Polysaccharide from Okra (Abelmoschus esculentus (L.) Moench) Improves Antioxidant Capacity via PI3K/AKT Pathways and Nrf2 Translocation in a Type 2 Diabetes Model

 ,
,
Abstract
:1. Introduction
2. Results
2.1. Physicochemical Property Analysis of OP
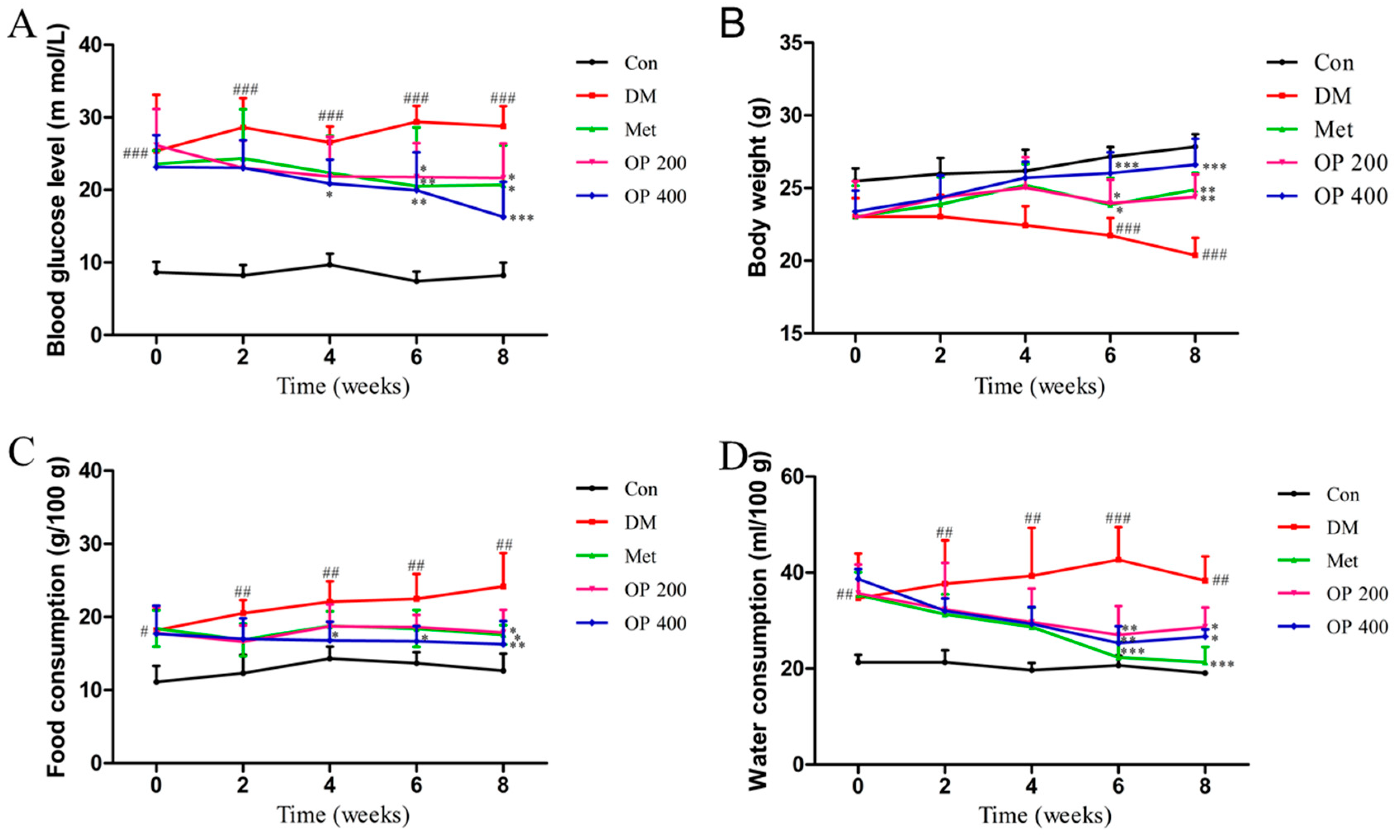
2.2. Effects of OP on Fasting Blood Glucose (FBG) Levels, Body Weight, Food, and Water Consumption
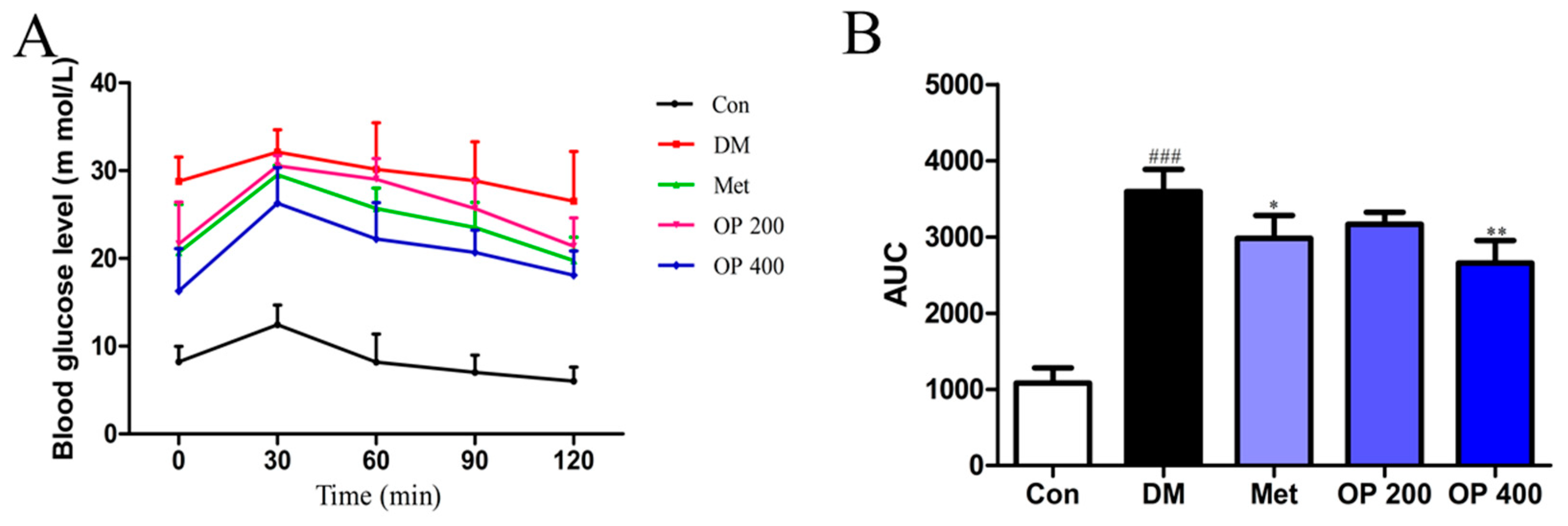
2.3. Effects of OP on an Oral Glucose Tolerance Test (OGTT)
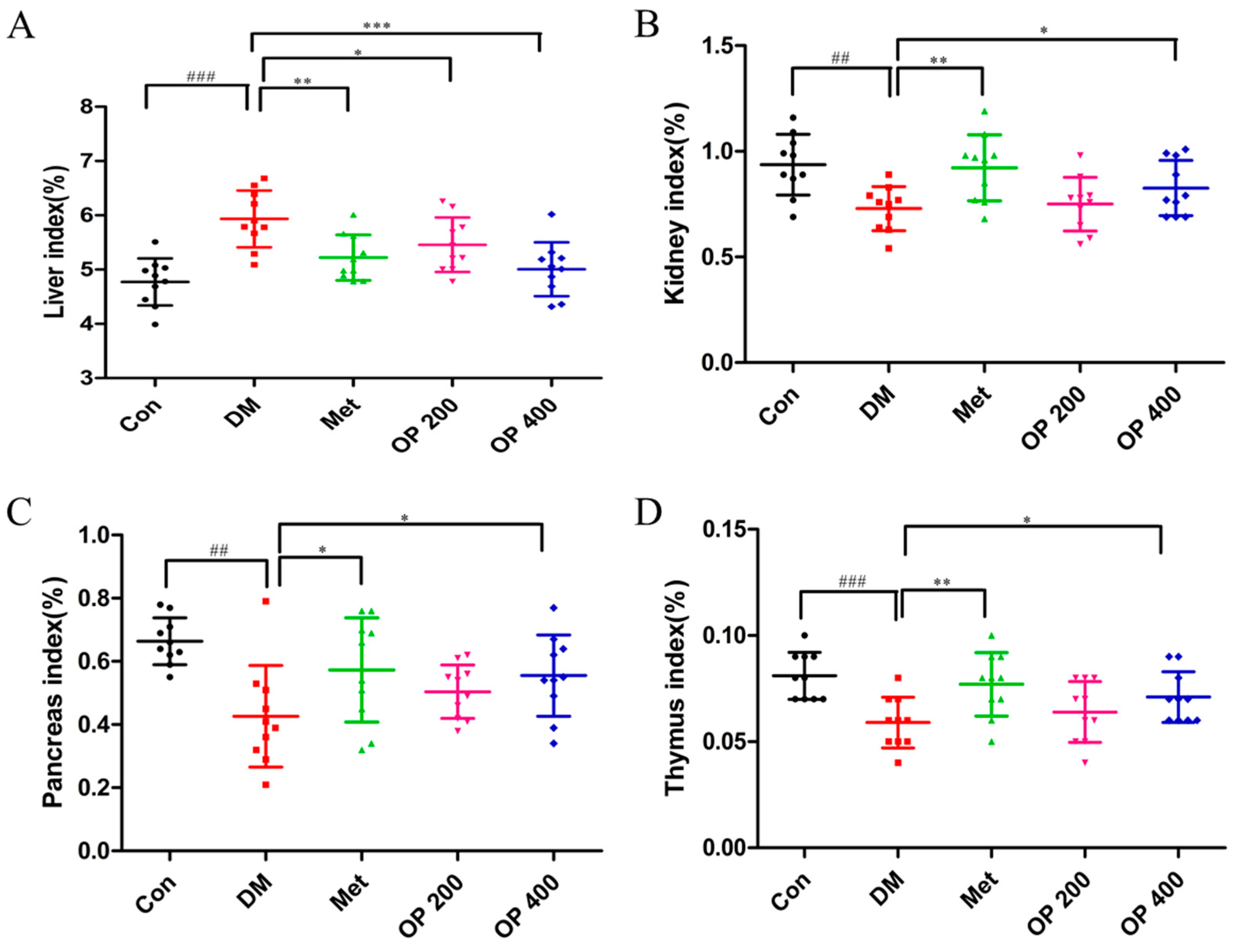
2.4. Effects of OP on Organ Index
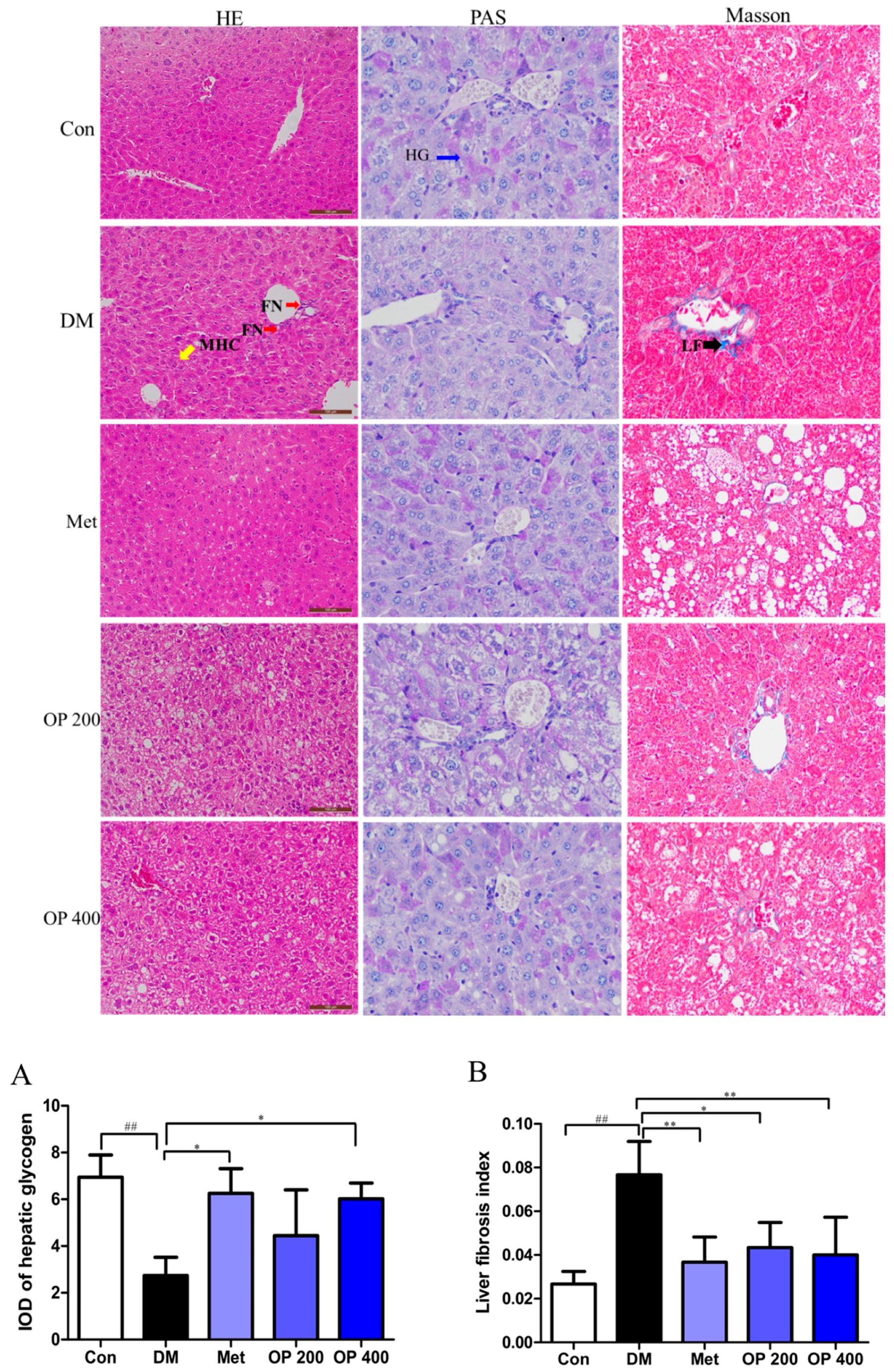
2.5. Effect of OP on the T2DM-Induced Histopathological Changes in the Liver
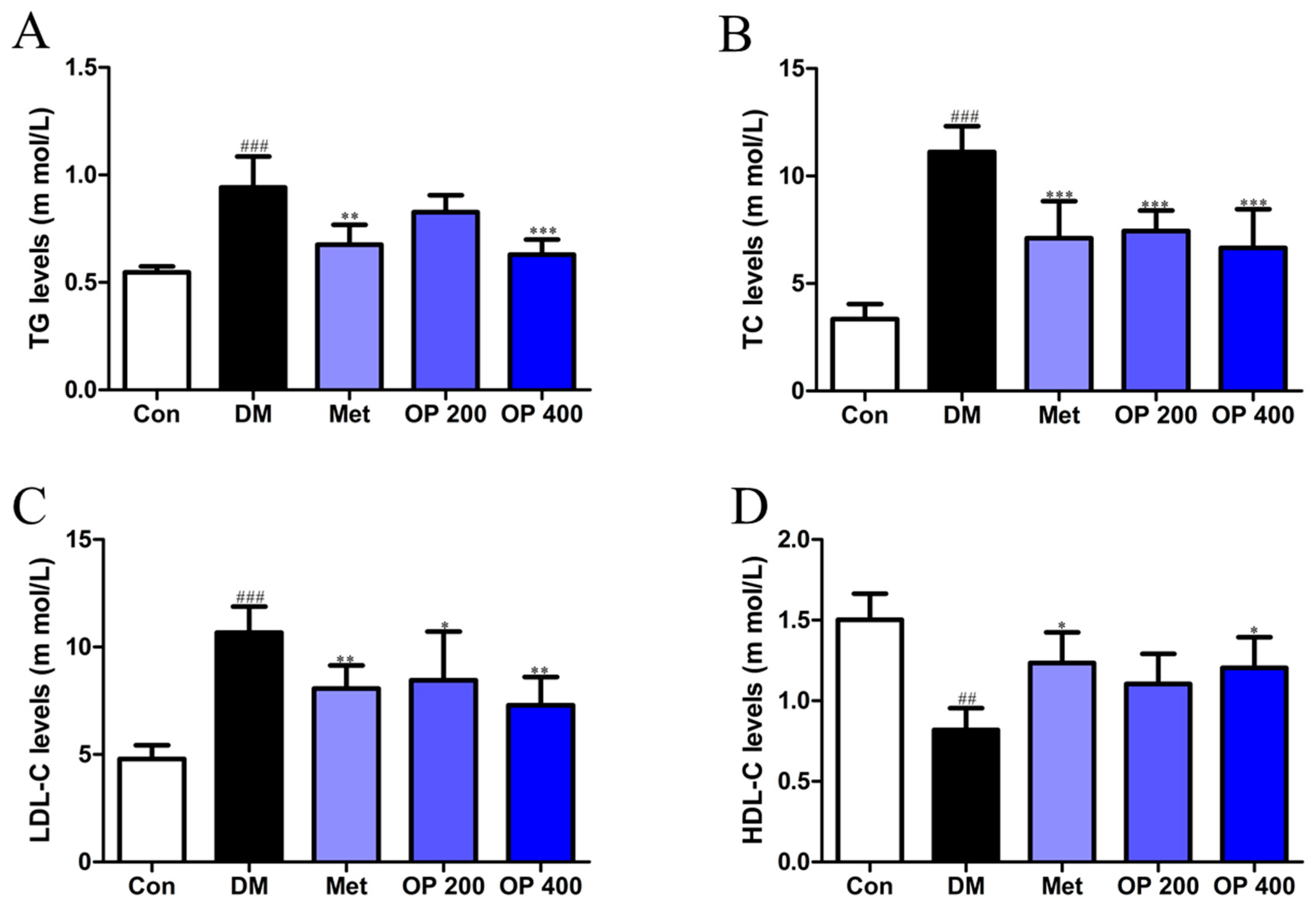
2.6. Effects of OP on Serum Lipid Metabolism of T2DM Mice
2.7. Effects of OP on Superoxide Dismutase (SOD), Catalase (CAT), Glutathione Peroxidase (GSH-Px), and Malondialdehyde (MDA) in Serum and Liver
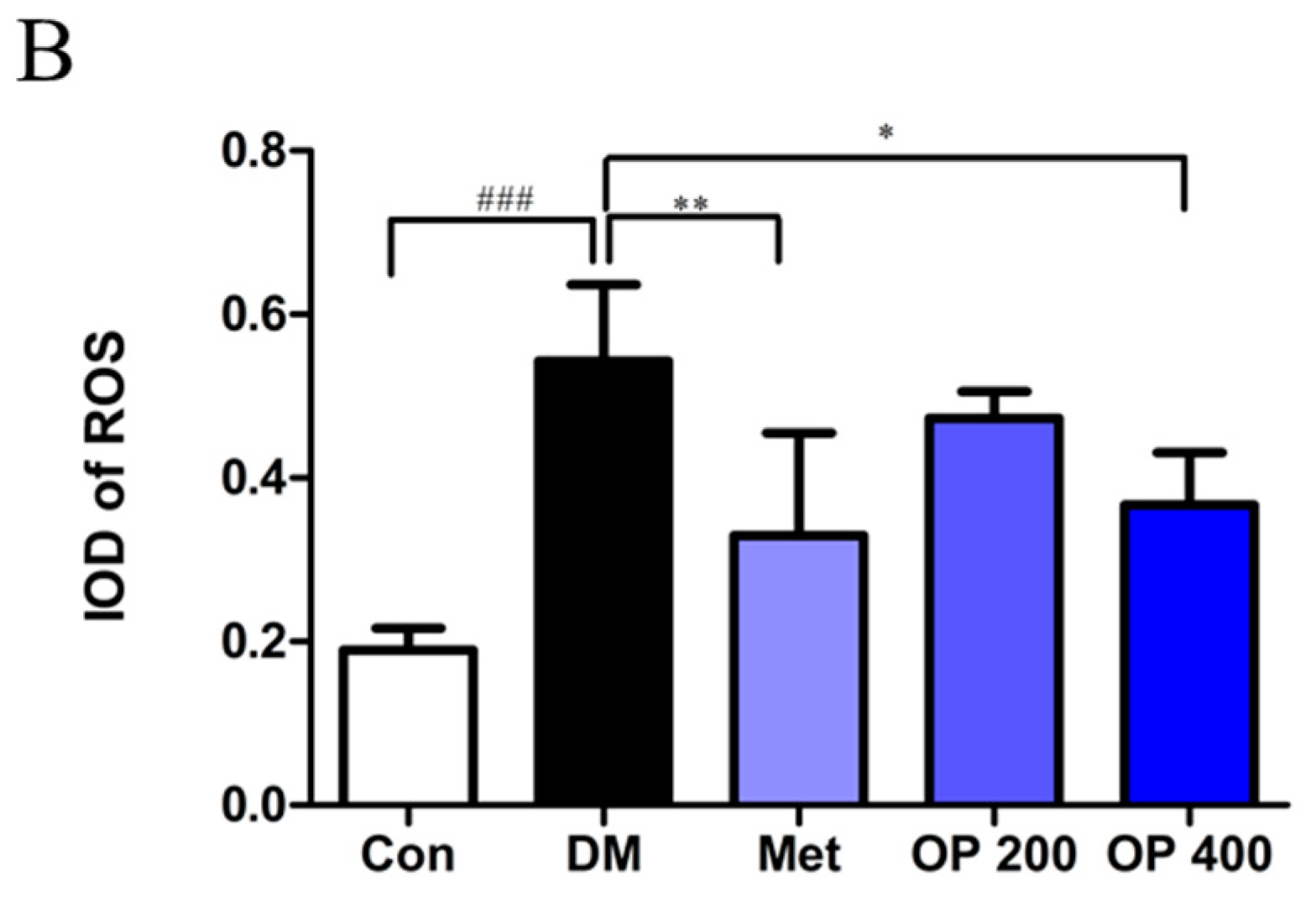
2.8. Effect of OP on ROS in Liver
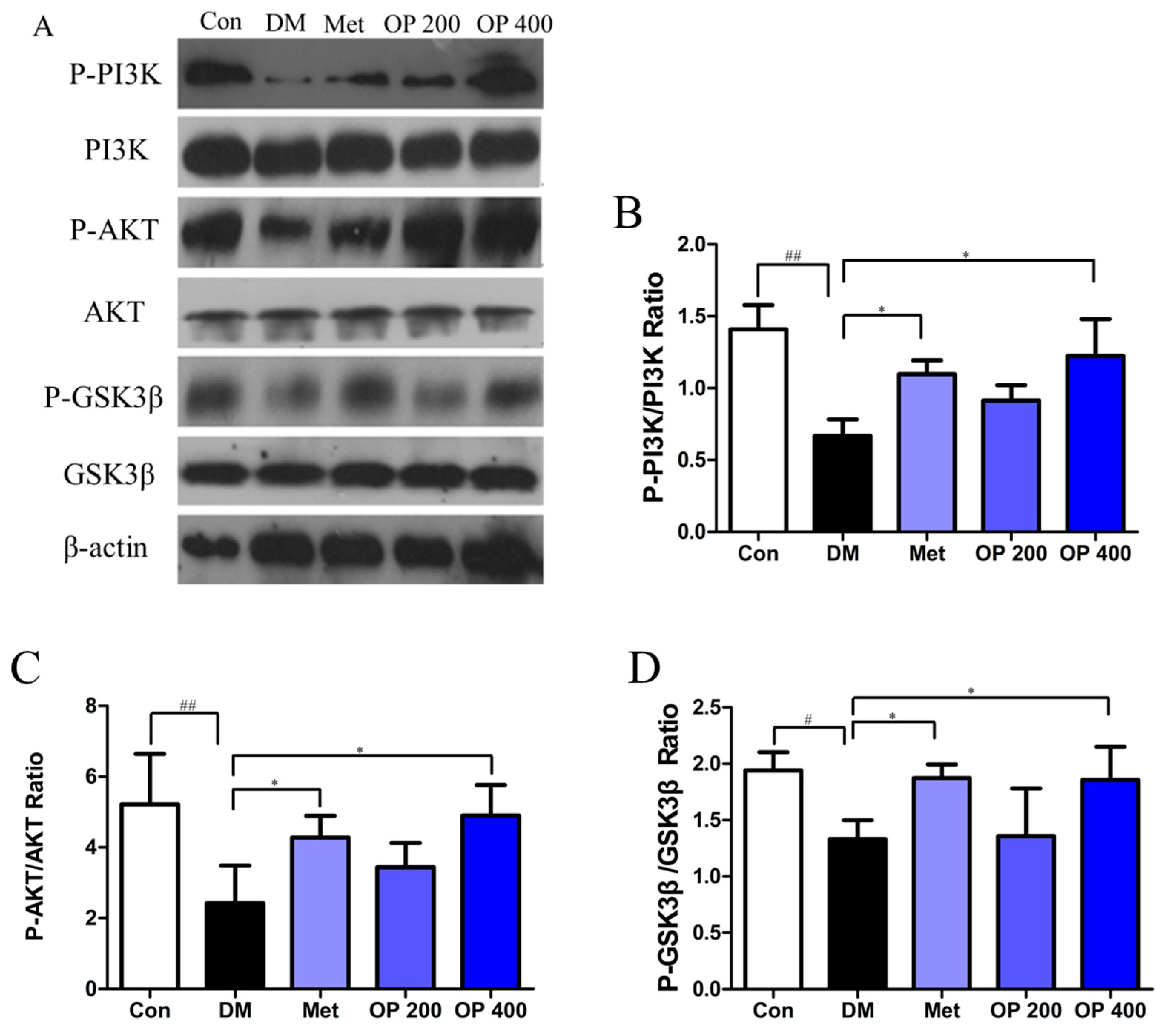
2.9. Effect of OP on P-PI3K, PI3K, P-AKT, AKT, P-GSK3β, and GSK3β in Liver
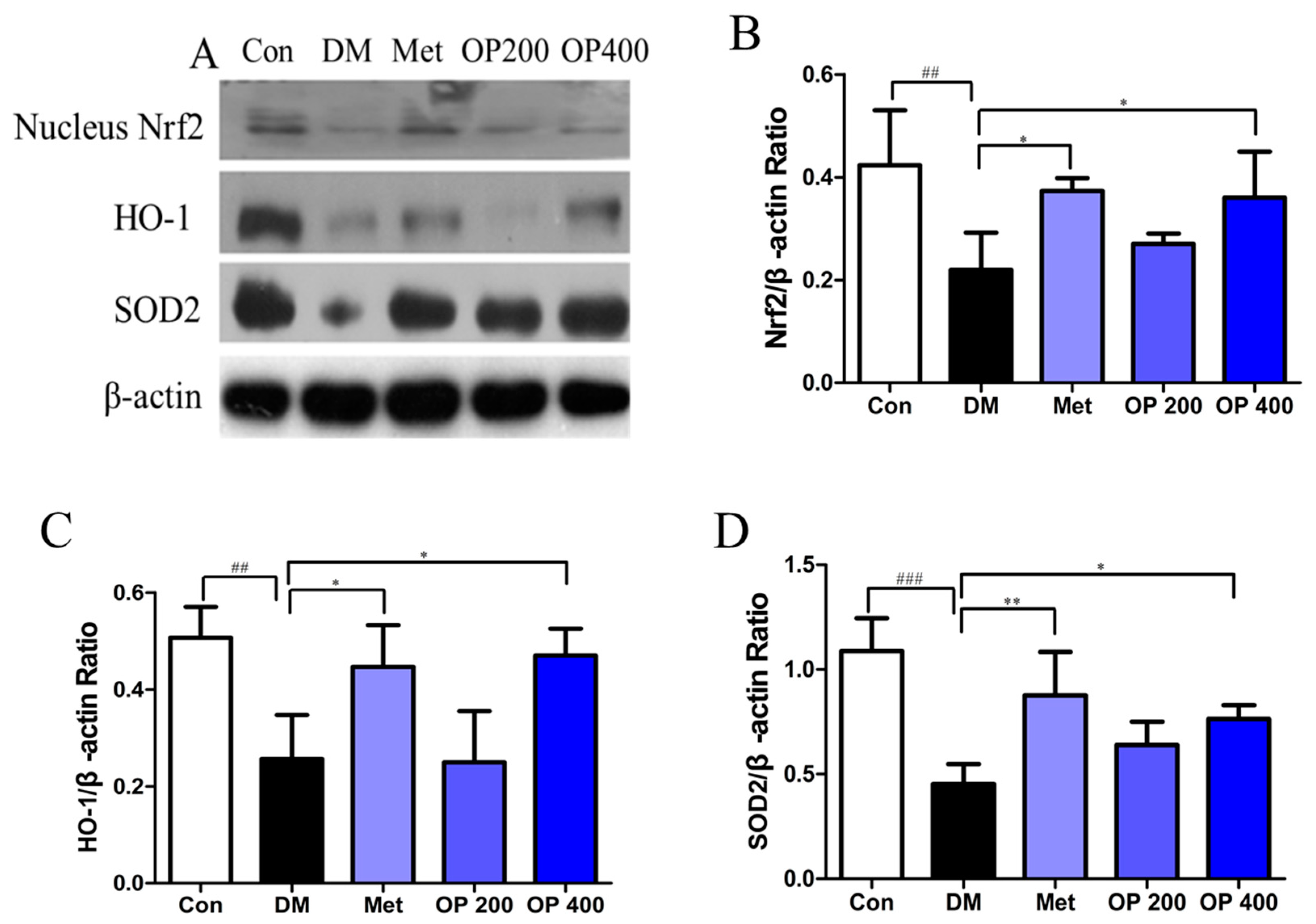
2.10. Effect of OP on the Expression of Nrf2, Heme Oxygenase-1 (HO-1), and Superoxide Dismutase 2 (SOD2) in the Liver
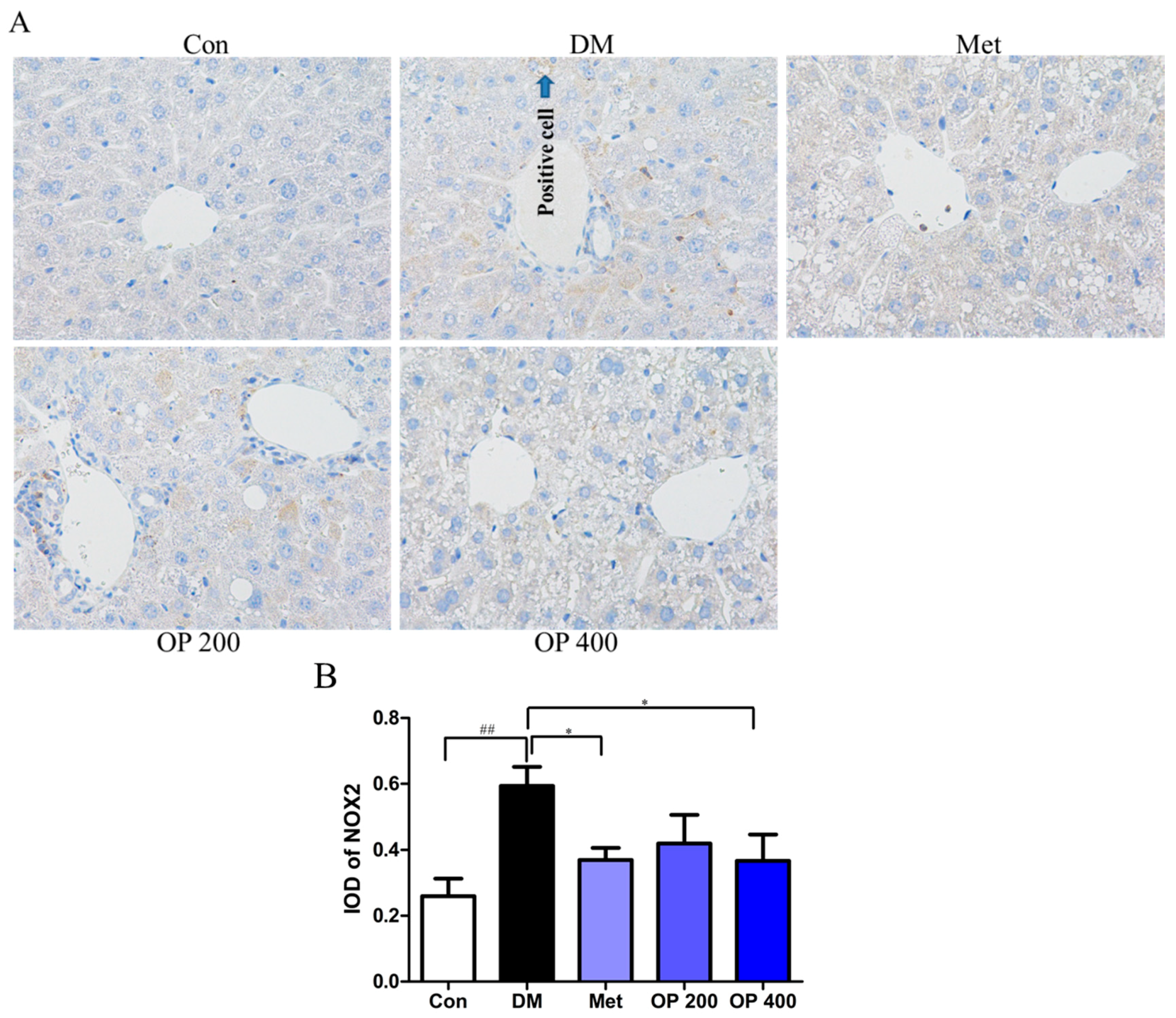
2.11. Effect of OP on the Nicotinamide Adenine Dinucleotide Phosphate Oxidases 2 (NOX2) Expression in the Liver
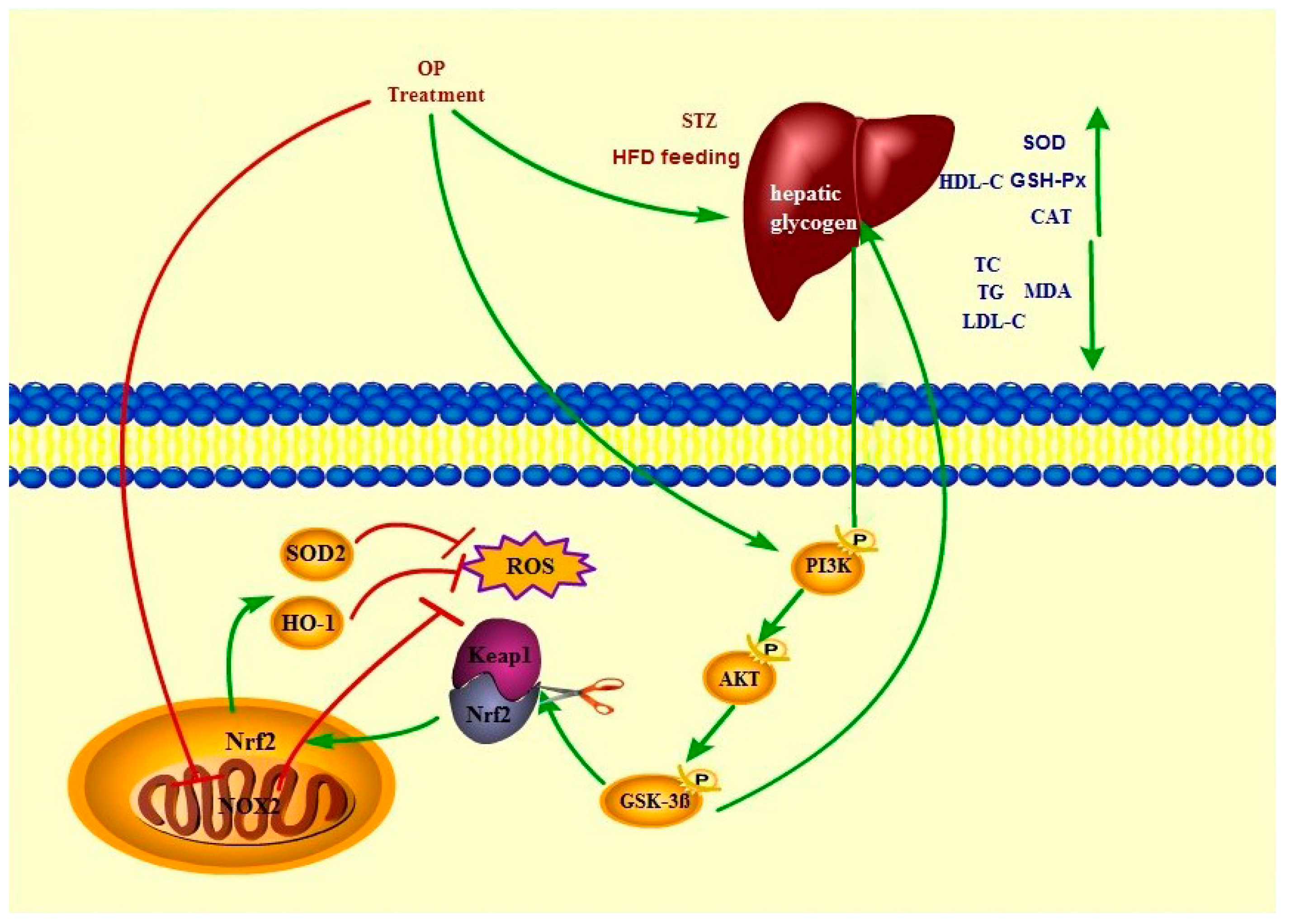
3. Discussion
4. Materials and Methods
4.1. Extracted and Purified Polysaccharides from Okra (OP)
4.2. The Determination of Composition of OP
4.3. Animal Diets and Experimental Design
4.4. Oral Glucose Tolerance Tests (OGTT)
4.5. Serum Collection and Liver Tissues Preparation
4.6. Biochemical Analysis
4.6.1. Enzyme linked immunosorbent assay (ELISA) Assay Kits
4.6.2. Western Blot
4.6.3. Immunohistochemical (IHC) and Immunofluorescence Analyses
4.6.4. Histopathological Examination
4.7. Statistical Analysis
Supplementary Materials
Author Contributions
Funding
Conflicts of Interest
References
- International Diabetes Federation. IDF Diabetes Atlas. 2017. Available online: http://www.idf.org/diabetesatlas (accessed on 17 May 2019).
- Wang, H.Y.; Kan, W.C.; Cheng, T.J.; Yu, S.H.; Chang, L.H.; Chuu, J.J. Differential anti-diabetic effects and mechanism of action of charantin-rich extract of Taiwanese Momordica charantia between type 1 and type 2 diabetic mice. Food Chem. Toxicol. 2014, 69, 347–356. [Google Scholar] [CrossRef]
- Rochester, C.D.; Akiyode, O. Novel and emerging diabetes mellitus drug therapies for the type 2 diabetes patient. World J. Diabetes 2014, 5, 305–315. [Google Scholar] [CrossRef]
- Manickavasagam, M.; Subramanyam, K.; Ishwarya, R.; Elayaraja, D.; Ganapathi, A. Assessment of factors influencing the tissue culture-independent Agrobacterium-mediated in planta genetic transformation of okra [Abelmoschus esculentus (L.) Moench]. Plant. Cell Tiss. Org. 2015, 123, 309–320. [Google Scholar] [CrossRef]
- Ezureike, U.F.; Prieto, J.M. The use of plants in the traditional management of diabetes in Nigeria: Pharmacological and toxicological considerations. J. Ethnopharmacol. 2014, 155, 857–924. [Google Scholar] [CrossRef] [Green Version]
- Ong, K.W.; Hsu, A.; Tan, B.K.H. Anti-diabetic and anti-lipidemic effects of chlorogenic acid are mediated by ampk activation. Biochem. Pharmacol. 2013, 85, 1341–1351. [Google Scholar] [CrossRef]
- Nichols, K.W.; Kimes, T.M.; Harp, J.B.; Kou, T.D.; Brodovicz, K.G. Glycemic Response and Attainment of A1C Goals Following Newly Initiated Insulin Therapy for Type 2 Diabetes. Diabetes Care 2012, 35, 495–497. [Google Scholar] [CrossRef] [Green Version]
- Wu, J.J.; Shi, S.S.; Wang, H.J.; Wang, S.C. Mechanisms underlying the effect of polysaccharides in the treatment of type 2 diabetes: A review. Carbohyd. Polym. 2016, 144, 474–494. [Google Scholar] [CrossRef]
- Arapitsas, P. Identification and quantification of polyphenolic compounds from okra seeds and skins. Food Chem. 2008, 110, 1041–1045. [Google Scholar] [CrossRef]
- Fan, S.; Guo, L.; Zhang, Y.; Sun, Q.; Yang, B.; Huang, C. Okra polysaccharide improves metabolic disorders in high-fat diet-induced obese C57BL/6 mice. Mol. Nutr. Food Res. 2013, 57, 2075–2078. [Google Scholar] [CrossRef]
- Liu, J.; Zhao, Y.P.; Wu, Q.X.; John, A.; Jiang, Y.M.; Yang, J.L.; Liu, H.L.; Yang, B. Structure characterisation of polysaccharides in vegetable “okra” and evaluation of hypoglycemic activity. Food Chem. 2018, 242, 211–216. [Google Scholar] [CrossRef]
- Wang, J.; Wang, C.; Li, S.Q.; Li, W.W.; Yuan, G.Q.; Pan, Y.X.; Chen, H.X. Anti-diabetic effects of Inonotus obliquus polysaccharides in streptozotocin-induced type 2 diabetic mice and potential mechanism via PI3K-Akt signal pathway. Biomed. Pharmacother. 2017, 95, 1669–1677. [Google Scholar] [CrossRef]
- Logie, L.; Ruiz-alcaraz, A.J.; Keane, M.; Woods, Y.L.; Bain, J.; Marquez, R.; Alessi, D.R.; Sutherland, C. Characterization of a protein kinase B inhibitor in vitro and in insulin-treated liver cells. Diabetes 2007, 56, 2218–2227. [Google Scholar] [CrossRef]
- Li, C.; Xia, J.Y.; Wan, Q. Interaction of the Nrf2/ARE and IRS-PI3K/AKT pathways in pancreatic β-cells and the underlying mechanisms. Acta Medica Mediterr. 2018, 34, 1079–1084. [Google Scholar]
- Fernández-Millán, E.; Martín, M.A.; Goya, L.; Lizárraga-Mollinedo, E.; Escrivá, F.; Ramos, S.; Álvarez, C. Glucagon-like peptide-1 improves beta-cell antioxidant capacity via extracellular regulated kinases pathway and Nrf2 translocation. Free Radic. Biol. Med. 2016, 95, 16–26. [Google Scholar] [CrossRef] [Green Version]
- Islam, M.T. Phytochemical information and pharmacological activities of Okra (Abelmoschus esculentus): A literature-based review. Phytother. Res. 2019, 33, 72–80. [Google Scholar] [CrossRef]
- Yuan, Q.; Lin, S.; Fu, Y.; Nie, X.R.; Liu, W.; Su, Y.; Han, Q.H.; Zhao, L.; Zhang, Q.; Lin, D.R.; et al. Effects of extraction methods on the physicochemical characteristics and biological activities of polysaccharides from okra (Abelmoschus esculentus). Int. J. Biol. Macromol. 2019, 127, 178–186. [Google Scholar] [CrossRef]
- Lee, H.; Lim, Y. Tocotrienol-rich fraction supplementation reduces hyperglycemia-induced skeletal muscle damage through regulation of insulin signaling and oxidative stress in type 2 diabetic mice. J. Nutr. Biochem. 2018, 57, 77–85. [Google Scholar] [CrossRef]
- Chung, I.M.; Kim, E.H.; Yeo, M.A.; Kim, S.J.; Seo, M.C.; Moon, H.I. Antidiabetic effects of three Korean sorghum phenolic extracts in normal and streptozotocin-induced diabetic rats. Food Res. Int. 2011, 44, 127–132. [Google Scholar] [CrossRef]
- Chen, J.C.; Li, L.; Zhou, X.; Sun, P.Y.; Li, B.; Zhang, X. Preliminary characterization and antioxidant and hypoglycemic activities in vivo of polysaccharides from Huidouba. Food Funct. 2018, 9, 6337–6348. [Google Scholar] [CrossRef]
- Heeba, G.H.; Hamza, A.A. Rosuvastatin ameliorates diabetes-induced reproductive damage via suppression of oxidative stress, inflammatory and apoptotic pathways in male rats. Life Sci. 2015, 141, 13–19. [Google Scholar] [CrossRef]
- Fatani, A.J.; Alrejaie, S.S.; Abuohashish, H.M.; Alassaf, A.; Parmar, M.Y.; Ahmed, M.M. Lutein Dietary Supplementation Attenuates Streptozotocin-induced testicular damage and oxidative stress in diabetic rats. BMC Complement. Altern. Med. 2015, 15, 204. [Google Scholar] [CrossRef]
- Dixit, Y.; Kar, A. Protective role of three vegetable peels in alloxan induced diabetes mellitus in male mice. Plant Foods Hum. Nutr. 2010, 65, 284–289. [Google Scholar] [CrossRef]
- Li, S.; Chen, H.; Wang, J.; Wang, X.; Hu, B.; Lv, F. Involvement of the PI3K/Akt signal pathway in the hypoglycemic effects of tea polysaccharides on diabetic mice. Int. J. Biol. Macromol. 2015, 81, 967–974. [Google Scholar] [CrossRef]
- Wang, L.Y.; Wang, Y.; Xu, D.S.; Ruan, K.F.; Feng, Y.; Wang, S. MDG-1, a polysaccharide from Ophiopogon japonicus exerts hypoglycemic effects through the PI3K/Akt pathway in a diabetic KKAy mouse model. J. Ethnopharmacol. 2012, 143, 347–354. [Google Scholar] [CrossRef]
- Meenatchi, P.; Purushothaman, A.; Maneemegalai, S. Antioxidant, antiglycation and insulinotrophic properties of Coccinia grandis (L.) in vitro: Possible role in preven-tion of diabetic complications. J. Tradit. Complement. Med. 2015, 7, 54–64. [Google Scholar] [CrossRef]
- Yu, Y.; Bai, F.; Liu, Y.; Yang, Y.; Yuan, Q.; Zou, D.; Qu, S.; Tian, G.; Song, L.; Zhang, T. Fibroblast growth factor (FGF21) protects mouse liver against d-galactose-induced oxidative stress and apoptosis via activating Nrf2 and PI3K/Akt pathways. Mol. Cell. Biochem. 2015, 403, 287–299. [Google Scholar] [CrossRef]
- Sun, X.; Yang, Y.; Shi, J.; Wang, C.; Yu, Z.; Zhang, H. NOX4-and Nrf2-mediated oxidative stress induced by silver nanopar-ticles in vascular endothelial cells. J. Appl. Toxicol. 2017, 37, 1428–1437. [Google Scholar] [CrossRef]
- Wang, X.; Gu, C.S.; He, W.; Ye, X.L.; Chen, H.L.; Zhang, X.D.; Hai, C.X. Glucose oxidase induces insulin resistance via influencing multiple targets in vitro and in vivo: The central role of oxidative stress. Biochimie 2012, 94, 1705–1717. [Google Scholar] [CrossRef]
- Li, N.; Li, B.; Brun, T.; Deffert-Delbouille, C.; Mahiout, Z.; Daali, Y.; Ma, X.J.; Krause, K.H.; Maechler, P. NADPH oxidase NOX2 defines a new antagonistic role for reactive oxygen species and cAMP/PKA in the regulation of insulin secretion. Diabetes 2012, 61, 2842–2850. [Google Scholar] [CrossRef]
- DuBois, M.; Gilles, K.A.; Hamilton, J.K.; Rebers, P.A.; Smith, F. Colorimetric method for determination of sugars and related substances. Anal. Chem. 1956, 28, 350–356. [Google Scholar] [CrossRef]
- Bradford, M.M. A rapid and sensitive method for the quantitation of microgram quantities of protein utilizing the principle of protein-dye binding. Anal. Biochem. 1976, 72, 248–254. [Google Scholar] [CrossRef]
- Dodgson, K.S.; Price, R.G. A note on the determination of the ester sulphate content of sulphated polysaccharides. Biochem. J. 1962, 84, 106–110. [Google Scholar] [CrossRef] [Green Version]
- Chen, Z.; Zhang, W.; Tang, X.Y.; Fan, H.J.; Xie, X.J.; Wan, Q.; Wu, X.H.; Tang, J.Z. Extraction and characterization of polysaccharides from Semen Cassiae by microwave-assisted aqueous two-phase extraction coupled with spectroscopy and HPLC. Carbohyd. Polym. 2016, 144, 263–270. [Google Scholar] [CrossRef] [PubMed]
Sample Availability: Samples of the compounds are not available from the authors. |












{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Yield (%) a | Polysaccharide (%) | Protein (%) | Sulfate (%) | Mannose | Rhamnose | Glucuronic Acid | Galactosal Acid | Galactose | Arabia |
|---|---|---|---|---|---|---|---|---|---|
| 0.59 | 90.93 ± 0.76 | 0.69 ± 0.03 | 6.15 ± 0.19 | 3.4 ± 0.02 | 3.76 ± 0.01 | 24.19 ± 0.02 | 6.27 ± 0.01 | 8.73 ± 0.02 | 3.13 ± 0.02 |
| Parameter | Con | DM | Met | OP | |
|---|---|---|---|---|---|
| 200 mg/kg | 400 mg/kg | ||||
| SOD (U/mg prot) | 306.89 ± 73.9 | 201.87 ± 47.1 ## | 284.54 ± 19.5 * | 252.51 ± 46.5 | 274.18 ± 24.1 * |
| CAT (U/mg prot) | 65.13 ± 11.1 | 47.63 ± 7.79 ## | 57.80 ± 6.60 * | 50.32 ± 3.09 | 57.09 ± 6.91 * |
| GSH-Px (U/mg prot) | 562.53 ± 151 | 411.82 ± 26.4 # | 526.63 ± 59.1 * | 461.05 ± 90.2 | 530.08 ± 45.1 * |
| MDA (n mol/mg prot) | 2.71 ± 0.44 | 4.27 ± 0.31 ### | 3.17 ± 0.55 ** | 4.02 ± 0.71 | 3.48 ± 0.45 * |
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Liao, Z.; Zhang, J.; Liu, B.; Yan, T.; Xu, F.; Xiao, F.; Wu, B.; Bi, K.; Jia, Y. Polysaccharide from Okra (Abelmoschus esculentus (L.) Moench) Improves Antioxidant Capacity via PI3K/AKT Pathways and Nrf2 Translocation in a Type 2 Diabetes Model. Molecules 2019, 24, 1906. https://doi.org/10.3390/molecules24101906
Liao Z, Zhang J, Liu B, Yan T, Xu F, Xiao F, Wu B, Bi K, Jia Y. Polysaccharide from Okra (Abelmoschus esculentus (L.) Moench) Improves Antioxidant Capacity via PI3K/AKT Pathways and Nrf2 Translocation in a Type 2 Diabetes Model. Molecules. 2019; 24(10):1906. https://doi.org/10.3390/molecules24101906
Chicago/Turabian StyleLiao, Zhengzheng, Jingying Zhang, Bing Liu, Tingxu Yan, Fanxing Xu, Feng Xiao, Bo Wu, Kaishun Bi, and Ying Jia. 2019. "Polysaccharide from Okra (Abelmoschus esculentus (L.) Moench) Improves Antioxidant Capacity via PI3K/AKT Pathways and Nrf2 Translocation in a Type 2 Diabetes Model" Molecules 24, no. 10: 1906. https://doi.org/10.3390/molecules24101906