Oleanolic Acid Acetate Exerts Anti-Inflammatory Activity via IKKα/β Suppression in TLR3-Mediated NF-κB Activation

 , , , ,
, , , , {kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Materials and Methods
2.1. Preparation of OAA from V. angularis
2.2. Cell Culture
2.3. Toll-Like Receptor (TLR) Activation Assay
2.4. Cell Viability
2.5. Western Blot Analysis
2.6. RNA Isolation, Cdna Synthesis, and Quantitative Real-Time PCR
2.7. Immunoprecipitation and Biotin–Streptavidin Pull-Down
2.8. Statistical Analysis
3. Results
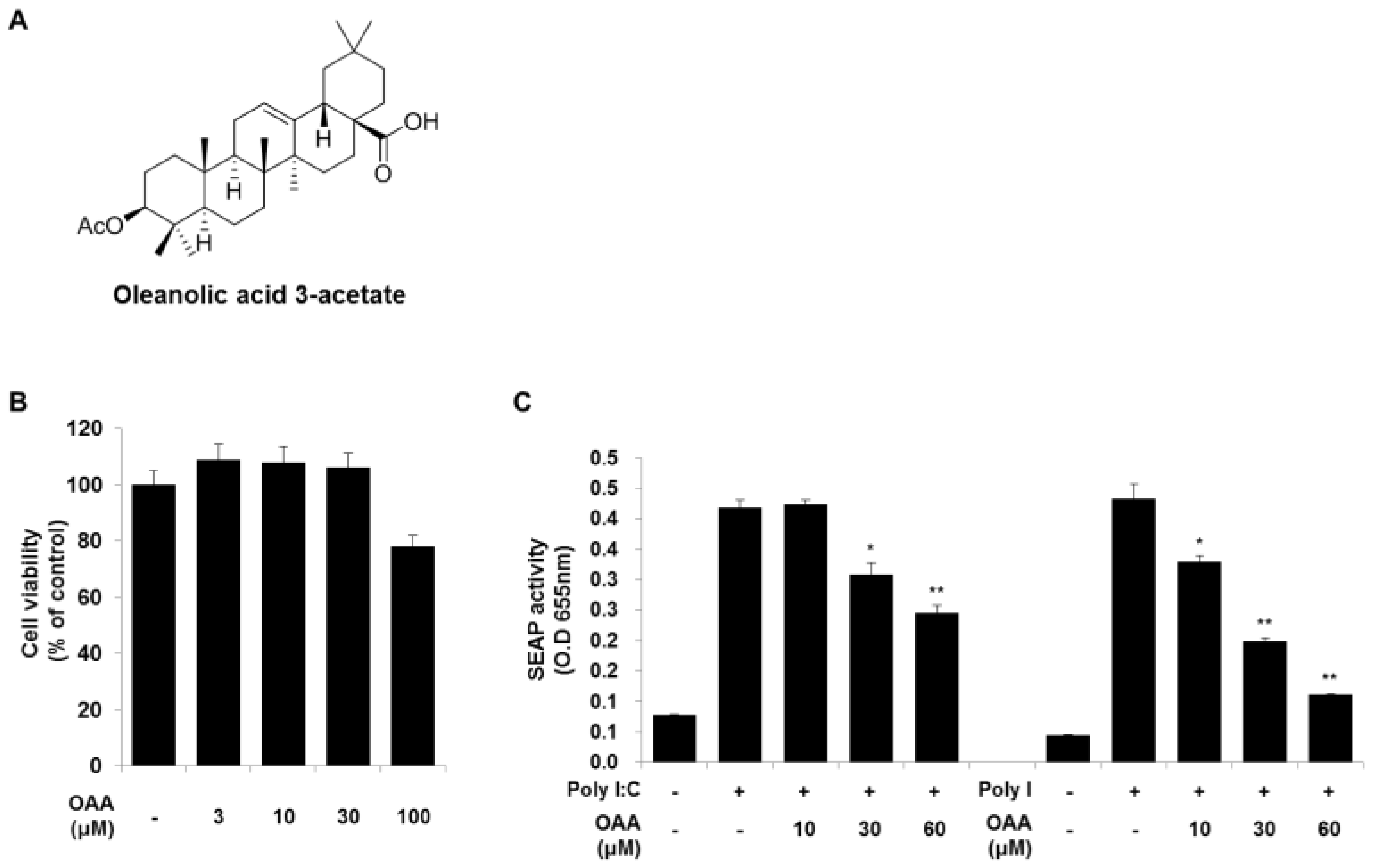
3.1. The Effect of OAA on TLR Activation in Human Monocytes
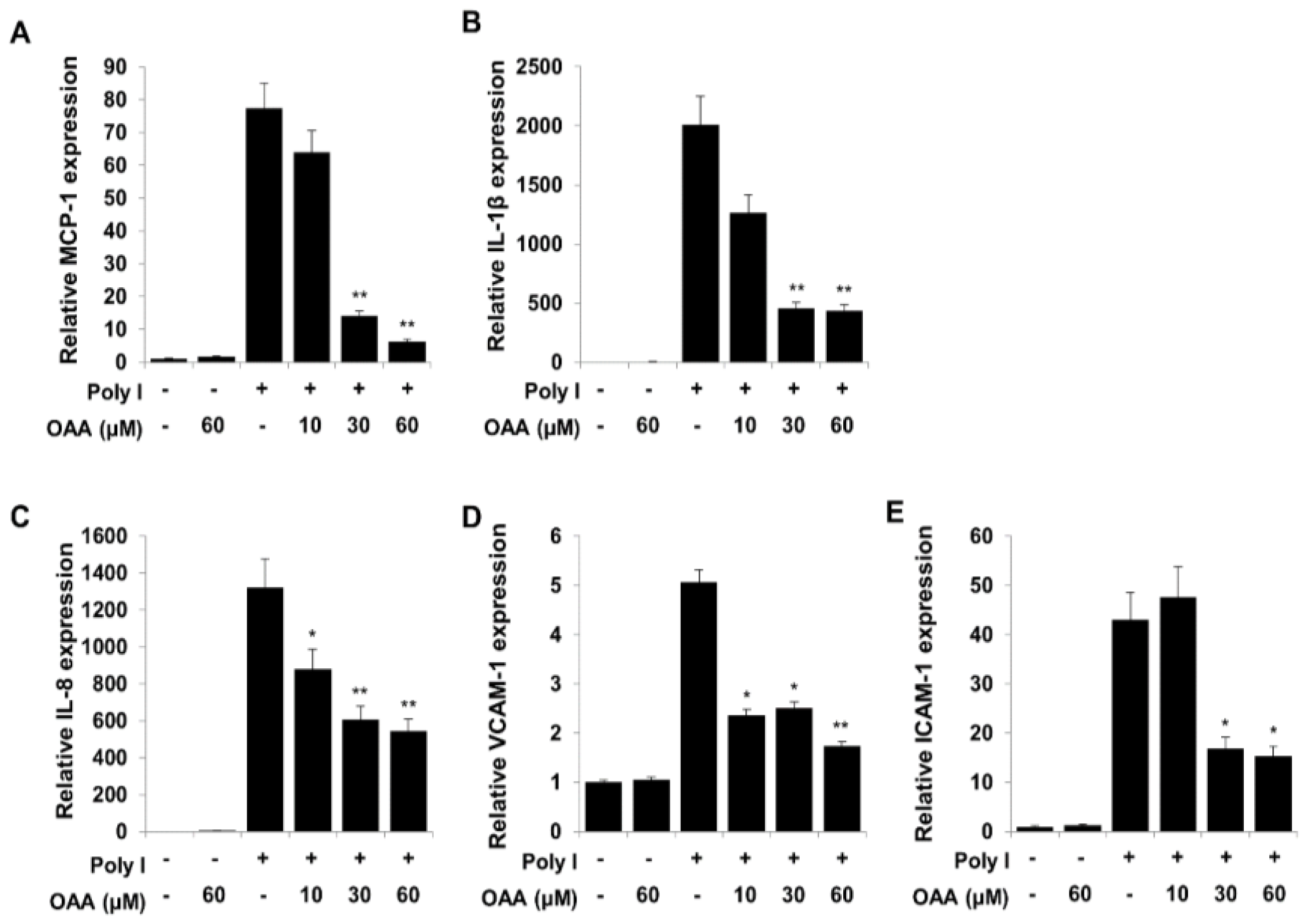
3.2. OAA Inhibits the TLR3-Mediated mRNA Expression of Proinflammatory Cytokines, Chemokines, and Proadhesive Molecules
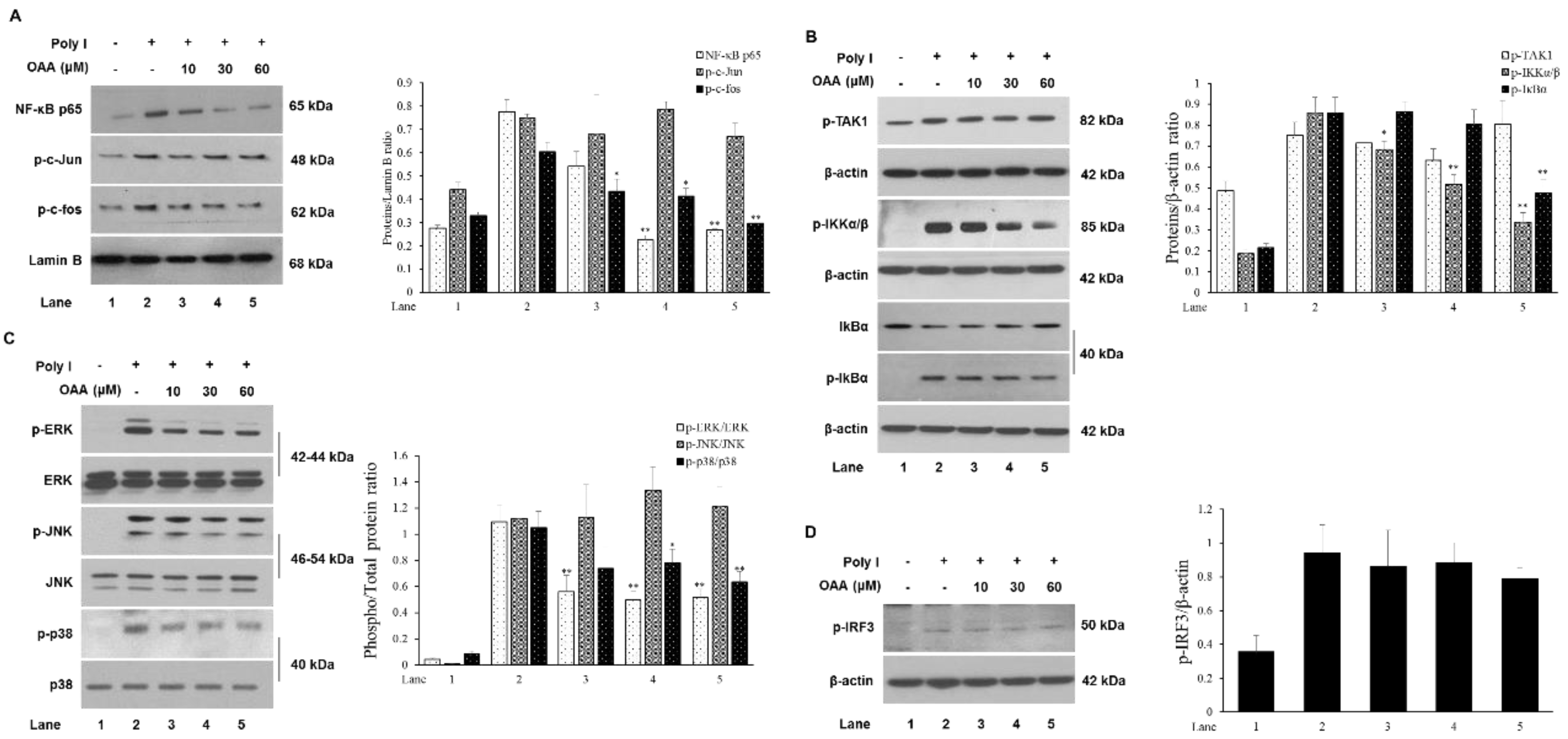
3.3. OAA Inhibits TLR3-Mediated Signaling
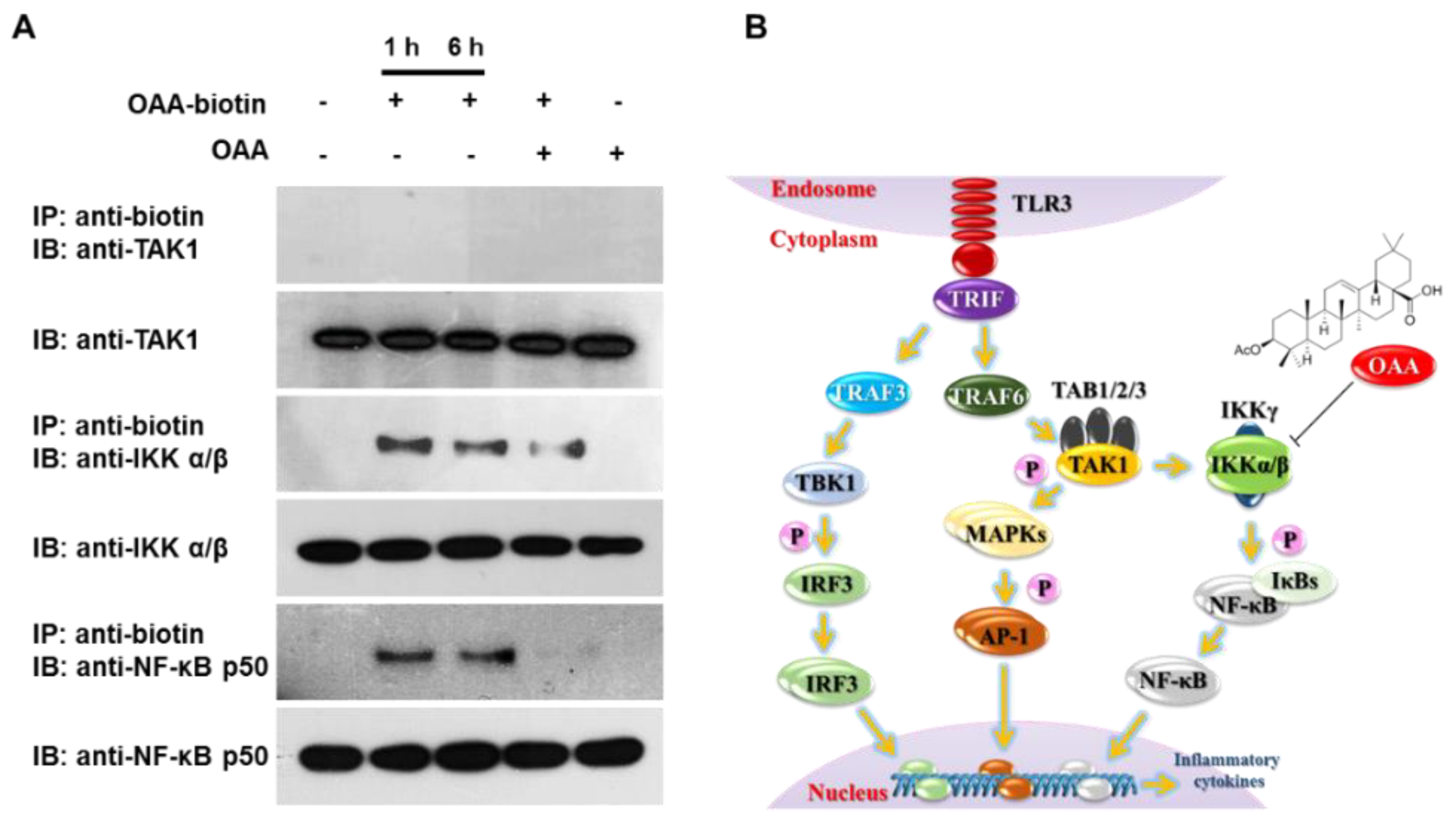
3.4. OAA Binds to IKKα/β Molecules
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Conflicts of Interest
References
- Aderem, A.; Underhill, D.M. Mechanisms of phagocytosis in macrophages. Annu. Rev. Immunol. 1999, 17, 593–623. [Google Scholar] [CrossRef] [PubMed]
- Aderem, A.; Ulevitch, R.J. Toll-like receptors in the induction of the innate immune response. Nature. 2000, 406, 782–787. [Google Scholar] [CrossRef] [PubMed]
- Takeda, K.; Akira, S. Microbial recognition by Toll-like receptors. J. Dermatol. Sci. 2004, 34, 73–82. [Google Scholar] [CrossRef] [PubMed]
- Vogel, S.N.; Fitzgerald, K.A.; Fenton, M.J. TLRs: differential adapter utilization by toll-like receptors mediates TLR-specific patterns of gene expression. Mol. Interv. 2003, 3, 466–477. [Google Scholar] [CrossRef] [PubMed]
- Takeda, K.; Akira, S. Toll-like receptors in innate immunity. Int. Immunol. 2005, 17, 1–14. [Google Scholar] [CrossRef]
- Sawa, Y.; Ueki, T.; Hata, M.; Iwasawa, K.; Tsuruga, E.; Kojima, H.; Ishikawa, H.; Yoshida, S. LPS-induced IL-6, IL-8, VCAM-1, and ICAM-1 expression in human lymphatic endothelium. J. Histochem. Cytochem. 2008, 56, 97–109. [Google Scholar] [CrossRef]
- Dong, T.; Li, C.; Wang, X.; Dian, L.; Zhang, X.; Li, L.; Chen, S.; Cao, R.; Huang, N.; He, S.; et al. Ainsliadimer A selectively inhibits IKKα/β by covalently binding a conserved cysteine. Nat. Commun. 2015, 6, 6522. [Google Scholar] [CrossRef]
- Surh, Y.J. Cancer chemoprevention with dietary phytochemicals. Nat. Rev. Cancer. 2003, 3, 768–780. [Google Scholar] [CrossRef]
- Bisoendial, R.J.; Stroes, E.S.; Tak, P.P. Where the immune response meets the vessel wall. Neth. J. Med. 2009, 67, 328–333. [Google Scholar]
- Reynolds, C.M.; Roche, H.M. Conjugated linoleic acid and inflammatory cell signaling. Prostaglandins Leukot. Essent. Fatty Acids. 2010, 82, 199–204. [Google Scholar] [CrossRef]
- Rico, M.C.; Rough, J.J.; Del Carpio-Cano, F.E.; Kunapuli, S.P.; DeLa Cadena, R.A. The axis of thrombospondin-1, transforming growth factor beta and connective tissue growth factor: An emerging therapeutic target in rheumatoid arthritis. Curr. Vasc. Pharmacol. 2010, 8, 338–343. [Google Scholar] [CrossRef] [PubMed]
- Oh, H.M.; Lee, S.W.; Yun, B.R.; Hwang, B.S.; Kim, S.N.; Park, C.S.; Jeoung, S.H.; Kim, H.K.; Lee, W.S.; Rho, M.C. Vigna angularis inhibits IL-6-induced cellular signaling and ameliorates collagen-induced arthritis. Rheumatology. 2014, 53, 56–64. [Google Scholar] [CrossRef] [PubMed]
- Mukai, Y.; Sato, S. Polyphenol-containing azuki bean (Vigna angularis) seed coats attenuate vascular oxidative stress and inflammation in spontaneously hypertensive rats. J. Nutr. Biochem. 2011, 22, 16–21. [Google Scholar] [CrossRef] [PubMed]
- Lim, H.J.; Jang, H.J.; Bak, S.G.; Lee, S.; Lee, S.W.; Lee, K.M.; Lee, S.J.; Rho, M.C. In vitro inhibitory effects of cirsiliol on IL-6-induced STAT3 activation through anti-inflammatory activity. Bioorg. Med. Chem. Lett. 2019, 29, 1586–1592. [Google Scholar] [CrossRef] [PubMed]
- Marshall-Clarke, S.; Downes, J.E.; Haga, I.R.; Bowie, A.G.; Borrow, P.; Pennock, J.L.; Grencis, R.K.; Rothwell, P. Polyinosinic acid is a ligand for toll-like receptor 3. J. Biol. Chem. 2007, 282, 24759–24766. [Google Scholar] [CrossRef] [PubMed]
- Luo, J.L.; Kamata, H.; Karin, M. IKK/NF-kappaB signaling: Balancing life and death—A new approach to cancer therapy. J. Clin. Investig. 2005, 115, 2625–2632. [Google Scholar] [CrossRef] [PubMed]
- Adhikari, A.; Xu, M.; Chen, Z.J. Ubiquitin-mediated activation of TAK1 and IKK. Oncogene 2007, 26, 3214–3226. [Google Scholar] [CrossRef] [Green Version]
- Eferl, R.; Wagner, E.F. A double-edged sword in tumorigenesis. Nat. Rev. Cancer. 2003, 3, 859–868. [Google Scholar] [CrossRef]
- Wang, A.; Al-Kuhlani, M.; Johnston, S.C.; Ojcius, D.M.; Chou, J.; Dean, D. Transcription factor complex AP-1 mediates inflammation initiated by Chlamydia pneumoniae infection. Cell Microbiol. 2013, 15, 779–794. [Google Scholar] [CrossRef]
- Oh, H.M.; Lee, S.W.; Park, M.H.; Kim, M.H.; Ryu, Y.B.; Kim, M.S.; Kim, H.H.; Park, K.H.; Lee, W.S.; Park, S.J.; et al. Norkurarinol inhibits toll-like receptor 3 (TLR3)-mediated pro-inflammatory signaling pathway and rotavirus replication. J. Pharmacol. Sci. 2012, 118, 161–170. [Google Scholar] [CrossRef]
- Brown, K.E.; Keller, P.J.; Ramialison, M.; Rembold, M.; Stelzer, E.H.; Loosli, F.; Wittbrodt, J. Nlcam modulates midline convergence during anterior neural plate morphogenesis. Dev. Biol. 2009, 339, 14–25. [Google Scholar] [CrossRef] [PubMed]
- Palusinska-Szysz, M.; Janczarek, M. Innate immunity to Legionella and toll-like receptors–review. Folia Microbiologica. 2010, 55, 508–514. [Google Scholar] [CrossRef] [PubMed]
- Alexopoulou, L.; Holt, A.C.; Medzhitov, R.; Flavell, R.A. Recognition of double-stranded RNA and activation of NF-κB by Toll-like receptor 3. Nature. 2001, 413, 732–738. [Google Scholar] [CrossRef] [PubMed]
- Ernst, M.K.; Dunn, L.L.; Rice, N.R. The PEST-like sequence of IκBα is responsible for inhibition of DNA binding but not for cytoplasmic retention of c-Rel or RelA homodimers. Mol. Cell Biol. 1995, 15, 872–882. [Google Scholar] [CrossRef]
- Choi, J.K.; Oh, H.M.; Lee, S.; Park, J.W.; Khang, D.W.; Lee, S.W.; Lee, W.S.; Rho, M.C.; Kim, S.H. Oleanolic acid acetate inhibits atopic dermatitis and allergic contact dermatitis in a murine model. Toxicol. Appl. Pharmacol. 2013, 269, 72–80. [Google Scholar] [CrossRef]
- Kim, M.S.; Han, J.Y.; Kim, S.H.; Jeon, D.; Kim, H.Y.; Lee, S.W.; Rho, M.C.; Lee, K. Oleanolic acid acetate attenuates polyhexamethylene guanidine phosphate-induced pulmonary inflammation and fibrosis in mice. Respir. Physiol. Neurobiol. 2018, 252–253. [Google Scholar] [CrossRef]
- Hwang, J.T.; Oh, H.M.; Kim, M.H.; Jeong, H.J.; Lee, C.; Jang, H.J.; Lee, S.W.; Park, C.S.; Rho, M.C. Simultaneous quantitation and validation of triterpenoids and phytosteroids in Phaseolus angularis Seeds. Molecules 2014, 19, 10309–10319. [Google Scholar] [CrossRef]
Sample Availability: Samples of the compounds are not available from the authors. |




© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Lim, H.J.; Jang, H.-J.; Kim, M.H.; Lee, S.; Lee, S.W.; Lee, S.-J.; Rho, M.-C. Oleanolic Acid Acetate Exerts Anti-Inflammatory Activity via IKKα/β Suppression in TLR3-Mediated NF-κB Activation. Molecules 2019, 24, 4002. https://doi.org/10.3390/molecules24214002
Lim HJ, Jang H-J, Kim MH, Lee S, Lee SW, Lee S-J, Rho M-C. Oleanolic Acid Acetate Exerts Anti-Inflammatory Activity via IKKα/β Suppression in TLR3-Mediated NF-κB Activation. Molecules. 2019; 24(21):4002. https://doi.org/10.3390/molecules24214002
Chicago/Turabian StyleLim, Hyung Jin, Hyun-Jae Jang, Mi Hwa Kim, Soyoung Lee, Seung Woong Lee, Seung-Jae Lee, and Mun-Chual Rho. 2019. "Oleanolic Acid Acetate Exerts Anti-Inflammatory Activity via IKKα/β Suppression in TLR3-Mediated NF-κB Activation" Molecules 24, no. 21: 4002. https://doi.org/10.3390/molecules24214002