Root Bark of Paeonia suffruticosa Extract and Its Component Methyl Gallate Possess Peroxynitrite Scavenging Activity and Anti-Inflammatory Properties through NF-κB Inhibition in LPS-treated Mice


Abstract
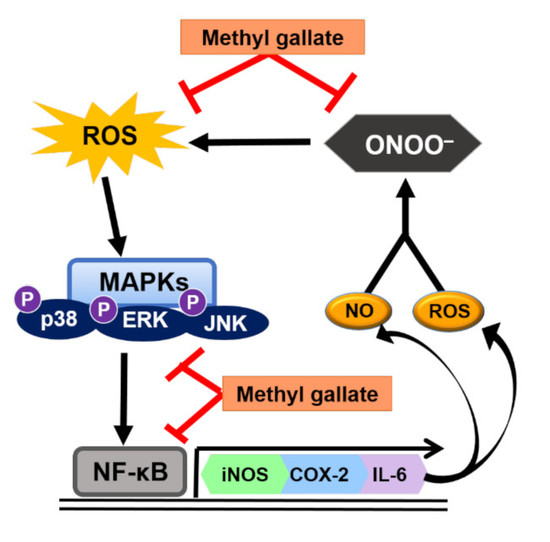
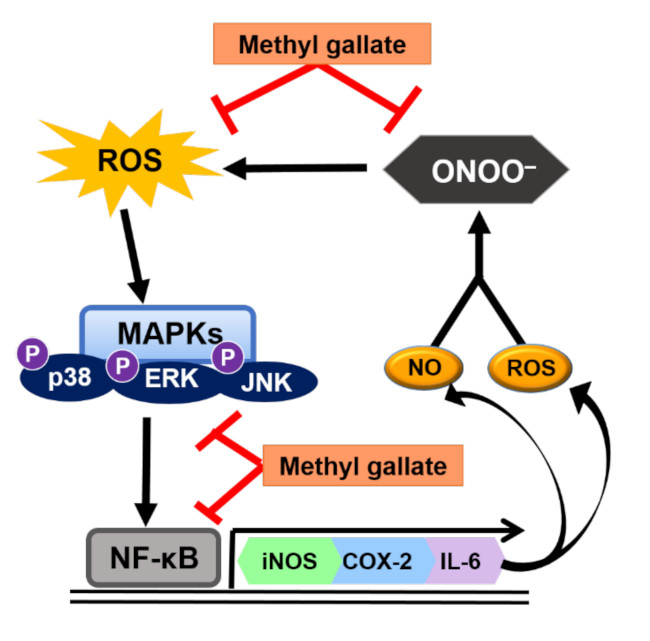
:
1. Introduction
2. Results
2.1. ONOO− Scavenging Activity of 70% EtOH Extracts from Korean Medicinal Plants
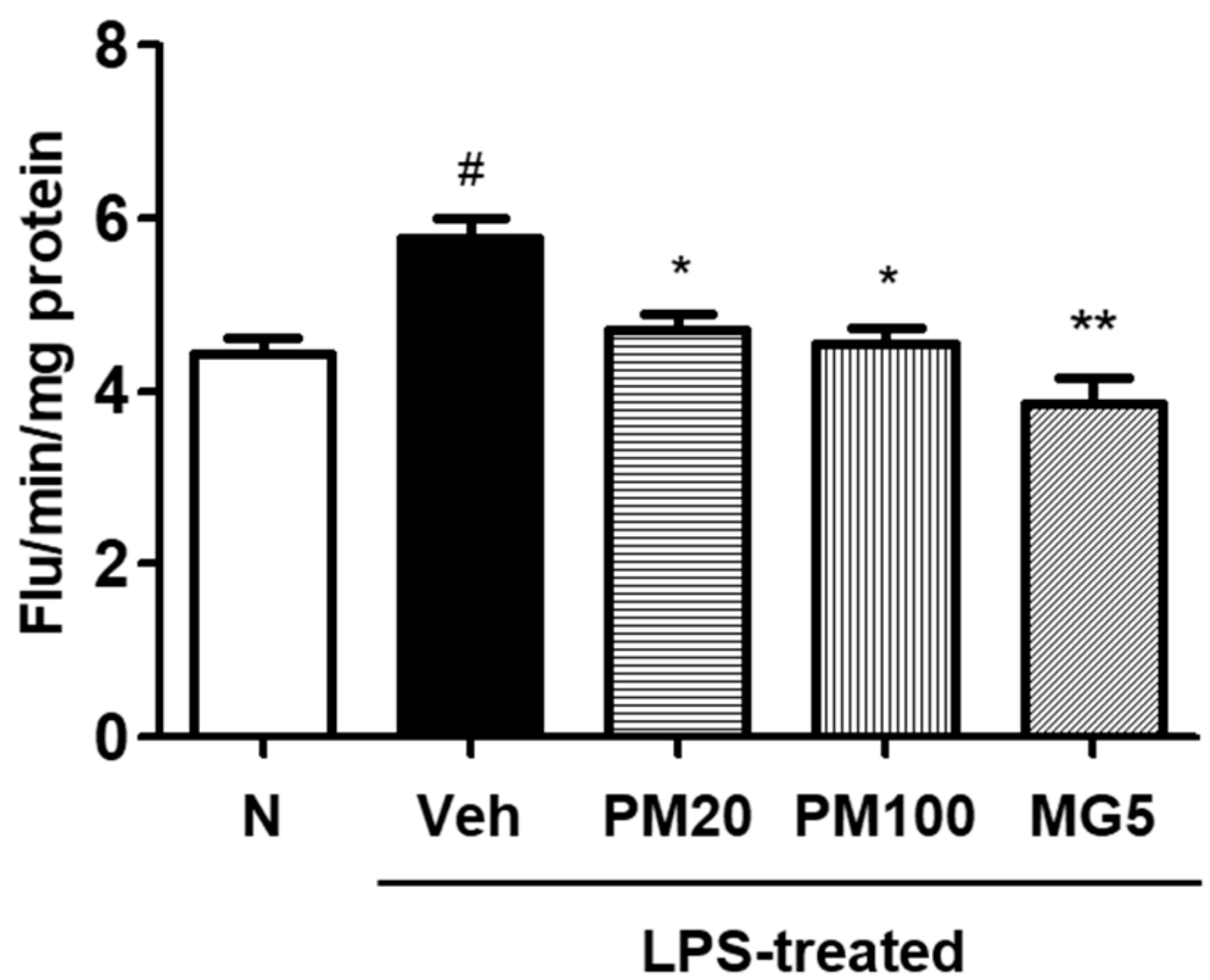
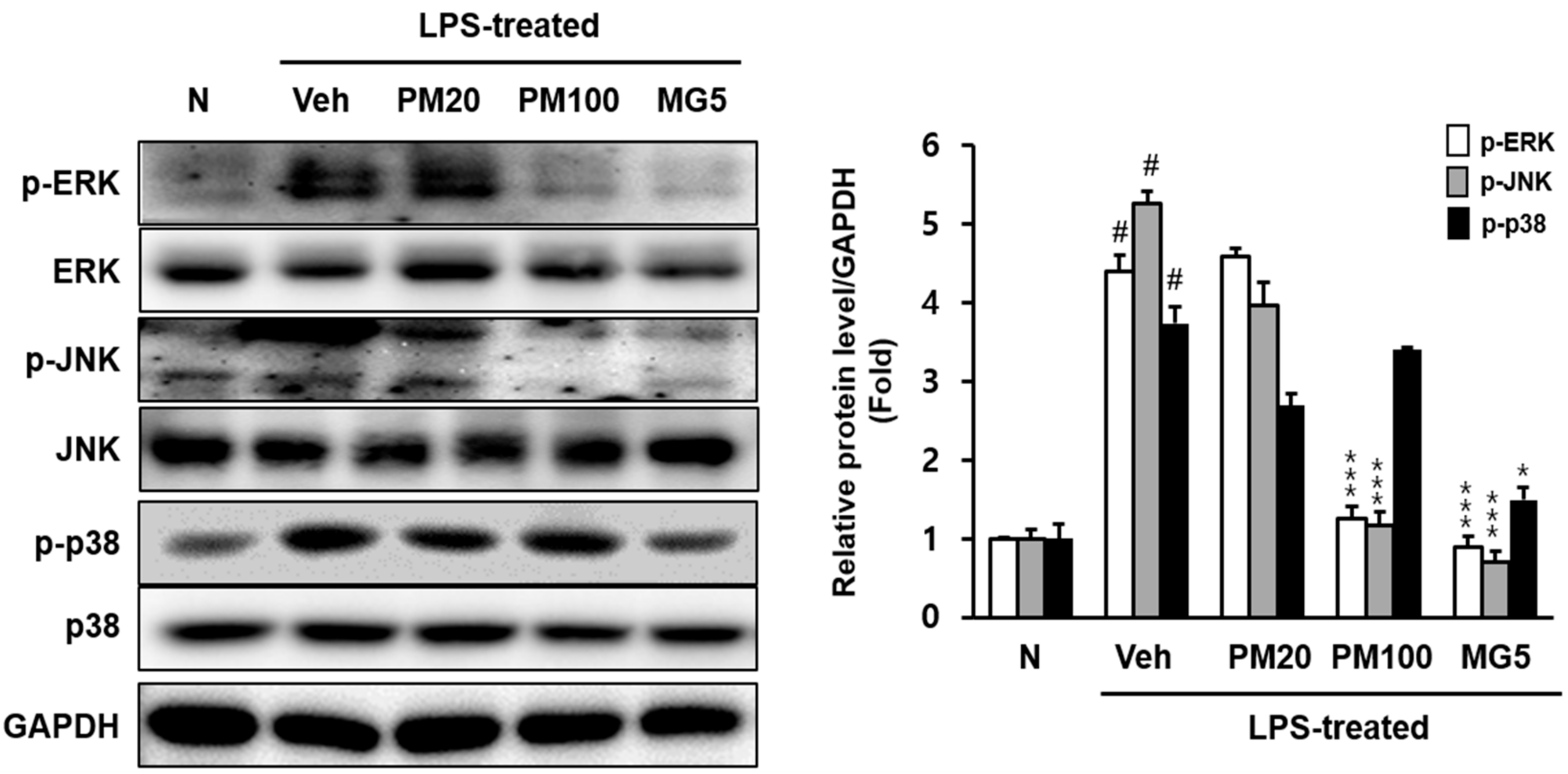
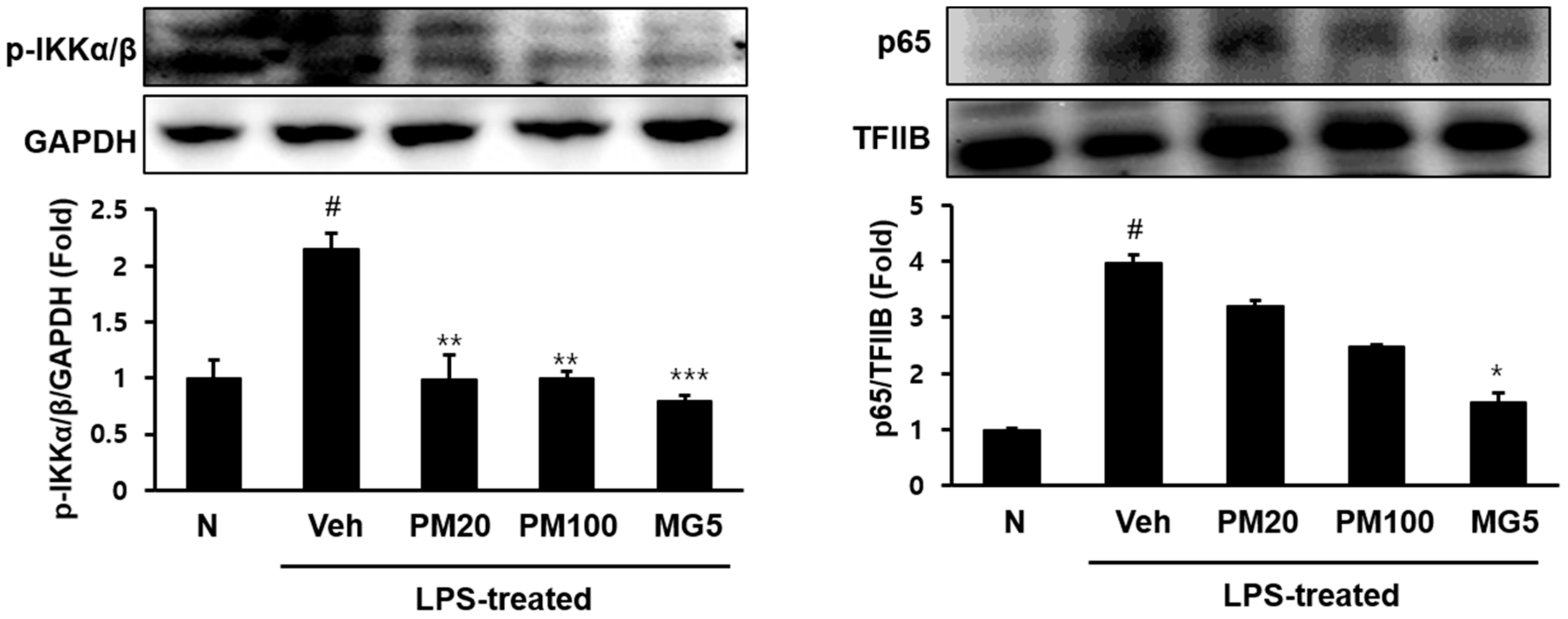
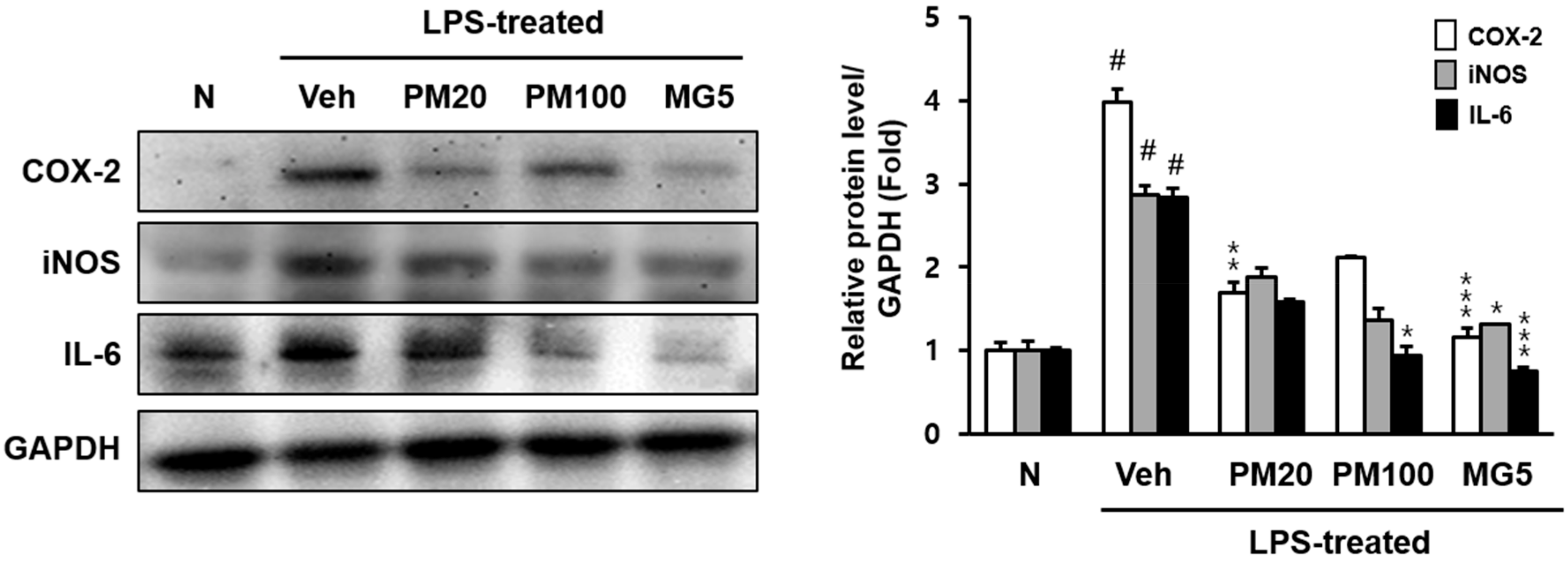
2.2. Effect of Root Bark of P. suffruticosa Extracts and Methyl Gallate on LPS-Induced Kidney Injury in Mice
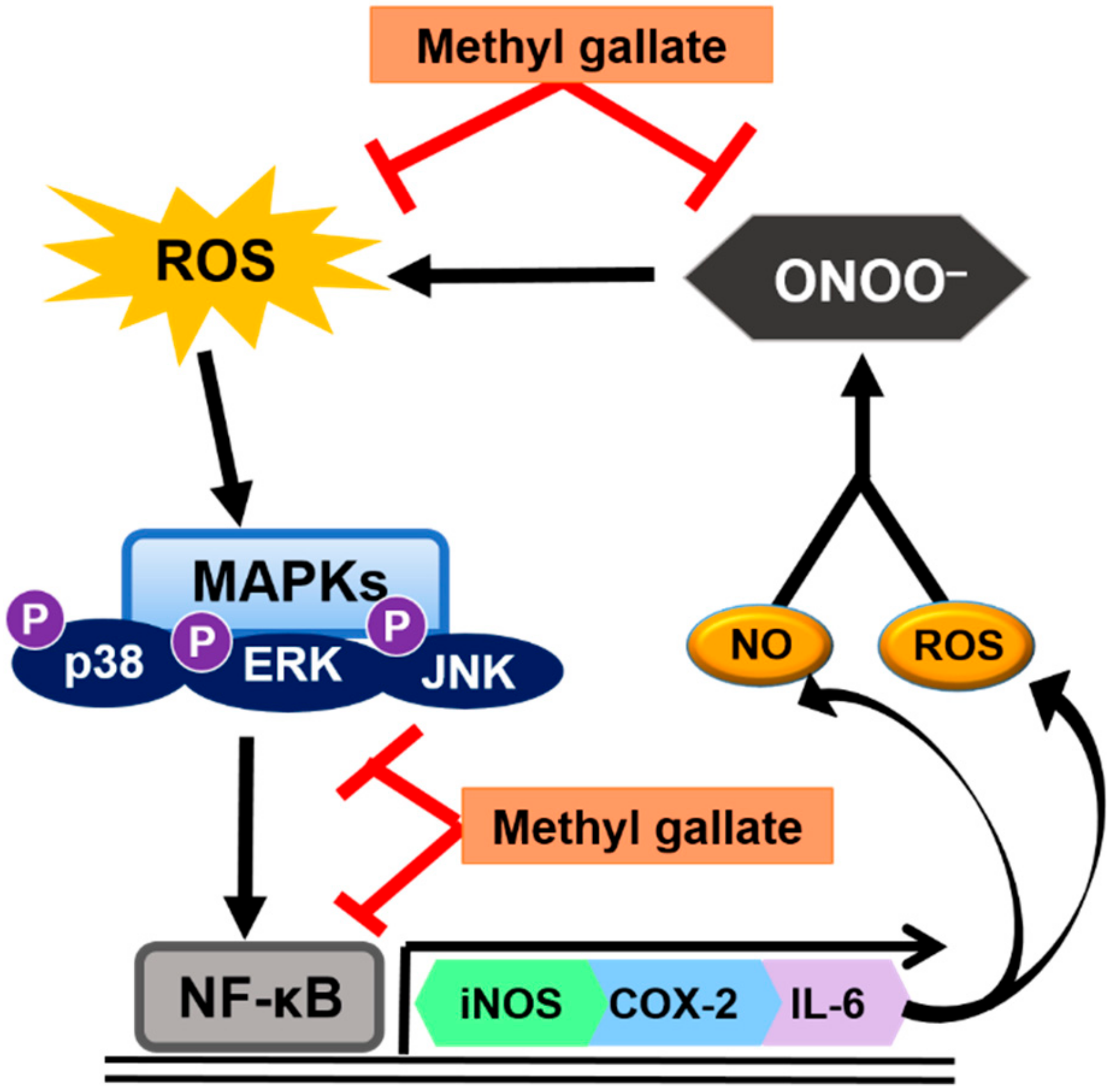
3. Discussion
4. Materials and Methods
4.1. Materials
4.2. Korean Medicinal Plants
4.3. Preparation of Extracts
4.4. Fractionation of the 70% EtOH Extract of Root Bark of P. suffruticosa
4.5. HPLC Analysis of the EtOAc Fraction of Root Bark of P. suffruticosa
4.6. Assay of ONOO− Levels
4.7. Experimental Animals and Treatment
4.8. Measurement of ROS Levels in the Kidney
4.9. Preparation of Cytosol and Nuclear Fractions
4.10. Western Blotting
4.11. Statistical Analyses
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Bashan, N.; Kovsan, J.; Kachko, I.; Ovadia, H.; Rudich, A. Positive and negative regulation of insulin signaling by reactive oxygen and nitrogen species. Physiol. Rev. 2009, 89, 27–71. [Google Scholar] [CrossRef] [PubMed]
- Beckman, J.S.; Koppenol, W.H. Nitric oxide, superoxide, and peroxynitrite: The good, the bad, and ugly. Am. J. Physiol. 1996, 271, C1424–C1437. [Google Scholar] [CrossRef] [PubMed]
- Mariotto, S.; Suzuki, Y.; Persichini, T.; Colasanti, M.; Suzuki, H.; Cantoni, O. Cross-talk between NO and arachidonic acid in inflammation. Curr. Med. Chem. 2007, 14, 1940–1944. [Google Scholar] [CrossRef] [PubMed]
- Cutler, R.G. Antioxidants and aging. Am. J. Clin. Nutr. 1991, 53, 373S–379S. [Google Scholar] [CrossRef] [PubMed]
- Meydani, M.; Lipman, R.D.; Han, S.N.; Wu, D.; Beharka, A.; Martin, K.R.; Bronson, R.; Cao, G.; Smith, D.; Meydani, S.N. The effect of long-term dietary supplementation with antioxidants. Ann. N. Y. Acad. Sci. 1998, 854, 352–360. [Google Scholar] [CrossRef] [PubMed]
- McCord, J.M.; Fridovich, I. The reduction of cytochrome c by milk xanthine oxidase. J. Biol. Chem. 1968, 243, 5753–5760. [Google Scholar] [PubMed]
- Nishikimi, M.; Appaji, N.; Yagi, K. The occurrence of superoxide anion in the reaction of reduced phenazine methosulfate and molecular oxygen. Biochem. Biophys. Res. Commun. 1972, 46, 849–854. [Google Scholar] [CrossRef]
- Bindoli, A.; Valente, M.; Cavallini, L. Inhibitory action of quercetin on xanthine oxidase and xanthine dehydrogenase activity. Pharmacol. Res. Commun. 1985, 17, 831–839. [Google Scholar] [CrossRef]
- Yokozawa, T.; Fujitsuka, N.; Oura, H.; Mori, A.; Kashiwagi, H. Determination of radical species in the kidney of rats with chronic renal failure by the spin trapping method. Nephron 1995, 70, 382–384. [Google Scholar] [CrossRef]
- Wu, H.K.; Sheu, S.J. Capillary electrophoretic determination of the constituents of paeoniae radix. J. Chromatogr. A 1996, 753, 139–146. [Google Scholar] [CrossRef]
- Lee, S.C.; Kwon, Y.S.; Son, K.H.; Kim, H.P.; Heo, M.Y. Antioxidative constituents from Paeonia lactiflora. Arch. Pharm. Res. 2005, 28, 775–783. [Google Scholar] [CrossRef] [PubMed]
- Hirokawa, S.; Nose, M.; Ishige, A.; Amagaya, S.; Oyama, T.; Ogihara, Y. Effect of Hachimi-jio-gan on scopolamine-induced memory impairment and on acetylcholine content in rat brain. J. Ethnopharmacol. 1996, 50, 77–84. [Google Scholar] [CrossRef]
- Okubo, T.; Nagai, F.; Seto, T.; Satoh, K.; Ushiyama, K.; Kano, I. The inhibition of phenylhydroquinone-induced oxidative DNA cleavage by constituents of Moutan Cortex and Paeoniae Radix. Biol. Pharm. Bull. 2000, 23, 199–203. [Google Scholar] [CrossRef] [PubMed]
- Qi, X.G. Protective mechanism of Salvia miltiorrhiza and Paeonia lactiflora for experimental liver damage. Zhong Xi Yi Jie He Za Zhi 1991, 11, 102–104. [Google Scholar] [PubMed]
- Zhang, Y. The effects of nifedipine, diltiazem, and Paeonia lactiflora Pall. on atherogenesis in rabbits. Zhonghua Xin Xue Guan Bing Za Zhi 1991, 19, 100–103. [Google Scholar] [PubMed]
- Sakai, Y.; Nagase, H.; Ose, Y.; Kito, H.; Sato, T.; Kawai, M.; Mizuno, M. Inhibitory action of peony root extract on the mutagenicity of benzo[a]pyrene. Mutat. Res. 1990, 244, 129–134. [Google Scholar] [CrossRef]
- Lin, H.C.; Ding, H.Y.; Ko, F.N.; Teng, C.M.; Wu, Y.C. Aggregation inhibitory activity of minor acetophenones from Paeonia species. Planta Med. 1999, 65, 595–599. [Google Scholar] [CrossRef]
- Lin, C.H.; Yeh, C.H.; Lin, L.J.; Wang, S.D.; Wang, J.S.; Kao, S.T. Immunomodulatory effect of Chinese herbal medicine formula sheng-fei-yu-chuan-tang in lipopolysaccharide-induced acute lung injury mice. Evid. Based Complement. Alternat. Med. 2013, 2013, 976342. [Google Scholar] [CrossRef]
- Moncada, S.; Palmer, R.M.; Higgs, E.A. Nitric oxide: Physiology, pathophysiology, and pharmacology. Pharmacol. Rev. 1991, 43, 109–142. [Google Scholar]
- Thiemermann, C.; Ruetten, H.; Wu, C.C.; Vane, J.R. The multiple organ dysfunction syndrome caused by endotoxin in the rat: Attenuation of liver dysfunction by inhibitors of nitric oxide synthase. Br. J. Pharmacol. 1995, 116, 2845–2851. [Google Scholar] [CrossRef]
- Boveris, A. Biochemistry of free radicals: From electrons to tissues. Medicina, B. Aires 1998, 58, 350–356. [Google Scholar]
- Leach, M.; Frank, S.; Olbrich, A.; Pfeilschifter, J.; Thiemermann, C. Decline in the expression of copper/zinc superoxide dismutase in the kidney of rats with endotoxic shock: Effects of the superoxide anion radical scavenger, tempol, on organ injury. Br. J. Pharmacol. 1998, 125, 817–825. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tsai, C.L.; Lin, Y.C.; Wang, H.M.; Chou, T.C. Baicalein, an active component of Scutellaria baicalensis, protects against lipopolysaccharide-induced acute lung injury in rats. J. Ethnopharmacol. 2014, 153, 197–206. [Google Scholar] [CrossRef] [PubMed]
- Park, J.H.; Kim, K.H.; Lee, W.R.; Han, S.M.; Park, K.K. Protective effect of melittin on inflammation and apoptosis in acute liver failure. Apoptosis 2012, 17, 61–69. [Google Scholar] [CrossRef] [PubMed]
- Manna, S.K.; Mukhopadhyay, A.; Aggarwal, B.B. Resveratrol suppresses TNF-induced activation of nuclear transcription factors NF-κB, activator protein-1, and apoptosis: Potential role of reactive oxygen intermediates and lipid peroxidation. J. Immunol. 2000, 164, 6509–6519. [Google Scholar] [CrossRef] [PubMed]
- Lawrence, T.; Fong, C. The resolution of inflammation: Anti-inflammatory roles of NF-κB. Int. J. Biochem. Cell Biol. 2010, 42, 519–523. [Google Scholar] [CrossRef]
- Speyer, C.L.; Neff, T.A.; Warner, R.L.; Guo, R.F.; Sarma, J.V.; Riedemann, N.C.; Murphy, M.E.; Murphy, H.S.; Ward, P.A. Regulatory effects of iNOS on acute lung inflammatory responses in mice. Am. J. Pathol. 2003, 163, 2319–2328. [Google Scholar] [CrossRef]
- Cho, E.J.; Yokozawa, T.; Kim, H.Y.; Shibahara, N.; Park, J.C. Rosa rugose attenuates diabetic oxidative stress in rats with streptozotocin-induced diabetes. Am. J. Chin. Med. 2004, 32, 487–496. [Google Scholar] [CrossRef]
- Kang, M.S.; Jang, H.S.; Oh, J.S.; Yang, K.H.; Choi, N.K.; Lim, H.S.; Kim, S.M. Effects of methyl gallate and gallic acid on the production of inflammatory mediators interleukin-6 and interleukin-8 by oral epithelial cells stimulated with Fusobacterium nucleatum. J. Microbiol. 2009, 47, 760–767. [Google Scholar] [CrossRef]
- Rosas, E.C.; Correa, L.B.; Pádua Tde, A.; Costa, T.E.; Mazzei, J.L.; Heringer, A.P.; Bizarro, C.A.; Kaplan, M.A.; Figueiredo, M.R.; Henriques, M.G. Anti-inflammatory effect of Schinus terebinthifolius Raddi hydroalcoholic extract on neutrophil migration in zymosan-induced arthritis. J. Ethnopharmacol. 2015, 175, 490–498. [Google Scholar] [CrossRef]
- Whang, W.K.; Park, H.S.; Ham, I.H.; Oh, M.; Namkoong, H.; Kim, H.K.; Hwang, D.W.; Hur, S.Y.; Kim, T.E.; Park, Y.G.; et al. Methyl gallate and chemicals structurally related to methyl gallate protect human umbilical vein endothelial cells from oxidative stress. Exp. Mol. Med. 2005, 37, 343–352. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lee, S.H.; Kim, J.K.; Kim, D.W.; Hwang, H.S.; Eum, W.S.; Park, J.; Han, K.H.; Oh, J.S.; Choi, S.Y. Antitumor activity of methyl gallate by inhibition of focal adhesion formation and Akt phosphorylation in glioma cells. Biochim. Biophys. Acta 2013, 1830, 4017–4029. [Google Scholar] [CrossRef] [PubMed]
- Acharyya, S.; Sarkar, P.; Saha, D.R.; Patra, A.; Ramamurthy, T.; Bag, P.K. Intracellular and membrane-damaging activities of methyl gallate isolated from Terminalia chebula against multidrug-resistant Shigella spp. J. Med. Microbiol. 2015, 64, 901–909. [Google Scholar] [CrossRef] [PubMed]
- Cho, E.J.; Yokozawa, T.; Rhyu, D.Y.; Kim, S.C.; Shibahara, N.; Park, J.C. Study on the inhibitory effects of Korean medicinal plants and their main compounds on the 1,1-diphenyl-2-picrylhydrazyl radical. Phytomedicine 2003, 10, 544–551. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hsieh, T.J.; Liu, T.Z.; Chia, Y.C.; Chern, C.L.; Lu, F.J.; Chuang, M.C.; Mau, S.Y.; Chen, S.H.; Syu, Y.H.; Chen, C.H. Protective effect of methyl gallate from Toona sinensis (Meliaceae) against hydrogen peroxide-induced oxidative stress and DNA damage in MDCK cells. Food Chem. Toxicol. 2004, 42, 843–850. [Google Scholar] [CrossRef]
- Kooy, N.W.; Royall, J.A.; Ischiropoulos, H.; Beckman, J.S. Peroxynitrite-mediated oxidation of dihydrorhodamine 123. Free Radic. Biol. Med. 1994, 16, 149–156. [Google Scholar] [CrossRef]
- Ali, S.F.; LeBel, C.P.; Bondy, S.C. Reactive oxygen species formation as a biomarker of methylmercury and trimethyltin neurotoxicity. Neurotoxicology 1992, 13, 637–648. [Google Scholar]
- Deng, L.; Lin-Lee, Y.C.; Claret, F.X.; Kuo, M.T. 2-Acetylaminofluorene up-regulates rat mdr1b expression through generating reactive oxygen species that activate NF-κB pathway. J. Biol. Chem. 2001, 276, 413–420. [Google Scholar] [CrossRef]
- Kim, J.M.; Lee, E.K.; Kim, D.H.; Yu, B.P.; Chung, H.Y. Kaempferol modulates pro-inflammatory NF-kappaB activation by suppressing advanced glycation endproducts-induced NADPH oxidase. Age 2010, 32, 197–208. [Google Scholar] [CrossRef]
Sample Availability: Samples of the compounds are available from the authors. |








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Herb Name | Family | Used Part |
|---|---|---|
| Lonicera japonica Thunb. | Caprifoliaceae | Whole |
| Lysimachia christinae Hance | Primulaceae | Whole |
| Scirpus yagara Ohwi | Cyperaceae | Root |
| Paeonia suffruticosa Andrew | Paeoniaceae | Root bark |
| Ailanthus altissima (Mill.) Swingle | Simaroubaceae | Leaf |
| Pueraria thunbergiana Benth | Leguminosae/Fabaceae | Root |
| Curcuma zedoaria (Christm.) Roscoe | Zingiberaceae | Root |
| Extract | IC50 Value (µg/mL) a |
|---|---|
| Lonicera japonica Thunb. | 7.92 ± 0.72 |
| Lysimachia christinae Hance | 13.19 ± 0.28 |
| Scirpus yagara Ohwi | 11.74 ± 0.77 |
| Paeonia suffruticosa Andrew | 7.69 ± 0.67 |
| Ailanthus altissima (Mill.) Swingle | 12.64 ± 0.37 |
| Pueraria thunbergiana Benth | 10.65 ± 0.04 |
| Curcuma zedoaria (Christm.) Roscoe | 10.13 ± 0.14 |
| l-Penicillamine b | 8.24 ± 0.24 |
| Sample | IC50 Values (µg/mL) a |
|---|---|
| 70% EtOH extract | 4.78 ± 0.13 |
| CH2Cl2 fraction | 1.78 ± 0.05 |
| EtOAc fraction | 0.25 ± 0.03 |
| n-BuOH fraction | 4.19 ± 0.23 |
| H2O fraction | 7.99 ± 0.79 |
| l-Penicillamine b | 9.00 ± 0.38 |
| Compound | Retention Time (min) | Calibration Equation a | Correlation Coefficient (r2) |
|---|---|---|---|
| Methyl gallate | 15.451 | y = 17.430x + 4.221 | 0.996 |
| Sample | Content (mg/g) | RSD a (%) |
|---|---|---|
| 70% EtOH extract | 5.07 ± 0.06 | 2.22 |
| EtOAc fraction | 39.62 ± 1.04 | 4.53 |
| Sample | IC50 Values (µM) a |
|---|---|
| Methyl gallate | 0.91 ± 0.26 |
| l-Penicillamine b | 8.79 ± 0.17 |
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Park, D.J.; Jung, H.J.; Park, C.H.; Yokozawa, T.; Jeong, J.-C. Root Bark of Paeonia suffruticosa Extract and Its Component Methyl Gallate Possess Peroxynitrite Scavenging Activity and Anti-Inflammatory Properties through NF-κB Inhibition in LPS-treated Mice. Molecules 2019, 24, 3483. https://doi.org/10.3390/molecules24193483
Park DJ, Jung HJ, Park CH, Yokozawa T, Jeong J-C. Root Bark of Paeonia suffruticosa Extract and Its Component Methyl Gallate Possess Peroxynitrite Scavenging Activity and Anti-Inflammatory Properties through NF-κB Inhibition in LPS-treated Mice. Molecules. 2019; 24(19):3483. https://doi.org/10.3390/molecules24193483
Chicago/Turabian StylePark, Dong Jin, Hee Jin Jung, Chan Hum Park, Takako Yokozawa, and Ji-Cheon Jeong. 2019. "Root Bark of Paeonia suffruticosa Extract and Its Component Methyl Gallate Possess Peroxynitrite Scavenging Activity and Anti-Inflammatory Properties through NF-κB Inhibition in LPS-treated Mice" Molecules 24, no. 19: 3483. https://doi.org/10.3390/molecules24193483