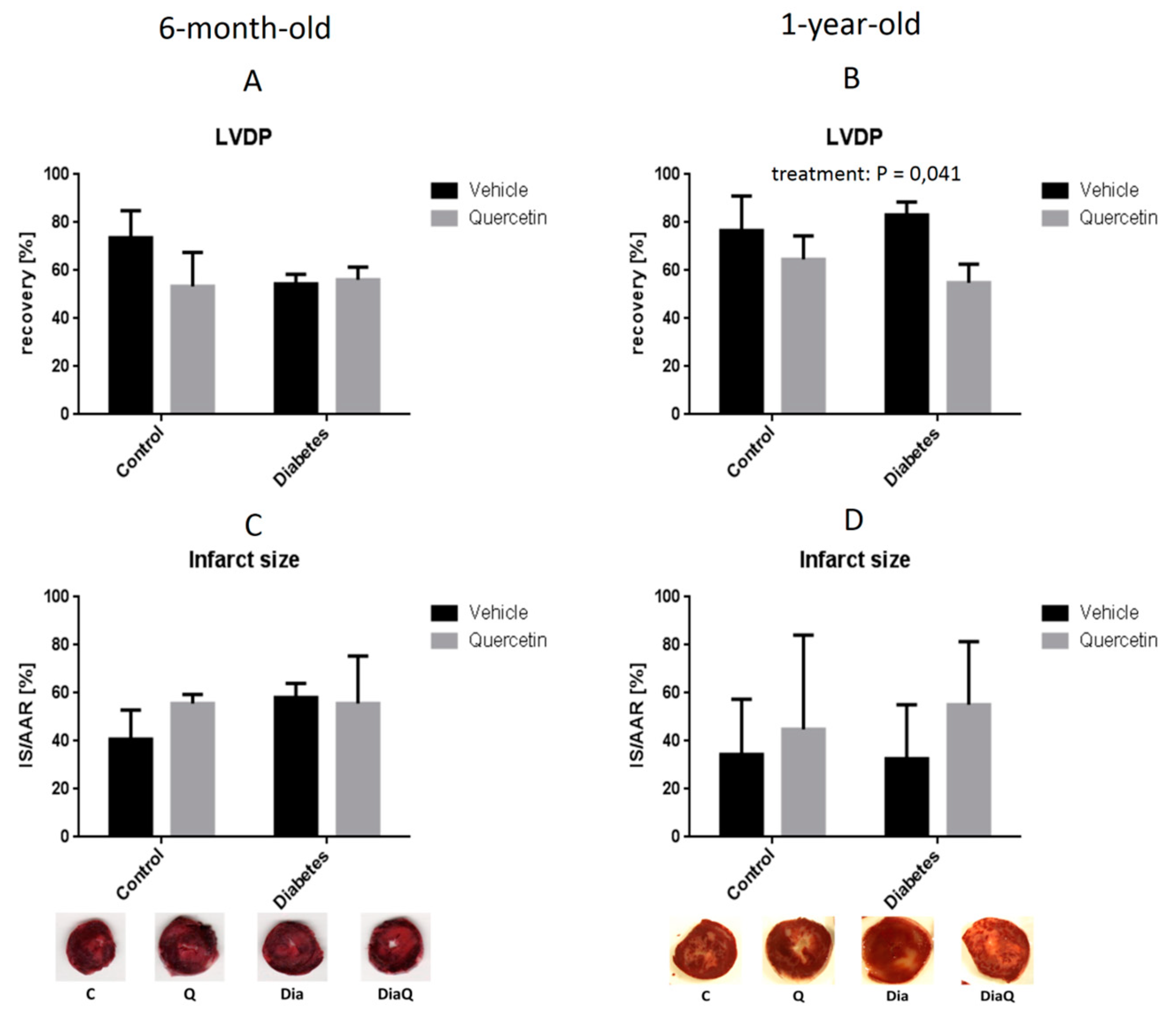
2.4. Effect of QCT on Post-Ischemic Recovery and Infarct Size in Isolated Hearts
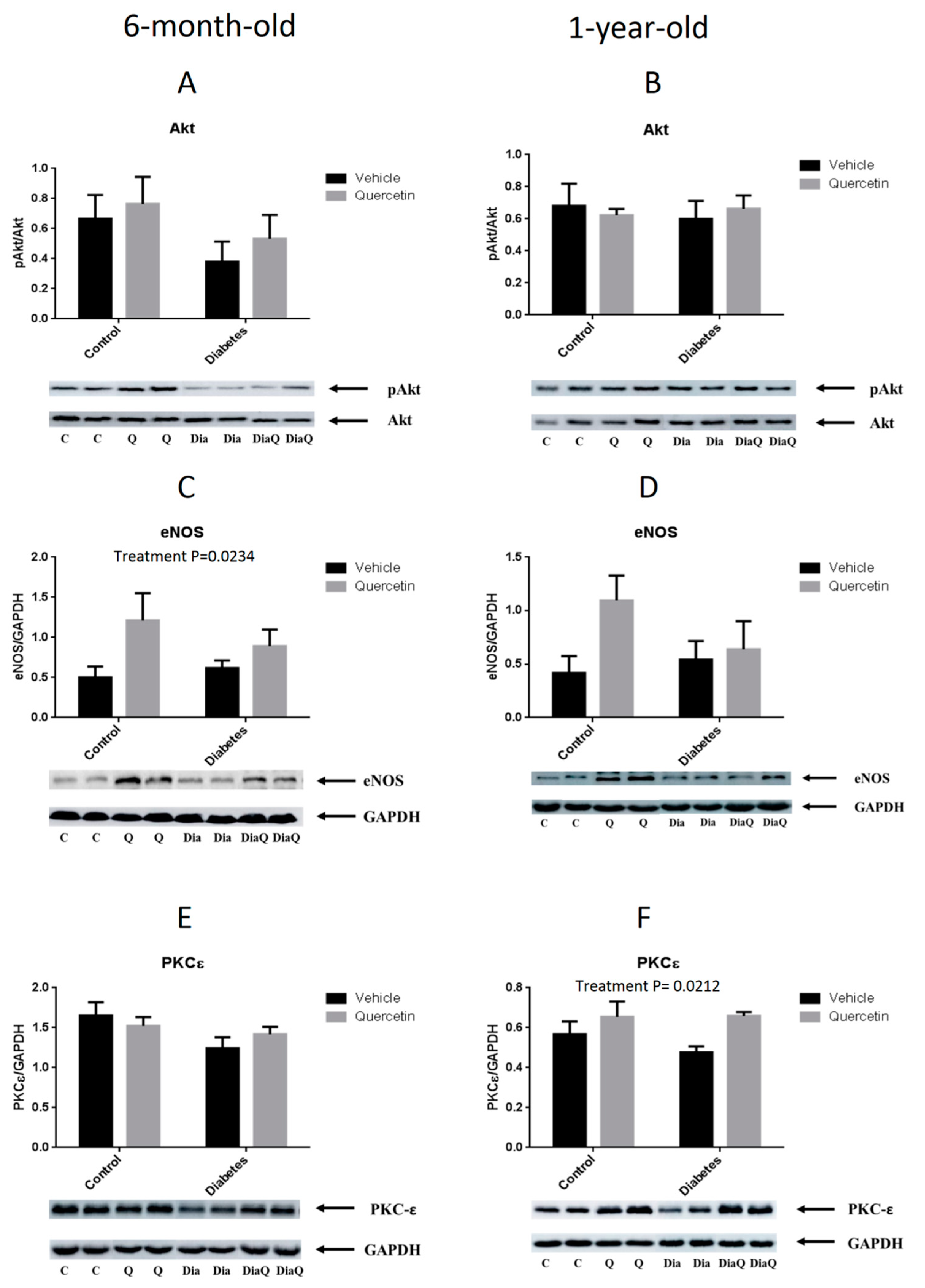
2.5. Effect of QCT on RISK Pathway Proteins Expression
3. Discussion
4. Materials and Methods
4.1. Materials
4.2. Experimental Model
4.3. Blood Pressure (BP) and Heart Rate (HR) Measurement
4.4. Perfusion Technique and Determination of Heart Function
4.5. Infarct Size Determination
4.6. Vascular Reactivity Measurements
4.7. Samples Collection
4.8. Preparation of Tissue Protein Fractions and Western Blot (WB) Analysis
4.9. Biochemical Evaluations in Blood Plasma
4.10. Statistical Evaluation
5. Conclusions
Supplementary Files
Supplementary File 1Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Li, Y.; Yao, J.; Han, C.; Yang, J.; Chaudhry, M.T.; Wang, S.; Liu, H.; Yin, Y. Quercetin, inflammation and immunity. Nutrients 2016, 8, 167. [Google Scholar] [CrossRef] [PubMed]
- Mlcek, J.; Jurikova, T.; Skrovankova, S.; Sochor, J. Quercetin and its anti-allergic immune response. Molecules 2016, 21, 623. [Google Scholar] [CrossRef] [PubMed]
- Patel, R.V.; Mistry, B.M.; Shinde, S.K.; Syed, R.; Singh, V.; Shin, H. Therapeutic potential of quercetin as a cardiovascular agent. Eur. J. Med. Chem. 2018, 155, 889–904. [Google Scholar] [CrossRef] [PubMed]
- Barreca, D.; Bellocco, E.; D’Onofrio, G.; Nabavi, S.F.; Daglia, M.; Rastrelli, L.; Nabavi, S.M. Neuroprotective effects of quercetin: from chemistry to medicine. CNS Neurol. Disord. Drug Targets 2016, 15, 964–975. [Google Scholar] [CrossRef] [PubMed]
- Reyes-Farias, M.; Carrasco-Pozo, C. The anti-cancer effect of quercetin: molecular implications in cancer metabolism. Int. J. Mol. Sci. 2019, 20, 3177. [Google Scholar] [CrossRef] [Green Version]
- Haddad, P.; Eid, H. The antidiabetic potential of quercetin: Underlying mechanisms. Curr. Med. Chem. 2016, 24, 355–364. [Google Scholar] [CrossRef] [PubMed]
- Xu, D.; Hu, M.-J.; Wang, Y.-Q.; Cui, Y.-L. Antioxidant activities of quercetin and its complexes for medicinal application. Molecules 2019, 24, 1123. [Google Scholar] [CrossRef] [Green Version]
- Dobrocsyova, V.; Krskova, K.; Capcarova, M.; Zorad, S. Modulation of adipogenesis and oxidative status by quercetin and ochratoxin a: Positive or negative impact on rat adipocyte metabolism? Molecules 2019, 24, 3726. [Google Scholar] [CrossRef] [Green Version]
- Marunaka, Y.; Marunaka, R.; Sun, H.; Yamamoto, T.; Kanamura, N.; Inui, T.; Taruno, A. Actions of quercetin, a polyphenol, on blood pressure. Molecules 2017, 22, 209. [Google Scholar] [CrossRef]
- Perez, A.; Gonzalez-Manzano, S.; Jimenez, R.; Perez-Abud, R.; Haro, J.M.; Osuna, A.; Santos-Buelga, C.; Duarte, J.; Perez-Vizcaino, F. The flavonoid quercetin induces acute vasodilator effects in healthy volunteers: Correlation with beta-glucuronidase activity. Pharmacol. Res. 2014, 89, 11–18. [Google Scholar] [CrossRef]
- Barteková, M.; Carnická, S.; Pancza, D.; Ondrejcáková, M.; Breier, A.; Ravingerová, T. Acute treatment with polyphenol quercetin improves postischemic recovery of isolated perfused rat hearts after global ischemia. Can. J. Physiol. Pharmacol. 2010, 88, 465–471. [Google Scholar] [CrossRef] [PubMed]
- Bartekova, M.; Radosinska, J.; Pancza, D.; Barancik, M.; Ravingerova, T. Cardioprotective effects of quercetin against ischemia-reperfusion injury are age-dependent. Physiol. Res. 2016, 65 (Suppl. 1), S101–S107. [Google Scholar]
- Dong, L.Y.; Chen, F.; Xu, M.; Yao, L.P.; Zhang, Y.J.; Zhuang, Y. Quercetin attenuates myocardial ischemia-reperfusion injury via downregulation of the HMGB1-TLR4-NF-κB signaling pathway. Am. J. Transl. Res. 2018, 10, 1273–1283. [Google Scholar] [PubMed]
- Dong, Q.; Chen, L.; Lu, Q.; Sharma, S.; Li, L.; Morimoto, S.; Wang, G. Quercetin attenuates doxorubicin cardiotoxicity by modulating Bmi-1 expression. Br. J. Pharmacol. 2014, 171, 4440–4454. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Barteková, M.; Šimončíková, P.; Fogarassyová, M.; Ivanová, M.; Okruhlicová, Ľ.; Tribulová, N.; Dovinová, I.; Barančík, M. Quercetin improves postischemic recovery of heart function in doxorubicin-treated rats and prevents doxorubicin-induced matrix metalloproteinase-2 activation and apoptosis induction. Int. J. Mol. Sci. 2015, 16, 8168–8185. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jin, H.-B.; Yang, Y.-B.; Song, Y.-L.; Zhang, Y.; Li, Y.-R. Protective roles of quercetin in acute myocardial ischemia and reperfusion injury in rats. Mol. Biol. Rep. 2012, 39, 11005–11009. [Google Scholar] [CrossRef] [PubMed]
- Liu, H.; Guo, X.; Chu, Y.; Lu, S. Heart protective effects and mechanism of quercetin preconditioning on anti-myocardial ischemia reperfusion (IR) injuries in rats. Gene 2014, 545, 149–155. [Google Scholar] [CrossRef]
- Liu, X.; Yu, Z.; Huang, X.; Gao, Y.; Wang, X.; Gu, J.; Xue, S. Peroxisome proliferator-activated receptor γ (PPARγ) mediates the protective effect of quercetin against myocardial ischemia-reperfusion injury via suppressing the NF-κB pathway. Am. J. Transl. Res. 2016, 8, 5169–5186. [Google Scholar]
- Li, C.; Wang, T.; Zhang, C.; Xuan, J.; Su, C.; Wang, Y. Quercetin attenuates cardiomyocyte apoptosis via inhibition of JNK and p38 mitogen-activated protein kinase signaling pathways. Gene 2016, 577, 275–280. [Google Scholar] [CrossRef]
- Shu, Z.; Yang, Y.; Yang, L.; Jiang, H.; Yu, X.; Wang, Y. Cardioprotective effects of dihydroquercetin against ischemia reperfusion injury by inhibiting oxidative stress and endoplasmic reticulum stress-induced apoptosis: Via the PI3K/Akt pathway. Food Funct. 2019, 10, 213–215. [Google Scholar] [CrossRef]
- Annapurna, A.; Reddy, C.S.; Akondi, R.B.; Rao, S.R.C. Cardioprotective actions of two bioflavonoids, quercetin and rutin, in experimental myocardial infarction in both normal and streptozotocin-induced type I diabetic rats. J. Pharm. Pharmacol. 2009, 61, 1365–1374. [Google Scholar] [CrossRef] [PubMed]
- Duarte, J.; Pérez-Palencia, R.; Vargas, F.; Angeles Ocete, M.; Pérez-Vizcaino, F.; Zarzuelo, A.; Tamargo, J. Antihypertensive effects of the flavonoid quercetin in spontaneously hypertensive rats. Br. J. Pharmacol. 2001, 133, 117–124. [Google Scholar] [CrossRef] [PubMed]
- Galindo, P.; Rodriguez-Gómez, I.; González-Manzano, S.; Dueñas, M.; Jiménez, R.; Menéndez, C.; Vargas, F.; Tamargo, J.; Santos-Buelga, C.; Pérez-Vizcaíno, F.; et al. Glucuronidated quercetin lowers blood pressure in spontaneously hypertensive rats via deconjugation. PLoS ONE 2012, 7, e32673. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Calabró, V.; Litterio, M.C.; Fraga, C.G.; Galleano, M.; Piotrkowski, B. Effects of quercetin on heart nitric oxide metabolism in l-NAME treated rats. Arch. Biochem. Biophys. 2018, 647, 47–53. [Google Scholar] [CrossRef]
- Galisteo, M.; García-Saura, M.F.; Jiménez, R.; Villar, I.C.; Wangensteen, R.; Zarzuelo, A.; Vargas, F.; Duarte, J. Effects of quercetin treatment on vascular function in deoxycorticosterone acetate-salt hypertensive rats. Comparative study with verapamil. Planta Med. 2004, 70, 334–341. [Google Scholar]
- García-Saura, M.F.; Galisteo, M.; Villar, I.C.; Bermejo, A.; Zarzuelo, A.; Vargas, F.; Duarte, J. Effects of chronic quercetin treatment in experimental renovascular hypertension. Mol. Cell. Biochem. 2005, 270, 147–155. [Google Scholar] [CrossRef]
- Egert, S.; Bosy-Westphal, A.; Seiberl, J.; Kürbitz, C.; Settler, U.; Plachta-Danielzik, S.; Wagner, A.E.; Frank, J.; Schrezenmeir, J.; Rimbach, G.; et al. Quercetin reduces systolic blood pressure and plasma oxidised low-density lipoprotein concentrations in overweight subjects with a high-cardiovascular disease risk phenotype: A double-blinded, placebo-controlled cross-over study. Br. J. Nutr. 2009, 102, 1065–1074. [Google Scholar] [CrossRef] [Green Version]
- Serban, M.C.; Sahebkar, A.; Zanchetti, A.; Mikhailidis, D.P.; Howard, G.; Antal, D.; Andrica, F.; Ahmed, A.; Aronow, W.S.; Muntner, P.; et al. Effects of quercetin on blood pressure: A systematic review and meta-analysis of randomized controlled trials. J. Am. Heart Assoc. 2016, 5, 1–16. [Google Scholar] [CrossRef] [Green Version]
- Zahedi, M.; Ghiasvand, R.; Feizi, A.; Asgari, G.; Darvish, L. Does quercetin improve cardiovascular risk factors and inflammatory biomarkers in women with type 2 diabetes: A double-blind randomized controlled clinical trial. Int. J. Prev. Med. 2013, 4, 777–785. [Google Scholar]
- Rivera, L.; Morón, R.; Sánchez, M.; Zarzuelo, A.; Galisteo, M. Quercetin ameliorates metabolic syndrome and improves the inflammatory status in obese zucker rats. Obesity 2008, 16, 2081–2087. [Google Scholar] [CrossRef]
- Khoo, N.K.H.; White, C.R.; Pozzo-Miller, L.; Zhou, F.; Constance, C.; Inoue, T.; Patel, R.P.; Parks, D.A. Dietary flavonoid quercetin stimulates vasorelaxation in aortic vessels. Free Radic. Biol. Med. 2010, 49, 339–347. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Monori-Kiss, A.; Monos, E.; Nádasy, G.L. Quantitative analysis of vasodilatory action of quercetin on intramural coronary resistance arteries of the rat in vitro. PLoS ONE 2014, 9, e105587. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Choi, S.; Ryu, K.H.; Park, S.H.; Jun, J.Y.; Shin, B.C.; Chung, J.H.; Yeum, C.H. Direct vascular actions of quercetin in aorta from renal hypertensive rats. Kidney Res. Clin. Pract. 2016, 35, 15–21. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yuan, T.-Y.; Niu, Z.-R.; Chen, D.; Chen, Y.-C.; Zhang, H.-F.; Fang, L.-H.; Du, G.-H. Vasorelaxant effect of quercetin on cerebral basilar artery in vitro and the underlying mechanisms study. J. Asian Nat. Prod. Res. 2018, 20, 477–487. [Google Scholar] [CrossRef] [PubMed]
- Shen, Y.; Ward, N.C.; Hodgson, J.M.; Puddey, I.B.; Wang, Y.; Zhang, D.; Maghzal, G.J.; Stocker, R.; Croft, K.D. Dietary quercetin attenuates oxidant-induced endothelial dysfunction and atherosclerosis in apolipoprotein E knockout mice fed a high-fat diet: A critical role for heme oxygenase-1. Free Radic. Biol. Med. 2013, 65, 908–915. [Google Scholar] [CrossRef] [PubMed]
- Roghani, M.; Baluchnejadmojarad, T.; Reza Vaez-Mahdavi, M.; Roghani-Dehkordi, F. Mechanisms underlying quercetin-induced vasorelaxation in aorta of subchronic diabetic rats: An in vitro study. Vascul. Pharmacol. 2004, 42, 31–35. [Google Scholar] [CrossRef]
- Ajay, M.; Achike, F.I.; Mustafa, A.M.; Mustafa, M.R. Effect of quercetin on altered vascular reactivity in aortas isolated from streptozotocin-induced diabetic rats. Diabetes Res. Clin. Pract. 2006, 73, 1–7. [Google Scholar] [CrossRef] [Green Version]
- Kunasegaran, T.; Mustafa, M.R.; Achike, F.I.; Murugan, D.D. Quercetin and pioglitazone synergistically reverse endothelial dysfunction in isolated aorta from fructose-streptozotocin (F-STZ)-induced diabetic rats. Eur. J. Pharmacol. 2017, 799, 160–170. [Google Scholar] [CrossRef]
- Gasparotto Junior, A.; dos Reis Piornedo, R.; Assreuy, J.; Da Silva-Santos, J.E. Nitric oxide and K ir 6.1 potassium channel mediate isoquercitrin-induced endothelium-dependent and independent vasodilation in the mesenteric arterial bed of rats. Eur. J. Pharmacol. 2016, 788, 328–334. [Google Scholar] [CrossRef]
- Hou, X.; Liu, Y.; Niu, L.; Cui, L.; Zhang, M. Enhancement of voltage-gated K+ channels and depression of voltage-gated ca2+ channels are involved in quercetin-induced vasorelaxation in rat coronary artery. Planta Med. 2014, 80, 465–472. [Google Scholar] [CrossRef]
- García-Prieto, C.F.; Pulido-Olmo, H.; Ruiz-Hurtado, G.; Gil-Ortega, M.; Aranguez, I.; Rubio, M.A.; Ruiz-Gayo, M.; Somoza, B.; Fernández-Alfonso, M.S. Mild caloric restriction reduces blood pressure and activates endothelial AMPK-PI3K-Akt-eNOS pathway in obese Zucker rats. Vascul. Pharmacol. 2015, 65–66, 3–12. [Google Scholar] [CrossRef] [PubMed]
- Luo, H.; Wang, X.; Chen, C.; Wang, J.; Zou, X.; Li, C.; Xu, Z.; Yang, X.; Shi, W.; Zeng, C. Oxidative stress causes imbalance of renal renin angiotensin system (ras) components and hypertension in obese zucker rats. J. Am. Heart Assoc. 2015, 4, 1–14. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Morales-Cano, D.; Callejo, M.; Barreira, B.; Mondejar-Parreño, G.; Esquivel-Ruiz, S.; Ramos, S.; Martín, M.Á.; Cogolludo, A.; Moreno, L.; Perez-Vizcaino, F. Elevated pulmonary arterial pressure in Zucker diabetic fatty rats. PLoS ONE 2019, 14, e0211281. [Google Scholar] [CrossRef] [PubMed]
- Kojda, G.; Harrison, D. Interactions between NO and reactive oxygen species: Pathophysiological importance in atherosclerosis, hypertension, diabetes and heart failure. Cardiovasc. Res. 1999, 43, 562–571. [Google Scholar] [CrossRef] [Green Version]
- Yan, L.; Zhang, J.D.; Wang, B.; Lv, Y.J.; Jiang, H.; Liu, G.L.; Qiao, Y.; Ren, M.; Guo, X.F. Quercetin inhibits left ventricular hypertrophy in spontaneously hypertensive rats and inhibits angiotensin II-induced H9C2 cells hypertrophy by enhancing PPAR-γ expression and suppressing AP-1 activity. PLoS ONE 2013, 8, e72548. [Google Scholar] [CrossRef] [Green Version]
- Galindo, P.; González-Manzano, S.; Zarzuelo, M.J.; Gómez-Guzmán, M.; Quintela, A.M.; González-Paramás, A.; Santos-Buelga, C.; Pérez-Vizcaíno, F.; Duarte, J.; Jiménez, R. Different cardiovascular protective effects of quercetin administered orally or intraperitoneally in spontaneously hypertensive rats. Food Funct. 2012, 3, 643. [Google Scholar] [CrossRef]
- Machha, A.; Achike, F.I.; Mustafa, A.M.; Mustafa, M.R. Quercetin, a flavonoid antioxidant, modulates endothelium-derived nitric oxide bioavailability in diabetic rat aortas. Nitric Oxide 2007, 16, 442–447. [Google Scholar] [CrossRef]
- Küng, C.F.; Lüscher, T.F. Different mechanisms of endothelial dysfunction with aging and hypertension in rat aorta. Hypertension 1995, 25, 194–200. [Google Scholar] [CrossRef]
- Vanhoutte, P. Ageing and endothelial dysfunction. Eur. Hear. J. Suppl. 2002, 4, A8–A17. [Google Scholar] [CrossRef] [Green Version]
- Puzserova, A.; Ilovska, V.; Balis, P.; Slezak, P.; Bernatova, I. Age-Related alterations in endothelial function of femoral artery in young SHR and WKY rats. Biomed. Res. Int. 2014, 2014, 1–12. [Google Scholar] [CrossRef] [Green Version]
- Duarte, J.; Jiménez, R.; O’Valle, F.; Galisteo, M.; Pérez-Palencia, R.; Vargas, F.; Pérez-Vizcaíno, F.; Zarzuelo, A.; Tamargo, J. Protective effects of the flavonoid quercetin in chronic nitric oxide deficient rats. J. Hypertens. 2002, 20, 1843–1854. [Google Scholar] [CrossRef] [PubMed]
- Sánchez, M.; Galisteo, M.; Vera, R.; Villar, I.C.; Zarzuelo, A.; Tamargo, J.; Pérez-Vizcaíno, F.; Duarte, J. Quercetin downregulates NADPH oxidase, increases eNOS activity and prevents endothelial dysfunction in spontaneously hypertensive rats. J. Hypertens. 2006, 24, 75–84. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Morales-Cano, D.; Menendez, C.; Moreno, E.; Moral-Sanz, J.; Barreira, B.; Galindo, P.; Pandolfi, R.; Jimenez, R.; Moreno, L.; Cogolludo, A.; et al. The flavonoid quercetin reverses pulmonary hypertension in rats. PLoS ONE 2014, 9, e114492. [Google Scholar] [CrossRef] [PubMed]
- Panchal, S.K.; Poudyal, H.; Brown, L. Quercetin ameliorates cardiovascular, hepatic, and metabolic changes in diet-induced metabolic syndrome in rats. J. Nutr. 2012, 142, 1026–1032. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Suganya, N.; Dornadula, S.; Chatterjee, S.; Mohanram, R.K. Quercetin improves endothelial function in diabetic rats through inhibition of endoplasmic reticulum stress-mediated oxidative stress. Eur. J. Pharmacol. 2018, 819, 80–88. [Google Scholar] [CrossRef]
- Matsumoto, T.; Kakami, M.; Noguchi, E.; Kobayashi, T.; Kamata, K. Imbalance between endothelium-derived relaxing and contracting factors in mesenteric arteries from aged OLETF rats, a model of Type 2 diabetes. Am. J. Physiol. Circ. Physiol. 2007, 293, H1480–H1490. [Google Scholar] [CrossRef]
- Sengupta, P. The laboratory rat: Relating its age with human’s. Int. J. Prev. Med. 2013, 4, 624–630. [Google Scholar]
- Griecsová, L.; Farkašová, V.; Gáblovský, I.; Khandelwal, V.K.M.; Bernátová, I.; Tatarková, Z.; Kaplan, P.; Ravingerová, T. Effect of maturation on the resistance of rat hearts against ischemia. Study of potential molecular mechanisms. Physiol. Res. 2015, 64 (Suppl. 5), S685–S696. [Google Scholar]
- Han, J.; Xuan, J.L.; Hu, H.R.; Chen, Z.W. Protective effect against myocardial ischemia reperfusion injuries induced by hyperoside preconditioning and its relationship with PI3K/Akt signaling pathway in rats. Zhongguo Zhong Yao Za Zhi 2015, 40, 118–123. [Google Scholar]
- Rossello, X.; Yellon, D.M. The RISK pathway and beyond. Basic Res. Cardiol. 2018, 113, 2. [Google Scholar] [CrossRef] [Green Version]
- Ledvenyiova, V.; Pancza, D.; Matejiková, J.; Ferko, M.; Bernatova, I.; Ravingerova, T. Impact of age and sex on response to ischemic preconditioning in the rat heart: Differential role of the PI3K-AKT pathway. Can. J. Physiol. Pharmacol. 2013, 91, 640–647. [Google Scholar] [CrossRef] [PubMed]
- Ondrejcakova, M.; Barancik, M.; Bartekova, M.; Ravingerova, T.; Jezova, D. Prolonged oxytocin treatment in rats affects intracellular signaling and induces myocardial protection against infarction. Gen. Physiol. Biophys. 2012, 31, 261–270. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zálešák, M.; Blažíček, P.; Gablovský, I.; Ledvényiová, V.; Barteková, M.; Ziegelhöffer, A.; Ravingerová, T. Impaired PI3K/Akt signaling as a potential cause of failure to precondition rat hearts under conditions of simulated hyperglycemia. Physiol. Res. 2015, 64, 633–641. [Google Scholar] [PubMed]
- Sasváriová, M.; Tyukos-Kaprinay, B.; Salvaras, L.; Belovičová, K.; Bögi, E.; Knezl, V.; Barteková, M.; Stankovičová, T.; Dubovický, M. Effect of pre-gestational stress and prenatal venlafaxine administration on cardiovascular system of rat offspring. Eur. Pharm. J. 2018, 65, 17–22. [Google Scholar] [CrossRef] [Green Version]
- Barteková, M.; Styk, J.; Pancza, D.; Kukan, M.; Šeboková, J.; Breier, A. Proteins released from liver after ischaemia induced and elevation of heart resistance against ischaemia-reperfusion injury: 1. Beneficial effect of protein fraction isolated from perfusate after ischaemia and reperfusion of liver. Gen. Physiol. Biophys. 2003, 22, 567–577. [Google Scholar] [PubMed]
- Barteková, M.; Sulová, Z.; Pancza, D.; Ravingerová, T.; Stankovicová, T.; Styk, J.; Breier, A. Proteins released from liver after ischaemia induced an elevation of heart resistance against ischaemia-reperfusion injury: 2. Beneficial effect of liver ischaemia in situ. Gen. Physiol. Biophys. 2004, 23, 489–497. [Google Scholar] [PubMed]
- Kristek, F.; Cacanyiova, S.; Gerova, M. Hypotrophic effect of long-term neuronal NO-synthase inhibition on heart and conduit arteries of the Wistar rats. J. Physiol. Pharmacol. 2009, 60, 21–27. [Google Scholar]
- Bradford, M.M. A rapid and sensitive method for the quantitation of microgram quantities of protein utilizing the principle of protein-dye binding. Anal. Biochem. 1976, 72, 248–254. [Google Scholar] [CrossRef]
Sample Availability: Samples of the compounds are not available from the authors. |









{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| 6-Month-Old Rats | 1-Year-Old Rats | |||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| LEAN | OBESE | Two-Way ANOVA | LEAN | OBESE | Two-Way ANOVA | |||||||||
| C | Q | Dia | DiaQ | Diabetes | Treatment | Interaction | C | Q | Dia | DiaQ | Diabetes | Treatment | Interaction | |
| BW (g) | 349 ± 7 | 346 ± 9 | 493 ± 8 | 487 ± 10 | **** | ns. | ns. | 406 ± 10 | 413 ± 8 | 520 ± 25 | 514 ± 24 | **** | ns. | ns. |
| BW/tibia (g/mm) | 9.2 ±0.2 | 8.9 ± 0.2 | 13.2 ± 0.2 | 12.9 ± 0.3 | **** | ns. | ns. | 10.2 ± 0.2 | 10.3 ± 0.2 | 13.7 ± 0.6 | 13.4 ± 0.7 | **** | ns. | ns. |
| HW/tibia (g/mm) | 0.03 ± 0.001 | 0.03 ± 0.001 | 0.04 ± 0.001 | 0.04 ± 0.001 | **** | ns. | ns. | 0.03 ± 0.001 | 0.03 ± 0.001 | 0.04 ± 0.001 | 0.04 ± 0.01 | ** | ns. | ns. |
| Glucose (mmol/L) | 7.3 ± 0.4 | 6.9 ± 0.2 | 11.2 ± 1.3 | 12.6 ± 1.9 | ** | ns. | ns. | 6.1 ± 0.1 | 5.9 ± 0.2 | 16.9 ± 1.6 | 18.5 ± 2.1 | **** | ns. | ns. |
| TAG (mmol/L) | 0.38 ± 0.13 | 0.40 ± 0.08 | 5.87 ± 0.89 | 5.76 ± 1.23 | **** | ns. | ns. | 0.24 ± 0.05 | 0.18 ± 0.03 | 3.66 ± 0.41 | 3.67 ± 0.39 | **** | ns. | ns. |
| Ch. (mmol/L) | 1.92 ± 0.09 | 1.63 ± 0.07 | 2.44 ± 0.28 | 2.7 ± 0.32 | ** | ns. | ns. | 2.85 ± 0.11 | 2.81 ± 0.08 | 4.86 ± 0.41 | 4.79 ± 0.25 | **** | ns. | ns. |
| HDL-Ch. (mmol/L) | 0.93 ± 0.08 | 0.67 ± 0.07 | 1.01 ± 0.18 | 0.97 ± 0.16 | ns. | ns. | ns. | 1.36 ± 0.04 | 1.37 ± 0.04 | 2.36 ± 0.13 | 2.37 ± 0.07 | **** | ns. | ns. |
| LDL-Ch. (mmol/L) | 0.46 ± 0.03 | 0.36 ± 0.02 | 0.34 ± 0.07 | 0.32 ± 0.06 | ns. | ns. | ns. | 0.82 ± 0.04 | 0.81 ± 0.02 | 1.05 ± 0.08 | 0.91 ± 0.07 | * | ns. | ns. |
| 6-Month-Old Rats | 1-Year-Old Rats | |||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| LEAN | OBESE | Two-Way ANOVA | LEAN | OBESE | Two-Way ANOVA | |||||||||
| C | Q | Dia | DiaQ | Diabetes | Treatment | Interaction | C | Q | Dia | DiaQ | Diabetes | Treatment | Interaction | |
| LVDP (mmHg) | 81 ± 7 | 72 ± 9 | 83 ± 8 | 71 ± 3 | ns. | ns. | ns. | 73 ± 5 | 90 ± 9 | 114 ± 9 | 109 ± 4 | *** | ns. | ns. |
| +(dp/dt)max (mmHg) | 1789 ± 179 | 1548 ± 241 | 1615 ±187 | 1383 ± 123 | ns. | ns. | ns. | 1522 ± 135 | 1909 ± 213 | 2143 ± 156 | 2048 ± 110 | * | ns. | ns. |
| −(dp/dt)max (mmHg) | 1287 ± 127 | 1187 ± 183 | 1317 ±154 | 1136 ± 112 | ns. | ns. | ns. | 1158 ± 143 | 1451 ± 198 | 1690 ± 128 | 1661 ± 72 | * | ns. | ns. |
| HR (beats/min) | 226 ± 28 | 237 ± 13 | 207 ± 18 | 173 ± 19 | ns. | ns. | ns. | 227 ± 29 | 210 ± 10 | 154 ± 12 | 162 ± 17 | ** | ns. | ns. |
| RPP (mmHg/min) | 18,583 ± 3047 | 16,867 ± 2304 | 17,495 ± 2975 | 12,300 ± 1629 | ns. | ns. | ns. | 16,533 ± 2298 | 19,060 ± 2675 | 17,341 ± 1494 | 17,794 ± 2107 | ns. | ns. | ns. |
| CF (mL) | 12 ± 2 | 16 ± 2 | 15 ± 1 | 12 ± 2 | ns. | ns. | ns. | 12 ± 2 | 12 ± 0 | 18 ± 1 | 16 ± 1 | *** | ns. | ns. |
| Experimental Group | 6-Month-Old | 1-Year-Old |
|---|---|---|
| Control (C) | n = 12 | n = 12 |
| QCT-treated (Q) | n = 12 | n = 13 |
| Diabetic (Dia) | n = 12 | n = 16 |
| Diabetic QCT-treated (DiaQ) | n = 12 | n = 17 |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Ferenczyova, K.; Kalocayova, B.; Kindernay, L.; Jelemensky, M.; Balis, P.; Berenyiova, A.; Zemancikova, A.; Farkasova, V.; Sykora, M.; Tothova, L.; et al. Quercetin Exerts Age-Dependent Beneficial Effects on Blood Pressure and Vascular Function, But Is Inefficient in Preventing Myocardial Ischemia-Reperfusion Injury in Zucker Diabetic Fatty Rats. Molecules 2020, 25, 187. https://doi.org/10.3390/molecules25010187
Ferenczyova K, Kalocayova B, Kindernay L, Jelemensky M, Balis P, Berenyiova A, Zemancikova A, Farkasova V, Sykora M, Tothova L, et al. Quercetin Exerts Age-Dependent Beneficial Effects on Blood Pressure and Vascular Function, But Is Inefficient in Preventing Myocardial Ischemia-Reperfusion Injury in Zucker Diabetic Fatty Rats. Molecules. 2020; 25(1):187. https://doi.org/10.3390/molecules25010187
Chicago/Turabian StyleFerenczyova, Kristina, Barbora Kalocayova, Lucia Kindernay, Marek Jelemensky, Peter Balis, Andrea Berenyiova, Anna Zemancikova, Veronika Farkasova, Matus Sykora, Lubomira Tothova, and et al. 2020. "Quercetin Exerts Age-Dependent Beneficial Effects on Blood Pressure and Vascular Function, But Is Inefficient in Preventing Myocardial Ischemia-Reperfusion Injury in Zucker Diabetic Fatty Rats" Molecules 25, no. 1: 187. https://doi.org/10.3390/molecules25010187