Sphingosine-1-Phosphate Improves the Biological Features of Mouse Bone Marrow-Derived EPCs Partially through PI3K/AKT/eNOS/NO Pathway

 ,
, {kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Results
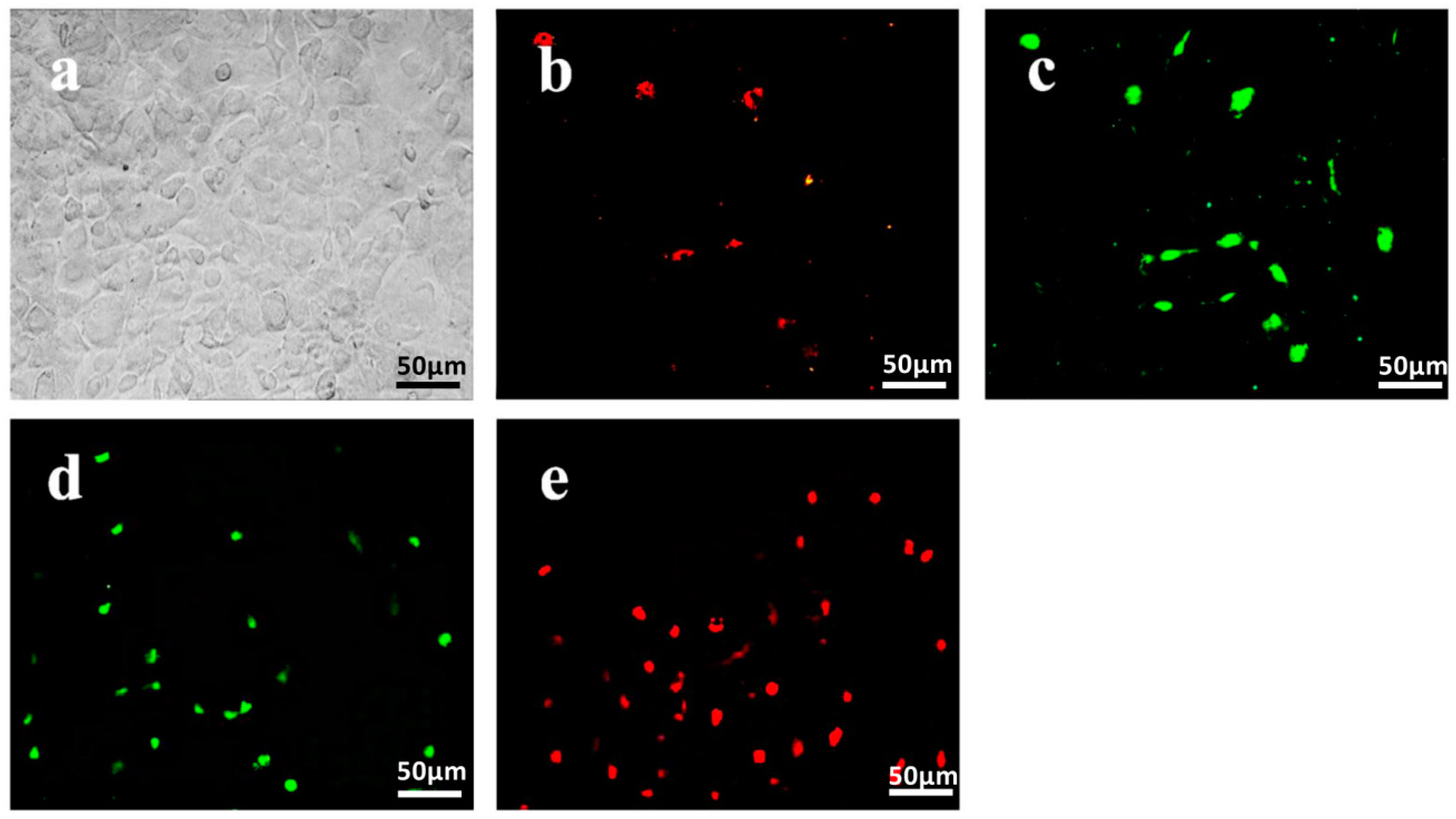
2.1. Isolation and Identification of Endothelial Progenitor Cells
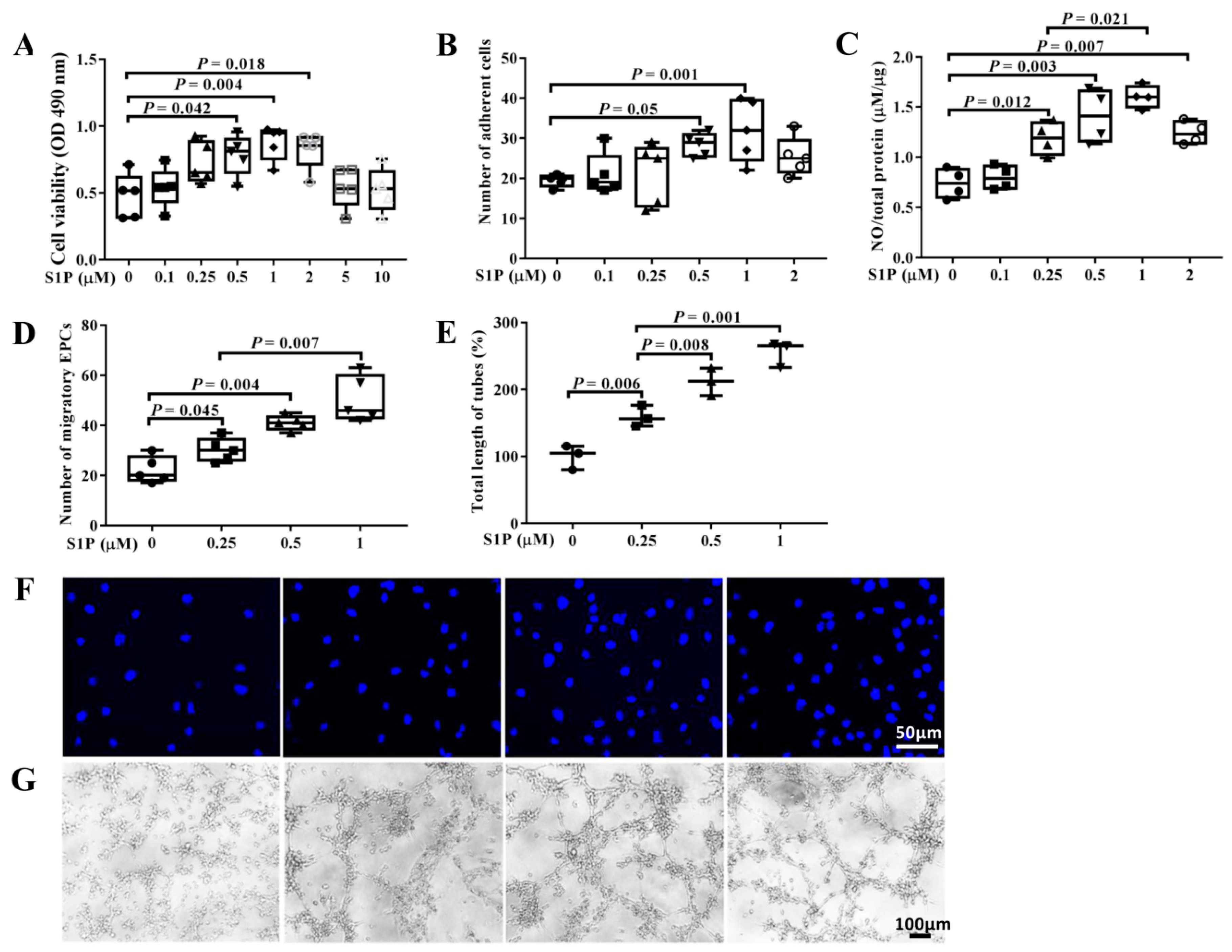
2.2. S1P Improves the Biological Features of EPCs
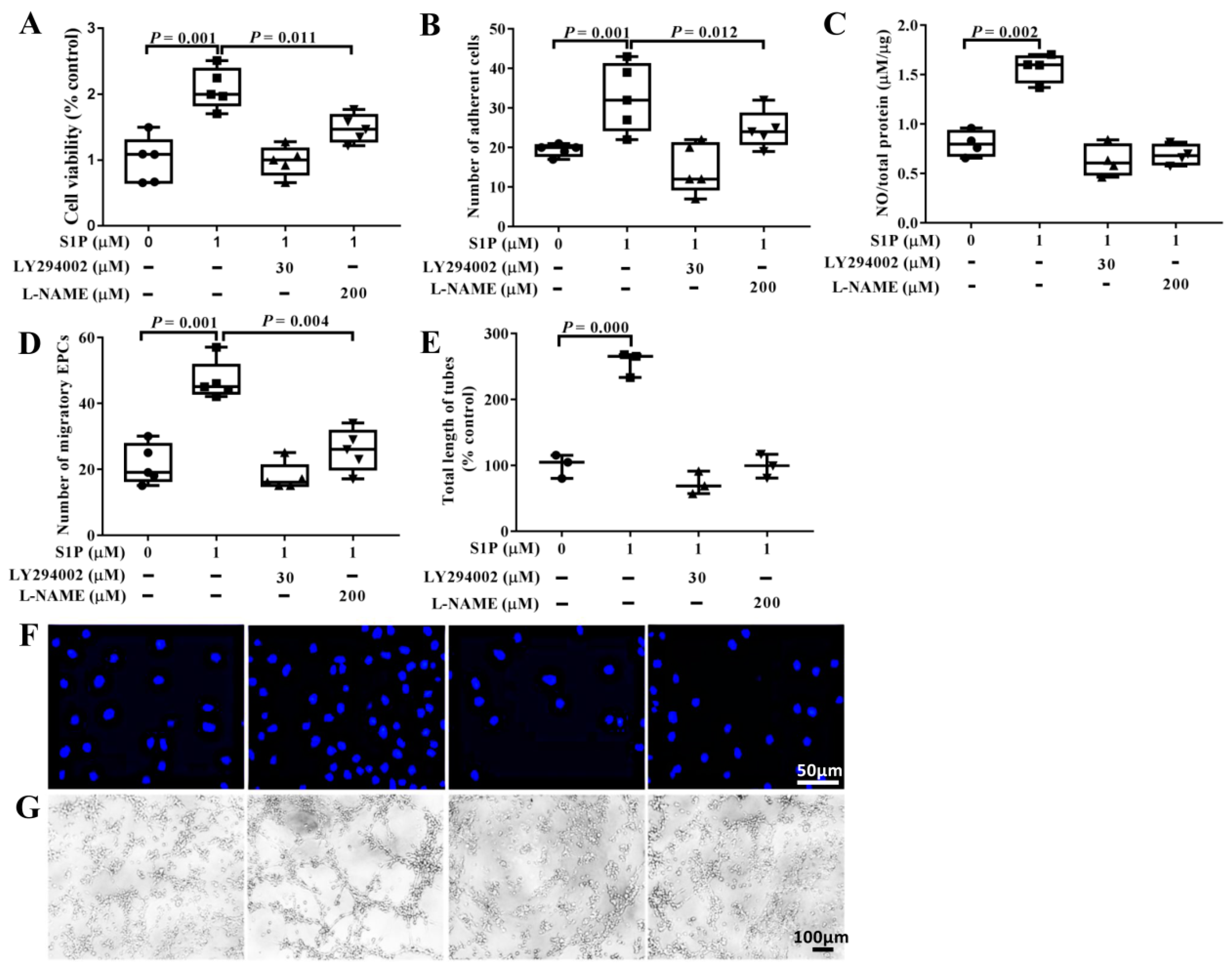
2.3. LY294002 and L-NAME Partialy Suppress S1P-Promoted Biological Features of EPCs
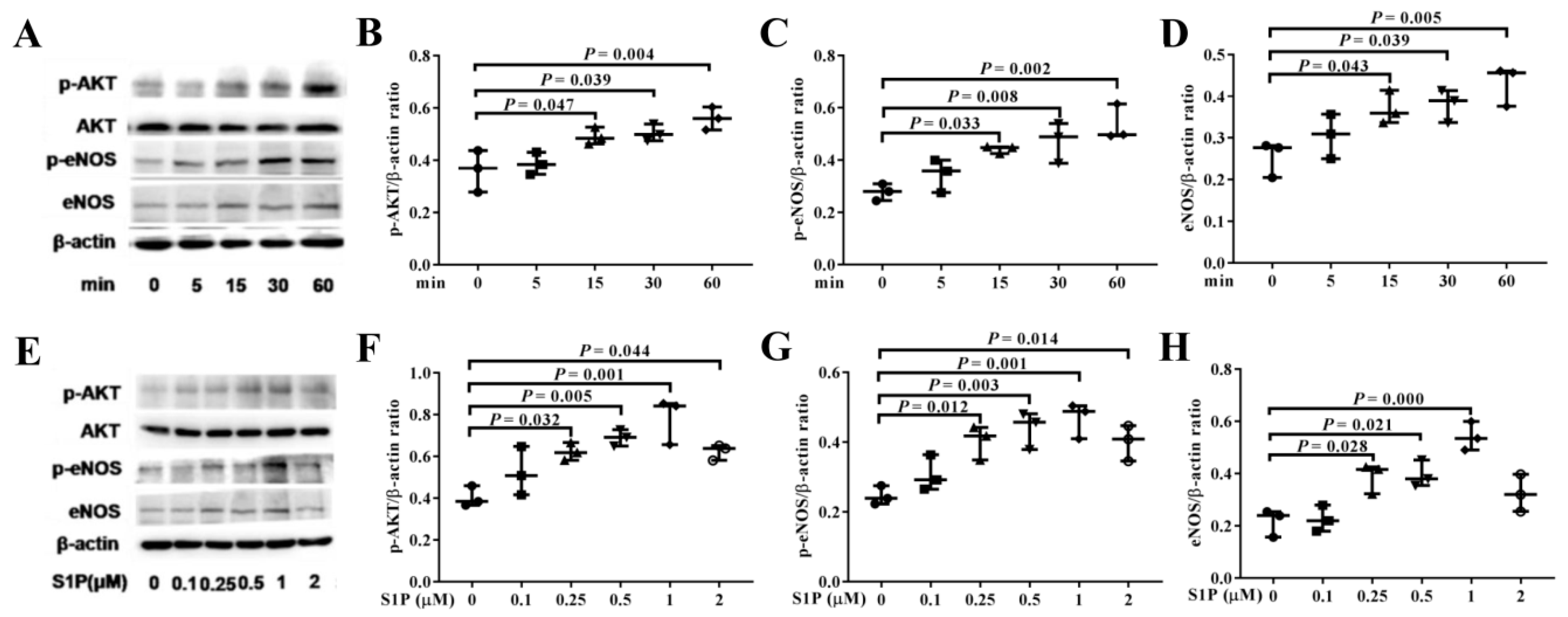
2.4. S1P Activates AKT and eNOS Phosphorylation
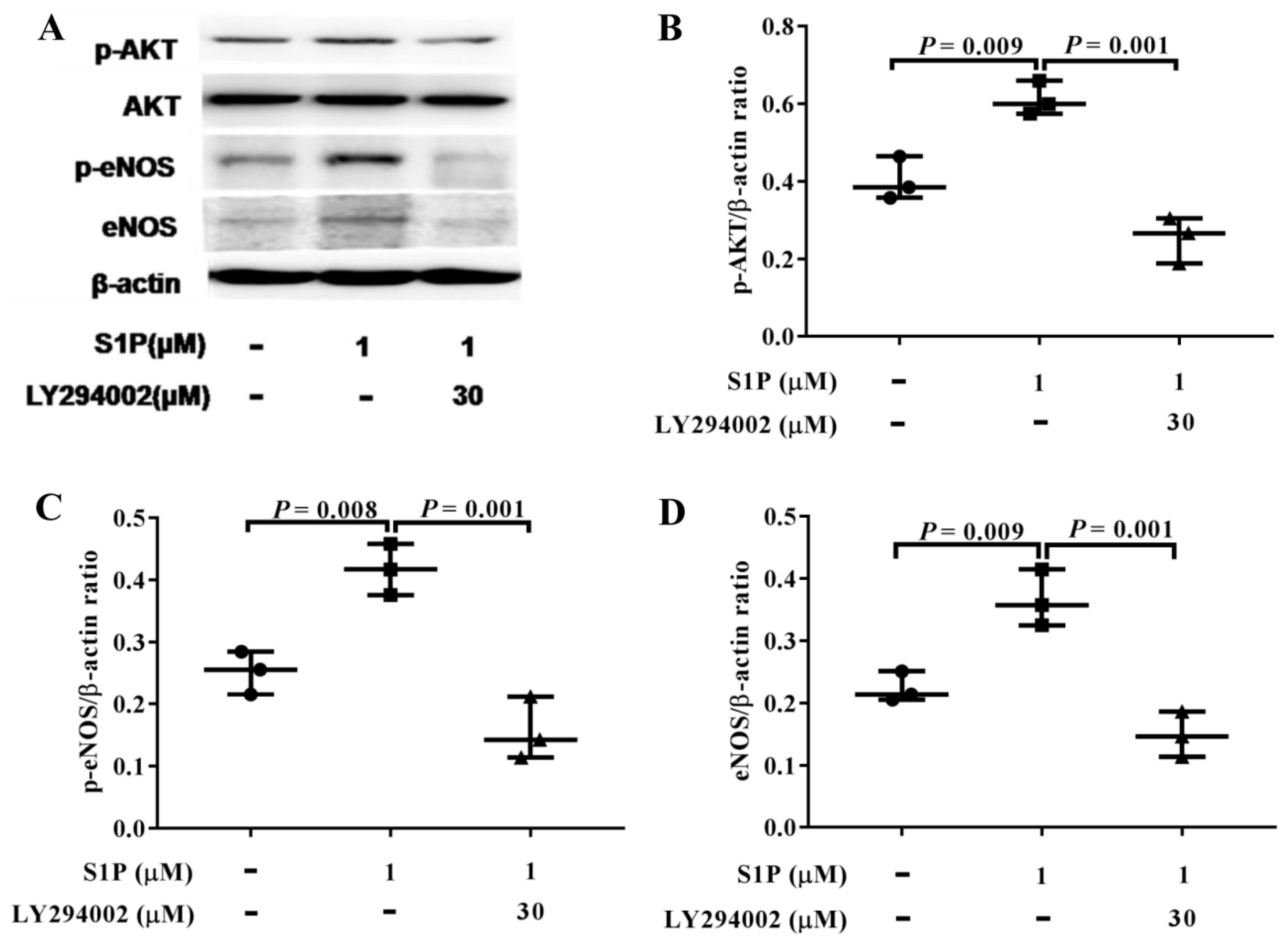
2.5. LY294002 Inhibits The Levels of p-AKT, p-eNOS, and eNOS Promoted by S1P
3. Discussion
4. Materials and Methods
4.1. Isolation and Culture of EPCs Derived from C57 Mice Bone Marrow
4.2. Immunocytochemistry
4.3. EPCs Treatment
4.4. Cell Viability of EPCs
4.5. EPCs Migration Assay
4.6. Cell Adhesion
4.7. Tube Formation Assay
4.8. Detection of NO in Medium
4.9. Western Blot Analyses
4.10. Statistical Analysis
Supplementary Materials
Author Contributions
Funding
Conflicts of Interest
References
- Schwartz, B.G.; Economides, C.; Mayeda, G.S.; Burstein, S.; Kloner, R.A. The endothelial cell in health and disease: Its function, dysfunction, measurement and therapy. Int. J. Impot. Res. 2010, 22, 77–90. [Google Scholar] [CrossRef] [PubMed]
- Xu, J.Y.; Lee, Y.K.; Wang, Y.; Tse, H.F. Therapeutic application of endothelial progenitor cells for treatment of cardiovascular diseases. Curr. Stem Cell Res. Ther. 2014, 9, 401–414. [Google Scholar] [CrossRef] [PubMed]
- Hill, J.M.; Zalos, G.; Halcox, J.P.; Schenke, W.H.; Waclawiw, M.A.; Quyyumi, A.A.; Finkel, T. Circulating endothelial progenitor cells, vascular function, and cardiovascular risk. N. Engl. J. Med. 2003, 348, 593–600. [Google Scholar] [CrossRef] [PubMed]
- Naito, H.; Kidoya, H.; Sakimoto, S.; Wakabayashi, T.; Takakura, N. Identification and characterization of a resident vascular stem/progenitor cell population in preexisting blood vessels. EMBO J. 2012, 31, 842–855. [Google Scholar] [CrossRef] [PubMed]
- Bitterli, L.; Afan, S.; Bühler, S.; DiSanto, S.; Zwahlen, M.; Schmidlin, K.; Yang, Z.; Baumgartner, I.; Diehm, N.; Kalka, C. Endothelial progenitor cells as a biological marker of peripheral artery disease. Vasc. Med. 2016, 21, 3–11. [Google Scholar] [CrossRef]
- Georgescu, A.; Alexandru, N.; Andrei, E.; Dragan, E.; Cochior, D.; Dias, S. Effects of transplanted circulating endothelial progenitor cells and platelet microparticles in atherosclerosis development. Biol. Cell 2016, 108, 219–243. [Google Scholar] [CrossRef]
- Gandy, K.A.; Obeid, L.M. Regulation of the sphingosine kinase/sphingosine 1-phosphate pathway. Handb. Exp. Pharmacol. 2013, 216, 275–303. [Google Scholar]
- Maceyka, M.; Spiegel, S. Sphingolipid metabolites in inflammatory disease. Nature 2014, 510, 58–67. [Google Scholar] [CrossRef] [Green Version]
- Zhao, J.; Garcia, D.; Gartung, A.; Lee, M.J. Sphingosine-1-phosphate receptor subtype 2 signaling in endothelial senescence-associated functional impairments and inflammation. Curr. Atheroscler. Rep. 2015, 17, 504. [Google Scholar] [CrossRef]
- Daum, G.; Grabski, A.; Reidy, M.A. Sphingosine1-phosphate: A regulator of arterial lesions. Thromb Vasc. Biol. 2009, 29, 1439–1443. [Google Scholar] [CrossRef]
- Seeger, F.H.; Haendeler, J.; Walter, D.H.; Rochwalsky, U.; Reinhold, J.; Urbich, C.; Rössig, L.; Corbaz, A.; Chvatchko, Y.; Zeiher, A.M.; et al. p38 mitogen-activated protein kinase downregulates endothelial progenitor cells. Circulation 2005, 111, 1184–1191. [Google Scholar] [CrossRef] [PubMed]
- Walter, D.H.; Haendeler, J.; Reinhold, J.; Rochwalsky, U.; Seeger, F.; Honold, J.; Hoffmann, J.; Urbich, C.; Lehmann, R.; Arenzana-Seisdesdos, F.; et al. Impaired CXCR4 signaling contributes to the reduced neovascularization capacity of endothelial progenitor cells from patients with coronary artery disease. Circ. Res. 2005, 97, 1142–1151. [Google Scholar] [CrossRef] [PubMed]
- Walter, D.H.; Rochwalsky, U.; Reinhold, J.; Seeger, F.; Aicher, A.; Urbich, C.; Spyridopoulos, I.; Chun, J.; Brinkmann, V.; Keul, P.; et al. Sphingosine-1-Phosphate Stimulates the Functional Capacity of Progenitor Cells by Activation of the CXCR4-Dependent Signaling Pathway via the S1P3 Receptor. Arterioscler. Thromb. Vasc. Biol. 2007, 27, 275–282. [Google Scholar] [CrossRef] [PubMed]
- Son, B.R.; Marquez-Curtis, L.A.; Kucia, M.; Wysoczynski, M.; Turner, A.R.; Ratajczak, J.; Ratajczak, M.Z.; Janowska-Wieczorek, A. Migration of bone marrow and cord blood mesenchymal stem cells in vitro is regulated by stromal-derived factor-1-CXCR4 and hepatocyte growth factor-c-met axes and involves matrix metalloproteinases. Stem Cells 2006, 24, 1254–1264. [Google Scholar] [CrossRef] [PubMed]
- Haynes, M.P.; Li, L.; Sinha, D.; Russell, K.S.; Hisamoto, K.; Baron, R.; Collinge, M.; Sessa, W.C.; Bender, J.R. Src kinase mediates phosphatidylinositol 3-kinase/Akt-dependent rapid endothelial nitric-oxide synthase activation by estrogen. J. Biol. Chem. 2003, 278, 2118–2123. [Google Scholar] [CrossRef] [PubMed]
- Chen, X.; Chen, Q.; Wang, L.; Li, G. Ghrelin induces cell migration through GHSR1a-mediated PI3K/Akt/eNOS/NO signaling pathway in endothelial progenitor cells. Metabolism 2013, 62, 743–752. [Google Scholar] [CrossRef] [PubMed]
- Yang, N.; Yao, S.; Wang, M.; Jiao, P.; Zhang, Y.; Qin, S. Apolipoprotein A-I mimetic peptide reverse D-4F improves the biological functions of mouse bone marrow-derived late EPCs via PI3K/AKT/eNOS pathway. Mol. Cell Biochem. 2013, 377, 229–236. [Google Scholar] [CrossRef]
- Kwon, S.M.; Lee, J.H.; Lee, S.H.; Jung, S.Y.; Kim, D.Y.; Kang, S.H.; Yoo, S.Y.; Hong, J.K.; Park, J.H.; Kim, J.H.; et al. Cross talk with hematopoietic cells regulates the endothelial progenitor cell differentiation of CD34 positive cells. PLoS ONE 2014, 9, e106310. [Google Scholar] [CrossRef]
- Asahara, T.; Murohara, T.; Sullivan, A.; Silver, M.; van der Zee, R.; Li, T.; Witzenbichler, B.; Schatteman, G.; Isner, J.M. Isolation of putative progenitor endothelial cells for angiogenesis. Science 1997, 275, 964–967. [Google Scholar] [CrossRef]
- Winkler, M.S.; Nierhaus, A.; Poppe, A.; Greiwe, G.; Gräler, M.H.; Daum, G. Sphingosine-1-Phosphate: A Potential Biomarker and Therapeutic Target for Endothelial Dysfunction and Sepsis? Shock 2017, 47, 666–672. [Google Scholar] [CrossRef]
- Yang, S.W.; Hennessy, R.R.; Khosla, S.; Lennon, R.; Loeffler, D.; Sun, T.; Liu, Z.; Park, K.H.; Wang, F.L.; Lerman, L.O.; et al. Circulating osteogenic endothelial progenitor cell counts: New biomarker for the severity of coronary artery disease. Int. J. Cardiol. 2017, 227, 833–839. [Google Scholar] [CrossRef] [PubMed]
- Asahara, T.; Kawamoto, A.; Masuda, H. Concise review: Circulating endothelial progenitor cells for vascular medicine. Stem Cells 2011, 29, 1650–1655. [Google Scholar] [CrossRef] [PubMed]
- Haider, K.H.; Aziz, S.; Al-Reshidi, M.A. Endothelial progenitor cells for cellular angiogenesis and repair: Lessons learned from experimental animal models. Regen. Med. 2017, 12, 969–982. [Google Scholar] [CrossRef] [PubMed]
- Watt., J.; Kennedy, S.; Ahmed, N.; Hayhurst, J.; McClure, J.D.; Berry, C.; Wadsworth, R.M.; Oldroyd, K.G. The relationship between oxidised LDL, endothelial progenitor cells and coronary endothelial function in patients with CHD. Open Heart 2016, 3, e000342. [Google Scholar] [CrossRef] [PubMed]
- Sattler, K.J.; Elbasan, S.; Keul, P.; Elter-Schulz, M.; Bode, C.; Gräler, M.H.; Bröcker-Preuss, M.; Budde, T.; Erbel, R.; Heusch, G.; et al. Sphingosine 1-phosphate levels in plasma and HDL are altered in coronary artery disease. Basic Res. Cardiol. 2010, 105, 821–832. [Google Scholar] [CrossRef] [PubMed]
- Vasa, M.; Fichtlscherer, S.; Aicher, A.; Adler, K.; Urbich, C.; Martin, H.; Zeiher, A.M.; Dimmeler, S. Number and migratory activity of circulating endothelial progenitor cells inversely correlate with risk factors for coronary artery disease. Circ Res. 2001, 89, e1–e7. [Google Scholar] [CrossRef] [PubMed]
- Tepper, O.M.; Galiano, R.D.; Capla, J.M.; Kalka, C.; Gagne, P.J.; Jacobowitz, G.R.; Levine, J.P.; Gurtner, G.C. Human endothelial progenitor cells from type II diabetics exhibit impaired proliferation, adhesion, and incorporation into vascular structures. Circulation 2002, 106, 2781–2786. [Google Scholar] [CrossRef] [PubMed]
- Hiroki, J.; Shimokawa, H.; Higashi, M.; Morikawa, K.; Kandabashi, T.; Kawamura, N.; Kubota, T.; Ichiki, T.; Amano, M.; Kaibuchi, K.; et al. Inflammatory stimuli upregulate Rho-kinase in human coronary vascular smooth muscle cells. J. Mol. Cell Cardiol. 2004, 37, 537–546. [Google Scholar] [CrossRef]
- Napoli, C.; Nigris, F.D.; Williams-Ignarro, S.; Pignalosa, O.; Sica, V.; Ignarro, L.J. Nitric oxide and atherosclerosis: An update. Nitric Oxide 2006, 15265–15279. [Google Scholar] [CrossRef]
- Kitahara, M.; Kanaki, T.; Ishii, I.; Saito, Y. Atherosclerosis induced by chronic inhibition of the synthesis of nitric oxide in moderately hypercholesterolaemic rabbits is suppressed by pitavastatin. Br. J. Pharmacol. 2010, 159, 1418–1428. [Google Scholar] [CrossRef] [Green Version]
- Eberhardt, R.T.; Loscalzo, J. Nitric oxide in atherosclerosis: Nitric oxide and the cardiovascular system. Humana Press 2000, 273–295. [Google Scholar]
- Maceyka, M.; Harikumar, K.B.; Milstien, S.; Spiegel, S. Sphingosine-1-phosphate signaling and its role in disease. Trends Cell Biol. 2012, 22, 50–60. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rosen, H.; Stevens, R.C.; Hanson, M.; Roberts, E.; Oldstone, M.B. Sphingosine-1-Phosphate and Its Receptors: Structure, Signaling, and Influence. Annu. Rev. Biochem. 2013, 82, 637–662. [Google Scholar] [CrossRef]
- Soltau, I.; Mudersbach, E.; Geissen, M.; Schwedhelm, E.; Winkler, M.S.; Geffken, M.; Peine, S.; Schoen, G.; Debus, E.S.; Larena-Avellaneda, A.; et al. Serum-Sphingosine-1-Phosphate Concentrations Are Inversely Associated with Atherosclerotic Diseases in Humans. PLoS ONE 2016, 11, e0168302. [Google Scholar] [CrossRef] [PubMed]
- Kimura, T.; Boehmler, A.M.; Seitz, G.; Kuçi, S.; Wiesner, T.; Brinkmann, V.; Kanz, L.; Möhle, R. The sphingosine 1-phosphate receptor agonist FTY720 supports CXCR4-dependent migration and bone marrow homing of human CD34 progenitor cells. Blood 2004, 103, 4478–4486. [Google Scholar] [CrossRef] [PubMed]
- Zhao, Y.D.; Ohkawara, H.; Rehman, J.; Wary, K.K.; Vogel, S.M.; Minshall, R.D.; Zhao, Y.Y.; Malik, A.B. Bone marrow progenitor cells induce endothelial adherens junction integrity by sphingosine-1-phosphate-mediated Rac1 and Cdc42 signaling. Circ. Res. 2009, 105, 696–704. [Google Scholar] [CrossRef] [PubMed]
- Wang, H.; Cai, K.Y.; Li, W.; Huang, H. Sphingosine-1-Phosphate Induces the Migration and Angiogenesis of Epcs Through the Akt Signaling Pathway via Sphingosine-1-Phosphate Receptor 3/Platelet-Derived Growth Factor Receptor-β. Cell Mol. Biol. Lett. 2015, 20, 597–611. [Google Scholar] [CrossRef]
- Poitevin, S.; Cussac, D.; Leroyer, A.S.; Albinet, V.; Sarlon-Bartoli, G.; Guillet, B.; Hubert, L.; Andrieu-Abadie, N.; Couderc, B.; Parini, A.; et al. Sphingosine kinase 1 expressed by endothelial colony-forming cells has a critical role in their revascularization activity. Cardiovasc. Res. 2014, 103, 121–130. [Google Scholar] [CrossRef]
- Everaert, B.R.; Van Craenenbroeck, E.M.; Hoymans, V.Y.; Haine, S.E.; Van Nassauw, L.; Conraads, V.M.; Timmermans, J.P.; Vrints, C.J. Current perspective of pathophysiological and interventional effects on endothelial progenitor cell biology: Focus on PI3K/AKT/eNOS pathway. Int. J. Cardiol. 2010, 144, 350–366. [Google Scholar] [CrossRef]
- Katusic, Z.S.; Austin, S.A. Endothelial nitric oxide: Protector of a healthy mind. Eur. Heart J. 2014, 35, 888–894. [Google Scholar] [CrossRef]
- Morales-Ruiz, M.; Lee, M.J.; Zöllner, S.; Gratton, J.P.; Scotland, R.; Shiojima, I.; Walsh, K.; Hla, T.; Sessa, W.C. Sphingosine 1-phosphate activates Akt, nitric oxide production, and chemotaxis through a gi protein/phosphoinositide 3-kinase pathway in endothelial cells. J. Biol. Chem. 2001, 276, 19672–19677. [Google Scholar] [CrossRef] [PubMed]
- Wang, H.; Huang, H.; Ding, S.F. Sphingosine-1-phosphate promotes the proliferation and attenuates apoptosis of Endothelial progenitor cells via S1PR1/S1PR3/PI3K/Akt pathway. Cell Biol Int. 2018, 42, 1492–1502. [Google Scholar] [CrossRef] [PubMed]
- Nakagawa, H.; Nihonmatsu, N.; Ohta, S.; Hirobe, M. Effects of new endogenous nonprotein amino acids, 1, 2, 3, 4-tetrahydroisoquinoline-3-carboxylic acid derivatives, on behaviorof mice. Biochem. Biophys. Res. Commun. 1996, 225, 1027–1034. [Google Scholar] [CrossRef] [PubMed]
- Robinson, E.S.; Anderson, N.J.; Crosby, J.; Nutt, D.J.; Hudson, A.L. Endogenous beta-carbolines as clonidine-displacing substances. Ann. N. Y. Acad. Sci. 2003, 1009, 157–166. [Google Scholar] [CrossRef] [PubMed]
- Nakagawa, H.; Yamada, M.; Maeda, N.; Iwatsuki, K.; Hirayama, A.; Ikenaka, K. Clinical trial of intrathecal administration of 5-fluoro-2′-deoxyuridine for treatment of meningeal dissemination of malignant tumors. J. Neurooncol. 1999, 45, 175–183. [Google Scholar] [CrossRef]
Sample Availability: Samples of the compounds Sphingosine-1-Phosphate, LY294002 and L-NAME are available from the authors. |





© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Wang, X.; Zhan, E.; Lu, G.; Mu, Q.; Zhang, T.; Yang, N. Sphingosine-1-Phosphate Improves the Biological Features of Mouse Bone Marrow-Derived EPCs Partially through PI3K/AKT/eNOS/NO Pathway. Molecules 2019, 24, 2404. https://doi.org/10.3390/molecules24132404
Wang X, Zhan E, Lu G, Mu Q, Zhang T, Yang N. Sphingosine-1-Phosphate Improves the Biological Features of Mouse Bone Marrow-Derived EPCs Partially through PI3K/AKT/eNOS/NO Pathway. Molecules. 2019; 24(13):2404. https://doi.org/10.3390/molecules24132404
Chicago/Turabian StyleWang, Xia, Enxin Zhan, Guohua Lu, Qingjie Mu, Tianliang Zhang, and Nana Yang. 2019. "Sphingosine-1-Phosphate Improves the Biological Features of Mouse Bone Marrow-Derived EPCs Partially through PI3K/AKT/eNOS/NO Pathway" Molecules 24, no. 13: 2404. https://doi.org/10.3390/molecules24132404