Current Trends on Seaweeds: Looking at Chemical Composition, Phytopharmacology, and Cosmetic Applications

 , , , ,
, , , ,  ,
,  ,
,  , ,
, ,  and
and 
Abstract
:1. Introduction
2. Seaweeds: A Critical Perspective
2.1. Food and Nutritional Applications
2.2. Seaweeds as Biofertilizers
3. Macroalgae Metabolites Diversity
3.1. Primary Metabolites
3.1.1. Carbohydrates
Fucoidans
Agars
Carrageenans
Ulvans
Alginates
Laminaran
3.1.2. Minerals
3.1.3. Proteins
3.1.4. Lipids
3.2. Macroalgae Secondary Metabolites Diversity
3.2.1. Chlorophyta Secondary Metabolites
3.2.2. Ochrophyta Secondary Metabolites
3.2.3. Rhodophyta Secondary Metabolites
4. Seaweeds Bioactive Effects: From Pre-Clinical to Clinical Studies
4.1. Seaweeds: In Vitro Bioactive Effects
4.1.1. Antimicrobial Activity
4.1.2. Antioxidant Activity
4.1.3. Anti-Inflammatory Activity
4.1.4. Antiproliferative and Anti-Angiogenesis Activity
4.1.5. Anticoagulant Activity
4.2. Seaweeds: In Vivo Bioactive Effects
4.3. Seaweeds: Clinical Evidence
5. Seaweeds Quality Parameters and Health Safety
5.1. A Brief Overview of Regulatory Practices
5.2. Multidimensional Procedures to Overcome Seaweeds’ Antinutrients
5.3. Bioavailability of Seaweed Bioactive Compounds
6. Targeting Seaweed Potentialities for Cosmetic Purposes
6.1. Enzymes, Collagen, and Bioactive Peptides
6.2. Fatty Acids and Derivatives
6.3. Phlorotannins and Polyphenols
6.4. Polysaccharides
6.5. Carotenoid Pigments, Fucosterols, Tocopherols, and Terpenoids
7. Seaweeds as Promising Cosmetics: What will Happen?
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Bhadury, P.; Wright, P.C. Exploitation of marine algae: biogenic compounds for potential antifouling applications. Planta 2004, 219, 561–578. [Google Scholar] [CrossRef] [PubMed]
- Ale, M.T.; Meyer, A.S. Fucoidans from brown seaweeds: An update on structures, extraction techniques and use of enzymes as tools for structural elucidation. RSC Adv. 2013, 3, 8131–8141. [Google Scholar] [CrossRef]
- Irkin, L.C.; Yayintas, Ö. Pharmacological Properties and Therapeutic Benefits of Seaweeds (A Review). Int. J. Trend Sci. Res. Dev. 2018, 2, 1126–1131. [Google Scholar] [CrossRef]
- Pal, A.; Kamthania, M.C.; Kumar, A. Bioactive Compounds and Properties of Seaweeds: A Review. Open Access Libr. J. 2014, 1, e752. [Google Scholar] [CrossRef]
- Anantharaman, P. Manual on Identification of Seaweed. All India Coordinate Project on Survey and Inventori- zation of Coastal and Marine Biodiversity. J. Mar. Biol. Assoc. India 2002, 29, 1–9. [Google Scholar]
- Figueiredo, M.A.O.; Creed, J.C. Marine Algae and Plant. In Encyclopedia of Life Support Systems; Eolss Publishers Co. Ltd: Oxford, MS, USA, 2009; pp. 1–7. [Google Scholar]
- Harley, C.; Anderson, K.; Demes, K.; Jorve, J.; Kordas, R.; Coyle, T. Effects of climate change on global seaweed communities. Phychological Soc. Am. 2012. [Google Scholar] [CrossRef]
- Christie, H.; Norderhaug, K.M.; Fredriksen, S. Macrophytes as habitat for fauna. Mar. Ecol. Prog. Ser. 2009, 396, 221–233. [Google Scholar] [CrossRef]
- Van Der Strate, H.J.; Boele-Bos, S.A.; Olsen, J.L.; van de, Z.L.; Stam, W.T. Phylogeographic studies in the tropical seaweed Cladophoropsis membranacea Chlorophyta, Ulvophyceae reveal a cryptic species complex. J. Phycol. 2002, 38, 572–582. [Google Scholar] [CrossRef]
- Wernberg, T.; Smale, D.; Thomsen, M. A decade of climate change experiments on marine organisms: procedures, patterns and problems. Glob. Chang. Biol. 2012, 18, 1491–1498. [Google Scholar] [CrossRef]
- Raven, J. Responses of aquatic photosynthetic organisms to increased solar UV-B. J. Photochem. Photobiol. B Biol. 1991, 9, 239–244. [Google Scholar] [CrossRef]
- Hoegh-Guldberg, O.; Mumby, P.J.; Hooten, A.J.; Steneck, R.S.; Greenfield, P.; Gomez, E.; Harvell, C.D.; Sale, P.F.; Edwards, A.J.; Caldeira, K.; et al. Coral reefs under rapid climate change and ocean acidification. Science 2007, 318, 1737–1742. [Google Scholar] [CrossRef] [PubMed]
- Davison, I.; Pearson, G. Stress tolerance in intertidal seaweeds. J. Phycol. 1996, 32, 197–211. [Google Scholar] [CrossRef]
- Swarnalatha, G. Comparison of nutritive value of seaweed with other terrestrial foods—a review. Int. J. Food Nutr. Sci. 2018, 7, 61–70. [Google Scholar]
- Lavanya, M.; Thangamathi, P.; Ananth, S.; Gnanasoundari, A. Properties of seaweeds: A review. Int. Jounal Curr. Innov. Res. 2017, 3, 843–846. [Google Scholar]
- Murata, M.; Nakazoe, J. Production and use of marine algae in Japan. Japan Agric. Res. Q. 2001, 35, 281–290. [Google Scholar] [CrossRef]
- Mahadevan, K. Seaweeds: a sustainable food source. In Seaweed Sustainability; Academic press: Cambridge, MA, USA, 2015; pp. 347–364. [Google Scholar]
- Madhusudan, C.; Manoj, S.; Rhaul, K.; Rishi, C. Seaweeds: A Diet with nutritional, medicinal and industrial value. Res. J. Med. Plant 2011, 5, 153–157. [Google Scholar] [CrossRef]
- Rhein-Knudsen, N.; Ale, M.T.; Meyer, A.S. Seaweed hydrocolloid production: an update on enzyme assisted extraction and modification technologies. Mar. Drugs 2015, 13, 3340–3359. [Google Scholar] [CrossRef]
- Pereira, L.; Sousa, A.; Coelho, H.; Amado, A.; Ribeiro-Claro, P. Use of FTIR, FT-Raman and 13C-NMR spectroscopy for identification of some seaweed phycocolloids. Biomol. Eng. 2003, 20, 223–228. [Google Scholar] [CrossRef]
- Kumar, C.S.C.S.; Ganesan, P.; Suresh, P.V.V.; Bhaskar, N. Seaweeds as a Source of Nutritionally Beneficial Compounds: A Review. J. Food Sci. Technol. 2008, 45, 1–13. [Google Scholar]
- Lahaye, M. Marine algae as sources of fibres: Determination of soluble and insoluble dietary fibrecontents in some ‘sea vegetables. ’ J. Sci. Food Agric. 1991, 54, 587–594. [Google Scholar] [CrossRef]
- Kolanjinathan, K.; Ganesh, P.; Saranraj, P. Pharmacological Importance of Seaweeds: A Review. World J. Fish Mar. Sci. 2014, 6, 1–15. [Google Scholar]
- Fleurence, J.; Coeur, C. Le Influence of mineralization methods on the determination of the mineral content of brown seaweed Undaria pinnatifida by Atomic absorption spectrophotometry. Hydrobiologia 1993, 260–261, 531–534. [Google Scholar] [CrossRef]
- Mabeau, S.; Fleurence, J. Seaweed in food products: biochemical and nutritional aspects. Trends Food Sci. Technol. 1993, 4, 103–107. [Google Scholar] [CrossRef]
- Suzuki, H.; Higuchi, T.; Sawa, K.; Ohtaki, S.; Tolli, J. Endemic coast goitre in Hokkaido, Japan. Acta Endocrinol. (Copenh). 1965, 50, 161–176. [Google Scholar] [CrossRef] [PubMed]
- Patel, R.V.; Pandya, K.Y.; Jasrai, R.T.; Brahmbhatt, N. A review: scope of utilizing seaweed as a biofertilizer in agriculture. Int. J. Adv. Res. 2017, 5, 2046–2054. [Google Scholar] [CrossRef]
- Marinho-Soriano, E.; Fonseca, P.C.; Carneiro, M.A.A.; Moreira, W.S.C. Seasonal variation in the chemical composition of two tropical seaweeds. Bioresour. Technol. 2006, 97, 2402–2406. [Google Scholar] [CrossRef]
- Khairy, H.M.; El-Shafay, S.M. Seasonal variations in the biochemical composition of some common seaweed species from the coast of Abu Qir Bay, Alexandria, Egypt. Oceanologia 2013, 55, 435–452. [Google Scholar] [CrossRef]
- Balboa, E.M.; Gallego-Fábrega, C.; Moure, A.; Domínguez, H. Study of the seasonal variation on proximate composition of oven-dried Sargassum muticum biomass collected in Vigo Ria, Spain. J. Appl. Phycol. 2016, 28, 1943–1953. [Google Scholar] [CrossRef]
- Neto, R.; Marçal, C.; Queirós, A.; Abreu, H.; Silva, A.; Cardoso, S. Screening of Ulva rigida, Gracilaria sp., Fucus vesiculosus and Saccharina latissima as functional ingredients. Int. J. Mol. Sci. 2018, 19, 2987. [Google Scholar] [CrossRef]
- Hessami, M.J.; Phang, S.-M.; Salleh, A.; Rabiei, R. Evaluation of tropical seaweeds as feedstock for bioethanol production. Int. J. Environ. Sci. Technol. 2018, 15, 977–992. [Google Scholar] [CrossRef]
- Catarino, M.D.; Silva, A.M.S.; Cardoso, S.M. Phycochemical constituents and biological activities of Fucus spp. Mar. Drugs 2018, 16, 249. [Google Scholar] [CrossRef] [PubMed]
- Holdt, S.; Kraan, S. Bioactive compounds in seaweed: functional food applications and legislation. J. Appl. Phycol. 2011, 23, 543–597. [Google Scholar] [CrossRef]
- Bikker, P.; van Krimpen, M.M.; van Wikselaar, P.; Houweling-Tan, B.; Scaccia, N.; van Hal, J.W.; Huijgen, W.J.J.; Cone, J.W.; López-Contreras, A.M. Biorefinery of the green seaweed Ulva lactuca to produce animal feed, chemicals and biofuels. J. Appl. Phycol. 2016, 28, 3511–3525. [Google Scholar] [CrossRef] [PubMed]
- Cabrita, A.R.J.; Maia, M.R.G.; Oliveira, H.M.; Sousa-Pinto, I.; Almeida, A.A.; Pinto, E.; Fonseca, A.J.M. Tracing seaweeds as mineral sources for farm-animals. J. Appl. Phycol. 2016, 28. [Google Scholar] [CrossRef]
- Syad, A.N.; Shunmugiah, K.P.; Kasi, P.D. Seaweeds as nutritional supplements: Analysis of nutritional profile, physicochemical properties and proximate composition of G. acerosa and S. wightii. Biomed. Prev. Nutr. 2013, 3, 139–144. [Google Scholar] [CrossRef]
- Schiener, P.; Black, K.D.; Stanley, M.S.; Green, D.H. The seasonal variation in the chemical composition of the kelp species Laminaria digitata, Laminaria hyperborea, Saccharina latissima and Alaria esculenta. J. Appl. Phycol. 2015, 27, 363–373. [Google Scholar] [CrossRef]
- Rupérez, P. Mineral content of edible marine seaweeds. Food Chem. 2002, 79, 23–26. [Google Scholar] [CrossRef]
- Biancarosa, I.; Belghit, I.; Bruckner, C.G.; Liland, N.S.; Waagbø, R.; Amlund, H.; Heesch, S.; Lock, E.J. Chemical characterization of 21 species of marine macroalgae common in Norwegian waters: benefits of and limitations to their potential use in food and feed. J. Sci. Food Agric. 2018, 98, 2035–2042. [Google Scholar] [CrossRef] [Green Version]
- Fleurence, J. Seaweed proteins. In Proteins in Food Processing; Yada, R.Y., Ed.; CRC Woodhead Publishing Limited: Cambridge, UK, 2017. [Google Scholar]
- Vieira, E.F.; Soares, C.; Machado, S.; Correia, M.; Ramalhosa, M.J.; Oliva-Teles, M.T.; Paula Carvalho, A.; Domingues, V.F.; Antunes, F.; Oliveira, T.A.C.; et al. Seaweeds from the Portuguese coast as a source of proteinaceous material: Total and free amino acid composition profile. Food Chem. 2018, 269, 264–275. [Google Scholar] [CrossRef] [Green Version]
- Astorga-España, M.S.; Rodríguez-Galdón, B.; Rodríguez-Rodríguez, E.M.; Díaz-Romero, C. Amino acid content in seaweeds from the Magellan Straits (Chile). J. Food Compos. Anal. 2016, 53, 77–84. [Google Scholar] [CrossRef]
- Kendel, M.; Wielgosz-Collin, G.; Bertrand, S.; Roussakis, C.; Bourgougnon, N.; Bedoux, G. Lipid composition, fatty acids and sterols in the seaweeds Ulva armoricana, and Solieria chordalis from Brittany (France): An analysis from nutritional, chemotaxonomic, and antiproliferative activity perspectives. Mar. Drugs 2015, 13, 5606–5628. [Google Scholar] [CrossRef] [PubMed]
- Khotimchenko, S.V. Lipids from the marine alga Gracilaria verrucosa. Chem. Nat. Compd. 2005, 41, 285–288. [Google Scholar] [CrossRef]
- Cardoso, C.; Ripol, A.; Cláudia Afonso, C.; Freire, M.; Varela, J.; Quental-Ferreira, H.; Pousão-Ferreira, P.; Bandarra, N. Fatty acid profiles of the main lipid classes of green seaweeds from fish pond aquaculture. Food Sci. Nutr. 2017, 5, 1186–1194. [Google Scholar] [CrossRef] [PubMed]
- Abdullah, N.S.; Muhamad, S.; Che Omar, I.C.; Abdullah, H. Fatty acids profiles of red seaweed, Gracilaria manilaensis. Exp. 2013, 11, 726–732. [Google Scholar]
- Seca, A.M.L.; Gouveia, V.L.M.; Barreto, C.; Silva, A.M.S.; Pinto, D.C.G.A. Comparative study by GC-MS and chemometrics on the chemical and nutritional profile of Fucus spiralis L. juvenile and mature life-cycle phases. J. Appl. Phycol. 2018, 30, 2539–2548. [Google Scholar] [CrossRef]
- Venkatesan, J.; Lowe, B.; Anil, S.; Manivasagan, P.; Al Kheraif, A.A.; Kang, K.H.; Kim, S.K. Seaweed polysaccharides and their potential biomedical applications. Starch 2015, 67, 381–390. [Google Scholar] [CrossRef]
- Tanna, B.; Mishra, A. Metabolites unravel nutraceutical potential of edible seaweeds: An emerging source of functional food. Compr. Rev. Food Sci. Food Saf. 2018, 17, 1613–1624. [Google Scholar] [CrossRef] [Green Version]
- Hsieh, M.H.; Chan, P.; Sue, Y.M.; Liu, J.C.; Liang, T.H.; Huang, T.Y.; Tomlinson, B.; Chow, M.S.S.; Kao, P.F.; Chen, Y.J. Efficacy and Tolerability of Oral Stevioside in Patients with Mild Essential Hypertension: A Two-Year, Randomized, Placebo-Controlled Study. Clin. Ther. 2003, 25, 2797–2808. [Google Scholar] [CrossRef] [Green Version]
- Cardoso, S.M.; Pereira, O.R.; Seca, A.M.L.; Pinto, D.C.G.A.; Silva, A.M.S. Seaweeds as preventive agents for cardiovascular diseases: From nutrients to functional foods. Mar. Drugs 2015, 13, 6838–6865. [Google Scholar] [CrossRef] [Green Version]
- Øverland, M.; Mydland, L.T.; Skrede, A. Marine macroalgae as a source of protein and bioactive compounds in feed for monogastric animals. J. Sci. Food Agric. 2019, 99, 13–24. [Google Scholar] [CrossRef] [Green Version]
- Bruhn, A.; Janicek, T.; Manns, D.; Nielsen, M.M.; Balsby, T.J.S.; Meyer, A.S.; Rasmussen, M.B.; Hou, X.; Saake, B.; Göke, C.; et al. Crude fucoidan content in two North Atlantic kelp species, Saccharina latissima and Laminaria digitata—seasonal variation and impact of environmental factors. J. Appl. Phycol. 2017, 29, 3121–3137. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Senthilkumar, K.; Manivasagan, P.; Venkatesan, J.; Kim, S.-K. Brown seaweed fucoidan: Biological activity and apoptosis, growth signaling mechanism in cancer. Int. J. Biol. Macromol. 2013, 60, 366–374. [Google Scholar] [CrossRef] [PubMed]
- Kwak, J.-Y. Fucoidan as a marine anticancer agent in preclinical development. Mar. Drugs 2014, 12, 851–870. [Google Scholar] [CrossRef] [PubMed]
- Fitton, J.H.; Stringer, D.N.; Karpiniec, S.S. Therapies from fucoidan: An update. Mar. Drugs 2015, 13, 5920–5946. [Google Scholar] [CrossRef] [Green Version]
- Huang, C.-Y.; Kuo, C.-H.; Lee, C.-H. Antibacterial and antioxidant capacities and attenuation of lipid accumulation in 3T3-L1 adipocytes by low-molecular-weight fucoidans prepared from compressional-puffing-pretreated Sargassum crassifolium. Mar. Drugs 2018, 16. [Google Scholar]
- Phull, A.R.; Kim, S.J. Fucoidan as bio-functional molecule: Insights into the antiinflammatory potential and associated molecular mechanisms. J. Funct. Foods 2017, 38, 415–426. [Google Scholar] [CrossRef]
- Wu, L.; Sun, J.; Su, X.; Yu, Q.; Yu, Q.; Zhang, P. A review about the development of fucoidan in antitumor activity: Progress and challenges. Carbohydr. Polym. 2016, 154, 96–111. [Google Scholar] [CrossRef]
- Koyanagi, S.; Tanigawa, N.; Nakagawa, H.; Soeda, S.; Shimeno, H. Oversulfation of fucoidan enhances its anti-angiogenic and antitumor activities. Biochem. Pharmacol. 2003, 65, 173–179. [Google Scholar] [CrossRef]
- Ale, M.T.; Mikkelsen, J.D.; Meyer, A.S. mportant determinants for fucoidan bioactivity: A critical review of structure-function relations and extraction methods for fucose-containing sulfated polysaccharides from brown seaweeds. Mar. Drugs 2011, 9, 2106–2130. [Google Scholar] [CrossRef] [Green Version]
- Wijesinghe, W.A.J.P.; Jeon, Y.-J. Biological activities and potential cosmeceutical applications of bioactive components from brown seaweeds: a review. Phytochem. Rev. 2011, 10, 431–443. [Google Scholar] [CrossRef]
- Shanura Fernando, I.P.; Asanka Sanjeewa, K.K.; Samarakoon, K.W.; Kim, H.-S.; Gunasekara, U.K.D.S.S.; Park, Y.-J.; Abeytunga, D.T.U.; Lee, W.W.; Jeon, Y.-J. The potential of fucoidans from Chnoospora minima and Sargassum polycystum in cosmetics: antioxidant, anti-inflammatory, skin-whitening, and antiwrinkle activities. J. Appl. Phycol. 2018, 30, 3223–3232. [Google Scholar] [CrossRef]
- Pereira, L. Seaweeds as source of bioactive substances and skin care therapy: Cosmeceuticals, algotheraphy, and thalassotherapy. Cosmetics 2018, 5, 68. [Google Scholar] [CrossRef] [Green Version]
- Gioele, C.; Marilena, S.; Valbona, A.; Nunziacarla, S.; Andrea, S.; Antonio, M. Gracilaria gracilis, source of agar: A short review. Curr. Org. Chem. 2017, 21, 380–386. [Google Scholar] [CrossRef]
- Gu, Y.; Cheong, K.-L.; Du, H. Modification and comparison of three Gracilaria spp. agarose with methylation for promotion of its gelling properties. Chem. Cent. J. 2017, 11. [Google Scholar] [CrossRef] [Green Version]
- Martín, L.A.; Rodriguez, M.C.; Matulewicz, M.C.; Fissore, E.N.; Gerschenson, L.N.; Leonardi, P.I. Seasonal variation in agar composition and properties from Gracilaria gracilis (Gracilariales, Rhodophyta) of the Patagonian coast of Argentina. Phycol. Res. 2013, 61, 163–171. [Google Scholar] [CrossRef]
- Lee, W.K.; Lim, Y.Y.; Leow, A.T.C.; Namasivayam, P.; Abdullah, J.O.; Ho, C.L. Factors affecting yield and gelling properties of agar. J. Appl. Phycol. 2016, 29, 1527–1540. [Google Scholar] [CrossRef]
- Lee, W.-K.; Lim, Y.-Y.; Leow, A.T.-C.; Namasivayam, P.; Abdullah, J.O.; Ho, C.-L. Biosynthesis of agar in red seaweeds: A review. Carbohydr. Polym. 2017, 164, 23–30. [Google Scholar] [CrossRef]
- Guan, J.; Li, L.; Mao, S. Applications of carrageenan in advanced drug delivery. In Seaweed Polysaccharides: Isolation, Biological and Biomedical Applications; Venkatesan, J., Anil, S., Kim, S.-K., Eds.; Elsevier: Cambridge, UK, 2017; pp. 283–303. [Google Scholar]
- McKim, J.M. Food additive carrageenan: Part I: A critical review of carrageenan in vitro studies, potential pitfalls, and implications for human health and safety. Crit. Rev. Toxicol. 2014, 44, 211–243. [Google Scholar] [CrossRef]
- Cunha, L.; Grenha, A. Sulfated seaweed polysaccharides as multifunctional materials in drug delivery applications. Mar. Drugs 2016, 14, 42. [Google Scholar] [CrossRef]
- Featherstone, S. Ingredients used in the preparation of canned foods. In A Complete Course in Canning and Related Processes. Microbiology, Packaging, HACCP and Ingredients; Elsevier: Amsterdam, The Netherlands, 2015; pp. 147–211. [Google Scholar]
- David, S.; Shani Levi, C.; Fahoum, L.; Ungar, Y.; Meyron-Holtz, E.G.; Shpigelman, A.; Lesmes, U. Revisiting the carrageenan controversy: do we really understand the digestive fate and safety of carrageenan in our foods? Food Funct. 2018, 9, 1344–1352. [Google Scholar] [CrossRef]
- Pangestuti, R.; Kim, S.K. Biological activities of carrageenan. Adv. Food Nutr. Res. 2014, 72, 113–124. [Google Scholar] [PubMed]
- Souza, R.B.; Frota, A.F.; Silva, J.; Alves, C.; Neugebauer, A.Z.; Pinteus, S.; Rodrigues, J.A.G.; Cordeiro, E.M.S.; de Almeida, R.R.; Pedrosa, R.; et al. In vitro activities of kappa-carrageenan isolated from red marine alga Hypnea musciformis: Antimicrobial, anticancer and neuroprotective potential. Int. J. Biol. Macromol. 2018, 112, 1248–1256. [Google Scholar] [CrossRef] [PubMed]
- Lahaye, M.; Robic, A. Structure and functional properties of ulvan, a polysaccharide from green seaweeds. Biomacromolecules 2007, 8, 1765–1774. [Google Scholar] [CrossRef] [PubMed]
- Robic, A.; Bertrand, D.; Sassi, J.F.; Lerat, Y.; Lahaye, M. Determination of the chemical composition of ulvan, a cell wall polysaccharide from Ulva spp. (Ulvales, Chlorophyta) by FT-IR and chemometrics. J. Appl. Phycol. 2009, 21, 451–456. [Google Scholar] [CrossRef]
- Kim, J.K.; Cho, M.L.; Karnjanapratum, S.; Shin, I.S.; You, S.G. In vitro and in vivo immunomodulatory activity of sulfated polysaccharides from Enteromorpha prolifera. Int. J. Biol. Macromol. 2011, 49, 1051–1058. [Google Scholar] [CrossRef] [PubMed]
- Tabarsa, M.; Han, J.H.; Kim, C.Y.; You, S.G. Molecular characteristics and immunomodulatory activities of water-soluble sulfated polysaccharides from Ulva pertusa. J. Med. Food 2012, 15, 135–144. [Google Scholar] [CrossRef]
- Li, N.; Liu, X.; He, X.; Wang, S.; Cao, S.; Xia, Z.; Xian, H.; Qin, L.; Mao, W. Structure and anticoagulant property of a sulfated polysaccharide isolated from the green seaweed Monostroma angicava. Carbohydr. Polym. 2017, 159, 195–206. [Google Scholar] [CrossRef]
- Li, X.; Xiong, F.; Liu, Y.; Liu, F.; Hao, Z.; Chen, H. Total fractionation and characterization of the water-soluble polysaccharides isolated from Enteromorpha intestinalis. Int. J. Biol. Macromol. 2018, 111, 319–325. [Google Scholar] [CrossRef]
- Liu, X.; Cao, S.; Qin, L.; He, M.; Sun, H.; Yang, Y.; Liu, X.; Mao, W. A sulfated heterorhamnan with novel structure isolated from the green alga Monostroma angicava. Carbohydr. Res. 2018, 466, 1–10. [Google Scholar] [CrossRef]
- Liu, X.; Hao, J.; He, X.; Wang, S.; Cao, S.; Qin, L.; Mao, W. A rhamnan-type sulfated polysaccharide with novel structure from Monostroma angicava Kjellm (Chlorophyta) and its bioactivity. Carbohydr. Polym. 2017, 173, 732–748. [Google Scholar] [CrossRef]
- Ching, S.H.; Bansal, N.; Bhandari, B. Alginate gel particles - A review of production techniques and physical properties. Crit. Rev. Food Sci. Nutr. 2017, 57, 1133–1152. [Google Scholar] [CrossRef] [PubMed]
- Lee, K.Y.; Mooney, D.J. Alginate: properties and biomedical applications. Prog. Polym. Sci. 2012, 37, 106–126. [Google Scholar] [CrossRef] [Green Version]
- Rhein-Knudsen, N.; Ale, M.T.; Ajalloueian, F.; Meyer, A.S. Characterization of alginates from Ghanaian brown seaweeds: Sargassum spp. and Padina spp. Food Hydrocoll. 2017, 71, 236–244. [Google Scholar] [CrossRef]
- Aderibigbe, B.A.; Buyana, B. Alginate in wound dressings. Pharmaceutics 2018, 10, 42. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kumar, V.; Latha, S.; Rose, M.H.; Sudha, P.N. Industrial applications of alginate. In Industrial Applications of Marine Biopolymers; Sudha, P.N., Ed.; CRC press: Boca Raton, FL, USA, 2017; pp. 545–575. [Google Scholar]
- Barzegar, M.; Erfani, F.; Jabbari, A. Chemical composition of 15 spinach (Spinacea oleracea L.) cultivars grown in Iran. Ital. J. Food Sci. 2007, 19. [Google Scholar]
- Zhang, Y.; Bao, Z.; Ye, X.; Xie, Z.; He, K.; Mergens, B.; Li, W.; Yatcilla, M.; Zheng, Q. Chemical investigation of major constituents in Aloe vera leaves and several commercial Aloe juice powders. J. AOAC Int. 2018, 101, 1741–1751. [Google Scholar] [CrossRef]
- Graiff, A.; Ruth, W.; Kragl, U.; Karsten, U. Chemical characterization and quantification of the brown algal storage compound laminarin — A new methodological approach. J. Appl. Phycol. 2016, 28, 533–543. [Google Scholar] [CrossRef]
- Rioux, L.-E.; Turgeon, S.L.; Beaulieu, M. Characterization of polysaccharides extracted from brown seaweeds. Carbohydr. Polym. 2007, 69, 530–537. [Google Scholar] [CrossRef]
- Nadeem, M.; Imran, M.; Khalique, A. Promising features of mango (Mangifera indica L.) kernel oil: a review. J. Food Sci. Technol. 2016, 53, 2185–2195. [Google Scholar] [CrossRef] [Green Version]
- Circuncisão, A.R.; Catarino, M.D.; Cardoso, S.M.; Silva, A.M.S. Minerals from macroalgae origin: Health benefits and risks for consumers. Mar. Drugs 2018, 16, 400. [Google Scholar] [CrossRef] [Green Version]
- USDA National Nutrient Database for Standard Reference. Available online: http://www.nal.usda.gov/fnic/foodcomp (accessed on 31 January 2019).
- Whelton, P.K. Sodium, potassium, blood pressure, and cardiovascular disease in humans. Curr. Hypertens. Rep. 2014, 16, e465. [Google Scholar] [CrossRef] [PubMed]
- Newberry, S.J.; Chung, M.; Anderson, C.A.M.; Chen, C.; Fu, Z.; Tang, A.; Zhao, N.; Booth, M.; Marks, J.; Hollands, S.; et al. Sodium and Potassium Intake: Effects on Chronic Disease Outcomes and Risks; AHRQ Comparative Effectiveness Reviews: Rockville, MD, USA, 2018.
- Then, M.; Szentmihályi, K.; Sárközi, A.; Illés, V.; Forgács, E. Effect of sample handling on alkaloid and mineral content of aqueous extracts of greater celandine (Chelidonium majus L.). J. Chromatogr. A 2000, 889, 69–74. [Google Scholar] [CrossRef]
- Beto, J.A. The role of calcium in human aging. Clin. Nutr. Res. 2015, 4, 1–8. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Baker, S.B.; Worthley, L.I. The essentials of calcium, magnesium and phosphate metabolism: part I. Physiology. Crit. Care Resusc. 2002, 4, 301–306. [Google Scholar] [PubMed]
- Loughrill, E.; Wray, D.; Christides, T.; Zand, N. Calcium to phosphorus ratio, essential elements and vitamin D content of infant foods in the UK: Possible implications for bone health. Matern. Child Nutr. 2017, 13, e12368. [Google Scholar] [CrossRef] [Green Version]
- Desideri, D.; Cantaluppi, C.; Ceccotto, F.; Meli, M.A.; Roselli, C.; Feduzi, L. Essential and toxic elements in seaweeds for human consumption. J. Toxicol. Environ. Heal. Part A 2016, 79, 112–122. [Google Scholar] [CrossRef]
- McCall, A.; Cummings, C.; Bhave, G.; Vanacore, R.; Page-McCaw, A.; Hudson, B.G. Bromine is an essential trace element for assembly of collagen IV scaffolds in tissue development and architecture. Cell 2014, 157, 1380–1392. [Google Scholar] [CrossRef] [Green Version]
- Rey-Crespo, F.; López-Alconso, M.; Miranda, M. The use of seaweed from the Galician coast as a mineral supplement in organic dairy cattle. Animal 2014, 8, 580–586. [Google Scholar] [CrossRef]
- Rajapakse, N.; Kim, S.K. Nutritional and digestive health benefits of seaweed. Adv. Food Nutr. Res. 2011, 64, 17–28. [Google Scholar]
- Rhee, S.S.; Braverman, L.E.; Pino, S.; He, X.; Pearce, E.N. High iodine content of Korean seaweed soup: A health risk for lactating women and their infants? Thyroid 2011, 21, 927–928. [Google Scholar] [CrossRef] [Green Version]
- Kadam, S.U.; Álvarez, C.; Tiwari, B.K.; O’Donnell, C.P. Extraction and characterization of protein from Irish brown seaweed Ascophyllum nodosum. Food Res. Int. 2016, 99, 1021–1027. [Google Scholar] [CrossRef] [PubMed]
- Harrysson, H.; Hayes, M.; Eimer, F.; Carlsson, N.G.; Toth, G.B.; Undeland, I. Production of protein extracts from Swedish red, green, and brown seaweeds, Porphyra umbilicalis Kutzing, Ulva lactuca Linnaeus, and Saccharina latissima (Linnaeus) J. V. Lamouroux using three different methods. J. Appl. Phycol. 2018, 30, 3565–3580. [Google Scholar] [CrossRef] [Green Version]
- Angell, A.R.; Mata, L.; de Nys, R.; Paul, N.A. The protein content of seaweeds: a universal nitrogen-to-protein conversion factor of five. J. Appl. Phycol. 2016, 28, 511–524. [Google Scholar] [CrossRef]
- Lourenço, S.O.; Barbarino, E.; De-Paula, J.C.; Pereira, L.O.; Marquez, U.M.L. Amino acid composition, protein content and calculation of nitrogen-to-protein conversion factors for 19 tropical seaweeds. Phycol. Res. 2002, 50, 233–241. [Google Scholar] [CrossRef]
- Bajpai, V.K. Korean seaweeds as a food of future: An update on use and risk factors. Indian J. Geomarine Sci. 2017, 46, 1253–1260. [Google Scholar]
- Mæhre, H.K.; Malde, M.K.; Eilertsen, K.E.; Elvevoll, E.O. Characterization of protein, lipid and mineral contents in common Norwegian seaweeds and evaluation of their potential as food and feed. J. Sci. Food Agric. 2014, 94, 3281–3290. [Google Scholar] [CrossRef]
- Marinho, G.S.; Holdt, S.L.; Angelidaki, I. Seasonal variations in the amino acid profile and protein nutritional value of Saccharina latissima cultivated in a commercial IMTA system. J. Appl. Phycol. 2015, 27, 1991–2000. [Google Scholar] [CrossRef]
- Ariede, M.B.; Candido, T.M.; Jacome, A.L.M.; Velasco, M.V.R.; Carvalho, J.C.M.; Baby, A.R. Cosmetic attributes of algae – A review. Algal Res. 2017, 25, 483–487. [Google Scholar] [CrossRef]
- Admassu, H.; Gasmalla, M.A.A.; Yang, R.; Zhao, W. Bioactive peptides derived from seaweed protein and their health benefits: Antihypertensive, antioxidant, and antidiabetic properties. J. Food Sci. 2018, 83, 6–16. [Google Scholar] [CrossRef] [Green Version]
- Hagino, H.; Saito, M. Use of Algal Proteins in Cosmetics. European Patent EP1433463B1, 18 December 2010. [Google Scholar]
- Hong, D.D.; Hien, H.T.M. Nutritional analysis of Vietnamese seaweeds for food and medicine. BioFactors 2004, 22, 323–325. [Google Scholar] [CrossRef]
- Lopes, G.; Pinto, E.; Andrade, P.B.; Valentão, P. Antifungal activity of phlorotannins against dermatophytes and yeasts: approaches to the mechanism of action and influence on Candida albicans virulence factor. PLoS One 2013, 8, 1–10. [Google Scholar] [CrossRef]
- Miyashita, K.; Mikami, N.; Hosokawa, M. Chemical and nutritional characteristics of brown seaweed lipids: A review. J. Funct. Foods 2013, 5, 1507–1517. [Google Scholar] [CrossRef]
- Sinurat, E.; Fadjriah, S. The Chemical Properties of Seaweed Caulerpa lentifera from Takalar, South Sulawesi. In IOP Conference Series: Materials Science and Engineering; IOP Publishing: Bristol, UK, 2019; pp. 546, 042043. [Google Scholar]
- Tang, Y.H.; Jiang, Y.; Meng, J.S.; Tao, J. A brief review of physiological roles, plant resources, synthesis, purification and oxidative stability of alpha-linolenic acid. Emirates J. Food Agric. 2018, 30, 341–356. [Google Scholar]
- Gammone, M.A.; Riccioni, G.; Parrinello, G.; D’Orazio, N. Omega-3 polyunsaturated fatty acids: Benefits and endpoints in sport. Nutrients 2019, 11, 46. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Simopoulos, A.P.; DiNicolantonio, J.J. The importance of a balanced ω-6 to ω-3 ratio in the prevention and management of obesity. Open Hear. 2016, 3. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ahmad, M.; Nangyal, H.; Sherwani, S.K.; Islam, Z.; Shah, S.H. Effect of heat stress on fatty acids profiles of Aloe vera and Bryophyllum pinnatum leaves. World Appl. Sci. J. 2013, 28, 1592–1596. [Google Scholar]
- Nadeem, M.; Anjum, F.M.; Khan, M.I.; Tehseen, S.; El-Ghorab, A.; Sultan, J.I. Nutritional and medicinal aspects of coriander (Coriandrum sativum L.): A review. Br. Food J. 2013, 115, 743–755. [Google Scholar] [CrossRef]
- Mikami, K.; Hosokawa, M. Biosynthetic pathway and health benefits of fucoxanthin, an algae-specific xanthophyll in brown seaweeds. Int. J. Mol. Sci. 2013, 14, 13763–13781. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Terasaki, M.; Kawagoe, C.; Ito, A.; Kumon, H.; Narayan, B.; Hosokawa, M.; Miyashita, K. Spatial and seasonal variations in the biofunctional lipid substances (fucoxanthin and fucosterol) of the laboratory-grown edible Japanese seaweed (Sargassum horneri Turner) cultured in the open sea. Saudi J. Biol. Sci. 2017, 24, 1475–1482. [Google Scholar] [CrossRef]
- Gammone, M.A.; Riccioni, G.; D’Orazio, N. Carotenoids: Potential allies of cardiovascular health? Food Nutr. Res. 2015, 59. [Google Scholar]
- Carvalhal, F.; Correia-da-Silva, M.; Sousa, E.; Pinto, M.; Kijjoa, A. Sources and biological activities of marine sulfated steroids. J. Mol. Endocrinol. 2018, 61, 211–231. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Li, X.-C.; Jacob, M.R.; Ding, Y.; Agarwal, A.K.; Smillie, T.J.; Khan, S.I.; Nagle, D.G.; Ferreira, D.; Clark, A.M. Capisterones A and B, which enhance fluconazole activity in Saccharomyces cerevisiae, from the marine green alga Penicillus capitatus. J. Nat. Prod. 2006, 69, 542–546. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ortiz, J.; Romero, N.; Robert, P.; Araya, J.; Lopez-Hernández, J.; Bozzo, C.; Navarrete, E.; Osorio, A.; Rios, A. Dietary fiber, amino acid, fatty acid and tocopherol contents of the edible seaweeds Ulva lactuca and Durvillaea antarctica. Food Chem. 2006, 99, 98–104. [Google Scholar] [CrossRef]
- Faulkner, D.J. Interesting aspects of marine natural products chemistry. Tetrahedron Lett. 1977, 33, 1421–1443. [Google Scholar] [CrossRef]
- Faulkner, D.J. Marine natural products: metabolites of marine algae and herbivorous marine molluscs. Nat. Prod. Rep. 1984, 1, 251–280. [Google Scholar] [CrossRef]
- Faulkner, D.J. Marine natural products. Nat. Prod. Rep. 2002, 19, 1–49. [Google Scholar]
- Blunt, J.W.; Copp, B.R.; Munro, M.H.G.; Northcote, P.T.; Prinsep, M.R. Marine natural products. Nat. Prod. Rep. 2003, 20, 1–48. [Google Scholar] [CrossRef]
- Blunt, J.W.; Carroll, A.R.; Copp, B.R.; Davis, R.A.; Keyzers, R.A.; Prinsep, M.R. Marine natural products. Nat. Prod. Rep. 2018, 35, 8–53. [Google Scholar] [CrossRef] [Green Version]
- Blunt, J.W.; Copp, B.R.; Munro, M.H.G.; Northcote, P.T.; Prinsep, M.R. Marine natural products. Nat. Prod. Rep. 2010, 27, 165–237. [Google Scholar] [CrossRef]
- Brodie, J.; Lewis, J. Unravelling the Algae: The past, present and future of algal systematics; CRC Press (Taylor & Francis Group): Boca Raton, FL, USA, 2007. [Google Scholar]
- Falkenberg, M.; Nakano, E.; Zambotti-Villela, L.; Zatelli, G.A.; Philippus, A.C.; Imamura, K.B.; Velasquez, A.M.A.; Freitas, R.P.; Tallarico, L.F.; Colepicolo, P.; et al. Bioactive compounds against neglected diseases isolated from macroalgae: a review. J. Appl. Phycol. 2018, 30, 1–27. [Google Scholar] [CrossRef] [Green Version]
- Blunt, J.W.; Copp, B.R.; Munro, M.H.G.; Northcote, P.T.; Prinsep, M.R. Marine natural products. Nat. Prod. Rep. 2011, 28, 196–268. [Google Scholar] [CrossRef] [PubMed]
- Blunt, J.W.; Copp, B.R.; Keyzers, R.A.; Munro, M.H.G.; Prinsep, M.R. Marine natural products. Nat. Prod. Rep. 2012, 29, 144–222. [Google Scholar] [CrossRef] [PubMed]
- Blunt, J.W.; Copp, B.R.; Keyzers, R.A.; Munro, M.H.G.; Prinsep, M.R. Marine natural products. Nat. Prod. Rep. 2013, 30, 237–323. [Google Scholar] [CrossRef] [PubMed]
- Blunt, J.W.; Copp, B.R.; Keyzers, R.A.; Munro, M.H.G.; Prinsep, M.R. Marine natural products. Nat. Prod. Rep. 2014, 31, 160–258. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Blunt, J.W.; Copp, B.R.; Keyzers, R.A.; Munro, M.H.G.; Prinsep, M.R. Marine natural products. Nat. Prod. Rep. 2015, 32, 116–211. [Google Scholar] [CrossRef] [Green Version]
- Blunt, J.W.; Copp, B.R.; Keyzers, R.A.; Munro, M.H.G.; Prinsep, M.R. Marine natural products. Nat. Prod. Rep. 2016, 33, 382–411. [Google Scholar] [CrossRef] [Green Version]
- Blunt, J.W.; Copp, B.R.; Keyzers, R.A.; Munro, M.H.G.; Prinsep, M.R. Marine natural products. Nat. Prod. Rep. 2017, 34, 235–294. [Google Scholar] [CrossRef] [Green Version]
- Blackman, A.J.; Wells, R.J. Flexilin and trifarin, terpene 1,4-diacetoxybuta-1,3-dienes from two Caulerpa species (Chlorophyta). Tetrahedron Lett. 1978, 19, 3063–3064. [Google Scholar] [CrossRef]
- Amico, V.; Oriente, G.; Piattelli, M.; Tringali, C.; Fattorusso, E.; Magno, S.; Mayol, L. Caulerpenyne, na unusual sesquiterpenoid from the green alga Caulerpa prolifera. Tetrahedron Lett. 1978, 19, 3593–3596. [Google Scholar] [CrossRef]
- Sun, H.H.; Fenical, W. Rhipocephalin and rhipocephenal; toxic feeding deterrents from the tropical marine alga. Tetrahedron Lett. 1979, 20, 685–688. [Google Scholar] [CrossRef]
- Paul, V.J.; Sun, H.H.; Fenical, W. Udoteal, a linear diterpenoid feeding deterrent from the tropical green alga Udotea flabellum. Phytochemistry 1982, 21, 468–469. [Google Scholar] [CrossRef]
- Paul, V.J.; Fenical, W. Chemical defense in tropical green algae, order Caulerpales. Mar. Ecol. Prog. Ser. 1986, 34, 157–169. [Google Scholar] [CrossRef]
- Paul, V.J.; Littler, M.M.; Littler, D.S.; Fenical, W. Evidence for chemical defense in tropical green alga Caulerpa ashmeadii (Caulerpaceae: Chlorophyta): Isolation of new bioactive sesquiterpenoids. J. Chem. Ecol. 1987, 13, 1171–1185. [Google Scholar] [CrossRef] [PubMed]
- Capon, R.J.; Ghisalberti, E.L.; Jefferies, P.R. Metabolites of the green algae, Caulerpa species. Phytochemistry 1983, 22, 1465–1467. [Google Scholar] [CrossRef]
- Guerriero, A.; Meinesz, A.; D’Ambrosio, M.; Pietra, F. Isolation of toxic and potentially toxic sesqui- and monoterpenes from the tropical green seaweed Caulerpa taxifolia which has invaded the region of Cap Martin and Monaco. Helv. Chim. Acta 1992, 75, 689–695. [Google Scholar] [CrossRef]
- Paul, V.J.; Fenical, W. Isolation of halimedatrial: Chemical defense adaptation in the calcareous reef-building alga Halimeda. Science (80-. ). 1983, 221, 747–749. [Google Scholar] [CrossRef]
- Paul, V.J.; Fenical, W. Novel bioactive diterpenoid metabolites from tropical marine algae of the genus Halimeda (Chlorophyta). Tetrahedron Lett. 1984, 40, 3053–3062. [Google Scholar] [CrossRef]
- Paul, V.J.; Fenical, W. Bioactive terpenoids from caribbean marine algae of the genera Penicillus and Udotea (chlorophyta). Tetrahedron Lett. 1984, 40, 2013–2918. [Google Scholar] [CrossRef]
- Paul, V.J.; Fenical, W.; Raffii, S.; Clardy, J. The isolation of new norcycloartene triterpenoids from the tropical marine alga (chlorophyta). Tetrahedron Lett. 1982, 23, 3459–3462. [Google Scholar] [CrossRef]
- Napoli, L.; Fattorusso, E.; Magno, S.; Mayol, L. The occurrence of (S)-(−)-squalene-2,3-epoxide in nature: Isolation from the green alga. Tetrahedron Lett. 1980, 21, 2917–2918. [Google Scholar] [CrossRef]
- Napoli, L.; Fattorusso, E.; Magno, S.; Mayol, L. Three squalene derivatives from Caulerpa prolifera. Phytochemistry 1982, 21, 782–784. [Google Scholar] [CrossRef]
- Hamann, M.T.; Otto, C.; Scheuer, P.J.; Dunbar, D.C. Kahalalides: Bioactive peptides from a marine mollusk Elysia rufescens and its algal diet Bryopsis sp. J. Org. Chem. 1996, 61, 6594–6600. [Google Scholar] [CrossRef] [PubMed]
- Hamann, M.T.; Scheuer, P.J. Kahalalide F: a bioactive depsipeptide from the sacoglossan mollusk Elysia rufescens and the green alga Bryopsis sp. J. Am. Chem. Soc. 1993, 115, 5825–5826. [Google Scholar] [CrossRef]
- López-Macià, À.; Jiménez, J.C.; Royo, M.; Giralt, E.; Albericio, F. Synthesis and structure determination of kahalalide F. J. Am. Chem. Soc. 2001, 123, 11398–11401. [Google Scholar] [CrossRef]
- Bonnard, I.; Manzanares, I.; Rinehart, K.L. Stereochemistry of kahalalide F. J. Nat. Prod. 2003, 66, 1466–1470. [Google Scholar] [CrossRef]
- Goetz, G.; Yoshida, W.Y.; Scheuer, P.J. The absolute stereochemistry of kahalalide F. Tetrahedron Lett. 1999, 55, 7739–7746. [Google Scholar] [CrossRef]
- Gao, J.; Caballero-George, C.; Wang, B.; Rao, K.V.; Shilabin, A.G.; Hamann, M.T. 5-OHKF and NorKA, depsipeptides from a Hawaiian collection of Bryopsis pennata: Binding properties for NorKA to the Human neuropeptide Y Y1 receptor. J. Nat. Prod. 2009, 72, 2172–2176. [Google Scholar] [CrossRef] [Green Version]
- Dmitrenok, A.; Iwashita, T.; Nakajima, T.; Sakamoto, B.; Namikoshi, M.; Nagai, H. New cyclic depsipeptides from the green alaga Bryopsis species; application of a carboxypeptidase hydrolysis reaction to the structure determination. Tetrahedron Lett. 2006, 62, 1301–1308. [Google Scholar] [CrossRef]
- Iwasaki, A.; Ohno, O.; Sumimoto, S.; Matsubara, T.; Shimada, S.; Sato, T.; Suenaga, K. Mebamamides A and B, cyclic lipopeptides isolated from the green alga Derbesia marina. J. Nat. Prod. 2015, 78, 901–908. [Google Scholar] [CrossRef]
- Jesus, A.; Correia-da-Silva, M.; Afonso, C.; Pinto, M.; Cidade, H. Isolation and potential biological applications of haloaryl secondary metabolites from macroalgae. Mar. Drugs 2019, 17, 73. [Google Scholar] [CrossRef] [Green Version]
- McConnell, O.J.; Hughes, P.A.; Targett, N.M. Diastereoisomers of cyclocymopol and cyclocymopol monomethyl ether from Cymopolia barbata. Phytochemistry 1982, 21, 2139–2141. [Google Scholar] [CrossRef]
- Estrada, D.M.; Martín, J.D.; Pérez, C. A new brominated monoterpenoid from Cymopolia barbata. J. Nat. Prod. 1987, 50, 735–737. [Google Scholar] [CrossRef]
- Wall, M.E.; Wani, M.C.; Manikumar, G.; Taylor, H.; Hughes, T.J.; Gaetano, K.; Gerwick, W.H.; McPhail, A.T.; McPhail, D.R. Plant antimutagenic agents, 7. Structure and antimutagenic properties of cymobarbatol and 4-isocymobarbatol, new cymopols from green alga (Cymopolia barbata). J. Nat. Prod. 1989, 52, 1092–1099. [Google Scholar] [CrossRef] [PubMed]
- Park, M.; Fenical, W.; Hay, M.E. Debromoisocymobarbatol, a new chromanol feeding deterrent from the marine alga Cymopolia barbata. Phytochemistry 1992, 31, 4115–4118. [Google Scholar] [CrossRef]
- Sun, H.H.; Paul, V.J.; Fenical, W. Avrainvilleol, a brominated diphenylmethane derivative with feeding deterrent properties from the tropical green alga Avrainvillea longicaulis. Phytochemistry 1983, 22, 743–745. [Google Scholar] [CrossRef]
- Carte, B.K.; Troupe, N.; Chan, J.A.; Westley, J.W.; Faulkner, D.J. Rawsonol, an inhibitor of HMG-CoA reductase from the tropical green alga Avrainvillea rawsoni. Phytochemistry 1989, 28, 2917–2919. [Google Scholar] [CrossRef]
- Chkhikvishvili, I.D.; Ramazanov, Z.M. Phenolic substances of brown algae and their antioxidant activity. Appl. Biochem. Microbiol. 1999, 36, 289–291. [Google Scholar] [CrossRef]
- Amsler, C.D.; Fairhead, V.A. Defensive and sensory chemical ecology of brown algae. Adv. Bot. Res. 2005, 43, 1–91. [Google Scholar]
- Lopes, G.; Sousa, C.; Silva, L.R.; Pinto, E.; Andrade, P.B.; Bernardo, J.; Mouga, T.; Valentão, P. Can phlorotannins purified extracts constitute a novel pharmacological alternative for microbial infections with associated inflammatory conditions? PLoS One 2012, 7. [Google Scholar] [CrossRef]
- Singh, I.P.; Sidana, J. Phlorotannins. In Functional ingredients from algae for foods and nutraceuticals; Woodhead Publishing, Ed.; Elsevier B.V: Sawston, Cambridge, 2013; pp. 181–204. [Google Scholar]
- Kirke, D.A.; Smyth, T.J.; Rai, D.K.; Kenny, O.; Stengel, D.B. The chemical and antioxidant stability of isolated low molecular weight phlorotannins. Food Chem. 2017, 221, 1104–1112. [Google Scholar] [CrossRef]
- Brunt, E.G.; Burgess, J.G. The promise of marine molecules as cosmetic active ingredients. Int. J. Cosmet. Sci. 2018, 40, 1–15. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Vallim, M.A.; De Paula, J.C.; Pereira, R.C.; Teixeira, V.L. The diterpenes from Dictytacean marine brown algae in the tropical Atlantic American region. Biochem. Syst. Ecol. 2005, 33, 1–16. [Google Scholar] [CrossRef]
- Chen, J.; Li, H.; Zhao, Z.; Xia, X.; Li, B.; Zhang, J.; Yan, X. Diterpenes from marine algae of the genus Dictyota. Mar. Drugs 2018, 16, 159. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gouveia, V.; Seca, A.M.L.; Barreto, M.C.; Pinto, D.C.G.A. Di- and sesquiterpenoids from Cystoseira genus: structure, intra-molecular transformations and biological activity. Mini-reviews Med. Chem. 2013, 13, 1150–1159. [Google Scholar] [CrossRef] [PubMed]
- Harper, M.K.; Bugni, T.S.; Copp, B.R.; James, R.D.; Lindsay, B.S.; Richardson, A.D.; Schnabel, P.C.; Tasdemir, D.; VanWagoner, R.M.; Verbitzki, S.M.; et al. Introduction to the chemical ecology of marine natural products. In Marine Chemical Ecology; McClintock, J.B., Baker, B.J., Eds.; CRC press: Boca Raton, FL, USA, 2001; pp. 3–70. [Google Scholar]
- Nylund, G.M.; Persson, F.; Lindegarth, M.; Cervin, G.; Hermansson, M.; Pavia, H. The red alga Bonnemaisonia asparagoides regulates epiphytic bacterial abundance and community composition by chemical defence. FEMS Microbiol. Ecol. 2010, 71, 84–93. [Google Scholar] [CrossRef] [Green Version]
- Salvador, N.; Gomez-Garreta, A.; Lavelli, L.; Ribera, M.A. Antimicrobial Activity of Iberian Macroalgae. Sci. Mar. 2007, 71, 101–113. [Google Scholar] [CrossRef] [Green Version]
- Machado, F.L.S.; Kaiser, C.R.; Costa, S.S.; Gestinari, L.M.; Soares, A.R. Biological activity of the secondary metabolite from marine algae of the genus Laurencia. Brazilian J. Pharmacogn. 2010, 20, 441–452. [Google Scholar] [CrossRef]
- Cabrita, M.T.; Vale, C.; Rauter, A.P. Halogenated compounds from marine algae. Mar. Drugs 2010, 8, 2301–2317. [Google Scholar] [CrossRef] [Green Version]
- Irie, T.; Suzuki, M.; Kurosawa, E.; Masamune, T. Laurinterol, debromolaurinterol and isolaurinterol, constituents of Laurencia intermedia Yamada. Tetrahedron Lett. 1970, 26, 3271–3277. [Google Scholar] [CrossRef]
- Fronczek, F.R.; Caccamese, S. Redetermination of pacefenol, a halogenated sesquiterpene from Mediterranean red alga Laurencia majuscula. Acta Crystallogr. 1986, 42, 1649–1651. [Google Scholar]
- Santos, A.O.; Veiga-Santos, P.; Ueda-Nakamura, T.; Filho, B.P.D.; Sudatti, D.B.; Bianco, É.M.; Pereira, R.C.; Nakamura, C.V. Effect of elatol, isolated from red seaweed Laurencia dendroidea, on Leishmania amazonensis. Mar. Drugs 2010, 8, 2733–2743. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Howard, B.M.; Fenical, W. Structures of the irieols, new dibromoditerpenoids of a unique skeletal class from the marine red alga Laurencia irieii. J. Org. Chem. 1978, 43, 4401–4408. [Google Scholar] [CrossRef]
- Suzuki, M.; Matsuo, Y.; Takahashi, Y.; Masuda, M. Callicladol, a novel cytotoxic bromotriterpene polyether from a Vietnamese species of the red algal genus Laurencia. Chem. Lett. 1995, 24, 1045–1046. [Google Scholar] [CrossRef]
- Wanke, T.; Philippus, A.C.; Zatelli, G.A.; Vieira, L.F.O.; Lhullier, C.; Falkenberg, M. C15 acetogenins from the Laurencia complex: 50 years of research – an overview. Brazilian J. Pharmacogn. 2015, 25, 569–578. [Google Scholar] [CrossRef] [Green Version]
- Fukuzawa, A.; Masamune, T. Laurepinnacin and isolaurepinnacin, new acetylenic cyclic ethers from the marine red alga Laurencia pinnata yamada. Tetrahedron Lett. 1981, 22, 4081–4084. [Google Scholar] [CrossRef]
- Fukuzawa, A.; Kurosawa, E. Laurallene, new bromoallene from the marine red alga Laurencia nipponica Yamada. Tetrahedron Lett. 1979, 20, 2797–2800. [Google Scholar] [CrossRef]
- Saitoh, T.; Suzuki, T.; Sugimoto, M.; Hagiwara, H.; Hoshi, T. Total synthesis of (+)-laurallene. Tetrahedron Lett. 2003, 44, 3175–3178. [Google Scholar] [CrossRef]
- Fuller, R.W.; Cardellina, J.H.; Jurek, J.; Scheuer, P.J.; Alvarado-Lindner, B.; McGuire, M.; Gray, G.N.; Steiner, J.R.; Clardy, J.; Menez, E.; et al. Isolation and structure/activity features of halomonrelated antitumor monoterpenes from the red alga Portieria hornemannii. J. Med. Chem. 1994, 37, 4407–4411. [Google Scholar] [CrossRef]
- Lane, A.L.; Mular, L.; Drenkard, E.J.; Shearer, T.L.; Engel, S.; Fredericq, S.; Fairchild, C.R.; Prudhomme, J.; Roch, K.L.; Hay, M.E.; et al. Ecological leads for natural product discovery: novel sesquiterpene hydroquinones from red macroalga Peyssonnelia sp. Tetrahedron Lett. 2010, 66, 455–461. [Google Scholar] [CrossRef] [Green Version]
- Young, R.M.; von Salm, J.L.; Amsler, M.O.; Lopez-Bautista, J.; Amsler, C.D.; McClintock, J.B.; Baker, B.J. Site-Specific Variability in the Chemical Diversity of the Antarctic Red Alga Plocamium cartilagineum. Mar. Drugs 2013, 11, 2126–2139. [Google Scholar] [CrossRef] [Green Version]
- De Nys, R.; Wright, A.D.; König, G.M.; Sticher, O. New halogenated furanones from the marine alga Delisea pulchra (cf. fimbriata). Tetrahedron Lett. 1993, 49, 11213–11220. [Google Scholar] [CrossRef]
- Rasmussen, T.B.; Manefield, M.; Andersen, J.B.; Eberl, L.; Anthoni, U.; Christophersen, C.; Steinberg, P.; Kjelleberg, S.; Givskov, M. How Delisea pulchra furanones affect quorum sensing and swarming motility in Serratia liquefaciens MG1. Microbiology 2000, 146, 3237–3244. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kjelleberg, S.; Steinberg, P. Surface waters in the sea. Microbiol. Today 2001, 28, 134–135. [Google Scholar]
- Witvrouw, M.; Reymen, D.; Haines, H.; Matulewicz, M.C.; Cerezo, A.; Coto, C.E.; De Clercq, E. Antiviral Activity of a Sulfated Polysaccharide from the Red Seaweed Nothogenia fastigiata. Biochem. Pharmacol. 1994, 47, 2187–2192. [Google Scholar]
- Khalil, H.P.S.A.; Lai, T.K.; Tye, Y.Y.; Rizal, S.; Chong, E.W.N.; Yap, S.W.; Hamzah, A.A.; Fazita, M.R.N.; Paridah, M.T. A review of extractions of seaweed hydrocolloids: Properties and applications. Express Polym. Lett. 2018, 12, 296–317. [Google Scholar] [CrossRef]
- Diogo, J.V.; Novo, S.G.; Gonzalez, M.J.; Ciancia, M.; Bratanich, A.C. Antiviral activity of lambda-carrageenan prepared from red seaweed (Gigartina skottsbergii) against BoHV-1 and SuHV-1. Res. Vet. Sci. 2015, 98, 142–144. [Google Scholar] [CrossRef]
- Jiao, G.L.; Yu, R.L.; Zhang, J.Z.; Ewart, H.S. Chemical structures and bioactivities of sulfated polysaccharides from marine algae. Mar. Drugs 2011, 9, 196–223. [Google Scholar] [CrossRef] [Green Version]
- Yamada, T.; Ogamo, A.; Saito, T.; Uchiyama, H.; Nakagawa, Y. Preparation of O-acylated low-molecular-weight carrageenans with potent anti-HIV activity and low anticoagulant effect. Carbohydr. Polym. 2000, 41, 115–120. [Google Scholar] [CrossRef]
- De Clercq, E. Current Lead Natural Products for the Chemotherapy of Human Immunodeficiency Virus (HIV) Infection. Med. Res. Rev. 2000, 20. [Google Scholar] [CrossRef]
- Wu, S.C.; Kang, S.K.; Kazlowski, B.; Wu, C.J.; Pan, C.L. Antivirus and Prebiotic Properties of Seaweed-Oligosaccharide-Lysates Derived from Agarase AS-II. J. Fish. Soc. Taiwan 2012, 39, 11–21. [Google Scholar]
- Bouhlal, R.; Haslin, C.; Chermann, J.C.; Colliec-Jouault, S.; Sinquin, C.; Simon, G.; Cerantola, S.; Riadi, H.; Bourgougnon, N. Antiviral Activities of Sulfated Polysaccharides Isolated from Sphaerococcus coronopifolius (Rhodophytha, Gigartinales) and Boergeseniella thuyoides (Rhodophyta, Ceramiales). Mar. Drugs 2011, 9, 1187–1209. [Google Scholar] [CrossRef] [PubMed]
- Cimino, P.; Bifulco, G.; Casapullo, A.; Bruno, I.; Gomez-Paloma, L.; Riccio, R. Isolation and NMR characterization of rosacelose, a novel sulfated polysaccharide from the sponge Mixylla rosacea. Carbohydr. Res. 2001, 334, 39–47. [Google Scholar] [CrossRef]
- Wang, H.; Ooi, E.V.; Ang, P.O., Jr. Antiviral activities of extracts from Hong Kong seaweeds. J. Zhejiang Univ. Sci. B 2008, 9, 969–976. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ahmadi, A.; Moghadamtousi, S.Z.; Abubakar, S.; Zandi, K. Antiviral Potential of Algae Polysaccharides Isolated from Marine Sources: A Review. Biomed Res. Int. 2015, 825203, 1–10. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Queiroz, K.C.S.; Medeiros, V.P.; Queiroz, L.S.; Abreu, L.R.; Rocha, H.A.; Ferreira, C.V.; Jucá, M.B.; Aoyama, H.; Leite, E.L. Inhibition of reverse transcriptase activity of HIV by polysaccharides of brown algae. Biomed. Pharmacother. 2008, 62, 303–307. [Google Scholar] [CrossRef] [PubMed]
- Akamatsu, E.; Shimanaga, M.; Kamei, Y. Isolation of an anti-influenza virus substance, MC26 from a marine brown alga, Sargassum piluliferum and its antiviral activity against influenza virus. Coast. Bioenvironment - Saga Univ. 2003, 1, 29–34. [Google Scholar]
- Mandal, P.; Mateu, C.G.; Chattopadhyay, K.; Pujol, C.A.; Damonte, E.B.; Ray, B. Structural features and antiviral activity of sulphated fucans from the brown seaweed Cystoseira indica. Antivir. Chem. Chemother. 2007, 18, 153–162. [Google Scholar] [CrossRef] [Green Version]
- Hidari, K.I.P.J.; Takahashi, N.; Arihara, M.; Nagaoka, M.; Morita, K.; Suzuki, T. Structure and anti-dengue virus activity of sulfated polysaccharide from a marine alga. Biochem. Biophys. Res. Commun. 2008, 376, 91–95. [Google Scholar] [CrossRef]
- Vairappan, C.S. Potent antibacterial activity of halogenated metabolites from Red algae Laurencia majuscule (Rhodomelaceae, Ceramiales). Biomol. Eng. 2003, 20, 255–259. [Google Scholar] [CrossRef]
- Vairappan, C.; Kawamoto, T.; Miwa, H.; Suzuki, M. Potent antibacterial activity of halogenated compounds against antibiotic-resistant bacteria. Planta Med. 2004, 70, 1087–1090. [Google Scholar] [CrossRef] [Green Version]
- Barreto, M.; Meyer, J.J.M. Isolation and antimicrobial activity of a lanosol derivative from Osmundaria serrata (Rhodophyta) and a visual exploration of its biofilm covering. South African J. Bot. 2006, 72, 521–528. [Google Scholar] [CrossRef] [Green Version]
- Srinivasakumar, K.P.; Rajashekhar, M. In vitro studies on bactericidal activity and sensitivity pattern of isolated marine microalgae against selective human bacterial pathogens. Indian J. Sci. Technol. 2009, 2, 58–64. [Google Scholar]
- Priyadharshini, S.; Bragadeeswaran, S.; Prabhu, K.; Rani, S.S. Antimicrobial and hemolytic activity of seaweed extracts Ulva fasciata (Delile 1813) from Mandapam, Southeast coast of India. Asian Pac. J. Trop. Biomed. 2011, S37–S39. [Google Scholar] [CrossRef]
- Kladi, M.; Vagias, C.; Stavri, M.; Mukhlesu Rahman, M.; Gibbons, S.; Roussis, V. C15 acetogenins with antistaphylococcal activity. Phytochem. Lett. 2008, 31–36. [Google Scholar] [CrossRef]
- Amiguet, V.T.; Jewell, L.E.; Mao, H.; Sharma, M.; Hudson, J.B.; Durst, T.; Allard, M.; Rochefort, G.; Arnason, J.T. Antibacterial properties of a glycolipid-rich extract and active principle from Nunavik collections of the macroalgae Fucus evanescens C. Agardh (Fucaceae). Can. J. Microbiol. 2011, 57, 745–749. [Google Scholar] [CrossRef] [PubMed]
- Lee, J.H.; Eom, S.H.; Lee, E.H.; Jung, Y.J.; Kim, H.J.; Jo, M.R.; Son, K.T.; Lee, H.J.; Kim, J.H.; Lee, M.S. In vitro antibacterial and synergistic effect of phlorotannins isolated from edible Brown seaweed Eisenia bicyclis against acne-related bacteria. Algae 2014, 29, 47–55. [Google Scholar] [CrossRef]
- Choi, J.S.; Ha, Y.M.; Joo, C.U.; Cho, K.K.; Kim, S.J.; Choi, I.S. Inhibition of oral pathogens and collagenase activity by seaweed extracts. J. Environ. Biol. 2012, 33, 115–121. [Google Scholar]
- Park, N.H.; Choi, J.S.; Hwang, S.Y.; Kim, Y.C.; Hong, Y.K.; Cho, K.K.; Choi, I.S. Antimicrobial activities of stearidonic and gamma-linolenic acids from the green seaweed Enteromorpha linza against several oral pathogenic bacteria. Bot. Stud. 2013, 54, 39–47. [Google Scholar] [CrossRef] [Green Version]
- Kim, Y.H.; Kim, J.H.; Jin, H.J.; Lee, S.Y. Antimicrobial activity of ethanol extracts of Laminaria japonica against oral microorganisms. Anaerobe 2013, 21, 34–38. [Google Scholar] [CrossRef]
- Lee, K.Y.; Jeong, M.R.; Choi, S.M.; Na, S.S.; Cha, J.D. Synergistic effect of fucoidan with antibiotics against oral pathogenic bacteria. Arch. Oral Biol. 2013, 58, 482–492. [Google Scholar] [CrossRef]
- Gupta, S.; Rajauria, G.; Abu-Ghannam, N. Study of the microbial diversity and antimicrobial properties of Irish edible brown seaweeds. Int. J. Food Sci. Technol. 2010, 45, 482–489. [Google Scholar] [CrossRef] [Green Version]
- Dussault, D.; Vu, K.; Vansach, T.; Horgen, F.; Lacroix, M. Antimicrobial effects of marine algal extracts and cyanobacterial pure compounds against five foodborne pathogens. Food Chem. 2016, 199, 114–118. [Google Scholar] [CrossRef] [PubMed]
- Krish, S.; Das, A. In-vitro bioactivity of marine seaweed, Cladophora rupestris. Int. J. Pharma Bio Sci. 2014, 5, 898–908. [Google Scholar]
- Boisvert, C.; Beaulieu, L.; Bonnet, C.; Pelletier, E. Assessment of the antioxidant and antibacterial activities of three species of edible seaweeds. J. Food Biochem. 2015, 39, 377–387. [Google Scholar] [CrossRef]
- Rodríguez-Martínez, A.V.; Sendón, R.; Abad, M.J.; González-Rodríguez, M.V.; Barros-Velázquez, J.; Aubourg, S.P.; Paseiro-Losada, P.; Rodríguez-Bernaldo de Quirós, A. Migration kinetics of sorbic acid from polylactic acid and seaweed based films into food simulants. Food Sci. Technol. 2016, 65, 630–636. [Google Scholar] [CrossRef]
- Sanaa, M.; Shanab, M. Antioxidant and antibiotic activities of some sea weeds (Egyptian isolates). Int. J. Agric. Biol. 2007, 9, 220–225. [Google Scholar]
- Senevirathne, M.; Kim, S.; Siriwardhana, N.; Ha, J.; Lee, K.; Jown, Y. Antioxidant potential of Ecklonia cava on Reactive oxygen species scavenging, metal chelating, reducing power and lipid peroxidation inhibition. Food Sci. Technol. Int. 2006, 12, 27–38. [Google Scholar] [CrossRef]
- Siriwardhana, N.; Lee, K.; Kim, S.; Ha, J.; Jeon, Y. Antioxidant activity of Hizikia fusiformis on reactive oxygen species scavenging and lipid peroxidation inhibition. Food Sci. Technol. Int. 2003, 9, 339–347. [Google Scholar] [CrossRef]
- Devi, K.P.; Suganthy, N.; Kesika, P.; Pandian, S.K. Bioprotective properties of seaweeds: In vitro evaluation of antioxidant activity and antimicrobial activity against food borne bacteria in relation to polyphenolic content. BMC Complement. Altern. Med. 2008, 8, 38. [Google Scholar] [CrossRef] [Green Version]
- Patra, J.K.; Rath, S.K.; Jena, K.; Rathod, V.K.; Thatoi, H. Evaluation of antioxidant and antimicrobial activity of seaweed (Sargassum sp.) extract: A study on inhibition of Glutathione-S-Transferase activity. Turkey J. Biol. 2008, 32, 119–125. [Google Scholar]
- Premalatha, B. Semecarpus anacardium L. nut extract administration induces the in vivo antioxidant defense system in Aflatoxin B mediated hepatocellular carcinoma. J. Ethnopharmacol. 2011, 66. [Google Scholar]
- Mariya, V.; Ravindran, V. Biomedical and pharmacological significance of marine macro algaereview. Indian J. Geo-Marine Sci. 2013, 42, 527–537. [Google Scholar]
- Heo, S.; Ko, S.; Cha, S.; Kang, D.; Park, H.; Choi, Y.; Kim, D.; Jung, W.; Jeon, Y. Effect of phlorotannins isolated from Ecklonia cava on melanogenesis and their protective effect against photo-oxidative stress induced by UV-B radiation. Toxicology 2009, 23, 1123–1130. [Google Scholar] [CrossRef] [PubMed]
- Heo, S.J.; Ko, S.C.; Kang, S.M.; Cha, S.H.; Lee, S.H.; Kang, D.H. Inhibitory effect of diphlorethohydroxycarmalol on melanogenesis and its protective effect against UV-B radiationinduced cell damage. Food Chem. Toxicol. 2010, 48, 1355–1361. [Google Scholar] [CrossRef] [PubMed]
- Qi, H.; Zhang, Q.; Zhao, T.; Chen, R.; Zhang, H.; Niu, X.; Li, Z. Antioxidant activity of different sulfate content derivatives of polysaccharide extracted from Ulva pertusa (Chlorophyta) in vitro. Int. J. Biol. Macromol. 2005, 37, 195–199. [Google Scholar] [CrossRef] [PubMed]
- Xue, C.; Chen, L.; Li, Z.; Cai, Y.; Lin, H.; Fang, Y. Antioxidative activities of low molecular fucoidans from kelp Laminaria japonica. Dev. Food Sci. 2004, 42, 139–145. [Google Scholar]
- Kumar, K.; Ganesane, K.; Rao, P. Antioxidant potential of solvent extracts of Kappaphycus alvarezii (Doty) Doty: an edible seaweed. Food Chem. 2008, 107, 289–295. [Google Scholar] [CrossRef]
- Li, K.; Li, X.; Ji, N.; Wang, B. Natural bromophenols from the marine red alga Polysiphonia urceolata (Rhodomelaceae): structural elucidation and DPPH radicalscavenging activity. Bioorganic Med. Chem. 2007, 15, 6627–6631. [Google Scholar] [CrossRef]
- Delgado, N.G.; Vázquez, A.I.F.; Sánchez, H.C.; del Valle, R.M.S.; Gómez, Y.S.; Alfonso, A.M.S. Antiinflammatory and antinociceptive activities of methanolic extract from red seaweed Dichotomaria obtusata. Brazilian J. Pharm. Sci. 2013, 1, 65–74. [Google Scholar] [CrossRef] [Green Version]
- Ferrucci, L.; Cherubini, A.; Bandinelli, S.; Bartali, B.; Corsi, A.; Lauretani, F.; Martin, A.; Andres-Lacueva, C.; Senin, U.; Guralnik, J.M.R. Relationship of plasma polyunsaturated fatty acids to circulating inflammatory markers. J. Clin. Endocrinol. Metab. 2006, 91, 439–446. [Google Scholar] [CrossRef]
- Khan, N.A.; Choi, J.S.; Lee, M.C.; Kim, E.; Nam, T.J.; Fujii, H.; Hong, Y.K. Antiinflammatory activities of methanol extracts from various seaweed species. J. Environ. Biol. 2008, 29, 465–469. [Google Scholar] [PubMed]
- Heo, S.J.; Yoon, W.J.; Kim, K.N.; Ahn, G.N.; Kang, S.M.; Kang, D.H. Evaluation of anti-inflammatory effect of fucoxanthin isolated from brown algae in lipopolysaccharide stimulated RAW 264.7 macrophages. Food Chem. Toxicol. 2010, 48, 2045–2051. [Google Scholar] [CrossRef] [PubMed]
- Enoki, T.; Sagawa, H.; Tominaga, T.; Nishiyama, E.; Komyama, N.; Sakai, T.; Yu, F.; Ikai, K.; Kato, I. Drugs, Foods or Drinks with the Use of Algae-Derived Physiologically Active Substances. U.S. Patent 0,105,029, 2003. [Google Scholar]
- Kim, K.J.; Lee, B.Y. Fucoidan from the sporophyll of Undaria pinnatifida suppresses adipocyte differentiation by inhibition of inflammation related cytokines in 3T3-L1 cells. Nutr. Res. 2012, 32, 439–447. [Google Scholar] [CrossRef]
- Park, H.; Han, M.; Park, C.; Jin, C.; Kim, G.; Choi, I.; Kim, N.; Nam, T.; Kwon, T.; Choi, Y. Anti-inflammatory effects of fucoidan through inhibition of NF-κB, MAPK and Akt activation in lipopolysaccharide-induced BV2 microglia cells. Food Chem. Toxicol. 2011, 49, 1745–1752. [Google Scholar] [CrossRef] [PubMed]
- Hosokawa, M.; Miyashita, T.; Nishikawa, S.; Tsukui, T.; Beppu, F.; Okada, T.; Miyashita, K. Fucoxanthin regulates adipocytokine mRNA expression in white adipose tissue of diabetic/obese KK-Ay mice. Arch. Biochem. Biophys. 2010, 504, 17–25. [Google Scholar] [CrossRef]
- Lee, S.H.; Han, J.S.; Heo, S.J.; Hwang, J.Y.; Jeon, Y.J. Protective effects of dieckol isolated from Ecklonia cava against high glucose-induced oxidative stress in human umbilical vein endothelial cells. Toxicology 2010, 24, 375–381. [Google Scholar] [CrossRef]
- Kim, M.M.; Rajapakse, N.; Kim, S.K. Anti-inflammatory effect of Ishige okamurae ethanolic extract via inhibition of NF-kappaB transcription factor in RAW 264.7 cells. Phyther. Res. 2009, 23, 628–634. [Google Scholar] [CrossRef]
- Lin, H.T.V.; Lu, W.J.; Tsai, G.; Chou, C.; Hsiao, H.; Hwang, P. Enhanced anti-inflammatory activity of brown seaweed Laminaria japonica by fermentation using Bacillus subtilis. Process Biochem. 2016, 51, 1945–1953. [Google Scholar] [CrossRef]
- Ramberg, J.E.; Nelson, E.D.; Sinnott, R.A. Immunomodulatory dietary polysaccharides: a systematic review of the literature. Nutr. J. 2010, 9. [Google Scholar] [CrossRef] [Green Version]
- Ye, H.; Wang, K.; Zhou, C.; Liu, J.; Zeng, X. Purification, antitumor and antioxidant activities in vitro of polysaccharides from the brown seaweed Sargassum pallidum. Food Chem. 2008, 111, 428–432. [Google Scholar] [CrossRef]
- Namvar, F.; Mohamad, R.; Baharara, J.; Zafar-Balanejad, S.; Fargahi, F.; Rahman, H.S. Antioxidant, Antiproliferative, and Antiangiogenesis Effects of Polyphenol-Rich Seaweed (Sargassum muticum). Biomed Res. Int. 2013, 2013, 1–9. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Castro, L.S.E.P.W.; de Sousa Pinheiro, T.; Castro, A.J.G.; Santos, M.D.S.N.; Soriano, E.M.; Leite, E.L. Potential anti-angiogenic, antiproliferative, antioxidant, and anticoagulant activity of anionic polysaccharides, fucans, extracted from brown algae Lobophora variegate. J. Appl. Phycol. 2015, 27, 1315. [Google Scholar] [CrossRef]
- Namvar, F.; Mohamed, S.; Fard, S.G.; Behravan, J.; Mustapha, N.M.; Alitheen, N.B.M.; Othman, F. Polyphenol-rich seaweed (Eucheuma cottonii) extract suppresses breast tumour via hormone modulation and apoptosis induction. Food Chem. 2012, 130, 376–382. [Google Scholar] [CrossRef]
- Yang, C.; Chung, D.; Shin, I.S.; Lee, H.; Kim, J.; Lee, Y.; You, S.G. Effects of Molecular Weight and Hydrolysis Conditions on Anticancer Activity of Fucoidans from Sporophyll of Undaria pinnatifida. Int. J. Biol. Macromol. 2008, 43, 433–437. [Google Scholar] [CrossRef]
- Michel, G.; Nyval-Collen, P.; Barbeyron, T.; Czjzek, M.; Helbert, W. Bioconversion of red seaweed galactans: a focus on bacterial agarases and carrageenases. Appl. Microbiol. Biotechnol. 2006, 71, 23–33. [Google Scholar] [CrossRef]
- Athukorala, Y.; Lee, K.-W.; Kim, S.-K.; Jeon, Y.-J. Anticoagulant activity of marine green and brown algae collected from Jeju Island in Korea. Bioresour. Technol. 2007, 98, 1711–1716. [Google Scholar] [CrossRef]
- De Zoysa, M.; Nikapitiya, C.; Jeon, Y.-J.; Jee, Y.; Lee, J. Anticoagulant activity of sulfated polysaccharide isolated from fermented brown seaweed Sargassum fulvellum. J. Appl. Phycol. 2008, 20, 67–74. [Google Scholar] [CrossRef]
- Júnior, S.Q.; Carneiro, V.H.A.; Fontenelle, T.P.C.; de Sousa Chaves, L.; Mesquita, J.X.; de Brito, T.V.; Prudêncio, R.S.; de Oliveira, J.S.; Medeiros, J.-V.R.; Aragão, K.S.; et al. Antioxidant and anti-inflammatory activities of methanol extract and its fractions from the brown seaweed Spatoglossum schroederi. J. Appl. Phycol. 2015, 27, 2367–2376. [Google Scholar] [CrossRef]
- Radhika, D.; Veerabahu, C.; Priya, R. Anti-inflammatory activities of some seaweed collected from the Gulf Of Mannar Coast, Tuticorin, South India. Int. J. Pharma Bio Sci. 2013, 4, 39–44. [Google Scholar]
- Albuquerque, I.R.L.; Cordeiro, S.L.; Gomes, D.L.; Dreyfuss, J.L.; Filgueira, L.G.A.; Leite, E.L.; Nader, H.B.; Rocha, H.A.O. Evaluation of anti-nociceptive and antiinflammatory activities of a heterofucan from Dictyota menstrualis. Mar. Drugs 2013, 11, 2722–2740. [Google Scholar] [CrossRef] [Green Version]
- Wang, R.; Paul, V.J.; Luesch, H. Seaweed extracts and unsaturated fatty acid constituents from the green alga Ulva lactuca as activators of the cytoprotective Nrf2-ARE pathway. Free Radic. Biol. Med. 2013, 57, 141–153. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kang, M.; Wijesinghe, W.; Lee, S.; Kang, S.; Ko, S.; Yang, X.; Kang, N.; Jeon, B.; Kim, J.; Lee, D.; et al. Dieckol isolated from brown seaweed Ecklonia cava attenuates type II diabetes in db/db mouse model. Food Chem. Toxicol. 2013, 53, 294–298. [Google Scholar] [CrossRef] [PubMed]
- Lee, S.; Lee, Y.S.; Jung, S.H.; Kang, S.S.; Shin, K.H. Anti-oxidant activities of fucosterol from the marine algae Pelvetia siliquosa. Arch. Pharm. Res. 2003, 26, 719–722. [Google Scholar] [CrossRef] [PubMed]
- Rivero, F.; Fallarero, A.; Castañeda, O.; Dajas, F.; Manta, E.; Areces, F.; Mancini Filho, J.; Vidal, A. Antioxidant activity in vivo and in vitro of Halimeda incrassata aqueous extracts. Food Sci. Technol. 2003, 23, 256–263. [Google Scholar] [CrossRef] [Green Version]
- Wu, X.; Yang, M.; Huang, X.; Yan, J.; Luo, Q. Effect of fucoidan on splenic lymphocyte apoptosis induced by radiation. Chinese J. Radiol. Med. Prot. 2003, 23, 43–50. [Google Scholar]
- Luo, D.; Zhang, Q.; Wang, H.; Cui, Y.; Sun, Z.; Yang, J.; Zheng, Y.; Jia, J.; Yu, F.; Wang, X.; et al. Fucoidan protects against dopaminergic neuron death in vivo and in vitro. Eur. J. Pharmacol. 2009, 617, 33–40. [Google Scholar] [CrossRef]
- Burney, M.; Mathew, L.; Gaikwad, A.; Nugent, E.K.; Gonzalez, A.O.; Smith, J.A. Evaluation Fucoidan Extracts From Undaria pinnatifida and Fucus vesiculosus in Combination with Anticancer Drugs in Human Cancer Orthotopic Mouse Models. Integr. Cancer Ther. 2018, 17, 755–761. [Google Scholar] [CrossRef] [Green Version]
- Mathew, L.; Burney, M.; Gaikwad, A.; Nyshadham, P.; Nugent, E.K.; Gonzalez, A.O.; Smith, J.A. Preclinical Evaluation of Safety of Fucoidan Extracts From Undaria pinnatifida and Fucus vesiculosus for Use in Cancer Treatment. Integr. Cancer Ther. 2017, 16, 572–584. [Google Scholar] [CrossRef] [Green Version]
- Zhou, G.; Sun, Y.; Xin, H.; Zhang, Y.; Li, Z.; Xu, Z. In vivo antitumor and immunomodulation activities of different molecular weight lambda-carrageenans from Chondrus ocellatus. Pharmacol. Res. 2004, 50, 47–53. [Google Scholar] [CrossRef]
- Syad, A.N.; Rajamohamed, B.S.; Shunmugaiah, K.P.; Kasi, P.D. Neuroprotective effect of the marine macroalga Gelidiellaacerosa: identification of active compounds through bioactivity-guided fractionation. Pharm. Biol. 2016, 54, 2073–2081. [Google Scholar] [CrossRef] [Green Version]
- Xu, H.; Kitajima, C.; Ito, H.; Miyazaki, T.; Baba, M.; Okuyama, T.; Okada, Y. Antidiabetic effect of polyphenols from brown alga Ecklonia kurome in genetically diabetic KK-A(y) mice. Pharm. Biol. 2012, 50, 393–400. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Liu, M.; Zhang, W.; Qiu, L.; Lin, X.K. Synthesis of butyl-isobutyl-phthalate and its interaction with α-glucosidase in vitro. J. Biochem. 2011, 149, 27–33. [Google Scholar] [CrossRef] [PubMed]
- Pantidos, N.; Boath, A.; Lund, V.; Conner, S.; McDougall, G.J. Phenolic-rich extracts from the edible seaweed, ascophyllum nodosum, inhibit α-amylase and α-glucosidase: Potential anti-hyperglycemic effects. J. Funct. Foods 2014, 10, 201–209. [Google Scholar] [CrossRef]
- Lee, S.; Park, M.; Heo, S.; Kang, S.; Ko, S.; Han, J.; Jeon, Y. Dieckol isolated from Ecklonia cava inhibits α-glucosidase and α-amylase in vitro and alleviates postprandial hyperglycemia in streptozotocin-induced diabetic mice. Food Chem. Toxicol. 2010, 48. [Google Scholar] [CrossRef]
- Park, M.H.; Nam, Y.H.; Han, J.S. Sargassum coreanum extract alleviates hyperglycemia and improves insulin resistance in db/db diabetic mice. Nutr. Res. Pract. 2015, 9, 472–479. [Google Scholar] [CrossRef] [Green Version]
- Maeda, H.; Hosokawa, M.; Sashima, T.; Murakami-Funayama, K.; Miyashita, K. Anti-obesity and anti-diabetic effects of fucoxanthin on diet-induced obesity conditions in a murine model. Mol. Med. Rep. 2009, 2, 897–902. [Google Scholar] [CrossRef]
- Barros-Gomes, J.; Nascimento, D.; Silveira, A.; Silva, R.; Gomes, D.; Melo, K.; Almeida-Lima, J.; Camara, R.; Silva, N.; Rocha, H. In Vivo Evaluation of the Antioxidant Activity and Protective Action of the Seaweed Gracilaria birdiae. Oxid. Med. Cell. Longev. 2018, 9354296, 1–12. [Google Scholar] [CrossRef] [Green Version]
- Kim, M.-J.; Chang, U.-J.; Lee, J.-S. Inhibitory effects of fucoidan in 3t3-l1 adipocyte differentiation. Mar. Biotechnol. 2009, 11, 557–562. [Google Scholar] [CrossRef]
- Hassan, S.; El-Twab, S.A.; Hetta, M.; Mahmoud, B. Improvement of lipid profile and antioxidant of hypercholesterolemic albino rats by polysaccharides extracted from the green alga Ulva lactuca Linnaeus. Saudi J. Biol. Sci. 2011, 18, 333–340. [Google Scholar] [CrossRef] [Green Version]
- Park, M.K.; Jung, U.; Roh, C. Fucoidan from marine brown algae inhibits lipid accumulation. Mar. Drugs 2011, 1359–1367. [Google Scholar] [CrossRef] [Green Version]
- Ha, A.W.; Kim, W.K. The effect of fucoxanthin rich power on the lipid metabolism in rats with a high fat diet. Nutr. Res. Pract. 2013, 7, 287–293. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kim, J.-Y.; Kwon, Y.M.; Kim, I.-S.; Kim, J.-A.; Yu, D.-Y.; Adhikari, B.; Lee, S.-S.; Choi, I.-S.; Cho, K.-K. Effects of the brown seaweed Laminaria japonica supplementation on serum concentrations of IgG, triglycerides, and cholesterol, and intestinal microbiota composition in rats. Front. Nutr. 2018, 5. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wang, L.; Wang, X.; Wu, H.; Liu, R. Overview on biological activities and molecular characteristics of sulfated polysaccharides from marine green algae in recent years. Mar. Drugs 2014, 12, 4984–5020. [Google Scholar] [CrossRef] [PubMed]
- Hayakawa, Y.; Hayashi, T.; Hayashi, K.; Osawa, T.; Niiya, K.; Sakuragawa, N. Activation of heparin cofactor II by calcium spirulan. J. Biol. Chem. 2000, 275, 11379–11382. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rocha, H.A.O.; Moraes, F.A.; Trindade, E.S.; Franco, C.R.C.; Torquato, R.J.S.; Veiga, S.S.; Valente, A.P.; Mourão, P.A.S.; Leite, E.L.; Nader, H.B.; et al. Structural and haemostatic activities of a sulfated galactofucan from the brown alga Spatoglossum schröederi. An ideal antithrombotic agent? J. Biol. Chem. 2005, 280, 41278–41288. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Liu, X.; Wang, S.; Cao, S.; He, X.; Qin, L.; He, M.; Yang, Y.; Hao, J.; Mao, W. Structural Characteristics and Anticoagulant Property In Vitro and In Vivo of a Seaweed Sulfated Rhamnan. Mar. Drugs 2018, 16, 243. [Google Scholar] [CrossRef] [Green Version]
- Maruyamaa, H.; Tamauchib, H.; Hashimotoc, M.; Nakano, T. Suppression of Th2 immune responses by Mekabu fucoidan from Undaria pinnatifida sporophylls. Int. Arch. Allergy Immunol. 2005, 137, 289–294. [Google Scholar] [CrossRef] [PubMed]
- Senni, K.; Gueniche, F.; Foucault-Bertaud, A.; Igondjo-Tchen, S.; Fioretti, F.; Colliec-Jouault, S.; Durand, P.; Guezennec, J.; Godeau, G.; Letourneur, D. Fucoidan a sulfated polysaccharide from brown algae is a potent modulator of connective tissue proteolysis. Arch. Biochem. Biophys. 2006, 445, 56–64. [Google Scholar] [CrossRef] [Green Version]
- Liu, X.; Shao, C.; Kong, W.; Fang, Y.; Wang, C. Evaluation of antitumor, immunomodulatory and free radical scavenging effects of a new herbal prescription seaweed complex preparation. J. Ocean Univ. China 2013, 12, 515–520. [Google Scholar] [CrossRef]
- Duraikannu, K.; Rani, K.S.; Anithajothi, R.; Umagowsalya, G.; Ramakritinan, C.M. In-vivo anticancer activity of red algae (Gelidiela acerosa and Acanthophora spicifera). Int. J. Pharm. Sci. Res. 2014, 5, 3347–3352. [Google Scholar]
- Zhao, C.; Yang, C.; Liu, B.; Lin, L.; Sarker, S.D.; Nahar, L.; Yu, H.; Cao, H.; Xiao, J. Bioactive compounds from marine macroalgae and their hypoglycemic benefits. Trends Food Sci. Technol. 2018, 72, 1–12. [Google Scholar] [CrossRef]
- Oh, J.-H.; Kim, J.; Lee, Y. Anti-inflammatory and antidiabetic effects of brown seaweeds in high-fat diet-induced obese mice. Nutr. Res. Pract. 2016, 10, 42–48. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Smit, A.J. Review Medicinal and pharmaceutical uses of seaweed natural products: A review. J. Appl. Phycol. 2004, 16. [Google Scholar] [CrossRef]
- Kim, M.; Kim, J.; Choi, W.; Lee, S. Effects of seaweed supplementation on blood glucose concentration, lipid profile, and antioxidant enzyme activities in patients with type 2 diabetes mellitus. Nutr. Res. Pract. 2008, 2, 62–67. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Takahashi, H.; Kawaguchi, M.; Kitamura, K.; Narumiya, S.; Kawamura, M.; Tengan, I.; Nishimoto, S.; Hanamure, Y.; Majima, Y.; Tsubura, S.; et al. An Exploratory Study on the anti-inflammatory Effects of Fucoidan in relation to Quality of Life in Advanced Cancer Patients. Integr. Cancer Ther. 2018, 17, 282291. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Teas, J.; Vena, S.; Cone, D.L.; Irhimeh, M. The consumption of seaweed as a protective factor in the etiology of breast cancer: proof of principle. J. Appl. Phycol. 2013, 25, 771–779. [Google Scholar] [CrossRef] [Green Version]
- Lee, L.; Wall, S.; Klepach, D.; Ge, L.; Zhang, Z.; Lee, R. Algisyl-LVRTM with coronary artery bypass grafting reduces left ventricular wall stress and improves function in the failing human heart. Int. J. Cardiol. 2013, 168, 2022–2028. [Google Scholar] [CrossRef] [Green Version]
- Anker, S.; Coats, A.; Cristian, G.; Dragomir, D.; Pusineri, E.; Piredda, M. A prospective comparison of alginate-hydrogel with standard medical therapy to determine impact on functional capacity and clinical outcomes in patients with advanced heart failure (AUGMENT-HF trial). Eur. Heart J. 2015, 36, 2297–2309. [Google Scholar] [CrossRef] [Green Version]
- Shanmugam, A.; Sudharsan, S.; Seedevi, P.; Ramasamy, P.; Subhapradha, N.; Vairamani, S. Heavy metal accumulation in seaweeds and sea grasses along southeast coast of India. J. Chem. Pharm. Res. 2012, 4, 4240–4244. [Google Scholar]
- Fantonalgo, R.N.; Falguisana, M.N. Detrimental Effects of the Selected Heavy Metals to Seaweeds: A Review. J. Environ. Earth Sci. 2017, 7, 30–39. [Google Scholar]
- Duinker, A.; Roiha, I.; Amlund, H.; Dahl, L.; Lock, E.-J.; Kögel, T.; Lunestad, B. Potential risks posed by macroalgae for application as feed and food – a Norwegian perspective. Natl. Inst. Nutr. Seaf. Res. 2016, 1–24. [Google Scholar]
- Pycke, B.; Faasse, M. Biochemical Composition and Quality Assessment of Native Macroalgae Collected along the Flemish Coast; Public Output report of the EnAlgae project: Oostende, Belgium, December 2015. [Google Scholar]
- Domínguez-González, R.; Romarís-Hortas, V.; García-Sartal, C.; Moreda-Piñeiro, A.; Barciela-Alonso, M.; Bermejo-Barrera, P. Evaluation of an in vitro method to estimate trace elements bioavailability in edible seaweeds. Talanta 2010, 82, 1668–1673. [Google Scholar] [CrossRef] [PubMed]
- Fleurence, J. Seaweeds as food. In Seaweed in Health and Disease Prevention; Fleurence, J., Levine, I., Eds.; Elsevier Inc.: Amsterdam, The Netherlandsc, 2016; pp. 149–167. [Google Scholar]
- Benevides, N.; Silva, S.D.S.; Magalhaes, S.; Melo, F.; Freitas, A.; Vasconcelos, I. Proximate analysis, toxic and antinutritional factors of ten Brazilian marine algae. Brazilian J. Veg. Physiol. 1998, 10, 31–36. [Google Scholar]
- Stévant, P.; Rebours, C.; Chapman, A. Seaweed aquaculture in Norway: recent industrial developments and future perspectives. Aquac. Int. 2017, 25, 1373–1390. [Google Scholar] [CrossRef] [Green Version]
- Oliveira, M.; Freitas, A.; Carvalho, A.; Sampaio, T.; Farias, D.; Teixeira, D.; Gouveia, S.; Pereira, J.; Sena, M. Nutritive and non-nutritive attributes of washed-up seaweeds from the coast of Ceará, Brazil. Food Chem. 2009, 115, 254–259. [Google Scholar] [CrossRef]
- Vasconcelos, I.; Oliveira, J. Antinutritional properties of plant lectins. Toxicon 2004, 44. [Google Scholar] [CrossRef] [PubMed]
- Rehman, Z.; Shah, W. Thermal heat processing effects on antinutrients, protein and starch digestibility of food legumes. Food Chem. 2004, 91. [Google Scholar] [CrossRef]
- Fleurence, J. The enzymatic degradation of algal cell walls: a useful approach to improving protein accessibility? J. Appl. Phycol. 1999, 11, 313–314. [Google Scholar] [CrossRef]
- Michalak, I.; Chojnacka, K. Algal extracts: Technology and advances. Eng. Life Sci. 2014, 14, 581–591. [Google Scholar] [CrossRef]
- Lahaye, M.; Vigouroux, J. Liquefaction of Dulse (Palmaria palmata (L.) Kuntze) by a commercial enzyme preparation and a purified endo-b-1,4 D-xylanase. J. Appl. Phycol. 1992, 4, 329–337. [Google Scholar] [CrossRef]
- Ardiansyah, D.; Hartinah, I. Wahidah Improvement of the nutritive quality of Sargassum powder through Aspergillus niger, Saccharomyces cerevisiae, and Lactobacillus spp. fermentations. AACL Bioflux 2018, 11, 753–764. [Google Scholar]
- Plaza, M.; Santoyo, S.; Jaime, L.; García-Blairsy Reina, G.; Herrero, M.; Señoráns, F.; Ibáñez, E. Screening for bioactive compounds from algae. J. Pharm. Biomed. Anal. 2010, 51, 450–455. [Google Scholar] [CrossRef] [PubMed]
- Maehre, H.; Edvinsen, G.; Eilertsen, K.; Elvevoll, E. Heat treatment increases the protein bioaccessibility in the red seaweed dulse (Palmaria palmata), but not in the brown seaweed winged kelp (Alaria esculenta). J. Appl. Phycol. 2016, 28, 581–590. [Google Scholar] [CrossRef] [Green Version]
- Santoso, J.; Gunji, S.; Yoshie-Stark, Y.; Suzuki, T. Mineral contents of Indonesian seaweeds and mineral solubility affected by basic cooking. Food Sci. Technol. Res. 2006, 12, 59–66. [Google Scholar] [CrossRef] [Green Version]
- Agatonovic-Kustrin, S.; Morton, D. Cosmeceuticals derived from bioactive substances found in marine algae. Oceanography 2013, 1. [Google Scholar]
- Jimenez-Escrig, A.; Sanchez-Muniz, F. Dietary fibre from edible seaweeds: chemical structure, physicochemical properties and effects on cholesterol metabolism. Nutr. Res. 2000, 20, 585–598. [Google Scholar] [CrossRef]
- García-Casal, M.; Pereira, A.; Leets, I.; Ramírez, J.; Quiroga, M. High iron content and bioavailability in humans from four species of marine algae. J. Nutr. 2007, 137, 2691–2695. [Google Scholar] [CrossRef]
- Burtin, P. Nutritional value of seaweeds. Electron. J. Environ. Agric. Food Chem. 2003, 2, 6. [Google Scholar]
- Uchida, M.; Murata, M. Fermentative preparation of single cell detritus from seaweed, Undaria pinnatifida, suitable as a replacement hatchery diet for unicellular algae. Aquaculture 2002, 207. [Google Scholar] [CrossRef]
- Fernando, I.P.S.; Nah, J.W.; Jeon, Y.J. Potential anti-inflammatory natural products from marine algae. Environ. Toxicol. Pharmacol. 2016, 48, 22–30. [Google Scholar] [CrossRef]
- European Commission CORDIS Database: EU Research Results. Available online: https://cordis.europa.eu/ (accessed on 31 January 2019).
- European Parliament and of the council of the European Union Cosmetic products should be safe under normal or reasonably foreseeable conditions of use. In particular, a risk- benefit reasoning should not justify a risk to human health. Off. J. Eur. Union 2009, 241–247.
- Hazozaki, T.; Laughlin, L.T.; Swanson, C.L. Laminaria Saccharina Extract and Vitamin b3 as Whitening Agents. WIPO (PCT) Patent WO2012011907A1, 26 January 2012. [Google Scholar]
- Ferdouse, F.; Holdt, S.L.; Smith, R.; Murúa, P.; Yang, Z. The global status of seaweed production, trade and utilization. FAO Globefish Res. Program. 2018, 124, 120. [Google Scholar]
- Ma’or, Z.; Yehuda, S.; Voss, W. Skin smoothing effects of Dead Sea minerals: Comparative profilometric evaluation of skin surface. Int. J. Cosmet. Sci. 1997, 19, 105–110. [Google Scholar] [CrossRef] [PubMed]
- Rossano, R.; Ungaro, N.; D’Ambrosio, A.; Liuzzi, G.M.; Riccio, P. Extracting and purifying R-phycoerythrin from Mediterranean red algae Corallina elongata Ellis & Solander. J. Biotechnol. 2003, 101, 289–293. [Google Scholar] [PubMed]
- Swatschek, D.; Schatton, W.; Kellermann, J.; Müller, W.E.G.; Kreuter, J. Marine sponge collagen: Isolation, characterization and effects on the skin parameters surface-pH, moisture and sebum. Eur. J. Pharm. Biopharm. 2002, 53, 107–113. [Google Scholar] [CrossRef]
- Hartmann, A.; Murauer, A.; Ganzera, M. Quantitative analysis of mycosporine-like amino acids in marine algae by capillary electrophoresis with diode-array detection. J. Pharm. Biomed. Anal. 2017, 138, 153–157. [Google Scholar] [CrossRef]
- Colabella, F.; Moline, M.; Libkind, D. UV Sunscreens of Microbial Origin: Mycosporines and Mycosporine- like Aminoacids. Recent Pat. Biotechnol. 2015, 8, 179–193. [Google Scholar] [CrossRef]
- Vanderlei, E.S.O.; Patoilo, K.K.N.R.; Lima, N.A.; Lima, A.P.S.; Rodrigues, J.A.G.; Silva, L.M.C.M.; Lima, M.E.P.; Lima, V.; Benevides, N.M.B. Antinociceptive and anti-inflammatory activities of lectin from the marine green alga Caulerpa cupressoides. Int. Immunopharmacol. 2010, 10, 1113–1118. [Google Scholar] [CrossRef]
- Sheih, I.C.; Fang, T.J.; Wu, T.K.; Lin, P.H. Anticancer and antioxidant activities of the peptide fraction from algae protein waste. J. Agric. Food Chem. 2010, 58, 1202–1207. [Google Scholar] [CrossRef]
- Wiencke, C.; torn Dieck, I. Temperature requirements of growth and temperature tolerance of macroalgae endemic to the Antarctic Region. Mar. Ecol. Prog. Ser. 2007, 54, 189–197. [Google Scholar] [CrossRef]
- Pulz, O.; Gross, W. Valuable products from biotechnology of microalgae. Appl. Microbiol. Biotechnol. 2004, 65, 635–648. [Google Scholar] [CrossRef] [PubMed]
- Urquiaga, I.; Leighton, F. Plant Polyphenol Antioxidants and Oxidative Stress. Biol. Res. 2000, 33, 55–64. [Google Scholar] [CrossRef] [PubMed]
- Singh, I.P.; Sidana, J.; Bharate, S.B.; Foley, W.J. Phloroglucinol compounds of natural origin: Synthetic aspects. Nat. Prod. Rep. 2010, 27, 393–416. [Google Scholar] [CrossRef] [PubMed]
- Stengel, D.B.; Connan, S.; Popper, Z.A. Algal chemodiversity and bioactivity: Sources of natural variability and implications for commercial application. Biotechnol. Adv. 2011, 29, 483–501. [Google Scholar] [CrossRef] [PubMed]
- Moon, H.E.; Islam, M.N.; Ahn, B.R.; Chowdhury, S.S.; Sohn, H.S.; Jung, H.A.; Choi, J.S. Protein Tyrosine Phosphatase 1B and α-Glucosidase Inhibitory Phlorotannins from Edible Brown Algae, Ecklonia stolonifera and Eisenia bicyclis. Biosci. Biotechnol. Biochem. 2011, 75, 1472–1480. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Fitton, J.H.; Irhimeh, M.; Falk, N. Macroalgal fucoidan extracts - a new opportunity for cosmetics.pdf. Cosmet. Toilet. 2007, 122, 55–64. [Google Scholar]
- Arjunan, N.; Kumari, H.L.J.; Singaravelu, C.M.; Kandasamy, R.; Kandasamy, J. Physicochemical investigations of biogenic chitosan-silver nanocomposite as antimicrobial and anticancer agent. Int. J. Biol. Macromol. 2016, 92, 77–87. [Google Scholar] [CrossRef]
- Aranaz, I.; Acosta, N.; Civera, C.; Elorza, B.; Mingo, J.; Castro, C.; Gandía, M.; de los, L.; Caballero, A.H. Cosmetics and cosmeceutical applications of chitin, chitosan and their derivatives. Polymers (Basel). 2018, 10, 213. [Google Scholar] [CrossRef] [Green Version]
- Lee, J.H.; Kim, H.H.; Ko, J.Y.; Jang, J.H.; Kim, G.H.; Lee, J.S.; Nah, J.W.; Jeon, Y.J. Rapid preparation of functional polysaccharides from Pyropia yezoensis by microwave-assistant rapid enzyme digest system. Carbohydr. Polym. 2016, 153, 512–517. [Google Scholar] [CrossRef]
- Palombo, P.; Fabrizi, G.; Ruocco, V.; Ruocco, E.; Fluhr, J.; Roberts, R.; Morganti, P. Beneficial long-term effects of combined oral/topical antioxidant treatment with the carotenoids lutein and zeaxanthin on human skin: A double-blind, placebo-controlled study. Skin Pharmacol. Physiol. 2007, 20, 199–210. [Google Scholar] [CrossRef]
- Kim, M.S.; Oh, G.H.; Kim, M.J.; Hwang, J.K. Fucosterol inhibits matrix metalloproteinase expression and promotes type-1 procollagen production in UVB-induced HaCaT cells. Photochem. Photobiol. 2013, 89, 911–918. [Google Scholar] [CrossRef]
- Peng, J.; Yuan, J.P.; Wu, C.F.; Wang, J.H. Fucoxanthin, a marine carotenoid present in brown seaweeds and diatoms: Metabolism and bioactivities relevant to human health. Mar. Drugs 2011, 9, 1806–1828. [Google Scholar] [CrossRef]
- Jensen, A. Tocopherol content of seaweed and seaweed meal: I.—Analytical methods and distribution of tocopherols in benthic algae. J. Sci. Food Agric. 1969, 20, 449–453. [Google Scholar] [CrossRef]
- Podhaisky, H.-P.; Wohlrab, W. Is the photoprotective effect of vitamin E based on its antioxidative capacity? J. Dermatol. Sci. 2002, 28, 84–86. [Google Scholar] [CrossRef]
- Alberts, D.S.; Goldman, R.; Xu, M.J.; Dorr, R.T.; Quinn, J.; Welch, K.; Guillen-Rodriguez, J.; Aickin, M.; Peng, Y.M.; Loescher, L.; et al. Disposition and metabolism of topically administered α-tocopherol acetate: A common ingredient of commercially available sunscreens and cosmetics. Nutr. Cancer 1996, 26, 193–201. [Google Scholar] [CrossRef]
- Correa, H.; Aristizabal, F.; Duque, C.; Kerr, R. Cytotoxic and antimicrobial activity of pseudopterosins and seco-pseudopterosins isolated from the octocoral Pseudopterogorgia elisabethae of San Andrés and Providencia islands (southwest Caribbean Sea). Mar. Drugs 2011, 9, 334–344. [Google Scholar] [CrossRef] [Green Version]
- Baldwin, J.E.; Williams, R.M. Biodiversity and Natural Product Diversity; Pergamon, Elsevier: Oxford, MS, USA, 2002; Volume 21. [Google Scholar]
- Pallela, R.; Na-Young, Y.; Kim, S.K. Anti-photoaging and photoprotective compounds derived from marine organisms. Mar. Drugs 2010, 8, 1189–1202. [Google Scholar] [CrossRef] [Green Version]
- Rampelotto, P.H.; Trincone, A. Grand Challenges in Marine Biotechnology. In Grand Challenges in Biology and Biotechnology; Springer International Publishing: Cham, Switzerland, 2018; p. 616. [Google Scholar]
- Dhargalkar, V.K.; Verlecar, X.N. Southern Ocean seaweeds: A resource for exploration in food and drugs. Aquaculture 2009, 287, 229–242. [Google Scholar] [CrossRef]








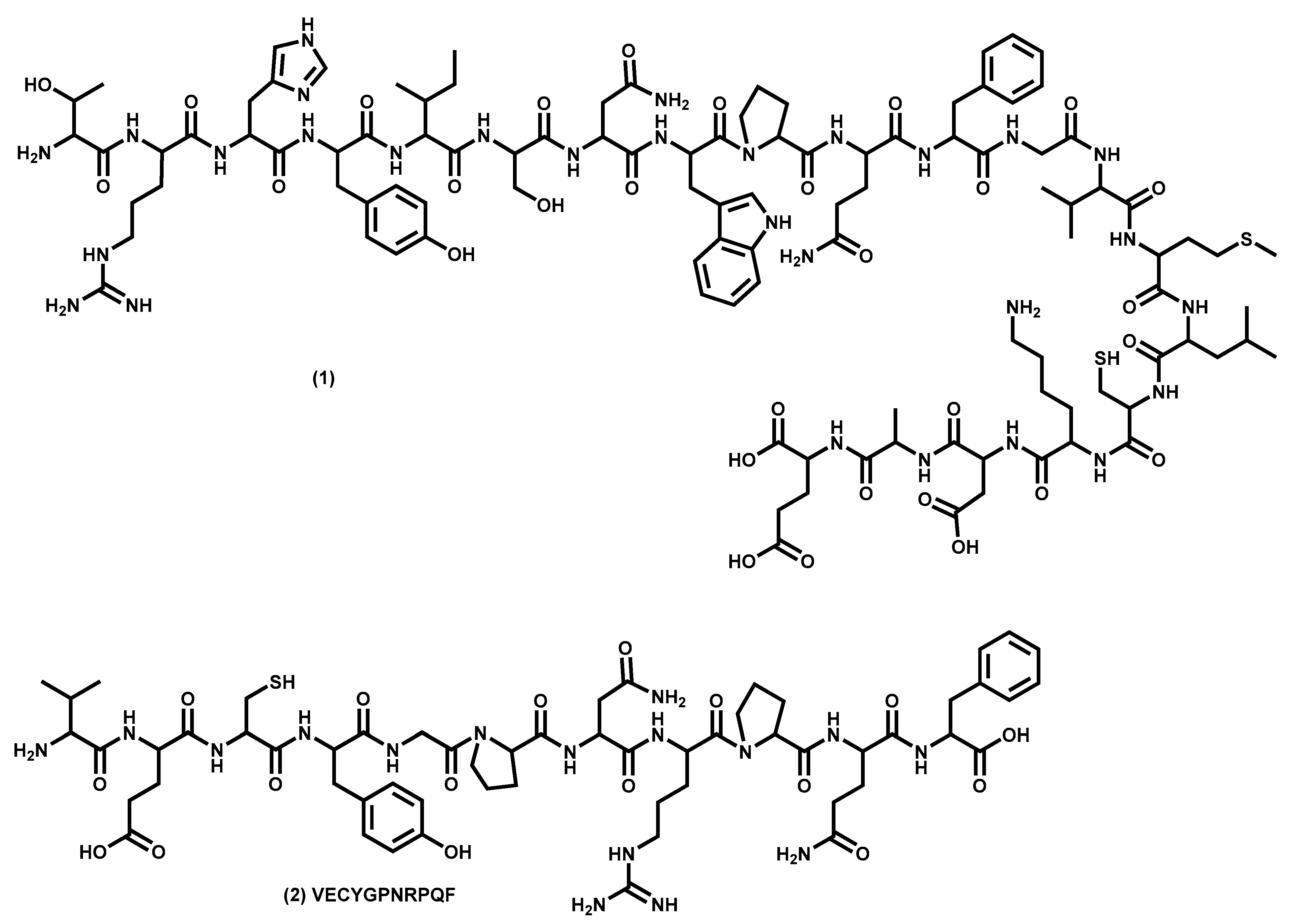{kind=link}
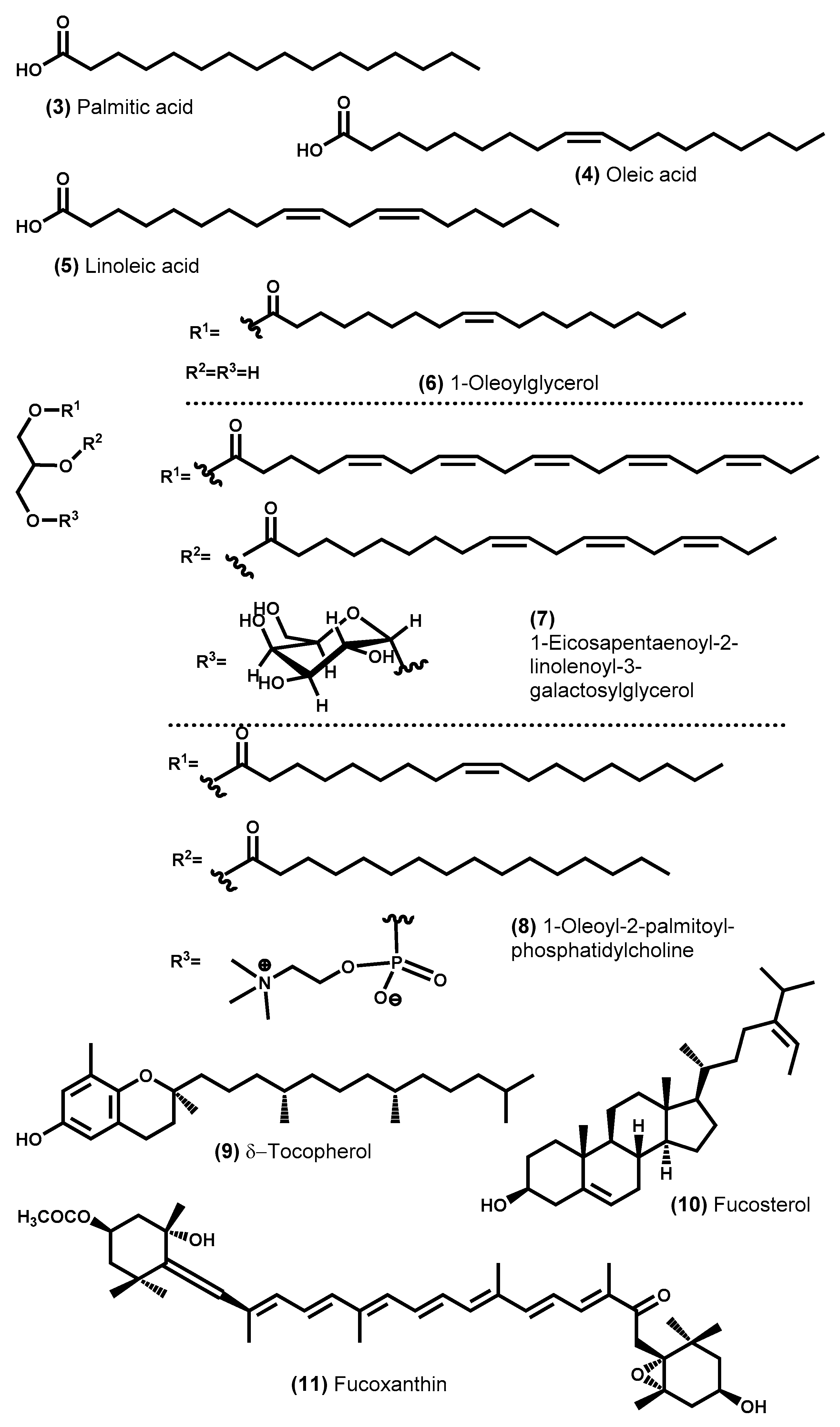{kind=link}
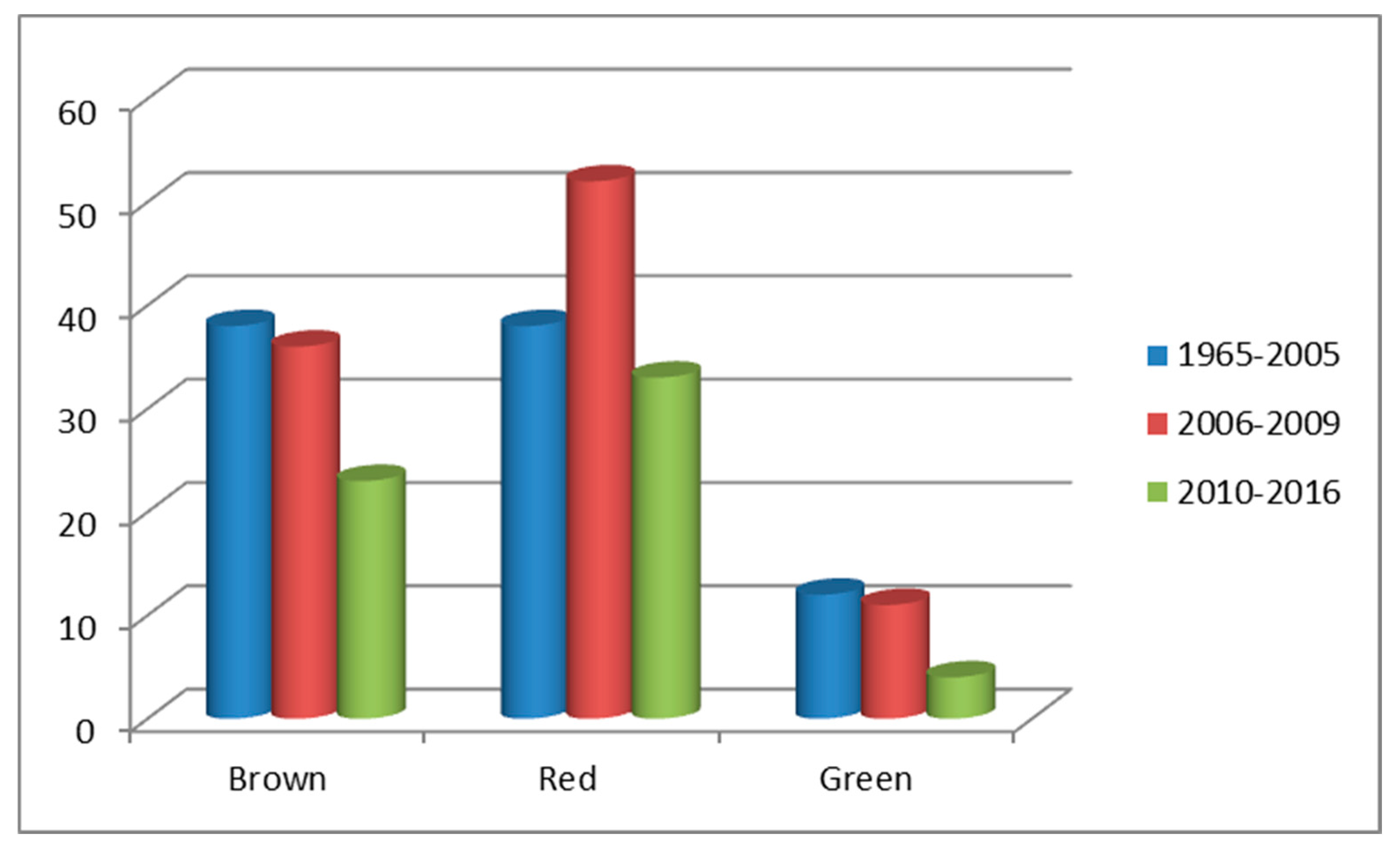{kind=link}
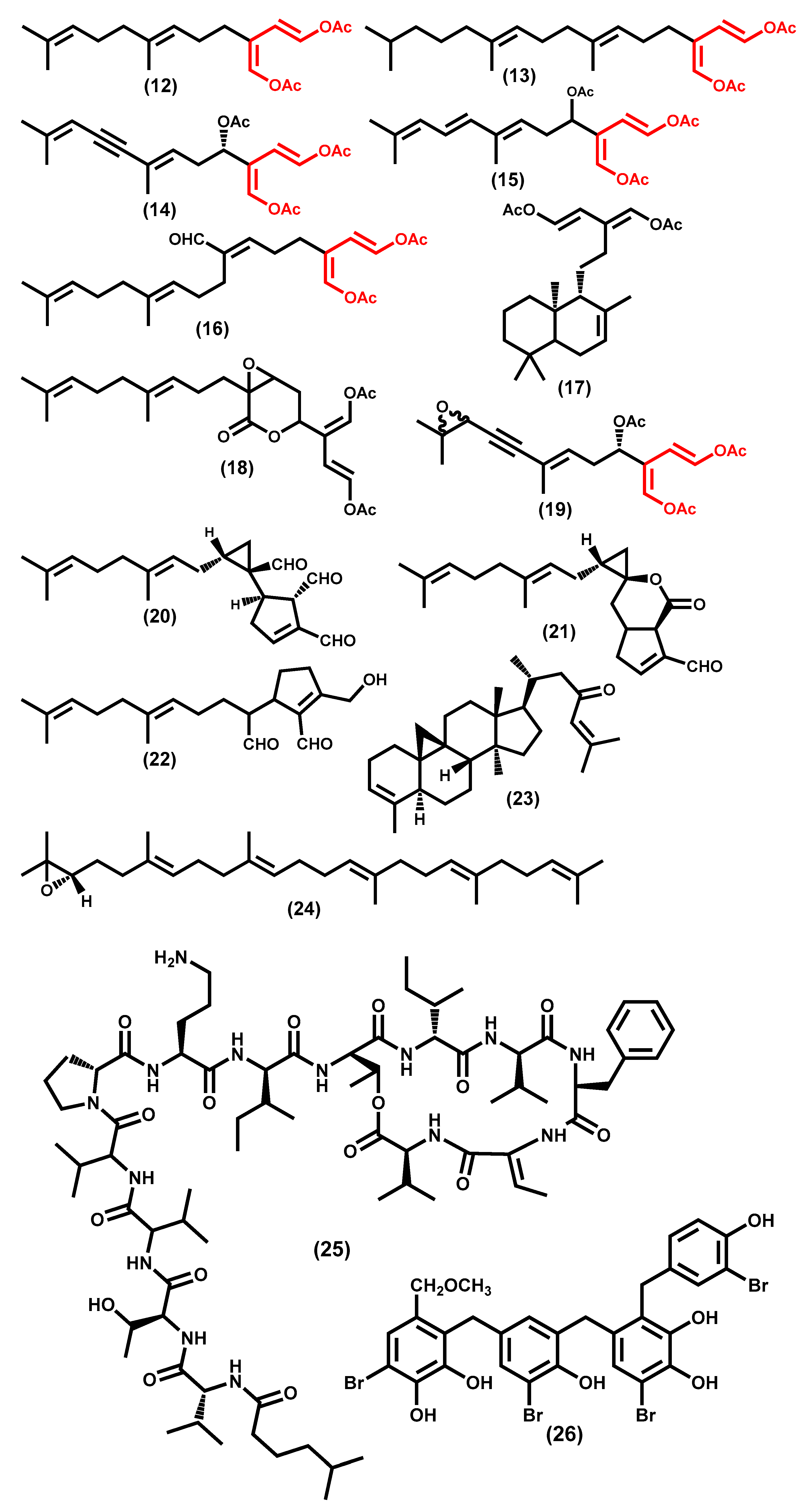{kind=link}
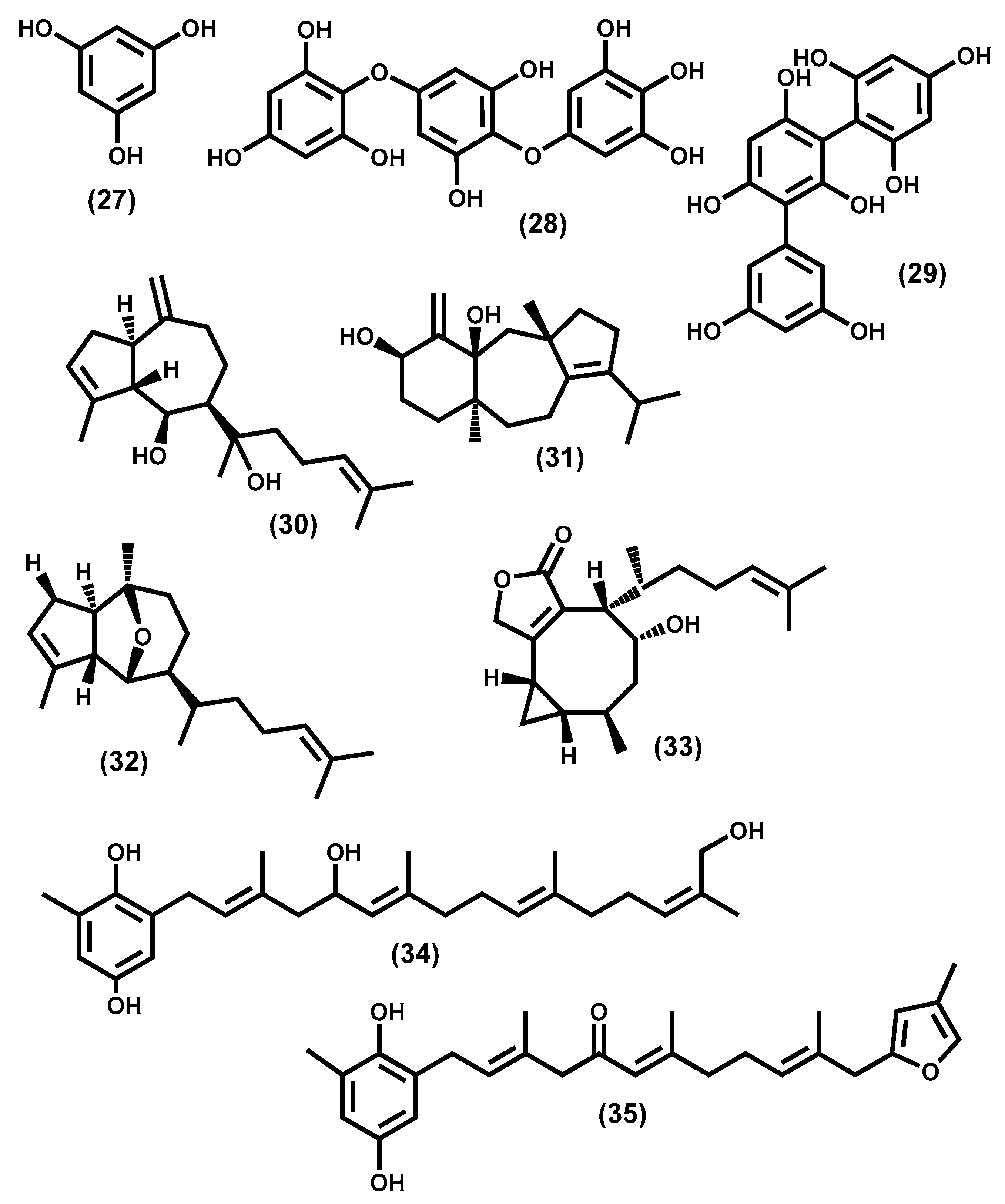{kind=link}
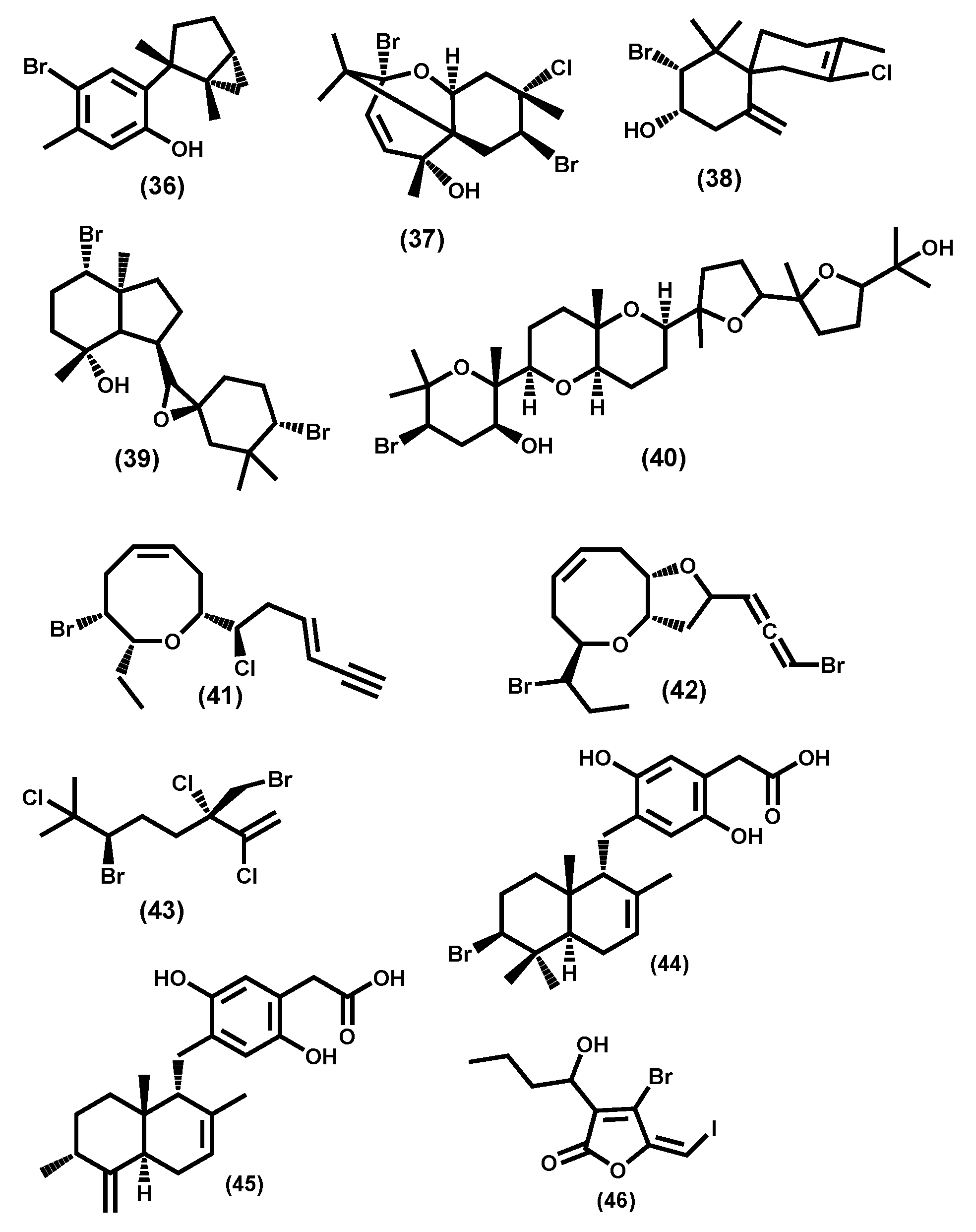{kind=link}
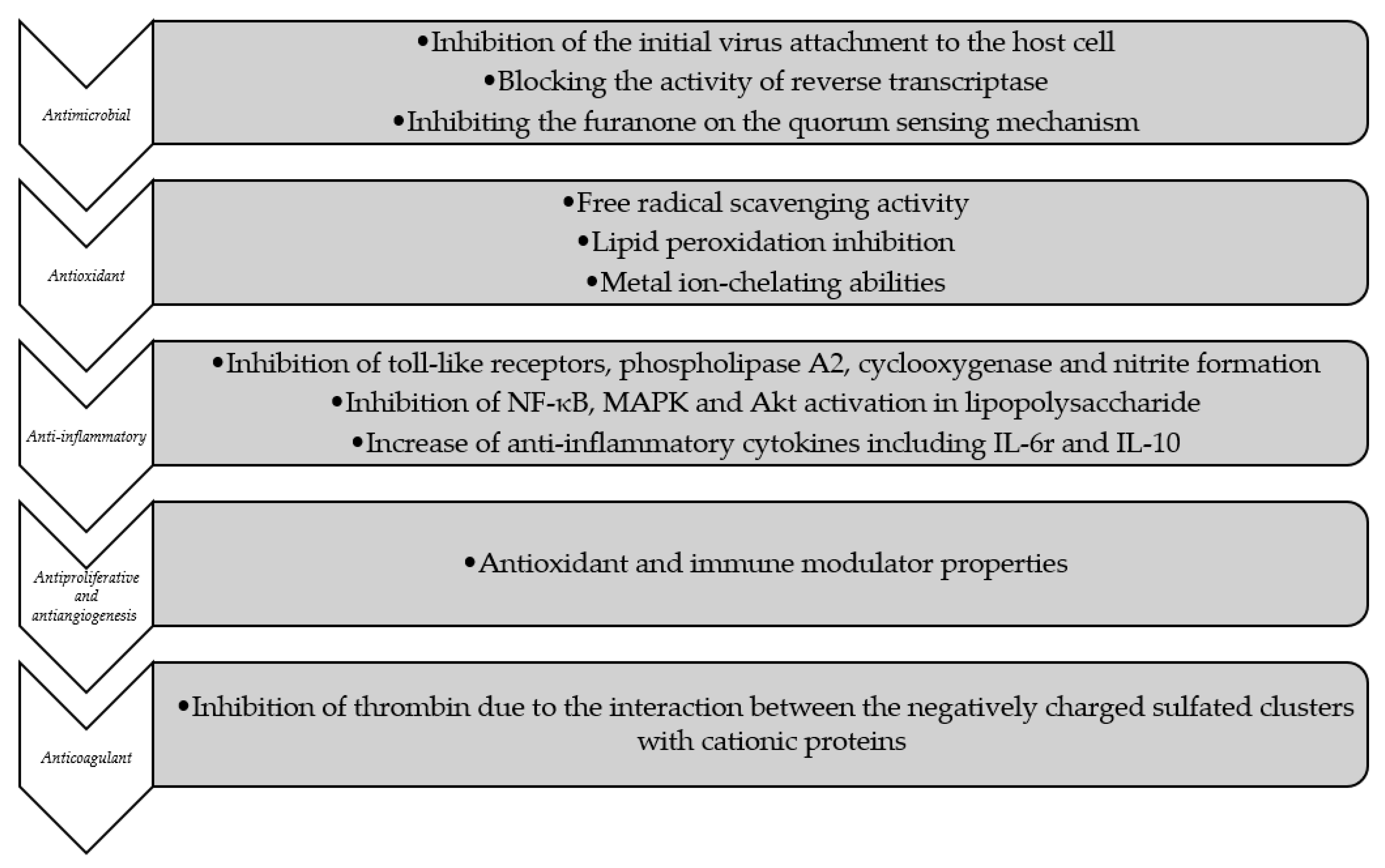{kind=link}
| Metabolites Classes | Green (Chlorophyta) | Brown (Ochrophyta) | Red (Rhodophyta) |
|---|---|---|---|
| Total Carbohydrates (% dw) | 29.8–58.1 [29,31,32] | 12.2–56.4 [31,32,33] | 34.6–71.2 [29,31,32] |
| Minerals (ash,% dw) | 11–73% [29,31,34,35,36] | 17–44% [31,34,36,37,38,39] | 7–37% [31,34,36,37,39] |
| Macrominerals (% dw) | 0.2–2.4 | 1.3–7.0 | 0.3–10 |
| Na | 1.1–2.8 | 0.9–11.5 | 0.2–9.2 |
| K | 0.09–4.9 | 0.4–3.0 | 0.2–1.9 |
| Ca | 0.4–3.7 | 0.1–1.2 | 0.1–1.7 |
| Mg | 0.09–0.25 | 0.07–0.4 | 0.1–0.7 |
| P | [31,35,36,40] | [31,36,39,40] | [31,36,39,40] |
| Trace elements (mg/kg dw) | 98–5800 | 16–1854 | 366–2110 |
| Fe | 3.0–32.7 | 6.8–154 | 23–72 |
| Zn | 0.5–32.7 | 0.8–38.6 | 1.99–34.6 |
| Cu | 26–180 | 2.7–547 | 4.1–392 |
| Mn | 23–480 | 59–11096 | 22–340 |
| I | [31,35,36,40] | [31,36,38,40] | [31,36,40] |
| Proteins (% dw) | 8.7–32.7 [29,31,41,42,43] | 4.3–24.0 [28,30,31,38,41,42,43,44] | 8.0–47.0 [28,29,31,41,42,43,44] |
| Lipids (total lipids%, dw) | 0.2–4.1 [16,29,31,34,44,45] | 0.3–4.5 [16,30,31,34,37] | 0.12–3.8 [16,29,31,34,37,44,45] |
| Saturated fatty acids (% of total fatty acid) | 34–77 [28,34,46] | 20–50 [31,34] | 25.5–76 [29,34,47] |
| Mono-unsaturated fatty acids (% of total fatty acid) | 12.2–23 [29,46] | 11–36 [30,34] | 10.6–35.7 [29,34,47] |
| Poly-unsaturated fatty acids (% of total fatty acid) | 6.6–39 [29,34,46] | 20–67 [30,34] | 12.1–68 [29,34,47] |
| Polysaccharides, Distribution (Function) | Monomeric Composition * (Glycosidic Linkage) | Other Groups Substituents | Other Characteristics |
|---|---|---|---|
| Fucoidans Brown algae (structural) | Type I: L-Fucose (α-1→3) Type II: L-Fucose alternating (α-1→3 and α-1→4) | Sulfated at C-4 (mainly) and C-2 Sulfated at C-2 and at alternating units also at C-3 | Molecular weight, degree of branching, sulfation ratio and presence of other monosaccharide (xylose, galactose, uronic acid) variable with the species |
| Agars Red algae (mainly structural) | Agarose type: β-d-Galactose, 3,6-anhydro-α-l-galactose (alternating β-1→4; α-1→3); Agaropectin: alternating d- and l-galactose (alternating β-1→4; α-1→3) | Sulfate, methoxy, and pyruvate substituents at C-2, C-6 and C-4 or C-3 | The degree of substitution like sulfation is species specific such as the proportion of agarose and agaropectin. |
| Carrageenans Red algae (mainly structural) | β-d-Galactose and 3,6-anhydro-α-d-galactose (alternating β-1→4 and α-1→3) | Sulfate group: only at C-4 of β-d-galactose (κ-carrageenan); accumulate with C-2 of 3,6-anhydro-α-d-galactose (ι-carrageenan) | Carrageenans content and type is species specific and depends on ecophysiological factors. There are other less abundant and less commercially relevant types of carrageenans (e.g., θ-, μ-, and β-carrageenans) |
| Ulvans Green algae (mainly structural) | A3s type: α-l-Rhamnose and β-d-glucuronic acid (1→4) B3s type: α-l-Rhamnose and α-l-iduronic acid (1→4) | Sulfate groups mainly at C-3 but also at both C-2 and C-3 at rhamnose units | Other sequences were determinate involving xylose and xylose-2-sulfate instead uronic acids residues (U3s and U2′s3s types) and a sequence like A3s but where glucuronic acid occur as branches on C-2 of rhamnose (A2g3s type) |
| Alginates Brown algae (mainly structural) | β-d-Mannuronic acid and α-l-guluronic acid (1→4) | Each monomer has a terminus COOH group | The monomers can be organized into homopolymers or hetero-polymers. The α-1→4 linkages give a bent and rigid chain while the β-1→4 linkage gives a flexible and linear chain. |
| Laminarin or laminaran Brown algae (storage) | d-Glucose (β-1→3) | Some residues of mannitol and/or uronic acids and occasional β-1→6-linkages in branching points as interchain linkages |
| Pharmacological Effects | Type of Seaweed | Ref. |
|---|---|---|
| Anti-inflammatory | Methanolic extracts of the brown seaweeds Padina tetrastomatica, Dictyota menstrualis, Sargassum wightii, and Spatoglossum schroederi | [272,273] |
| Red seaweeds Gracilaria edulis and Dichotomaria obtusata | [255,274] | |
| Green seaweed Caulerpa racemosa | [274] | |
| Antioxidant | Green seaweed Ulva lactuca | [275] |
| Brown alga Ecklonia cava and Pelvetia siliquosa | [276,277] | |
| Aqueous extract of the green seaweeds Halimeda incrassata | [278] | |
| Fucoidan from brown seaweeds | [279,280] | |
| Antitumor | Brown seaweeds Undaria pinnatifida and Fucus vesiculosus | [281,282] |
| Red seaweeds Chondrus ocellatus and Gelidiella acerosa | [278,283,284] | |
| Hypoglycemic | Brown seaweeds Ecklonia kurome, Ascophyllum nodosum, Eisenia bicyclis, Undaria pinnatifida, Ecklonia cava, and Saccharina japonica | [285,286,287,288,289,290] |
| Red seaweed Sargassum coreanum, Gracilaria birdiae, and Plocamium telfairiae | [289,291,292] | |
| Hypolipidemic | Brown seaweeds such as E. bicyclis and U. pinnatifida | [290] |
| Red seaweeds Gracilaria birdiae and Plocamium telfairiae | [291,292] | |
| Green seaweed Ulva lactuca and Caulerpa taxifolia | [293,294] | |
| Fucoxanthin and Fucoidan from brown seaweeds | [294,295,296] | |
| Anticoagulant | Brown seaweed Spatoglossum schroederi Blue-green algae Arthospira pratensis | [297,298,299] |
| Green seaweed Monostroma angicava | [300] |
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Salehi, B.; Sharifi-Rad, J.; Seca, A.M.L.; Pinto, D.C.G.A.; Michalak, I.; Trincone, A.; Mishra, A.P.; Nigam, M.; Zam, W.; Martins, N. Current Trends on Seaweeds: Looking at Chemical Composition, Phytopharmacology, and Cosmetic Applications. Molecules 2019, 24, 4182. https://doi.org/10.3390/molecules24224182
Salehi B, Sharifi-Rad J, Seca AML, Pinto DCGA, Michalak I, Trincone A, Mishra AP, Nigam M, Zam W, Martins N. Current Trends on Seaweeds: Looking at Chemical Composition, Phytopharmacology, and Cosmetic Applications. Molecules. 2019; 24(22):4182. https://doi.org/10.3390/molecules24224182
Chicago/Turabian StyleSalehi, Bahare, Javad Sharifi-Rad, Ana M. L. Seca, Diana C. G. A. Pinto, Izabela Michalak, Antonio Trincone, Abhay Prakash Mishra, Manisha Nigam, Wissam Zam, and Natália Martins. 2019. "Current Trends on Seaweeds: Looking at Chemical Composition, Phytopharmacology, and Cosmetic Applications" Molecules 24, no. 22: 4182. https://doi.org/10.3390/molecules24224182