Characterization, Antioxidant, Anti-Aging and Organ Protective Effects of Sulfated Polysaccharides from Flammulina velutipes


Abstract
:1. Introduction
2. Results
2.1. Structural Characterization
2.1.1. The Purity and Molecular Parameters
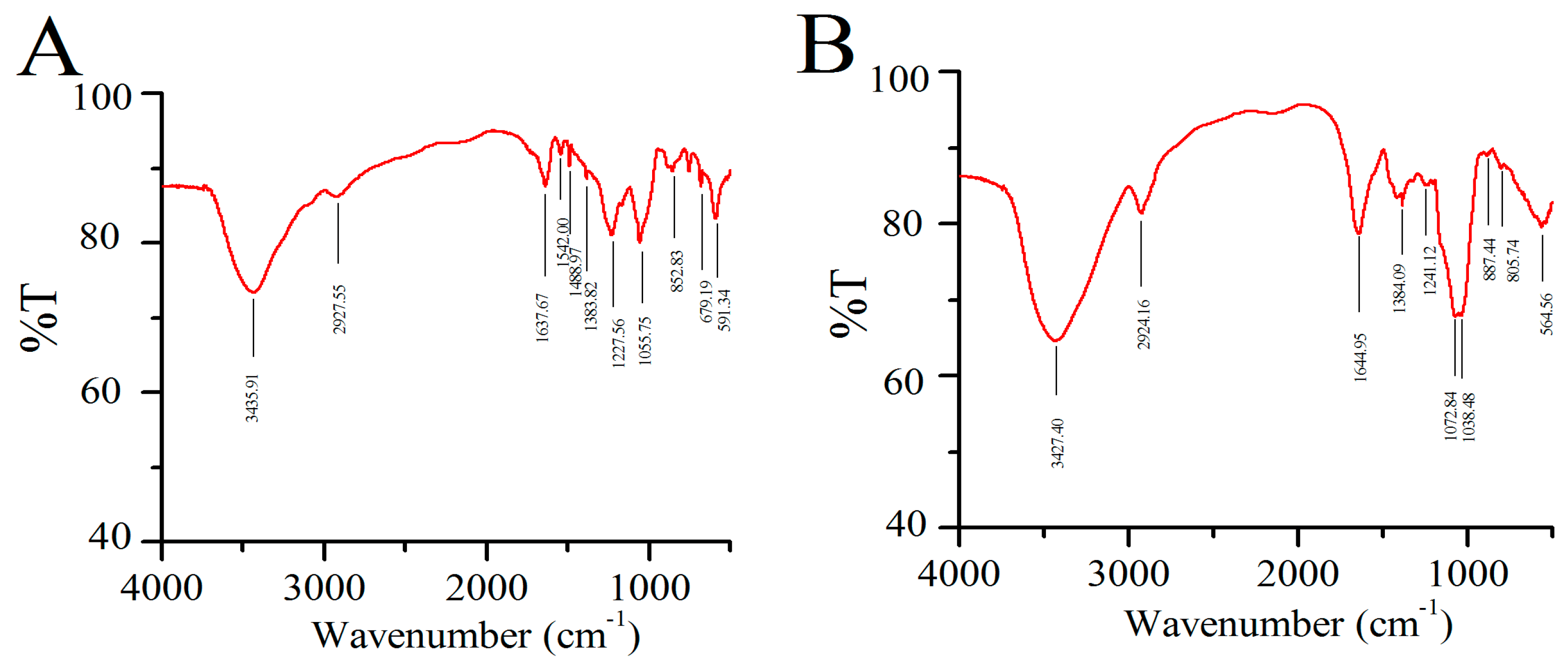
2.1.2. FT-IR Assay
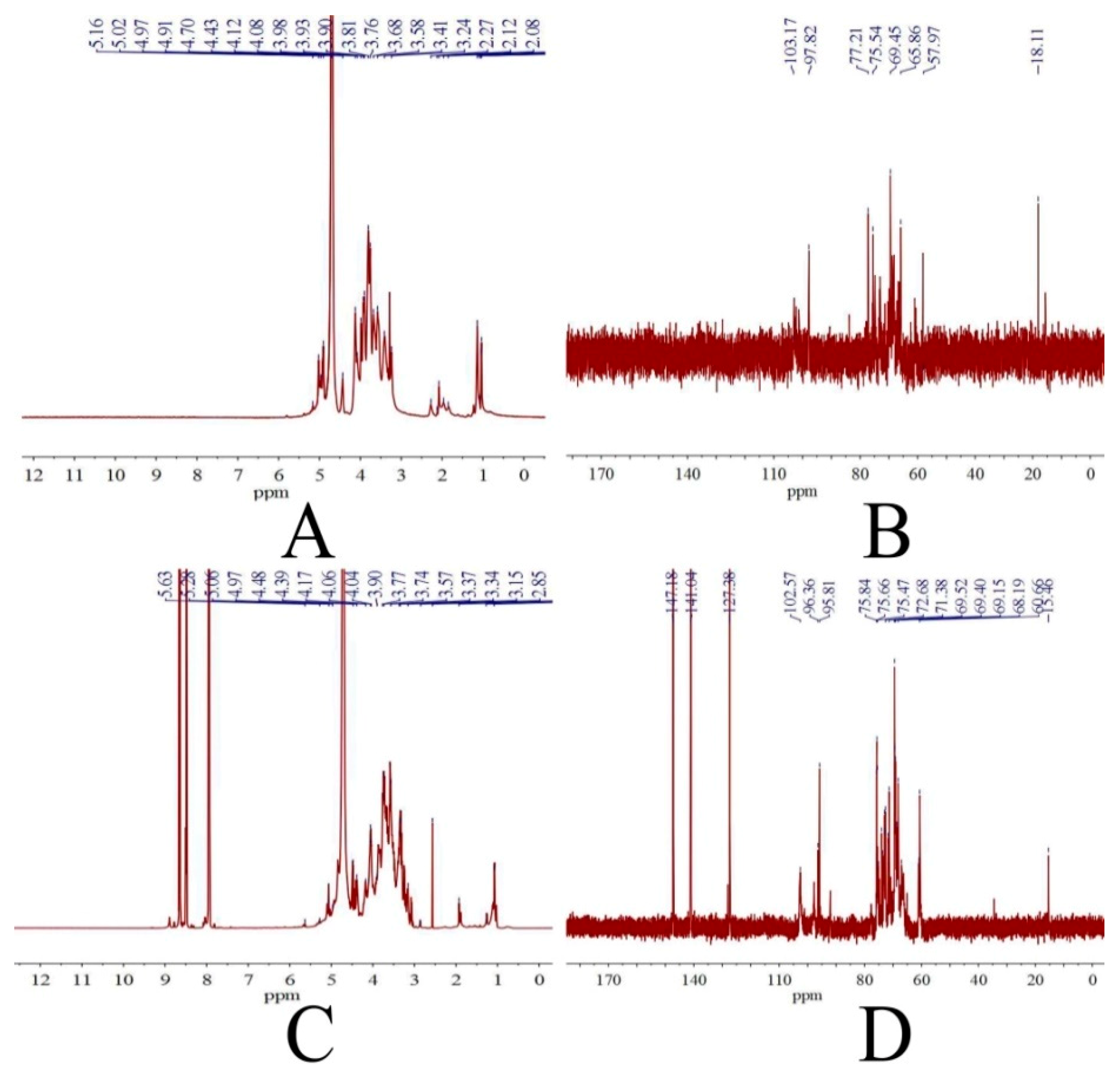
2.1.3. NMR Analysis
2.1.4. Monosaccharide Composition Analysis
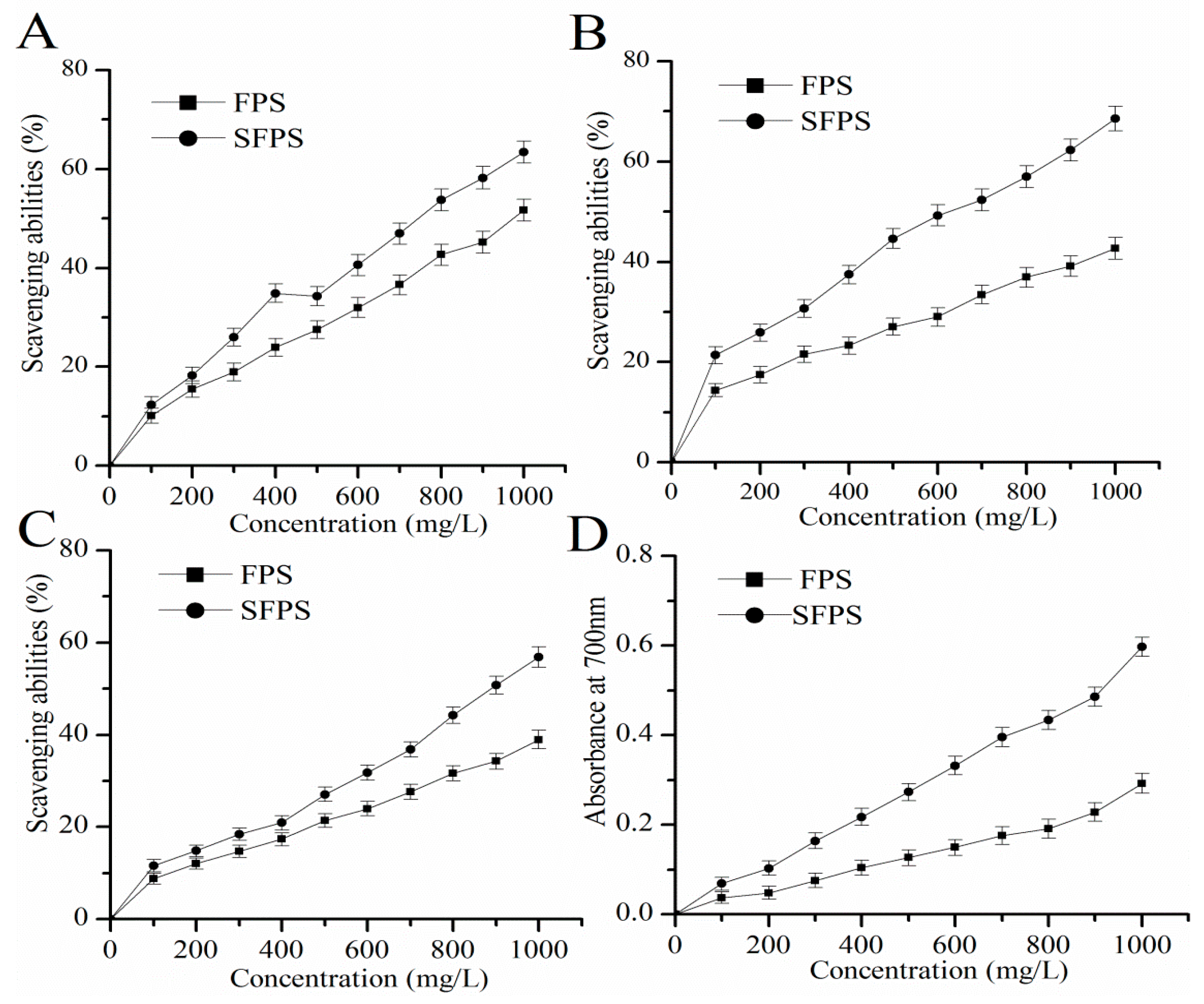
2.2. Antioxidant Capacities In Vitro
2.3. Acute and Subchronic Toxicity Analysis
2.4. Anti-Aging Capacities In Vivo
2.4.1. Body Weight and Organ Index
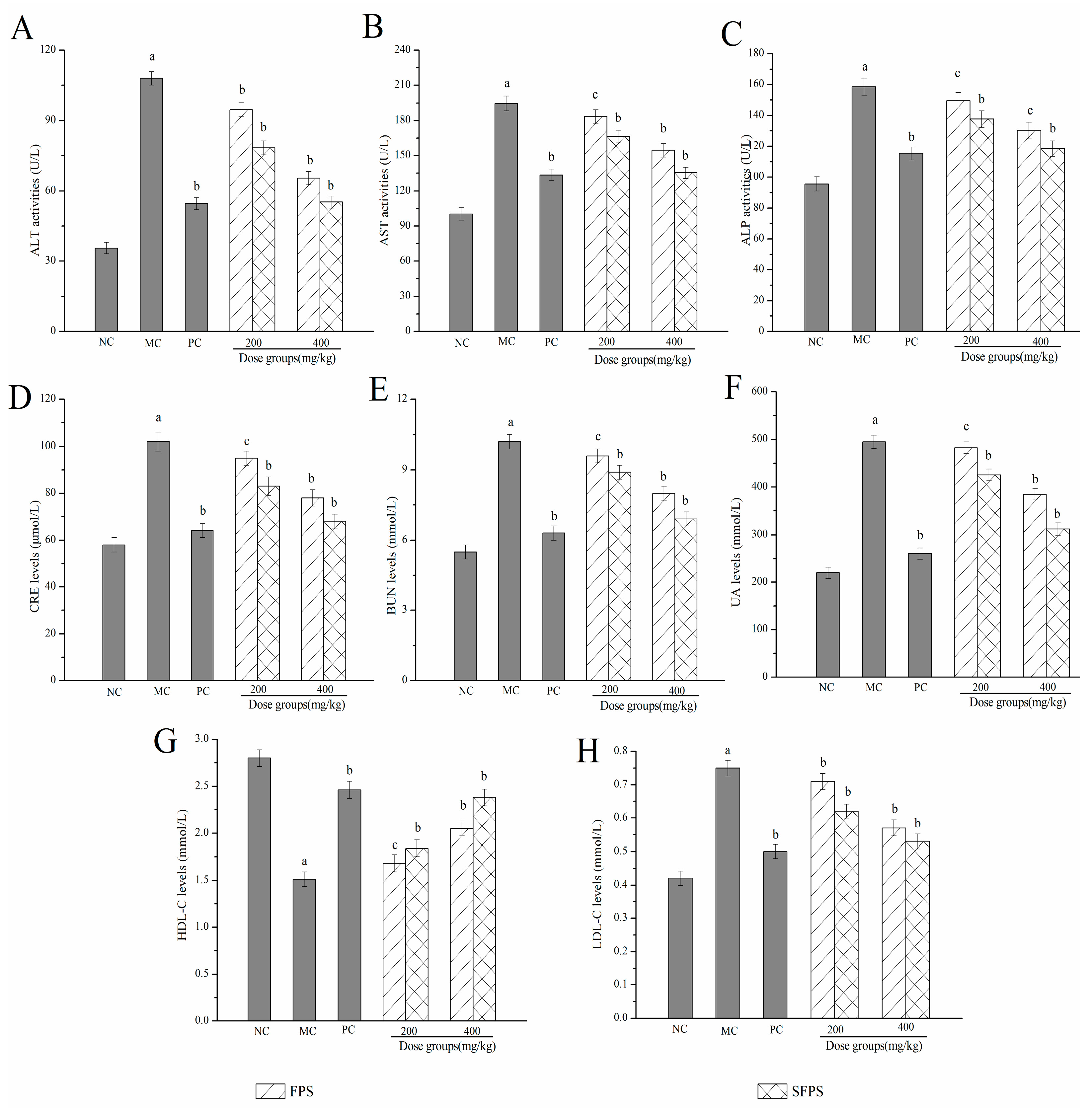
2.4.2. Effects of SFPS and FPS on Alanine Aminotransferase (ALT), Aspartate Aminotransferase (AST), Alkaline Phosphatase (ALP), Creatinine Levels (CRE), Blood Urea Nitrogen Levels (BUN), Uric Acid (UA), High Density Lipoprotein Cholesterol (HDL-C) and Low-Density Lipoprotein Cholesterol (LDL-C)
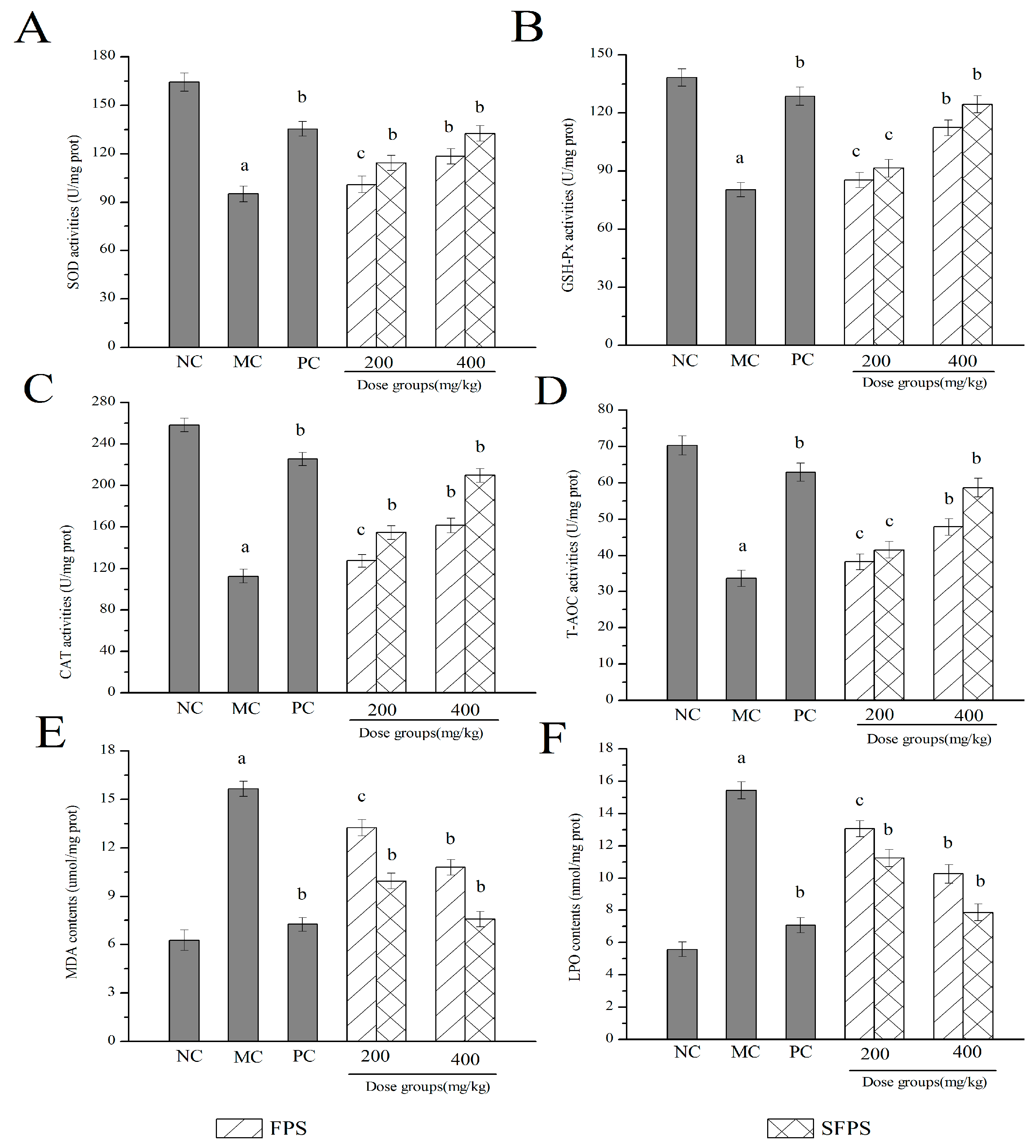
2.4.3. Effects of SFPS and FPS on Antioxidant Status
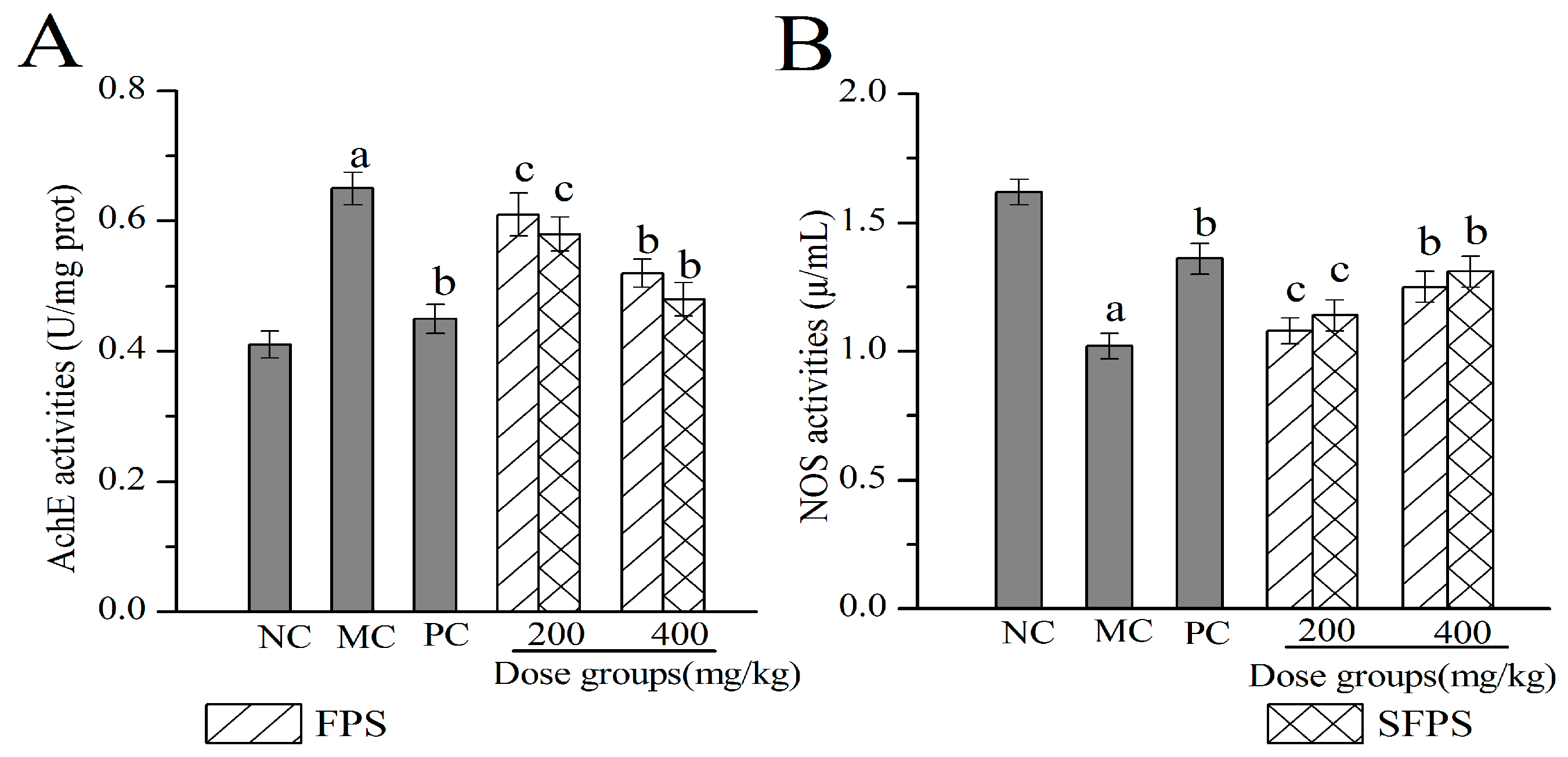
2.4.4. Effects of SFPS and FPS on Acetylcholin Esterase (Ach E) and Nitric Oxide Synthase (NOS) in the Brain
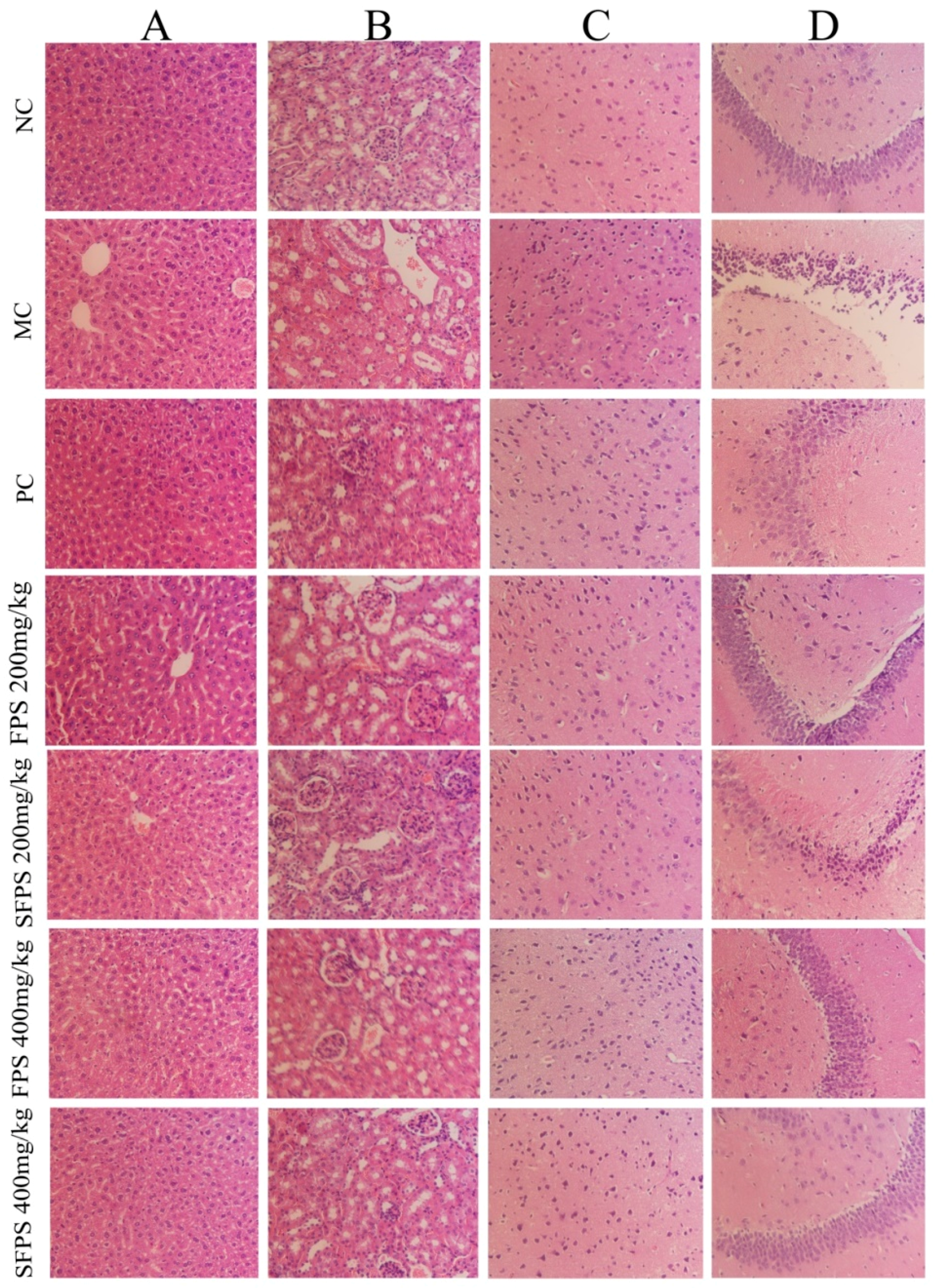
2.5. Histopathological Analysis
3. Discussion
4. Materials and Methods
4.1. Chemicals and Reagents
4.2. Preparation of FPS and SFPS
4.3. Degree of Substitution (DS)
4.4. Structural Characterization of FPS and SFPS
4.4.1. Ultraviolet (UV) Analysis and Molecular Weight Determination
4.4.2. Fourier-Transform Infrared (FT-IR) Spectroscopy Analysis
4.4.3. Nuclear Magnetic Resonance (NMR) Analysis
4.4.4. Monosaccharide Composition Analysis
4.5. In Vitro Antioxidant Activities
4.6. Acute and Subchronic Toxicity Experiment
4.7. Animal Experiments
4.8. Statistical Analysis
5. Conclusions
Author Contributions
Funding
Acknowledgments
Compliance with Ethical Standards
Conflicts of Interest
Abbreviations
References
- Sergiev, P.V.; Dontsova, O.A.; Berezkin, G.V. Theories of aging: An ever-evolving field. Acta Nat. 2015, 7, 9–18. [Google Scholar] [CrossRef]
- Kirkwood, T.B.L. Understanding the Odd Science of Aging. Cell 2005, 120, 437–447. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lan, Z.; Liu, J.P.; Chen, L.Y.; Fu, Q.; Luo, J.G.; Qu, R.; Ma, S. Danggui-Shaoyao-San ameliorates cognition deficits and attenuates oxidative stress-related neuronal apoptosis in d-galactose-induced senescent mice. J. Ethnopharmacol. 2012, 141, 386–395. [Google Scholar] [CrossRef] [PubMed]
- Haider, S.; Liaquat, L.; Shahzad, S.; Sadir, S.; Madiha, S.; Batool, Z.; Perveen, T. A high dose of short term exogenous d-galactose administration in young male rats produces symptoms simulating the natural aging process. Life Sci. 2015, 124, 110–119. [Google Scholar] [CrossRef] [PubMed]
- Vanhooren, V.; Libert, C. The mouse as a model organism in aging research: Usefulness, pitfalls and possibilities. Ageing Res. Rev. 2013, 12, 8–21. [Google Scholar] [CrossRef] [PubMed]
- Jin, S.L.; Yin, Y.G. In vivo antioxidant activity of total flavonoids from indocalamus leaves in aging mice caused by D-galactose. Food Chem. Toxicol. 2012, 50, 3814–3818. [Google Scholar] [CrossRef]
- Qu, Z.; Zhang, J.Z.; Yang, H.G.; Huo, L.Q.; Gao, J.; Chen, H.; Gao, W. Protective effect of tetrahydropalmatine against D-galactose induced memory impairment in rat. Physiol. Behav. 2015, 154, 114–125. [Google Scholar] [CrossRef]
- Gong, Y.S.; Guo, J.; Hu, K.; Gao, Y.Q.; Xie, B.J.; Sun, Z.D.; Hou, F.L. Ameliorative effect of lotus seedpod proanthocyanidins on cognitive impairment and brain aging induced by d-galactose. Exp. Gerontol. 2016, 74, 21–28. [Google Scholar] [CrossRef]
- Liochev, S.I. Reactive oxygen species and the free radical theory of aging. Free Radic. Biol. Med. 2013, 60, 1–4. [Google Scholar] [CrossRef]
- Schieber, M.; Chandel, N.S. ROS Function in Redox Signaling and Oxidative Stress. Curr. Biol. 2014, 24, 453–462. [Google Scholar] [CrossRef]
- Li, W.J.; Nie, S.P.; Peng, X.P.; Liu, X.Z.; Li, C.; Chen, Y.; Xie, M.Y. Ganoderma atrum Polysaccharide Improves Age-Related Oxidative Stress and Immune Impairment in Mice. J. Agric. Food Chem. 2012, 60, 1413–1418. [Google Scholar] [CrossRef] [PubMed]
- Fu, W.; Chen, J.L.; Cai, Y.L.; Lei, Y.F.; Chen, L.M.; Pei, L.; Ruan, J. Antioxidant, free radical scavenging, anti-inflammatory and hepatoprotective potential of the extract from Parathelypteris nipponica (Franch. et Sav.) Ching. J. Ethnopharmacol. 2010, 130, 521–528. [Google Scholar] [CrossRef] [PubMed]
- Young, S.I.; Woodside, J.V. Antioxidants in health and disease. J. Clin. Pathol. 2001, 54, 176–186. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wasser, S.P. Medicinal mushrooms as a source of antitumor and immunomodulating polysaccharides. Appl. Microbiol. Biotechnol. 2002, 60, 258–274. [Google Scholar] [PubMed]
- Jin, M.; Zhao, K.; Huang, Q.; Xu, C.; Shang, P. Isolation, structure and bioactivities of the polysaccharides from Angelica sinensis (Oliv.) Diels: A review. Carbohydr. Polym. 2012, 89, 713–722. [Google Scholar] [CrossRef] [PubMed]
- Dong, Y.R.; Cheng, S.J.; Qi, G.H.; Yang, Z.P.; Yin, S.Y.; Chen, G.T. Antimicrobial and antioxidant activities of Flammulina velutipes polysacchrides and polysacchride-iron(III) complex. Carbohydr. Polym. 2017, 161, 26–32. [Google Scholar] [CrossRef] [PubMed]
- Smiderle, F.R.; Ruthes, A.C.; van Arkel, J.; Chanput, W.; Iacomini, M.; Wichers, H.J.; Van Griensven, L.J. Polysaccharides from Agaricus bisporus and Agaricus brasiliensis show similarities in their structures and their immunomodulatory effects on human monocytic THP-1 cells. BMC Complement. Altern. Med. 2011, 11, 58. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Y.; Li, H.; Yang, X.D.; Jin, G.; Zhang, Y. Cognitive-enhancing effect of polysaccharides from Flammulina velutipes on Alzheimer’s disease by compatibilizing with ginsenosides. Int. J. Biol. Macromol. 2018, 112, 788–795. [Google Scholar] [CrossRef] [PubMed]
- Kizil, R.; Irudayaraj, J.; Seetharaman, K. Characterization of irradiated starches by using FT-Raman and FTIR spectroscopy. J. Agric. Food Chem. 2002, 50, 3912–3918. [Google Scholar] [CrossRef]
- Tu, W.S.; Zhu, J.H.; Bi, S.X.; Chen, D.H.; Song, L.Y.; Wang, L.S.; Yu, R. Isolation, characterization and bioactivities of a new polysaccharide from Annona squamosa and its sulfated derivative. Carbohydr. Polym. 2016, 152, 287–296. [Google Scholar] [CrossRef]
- Cai, W.R.; Gu, X.H.; Tang, J. Extraction, purification, and characterization of the polysaccharides from Opuntia milpa alta. Carbohydr. Polym. 2008, 71, 403–410. [Google Scholar] [CrossRef]
- Kong, Q.; Zhou, T.; Ge, J.; Wu, G.L.; Huang, J.Y.; Li, Q.H. Study on the relationship of sulfation and structural stability of polysaccharides from pumpkin pulp. Food Sci. Technol. 2009, 34, 160–163. [Google Scholar]
- Li, J.Y.; Yuan, P.F.; Wang, X.H.; Aipire, A.; Li, M.J.; Yang, J.; Zhang, F. Purification, characterization and bioactivities of polysaccharides from Pleurotus ferulae. Food Funct. 2017, 8, 1905–1914. [Google Scholar] [CrossRef] [PubMed]
- Liu, H.L.; Jiang, Y.M.; Yang, H.S.; Yang, B. Structure characteristics of an acidic polysaccharide purified from banana (Musa, nana, Lour.) pulp and its enzymatic degradation. Int. J. Biol. Macromol. 2017, 101, 299–303. [Google Scholar] [CrossRef] [PubMed]
- Meng, M.; Cheng, D.; Han, L.R.; Chen, Y.Y.; Wang, C.L. Isolation, purification, structural analysis and immunostimulatory activity of water-soluble polysaccharides from Grifola Frondosa fruiting body. Carbohydr. Polym. 2017, 157, 1134–1143. [Google Scholar] [CrossRef]
- Kou, L.; Du, M.Z.; Liu, P.J.; Zhang, B.H.; Zhang, Y.Z.; Yang, P.; Shang, M.Y.; Wang, X.D. Anti-Diabetic and Anti-Nephritic Activities of Grifola frondosa Mycelium Polysaccharides in Diet-Streptozotocin-Induced Diabetic Rats Via Modulation on Oxidative Stress. Appl. Biochem. Biotechnol. 2019, 187, 310–322. [Google Scholar] [CrossRef]
- Ullah, F.; Ali, T.; Ullah, N.; Kim, M.O. Caffeine prevents D-galactose-induced cognitive deficits, oxidative stress, neuroinflammation and neurodegeneration in the adult rat brain. Neurochem. Int. 2015, 90, 114–124. [Google Scholar] [CrossRef]
- Zhu, J.H.; Mu, X.Y.; Zeng, J.; Xu, C.Y.; Liu, J.; Zhang, M.S.; Wang, Y. Ginsenoside Rg1 Prevents Cognitive Impairment and Hippocampus Senescence in a Rat Model of D-Galactose-Induced Aging. PLoS ONE 2014, 9, e101291. [Google Scholar] [CrossRef]
- Childs, B.G.; Gluscevic, M.; Baker, D.J.; Laberge, R.M.; Marquess, D.; Dananberg, J.; van Deursen, J.M. Senescent cells: An emerging target for diseases of ageing. Nat. Rev. Drug Discov. 2017, 16, 718–735. [Google Scholar] [CrossRef]
- Xia, Z.Q. Preparation of the oligosaccharides derived from Flammulina velutipes and their antioxidant activities. Carbohydr. Polym. 2015, 118, 41–43. [Google Scholar] [CrossRef]
- Ma, Z.; Cui, F.Y.; Gao, X.; Zhang, J.J.; Zheng, L.; Jia, L. Purification, characterization, antioxidant activity and anti-aging of exopolysaccharides by Flammulina velutipes SF-06. Antonie Van Leeuwenhoek 2015, 107, 73–82. [Google Scholar] [CrossRef] [PubMed]
- Ziech, D.; Franco, R.; Pappa, A.; Panayiotidis, M.I. Reactive Oxygen Species (ROS)––Induced genetic and epigenetic alterations in human carcinogenesis. Mutat. Res. 2011, 711, 167–173. [Google Scholar] [CrossRef] [PubMed]
- Wang, Z.Y.; Wang, C.Y.; Quan, Y. Extraction of polysaccharides from Phellinus nigricans mycelia and their antioxidant activities in vitro. Carbohydr. Polym. 2014, 99, 110–115. [Google Scholar] [CrossRef] [PubMed]
- Meir, S.; Kanner, J.; Akiri, B.; Philosoph-Hadas, S. Determination and Involvement of Aqueous Reducing Compounds in Oxidative Defense Systems of Various Senescing Leaves. J. Agric. Food Chem. 1995, 43, 1813–1819. [Google Scholar] [CrossRef]
- Xiong, Q.P.; Li, X.; Zhou, R.Z.; Hao, H.R.; Li, S.L.; Jing, Y.; Shi, Y. Extraction, characterization and antioxidant activities of polysaccharides from E. corneum gigeriae galli. Carbohydr. Polym. 2014, 108, 247–256. [Google Scholar] [CrossRef] [PubMed]
- Cheng, D.Y.; Kong, H. The Effect of Lycium Barbarum Polysaccharide on Alcohol-Induced Oxidative Stress in Rats. Molecules 2011, 16, 2542–2550. [Google Scholar] [CrossRef]
- Chiu, C.H.; Chyau, C.C.; Chen, C.C.; Lin, C.H.; Cheng, C.H.; Mong, M.C. Polysaccharide extract of Cordyceps sobolifera attenuates renal injury in endotoxemic rats. Food Chem. Toxicol. 2014, 69, 281–288. [Google Scholar] [CrossRef]
- Ding, Q.Y.; Yang, D.; Zhang, W.N.; Lu, Y.M.; Zhang, M.Z.; Wang, L.M.; Chen, Y. Antioxidant and anti-aging activities of the polysaccharide TLH-3 from Tricholoma lobayense. Int. J. Biol. Macromol. 2015, 85, 133–140. [Google Scholar] [CrossRef] [PubMed]
- Govindan, S.; Johnson, E.E.R.; Christopher, J.; Shanmugam, J.; Thirumalairaj, V. Antioxidant and anti-aging activities of polysaccharides from Calocybe indica var. APK2. Exp. Toxicol. Pathol. 2016, 68, 329–334. [Google Scholar] [CrossRef]
- Yao, D.C.; Shi, W.B.; Gou, Y.L.; Zhou, X.R.; Yee Aw, T.; Zhou, Y.K.; Liu, Z. Fatty acid-mediated intracellular iron translocation: A synergistic mechanism of oxidative injury. Free Radic. Biol. Med. 2005, 39, 1385–1398. [Google Scholar] [CrossRef]
- Lu, Y.; Ye, M.; Song, S.; Li, L.; Shaikh, F.; Li, J.H. Isolation, purification, and anti-aging activity of melanin from Lachnum singerianum. Appl. Biochem. Biotechnol. 2014, 174, 762–771. [Google Scholar] [CrossRef] [PubMed]
- Young, S.I. Measurement of total antioxidant capacity. J. Clin. Pathol. 2001, 54, 339. [Google Scholar] [CrossRef] [PubMed]
- Zlokovic, B.V. Neurovascular pathways to neurodegeneration in Alzheimer’s disease and other disorders. Nat. Rev. Neurosci. 2011, 12, 723–738. [Google Scholar] [CrossRef] [PubMed]
- Ji, S.L.; Li, S.S.; Zhao, X.X.; Kang, N.X.; Cao, K.K.; Zhu, Y.Y.; Liu, Y. Protective role of phenylethanoid glycosides, Torenoside B and Savatiside A, in Alzheimer’s disease. Exp. Ther. Med. 2019, 17, 3755–3767. [Google Scholar] [CrossRef] [PubMed]
- Liu, J.; Wen, X.Y.; Zhang, X.Q.; Pu, H.M.; Kan, J.; Jin, C.H. Extraction, characterization and in vitro antioxidant activity of polysaccharides from black soybean. Int. J. Biol. Macromol. 2015, 72, 1182–1190. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Y.; Gu, M.; Wang, K.P.; Chen, Z.X.; Dai, L.Q.; Liu, J.Y.; Fang, Z. Structure, chain conformation and antitumor activity of a novel polysaccharide from Lentinus edodes. Fitoterapia 2010, 81, 1163–1170. [Google Scholar]
- Li, S.S.; Li, J.; Zhang, J.J.; Wang, W.S.; Wang, X.X.; Jing, H.J.; Jia, L. The Antioxidative, Antiaging, and Hepatoprotective Effects of Alkali-Extractable Polysaccharides by Agaricus bisporus. Evid. Based Complement. Altern. Med. 2017, 2017, 1–12. [Google Scholar] [CrossRef] [PubMed]
- Jia, X.; Wang, C.; Bai, Y.; Yu, J.; Xu, C. Sulfation of the Extracellular Polysaccharide Produced by the King Oyster Culinary-Medicinal Mushroom, Pleurotus eryngii (Agaricomycetes), and Its Antioxidant Properties In Vitro. Int. J. Med. Mushrooms 2017, 19, 355–362. [Google Scholar] [CrossRef]
- Miao, S.S.; Mao, X.H.; Pei, R.; Miao, S.P.; Xiang, C.; Lv, Y.J.; Liu, Y. Antitumor activity of polysaccharides from Lepista sordida against laryngocarcinoma in vitro and in vivo. Int. J. Biol. Macromol. 2013, 60, 235–240. [Google Scholar] [CrossRef]
- Zhang, J.; Liu, Y.J.; Park, H.S.; Xia, Y.; Kim, G. Antitumor activity of sulfated extracellular polysaccharides of Ganoderma lucidum from the submerged fermentation broth. Carbohydr. Polym. 2012, 87, 1539–1544. [Google Scholar] [CrossRef]
- Xu, Y.F.; Song, S.; Wei, Y.X.; Wang, F.X.; Zhao, M.; Guo, J.; Zhang, J. Sulfated modification of the polysaccharide from Sphallerocarpus gracilis and its antioxidant activities. Int. J. Biol. Macromol. 2016, 87, 180–190. [Google Scholar] [CrossRef] [PubMed]
- Sun, X.M.; Wang, H.H.; Han, X.F.; Chen, S.W.; Zhu, S.; Dai, J. Fingerprint analysis of polysaccharides from different Ganoderma by HPLC combined with chemometrics methods. Carbohydr. Polym. 2014, 114, 432–439. [Google Scholar] [CrossRef] [PubMed]
- Smirnoff, N.; Cumbes, Q.J. Hydroxyl radical scavenging activity of compatible solutes. Phytochemistry 1989, 28, 1057–1060. [Google Scholar] [CrossRef]
- Cumby, N.; Zhong, Y.; Naczk, M.; Shahidi, F. Antioxidant activity and water-holding capacity of canola protein hydrolysates. Food Chem. 2008, 109, 144–148. [Google Scholar] [CrossRef] [PubMed]
- Zhang, C.; Li, S.S.; Zhang, J.J.; Hu, C.L.; Che, G.; Zhou, M.; Jia, L. Antioxidant and hepatoprotective activities of intracellular polysaccharide from Pleurotus eryngii SI-04. Int. J. Biol. Macromol. 2016, 91, 568–577. [Google Scholar] [CrossRef] [PubMed]
- Kong, S.Z.; Li, J.C.; Li, S.D.; Liao, M.N.; Li, C.P.; Zheng, P.J.; Guo, M.H.; Tan, W.X.; Zheng, Z.H.; Hu, Z. Anti-Aging Effect of Chitosan Oligosaccharide on d-Galactose-Induced Subacute Aging in Mice. Mar. Drugs 2018, 16, 181. [Google Scholar] [CrossRef] [PubMed]
Sample Availability: Samples of the compounds are available from the authors. |










{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Groups | Body Weight (g) | Organ Index (mg/g) | ||
|---|---|---|---|---|
| Liver Index | kidney Index | Brain Index | ||
| NC | 45.91 ± 2.46 | 5.99 ± 0.46 | 4.61 ± 0.43 | 1.25 ± 0.08 |
| MC | 37.86 ± 2.28 a | 4.53 ± 0.41 a | 4.09 ± 0.35 a | 1.02 ± 0.05 a |
| PC | 42.78 ± 3.02 b | 5.38 ± 0.38 b | 4.58 ± 0.42 b | 1.21 ± 0.04 b |
| FPS | ||||
| 200 mg/kg/d | 38.78 ± 2.32 c | 4.70 ± 0.39 c | 4.12 ± 0.36 c | 1.08 ± 0.04 c |
| 400 mg/kg/d | 40.94 ± 2.31 c | 5.07 ± 0.45 c | 4.36 ± 0.40 b | 1.15 ± 0.05 c |
| SFPS | ||||
| 200 mg/kg/d | 39.86 ± 2.30 c | 4.84 ± 0.36 c | 4.26 ± 0.34 c | 1.11 ± 0.04 c |
| 400 mg/kg/d | 42.55 ± 3.05 b | 5.26 ± 0.37 b | 4.52 ± 0.44 b | 1.19 ± 0.07 b |
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Yuan, F.; Gao, Z.; Liu, W.; Li, H.; Zhang, Y.; Feng, Y.; Song, X.; Wang, W.; Zhang, J.; Huang, C.; et al. Characterization, Antioxidant, Anti-Aging and Organ Protective Effects of Sulfated Polysaccharides from Flammulina velutipes. Molecules 2019, 24, 3517. https://doi.org/10.3390/molecules24193517
Yuan F, Gao Z, Liu W, Li H, Zhang Y, Feng Y, Song X, Wang W, Zhang J, Huang C, et al. Characterization, Antioxidant, Anti-Aging and Organ Protective Effects of Sulfated Polysaccharides from Flammulina velutipes. Molecules. 2019; 24(19):3517. https://doi.org/10.3390/molecules24193517
Chicago/Turabian StyleYuan, Fangfang, Zheng Gao, Wenbo Liu, Huaping Li, Yiwen Zhang, Yanbo Feng, Xinling Song, Wenshuai Wang, Jianjun Zhang, Chunyan Huang, and et al. 2019. "Characterization, Antioxidant, Anti-Aging and Organ Protective Effects of Sulfated Polysaccharides from Flammulina velutipes" Molecules 24, no. 19: 3517. https://doi.org/10.3390/molecules24193517