
Low Levels of IgM and IgA Recognizing Acetylated C1-Inhibitor Peptides Are Associated with Systemic Lupus Erythematosus in Taiwanese Women


Abstract
:
1. Introduction
2. Results
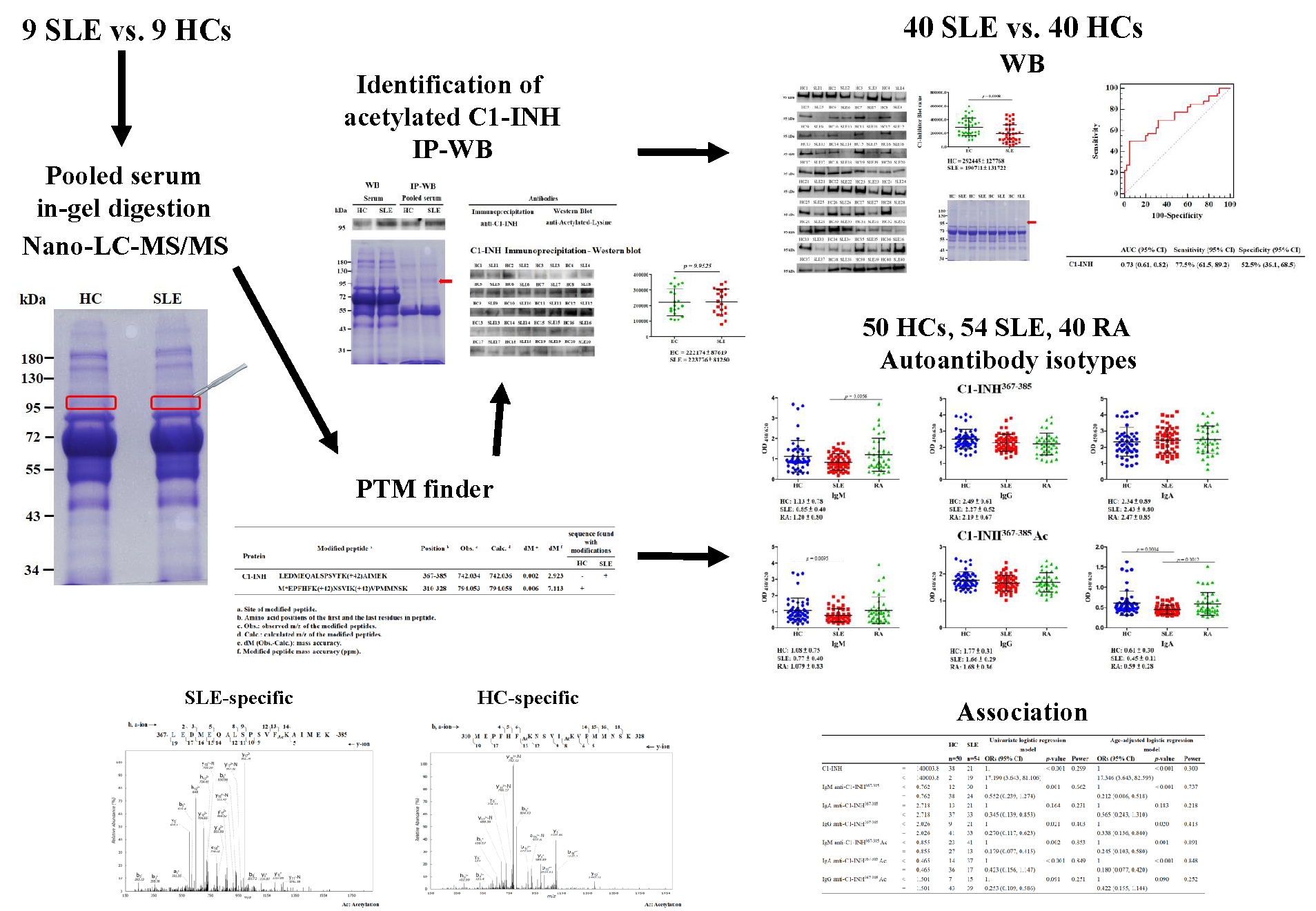
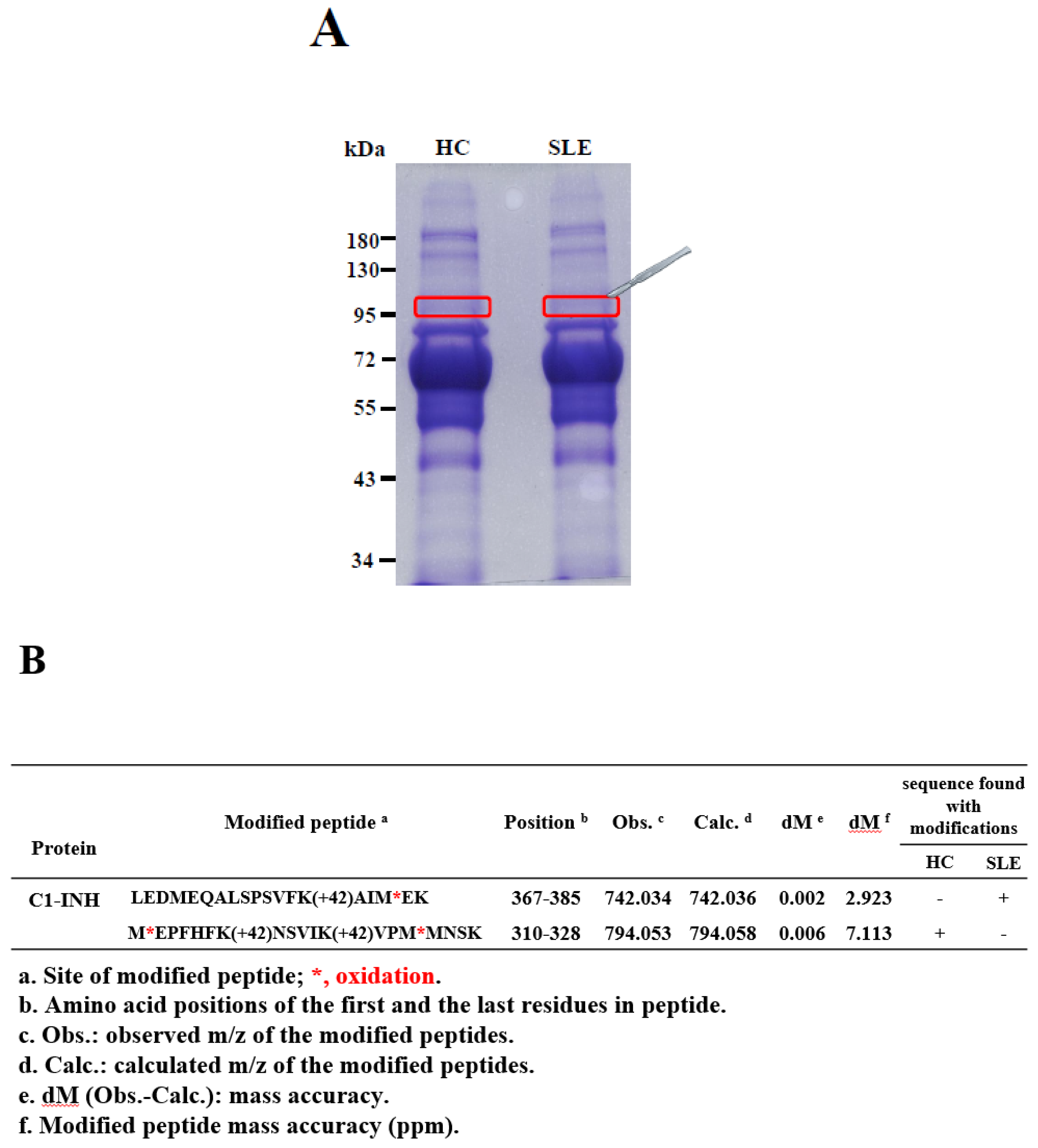
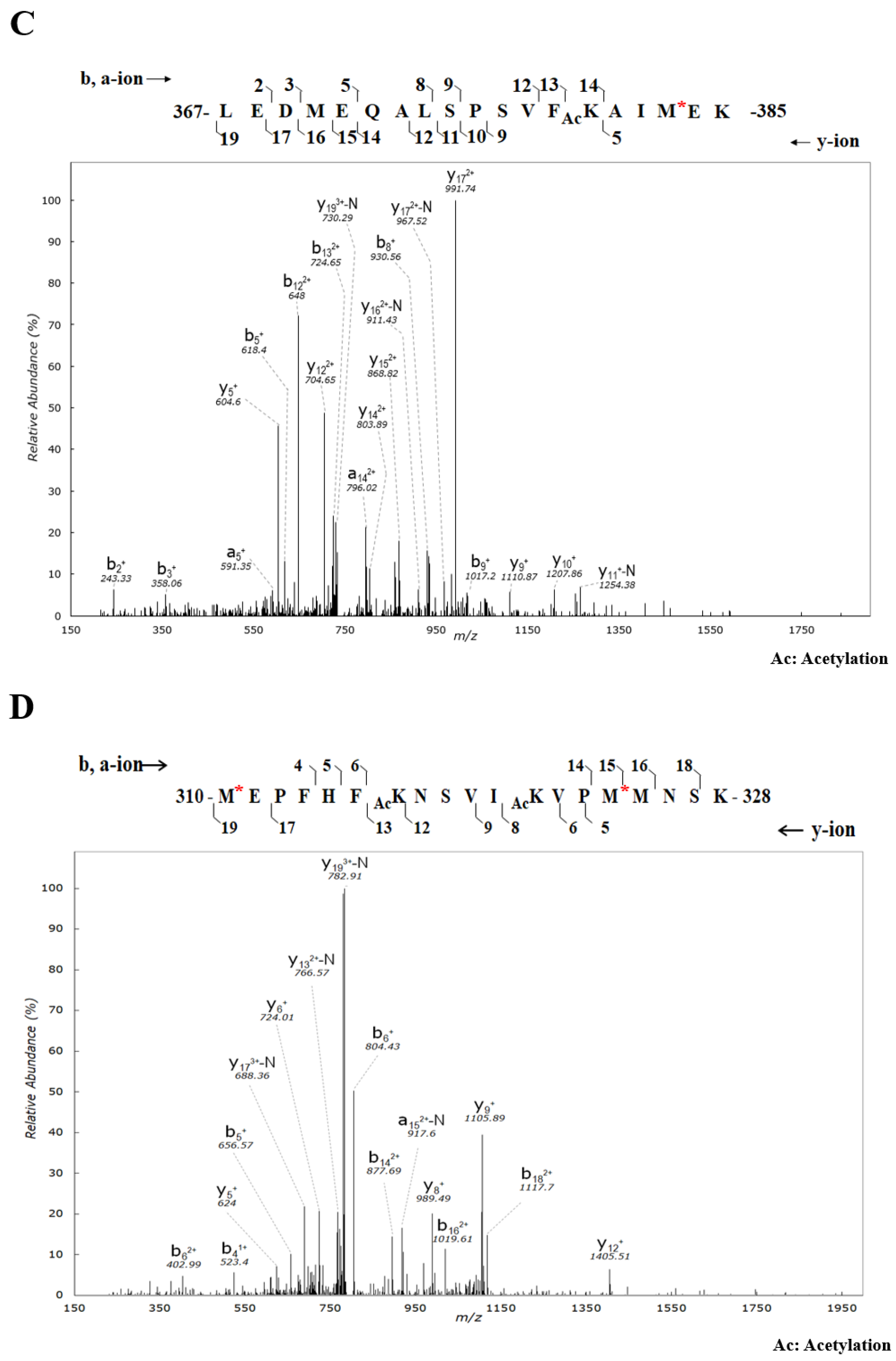
2.1. Identification and Validation of Novel Ac Modifications of Serum C1-INH
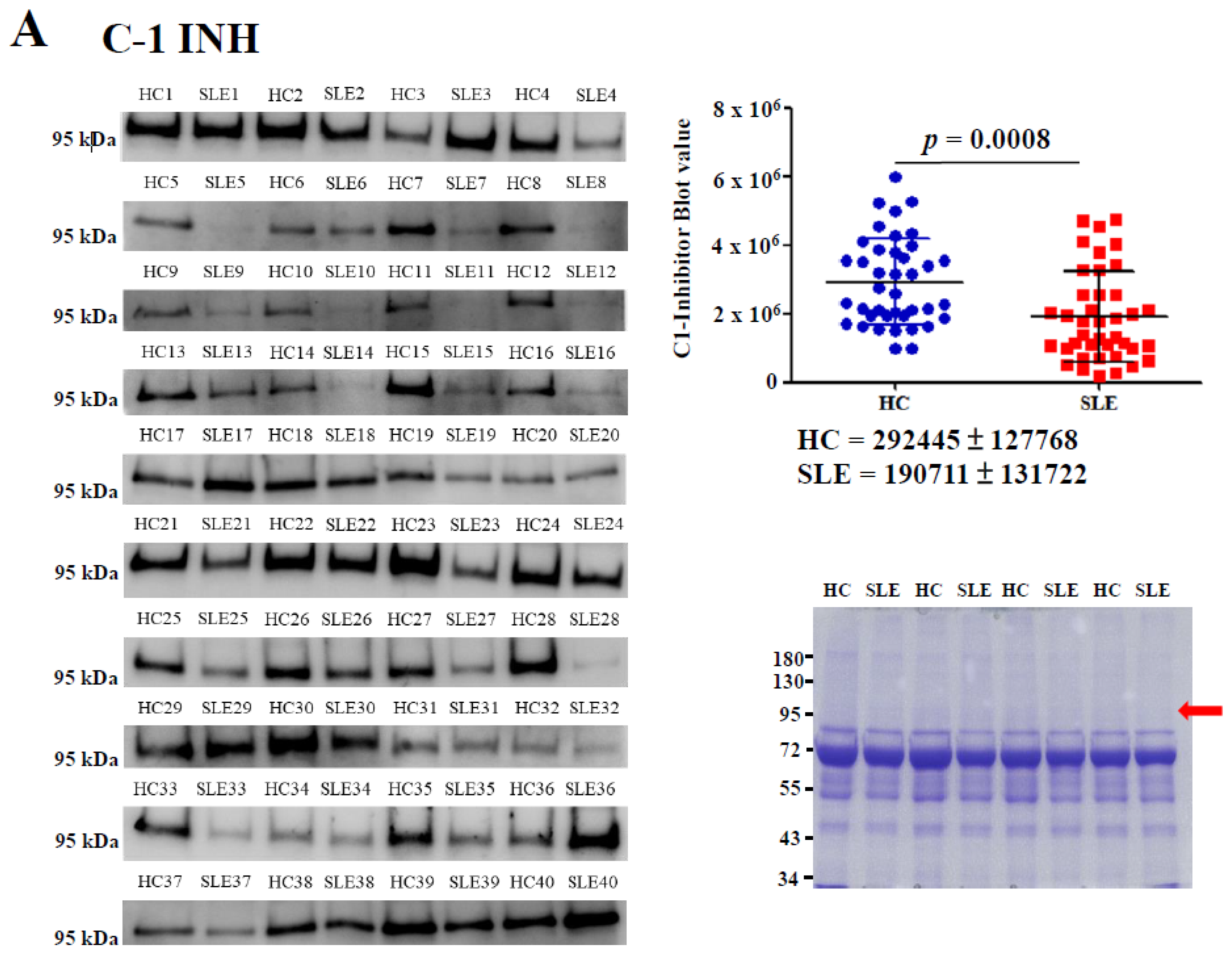
2.2. Determination of Serum C1-INH Levels Using Western Blotting
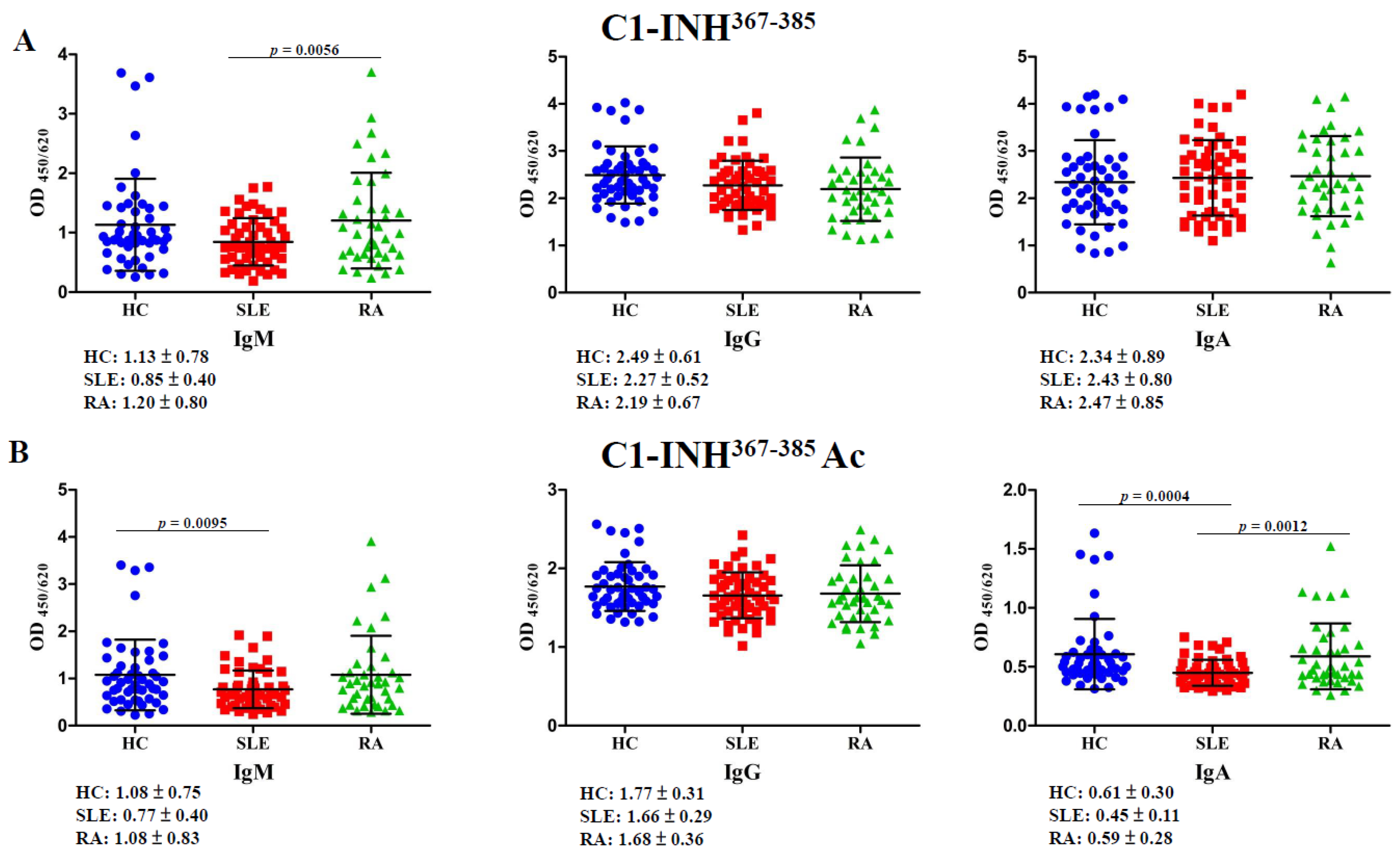
2.3. Autoantibodies Against C1-INH367–385 and C1-INH367–385 Ac Peptides
2.4. Determination of Serum Ac-Protein Adducts
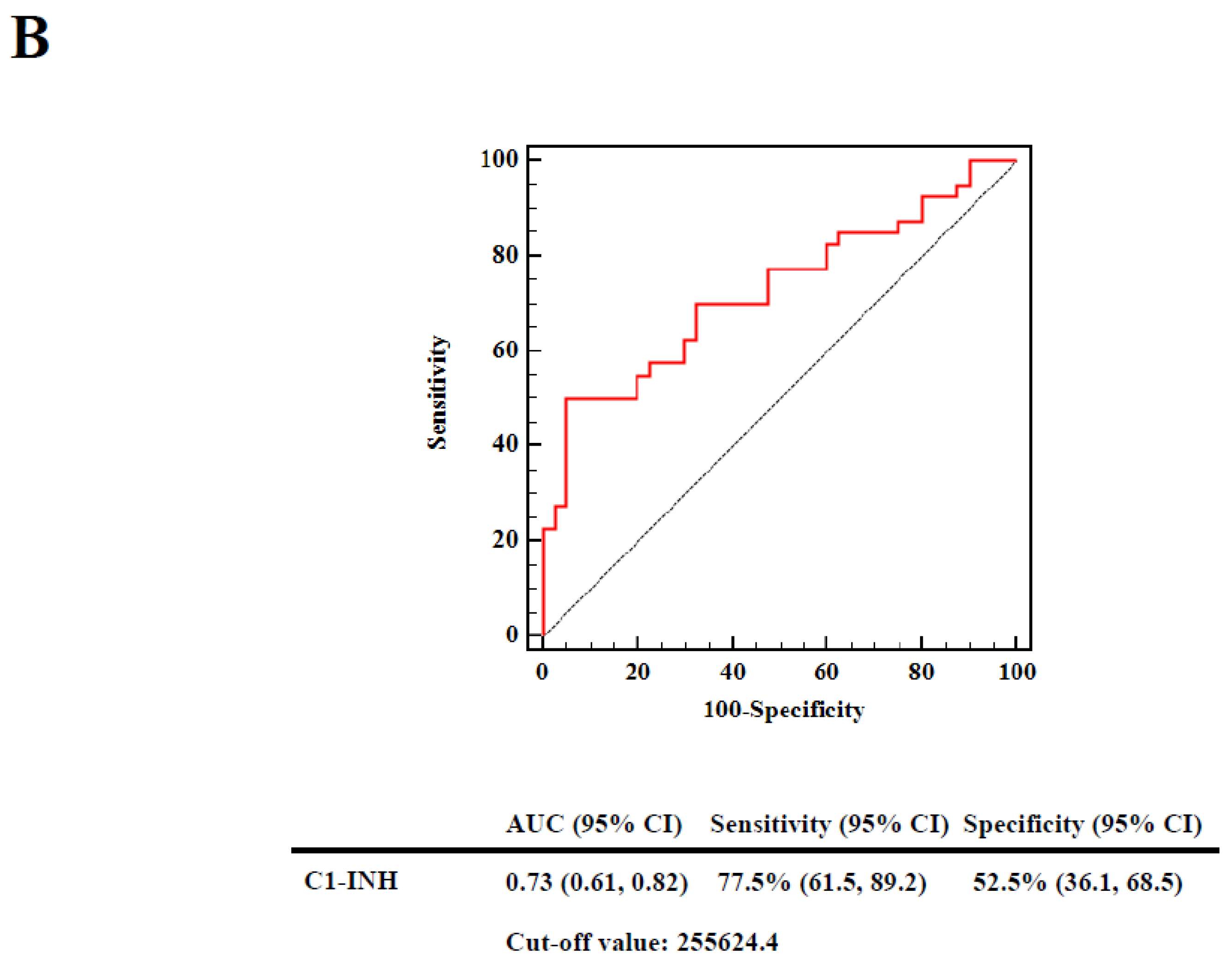
2.5. Associations of Decreased C1-INH Levels, Elevated Ac-Protein Adduct Levels, and Reduced Autoantibody Titers against C1-INH367–385 and C1-INH367–385 Ac Peptides with Risks in Patients with SLE
3. Discussion
4. Materials and Methods
4.1. Patients and Controls
4.2. In-Gel Digestion and PTM Identification Using Nano-LC-MS/MS
4.3. IP and Western Blotting
4.4. Measurement of Autoantibodies against C1-INH367–385 and C1-INH367–385 Ac Peptides
4.5. Determination of Serum Ac-Protein Adducts
4.6. Statistical Analyses
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
Abbreviations
References
- Yeh, K.W.; Yu, C.H.; Chan, P.C.; Horng, J.T.; Huang, J.L. Burden of systemic lupus erythematosus in taiwan: A population-based survey. Rheumatol. Int. 2013, 33, 1805–1811. [Google Scholar] [CrossRef]
- Adhami, E. Calculating the etiology of systemic lupus erythematosus. Med. Hypotheses 2004, 62, 237–246. [Google Scholar] [CrossRef]
- Parks, C.G.; de Souza Espindola Santos, A.; Barbhaiya, M.; Costenbader, K.H. Understanding the role of environmental factors in the development of systemic lupus erythematosus. Best Pract. Res. Clin. Rheumatol. 2017, 31, 306–320. [Google Scholar] [CrossRef] [PubMed]
- Lewis, M.J.; Jawad, A.S. The effect of ethnicity and genetic ancestry on the epidemiology, clinical features and outcome of systemic lupus erythematosus. Rheumatology (Oxford) 2017, 56, i67–i77. [Google Scholar] [CrossRef]
- Mistry, P.; Kaplan, M.J. Cell death in the pathogenesis of systemic lupus erythematosus and lupus nephritis. Clin. Immunol. 2017, 185, 59–73. [Google Scholar] [CrossRef]
- Rutter-Locher, Z.; Smith, T.O.; Giles, I.; Sofat, N. Association between systemic lupus erythematosus and periodontitis: A systematic review and meta-analysis. Front. Immunol. 2017, 8, 1295. [Google Scholar] [CrossRef]
- Cozzani, E.; Drosera, M.; Gasparini, G.; Parodi, A. Serology of lupus erythematosus: Correlation between immunopathological features and clinical aspects. Autoimmune Dis. 2014, 2014, 321359. [Google Scholar] [CrossRef]
- Alzolibani, A.A.; Al Robaee, A.A.; Al-Shobaili, H.A.; Rasheed, Z. 4-hydroxy-2-nonenal modified histone-h2a: A possible antigenic stimulus for systemic lupus erythematosus autoantibodies. Cell Immunol. 2013, 284, 154–162. [Google Scholar] [CrossRef] [PubMed]
- Khan, M.A.; Dixit, K.; Moinuddin; Arif, Z.; Alam, K. Studies on peroxynitrite-modified h1 histone: Implications in systemic lupus erythematosus. Biochimie 2014, 97, 104–113. [Google Scholar] [CrossRef] [PubMed]
- Ben Mansour, R.; Lassoued, S.; Elgaied, A.; Haddouk, S.; Marzouk, S.; Bahloul, Z.; Masmoudi, H.; Attia, H.; Aifa, M.S.; Fakhfakh, F. Enhanced reactivity to malondialdehyde-modified proteins by systemic lupus erythematosus autoantibodies. Scand. J. Rheumatol. 2010, 39, 247–253. [Google Scholar] [CrossRef]
- Doyle, H.A.; Aswad, D.W.; Mamula, M.J. Autoimmunity to isomerized histone h2b in systemic lupus erythematosus. Autoimmunity 2013, 46, 6–13. [Google Scholar] [CrossRef] [PubMed]
- Dieker, J.; Berden, J.H.; Bakker, M.; Briand, J.P.; Muller, S.; Voll, R.; Sjowall, C.; Herrmann, M.; Hilbrands, L.B.; van der Vlag, J. Autoantibodies against modified histone peptides in sle patients are associated with disease activity and lupus nephritis. PLoS ONE 2016, 11, e0165373. [Google Scholar] [CrossRef]
- Ziegelasch, M.; van Delft, M.A.; Wallin, P.; Skogh, T.; Magro-Checa, C.; Steup-Beekman, G.M.; Trouw, L.A.; Kastbom, A.; Sjowall, C. Antibodies against carbamylated proteins and cyclic citrullinated peptides in systemic lupus erythematosus: Results from two well-defined european cohorts. Arthritis Res. Ther. 2016, 18, 289. [Google Scholar] [CrossRef] [PubMed]
- van Bavel, C.C.; Dieker, J.W.; Kroeze, Y.; Tamboer, W.P.; Voll, R.; Muller, S.; Berden, J.H.; van der Vlag, J. Apoptosis-induced histone h3 methylation is targeted by autoantibodies in systemic lupus erythematosus. Ann. Rheum. Dis. 2011, 70, 201–207. [Google Scholar] [CrossRef] [PubMed]
- Khan, F.; Ali, R. Antibodies against nitric oxide damaged poly l-tyrosine and 3-nitrotyrosine levels in systemic lupus erythematosus. J. Biochem. Mol. Biol. 2006, 39, 189–196. [Google Scholar] [CrossRef] [PubMed]
- Dieker, J.W.; Fransen, J.H.; van Bavel, C.C.; Briand, J.P.; Jacobs, C.W.; Muller, S.; Berden, J.H.; van der Vlag, J. Apoptosis-induced acetylation of histones is pathogenic in systemic lupus erythematosus. Arthritis Rheum. 2007, 56, 1921–1933. [Google Scholar] [CrossRef]
- Jazwinska, E.C.; Gatenby, P.A.; Dunckley, H.; Serjeantson, S.W. C1 inhibitor functional deficiency in systemic lupus erythematosus (sle). Clin. Exp. Immunol. 1993, 92, 268–273. [Google Scholar] [CrossRef] [PubMed]
- Nettis, E.; Colanardi, M.C.; Loria, M.P.; Vacca, A. Acquired c1-inhibitor deficiency in a patient with systemic lupus erythematosus: A case report and review of the literature. Eur. J. Clin. Invest. 2005, 35, 781–784. [Google Scholar] [CrossRef]
- Nakamura, S.; Yoshinari, M.; Saku, Y.; Hirakawa, K.; Miishima, C.; Murai, K.; Tokiyama, K.; Fujishima, M. Acquired c1 inhibitor deficiency associated with systemic lupus erythematosus affecting the central nervous system. Ann. Rheum Dis. 1991, 50, 713–716. [Google Scholar] [CrossRef]
- Davis, A.E., 3rd; Mejia, P.; Lu, F. Biological activities of c1 inhibitor. Mol. Immunol. 2008, 45, 4057–4063. [Google Scholar] [CrossRef]
- Meszaros, T.; Fust, G.; Farkas, H.; Jakab, L.; Temesszentandrasi, G.; Nagy, G.; Kiss, E.; Gergely, P.; Zeher, M.; Griger, Z.; et al. C1-inhibitor autoantibodies in sle. Lupus 2010, 19, 634–638. [Google Scholar] [CrossRef] [PubMed]
- Mandle, R.; Baron, C.; Roux, E.; Sundel, R.; Gelfand, J.; Aulak, K.; Davis, A.E., 3rd; Rosen, F.S.; Bing, D.H. Acquired c1 inhibitor deficiency as a result of an autoantibody to the reactive center region of c1 inhibitor. J. Immunol. 1994, 152, 4680–4685. [Google Scholar] [PubMed]
- Drazic, A.; Myklebust, L.M.; Ree, R.; Arnesen, T. The world of protein acetylation. Biochim. Biophys Acta 2016, 1864, 1372–1401. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gil, J.; Ramirez-Torres, A.; Encarnacion-Guevara, S. Lysine acetylation and cancer: A proteomics perspective. J. Proteomics 2017, 150, 297–309. [Google Scholar] [CrossRef] [PubMed]
- Spange, S.; Wagner, T.; Heinzel, T.; Kramer, O.H. Acetylation of non-histone proteins modulates cellular signalling at multiple levels. Int. J. Biochem. Cell Biol. 2009, 41, 185–198. [Google Scholar] [CrossRef] [PubMed]
- Davis, A.E., 3rd. Structure and function of c1 inhibitor. Behring Inst. Mitt. 1989, 142–150. [Google Scholar]
- Wallace, E.M.; Perkins, S.J.; Sim, R.B.; Willis, A.C.; Feighery, C.; Jackson, J. Degradation of c1-inhibitor by plasmin: Implications for the control of inflammatory processes. Mol. Med. 1997, 3, 385–396. [Google Scholar] [CrossRef]
- Brown, E.W.; Ravindran, S.; Patston, P.A. The reaction between plasmin and c1-inhibitor results in plasmin inhibition by the serpin mechanism. Blood Coagul Fibrinolysis 2002, 13, 711–714. [Google Scholar] [CrossRef]
- Wagenaar-Bos, I.G.; Hack, C.E. Structure and function of c1-inhibitor. Immunol. Allergy Clin. North. Am. 2006, 26, 615–632. [Google Scholar] [CrossRef]
- Ansari, A.A.; Kidwai, S.A.; Salahuddin, A. Acetylation of amino groups and its effect on the conformation and immunological activity of obalbumin. J. Biol. Chem. 1975, 250, 1625–1632. [Google Scholar]
- Dhillon, P.K.; Adams, M.J. Thrombosis in systemic lupus erythematosus: Role of impaired fibrinolysis. Semin Thromb Hemost 2013, 39, 434–440. [Google Scholar] [CrossRef]
- van Bavel, C.C.; Dieker, J.; Muller, S.; Briand, J.P.; Monestier, M.; Berden, J.H.; van der Vlag, J. Apoptosis-associated acetylation on histone h2b is an epitope for lupus autoantibodies. Mol. Immunol. 2009, 47, 511–516. [Google Scholar] [CrossRef]
- Alaskhar Alhamwe, B.; Khalaila, R.; Wolf, J.; von Bulow, V.; Harb, H.; Alhamdan, F.; Hii, C.S.; Prescott, S.L.; Ferrante, A.; Renz, H.; et al. Histone modifications and their role in epigenetics of atopy and allergic diseases. Allergy Asthma Clin. Immunol. 2018, 14, 39. [Google Scholar] [CrossRef]
- Cheung, W.L.; Ajiro, K.; Samejima, K.; Kloc, M.; Cheung, P.; Mizzen, C.A.; Beeser, A.; Etkin, L.D.; Chernoff, J.; Earnshaw, W.C.; et al. Apoptotic phosphorylation of histone h2b is mediated by mammalian sterile twenty kinase. Cell 2003, 113, 507–517. [Google Scholar] [CrossRef]
- Suzuki, T.; Burlingame, R.W.; Casiano, C.A.; Boey, M.L.; Rubin, R.L. Antihistone antibodies in systemic lupus erythematosus: Assay dependency and effects of ubiquitination and serum DNA. J. Rheumatol. 1994, 21, 1081–1091. [Google Scholar]
- Dieker, J.; Muller, S. Epigenetic histone code and autoimmunity. Clin. Rev. Allergy Immunol. 2010, 39, 78–84. [Google Scholar] [CrossRef]
- Lloyd, K.A.; Wigerblad, G.; Sahlstrom, P.; Garimella, M.G.; Chemin, K.; Steen, J.; Titcombe, P.J.; Marklein, B.; Zhou, D.; Stalesen, R.; et al. Differential acpa binding to nuclear antigens reveals a pad-independent pathway and a distinct subset of acetylation cross-reactive autoantibodies in rheumatoid arthritis. Front. Immunol. 2018, 9, 3033. [Google Scholar] [CrossRef]
- Varga, L.; Szeplaki, G.; Visy, B.; Fust, G.; Harmat, G.; Miklos, K.; Nemeth, J.; Cervenak, L.; Karadi, I.; Farkas, H. C1-inhibitor (c1-inh) autoantibodies in hereditary angioedema. Strong correlation with the severity of disease in c1-inh concentrate naive patients. Mol. Immunol. 2007, 44, 1454–1460. [Google Scholar] [CrossRef]
- Alsenz, J.; Bork, K.; Loos, M. Autoantibody-mediated acquired deficiency of c1 inhibitor. N. Engl. J. Med. 1987, 316, 1360–1366. [Google Scholar] [CrossRef]
- He, S.; Tsang, S.; North, J.; Chohan, N.; Sim, R.B.; Whaley, K. Epitope mapping of c1 inhibitor autoantibodies from patients with acquired c1 inhibitor deficiency. J. Immunol. 1996, 156, 2009–2013. [Google Scholar]
- Hodkinson, J.P.; Bangs, C.; Wartenberg-Demand, A.; Bauhofer, A.; Langohr, P.; Buckland, M.S.; Guzman, D.; Yong, P.F.K.; Kiani-Alikhan, S. Low iga and igm is associated with a higher prevalence of bronchiectasis in primary antibody deficiency. J. Clin. Immunol. 2017, 37, 329–331. [Google Scholar] [CrossRef]
- Traverso, N.; Patriarca, S.; Balbis, E.; Furfaro, A.L.; Cottalasso, D.; Pronzato, M.A.; Carlier, P.; Botta, F.; Marinari, U.M.; Fontana, L. Anti malondialdehyde-adduct immunological response as a possible marker of successful aging. Exp. Gerontology 2003, 38, 1129–1135. [Google Scholar] [CrossRef]
- Nowakowska-Plaza, A.; Potaczek, D.P.; Gluszko, P.; Undas, A. Antibodies to n-homocysteinylated albumin and haemoglobin in patients with rheumatoid arthritis: A potential new marker of disease severity. Scand. J. Rheumatol 2014, 43, 17–21. [Google Scholar] [CrossRef]
- Padjas, A.; Undas, A.; Swadzba, J.; Musial, J. Antibodies to n-homocysteinylated albumin in patients with systemic lupus erythematosus. Pol. Arch. Med. Wewn 2007, 117, 20–25. [Google Scholar] [CrossRef]
- Liao, C.C.; Chang, Y.S.; Cheng, C.W.; Chi, W.M.; Tsai, K.L.; Chen, W.J.; Kung, T.S.; Tai, C.C.; Lin, Y.F.; Lin, H.T.; et al. Corrigendum to “isotypes of autoantibodies against differentially expressed novel malondialdehyde-modifiedpeptide adducts in serum of taiwanese women with rheumatoid arthritis”. J. Proteomics 2019, 194, 226. [Google Scholar] [CrossRef]
- Pruijn, G.J. Citrullination and carbamylation in the pathophysiology of rheumatoid arthritis. Front. Immunol. 2015, 6, 192. [Google Scholar] [CrossRef]
- Trouw, L.A.; Rispens, T.; Toes, R.E.M. Beyond citrullination: Other post-translational protein modifications in rheumatoid arthritis. Nat. Rev. Rheumatol 2017, 13, 331–339. [Google Scholar] [CrossRef]
- Figueiredo, C.P.; Bang, H.; Cobra, J.F.; Englbrecht, M.; Hueber, A.J.; Haschka, J.; Manger, B.; Kleyer, A.; Reiser, M.; Finzel, S.; et al. Antimodified protein antibody response pattern influences the risk for disease relapse in patients with rheumatoid arthritis tapering disease modifying antirheumatic drugs. Ann. Rheum Dis. 2017, 76, 399–407. [Google Scholar] [CrossRef]
- Aletaha, D.; Neogi, T.; Silman, A.J.; Funovits, J.; Felson, D.T.; Bingham, C.O., 3rd; Birnbaum, N.S.; Burmester, G.R.; Bykerk, V.P.; Cohen, M.D.; et al. 2010 rheumatoid arthritis classification criteria: An american college of rheumatology/european league against rheumatism collaborative initiative. Arthritis Rheumatism 2010, 62, 2569–2581. [Google Scholar]
- Arnett, F.C.; Edworthy, S.M.; Bloch, D.A.; McShane, D.J.; Fries, J.F.; Cooper, N.S.; Healey, L.A.; Kaplan, S.R.; Liang, M.H.; Luthra, H.S.; et al. The american rheumatism association 1987 revised criteria for the classification of rheumatoid arthritis. Arthritis Rheumatism 1988, 31, 315–324. [Google Scholar] [Green Version]
- Hochberg, M.C. Updating the american college of rheumatology revised criteria for the classification of systemic lupus erythematosus. Arthritis Rheumatism 1997, 40, 1725. [Google Scholar]
- Chang, Y.S.; Pan, C.H.; Chang, C.C.; Tsai, K.L.; Chou, H.W.; Chen, J.H.; Lin, S.H.; Lu, Y.Y.; Tai, C.C.; Lin, Y.F.; et al. Low levels of igg recognizing the alpha-1-antitrypsin peptide and its association with taiwanese women with primary sjogren’s syndrome. Int. J. Mol. Sci. 2017, 18, 2750. [Google Scholar] [CrossRef]
- Uen, Y.H.; Lin, K.Y.; Sun, D.P.; Liao, C.C.; Hsieh, M.S.; Huang, Y.K.; Chen, Y.W.; Huang, P.H.; Chen, W.J.; Tai, C.C.; et al. Comparative proteomics, network analysis and post-translational modification identification reveal differential profiles of plasma con a-bound glycoprotein biomarkers in gastric cancer. J. Proteomics 2013, 83, 197–213. [Google Scholar] [CrossRef]
- Eng, J.K.; McCormack, A.L.; Yates, J.R. An approach to correlate tandem mass spectral data of peptides with amino acid sequences in a protein database. J. Am. Soc. Mass. Spectrom. 1994, 5, 976–989. [Google Scholar] [CrossRef] [Green Version]
Sample Availability: Not available. |







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Risk Factors | Cut-off | HC | SLE | Univariate Logistic Regression Model a | Age-Adjusted Logistic Regression Model | |||||
|---|---|---|---|---|---|---|---|---|---|---|
| N = 50 | N = 54 | ORs (95% CI) | p-Value | Power | ORs (95% CI) | p-Value | Power | |||
| C1-INH | > | 255624.4 | 21 | 10 | 1 | 0.013 | 0.697 | 1 | 0.015 | 0.684 |
| ≤ | 255624.4 | 19 | 30 | 3.315 (1.286, 8.548) | 3.250 (1.256, 8.409) | |||||
| Acetylation-protein adduct | ≤ | 0.299 | 25 | 16 | 1 | 0.035 | 0.568 | 1 | 0.035 | 0.570 |
| > | 0.299 | 25 | 38 | 2.375 (1.062, 5.314) | 2.381 (1.063, 5.334) | |||||
| IgM anti-C1-INH367-385 | ≥ | 0.762 | 38 | 24 | 1 | 0.001 | 0.663 | 1 | <0.001 | 0.737 |
| < | 0.762 | 12 | 30 | 3.958 (1.705, 9.189) | 4.725 (1.929, 11.573) | |||||
| IgG anti-C1-INH367-385 | ≥ | 2.026 | 41 | 33 | 1 | 0.021 | 0.403 | 1 | 0.020 | 0.413 |
| < | 2.026 | 9 | 21 | 2.899 (1.172, 7.169) | 2.957 (1.190, 7.349) | |||||
| IgA anti-C1-INH367-385 | < | 2.718 | 37 | 33 | 1 | 0.164 | 0.231 | 1 | 0.183 | 0.219 |
| ≥ | 2.718 | 13 | 21 | 1.811 (0.785, 4.178) | 1.771 (0.763, 4.108) | |||||
| IgM anti-C1-INH367-385 Ac | ≥ | 0.855 | 27 | 13 | 1 | 0.002 | 0.853 | 1 | 0.001 | 0.892 |
| < | 0.855 | 23 | 41 | 3.702 (1.605, 8.538) | 4.089 (1.725, 9.694) | |||||
| IgG anti-C1-INH367-385 Ac | ≥ | 1.501 | 43 | 39 | 1 | 0.091 | 0.250 | 1 | 0.090 | 0.252 |
| < | 1.501 | 7 | 15 | 2.362 (0.872, 6.397) | 2.371 (0.874, 6.432) | |||||
| IgA anti-C1-INH367-385 Ac | ≥ | 0.465 | 36 | 17 | 1 | <0.001 | 0.850 | 1 | <0.001 | 0.848 |
| < | 0.465 | 14 | 37 | 5.597 (2.409, 13.005) | 5.566 (2.380, 13.016) | |||||
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Tsai, K.-L.; Liao, C.-C.; Chang, Y.-S.; Huang, C.-W.; Huang, Y.-C.; Chen, J.-H.; Lin, S.-H.; Tai, C.-C.; Lin, Y.-F.; Lin, C.-Y. Low Levels of IgM and IgA Recognizing Acetylated C1-Inhibitor Peptides Are Associated with Systemic Lupus Erythematosus in Taiwanese Women. Molecules 2019, 24, 1645. https://doi.org/10.3390/molecules24091645
Tsai K-L, Liao C-C, Chang Y-S, Huang C-W, Huang Y-C, Chen J-H, Lin S-H, Tai C-C, Lin Y-F, Lin C-Y. Low Levels of IgM and IgA Recognizing Acetylated C1-Inhibitor Peptides Are Associated with Systemic Lupus Erythematosus in Taiwanese Women. Molecules. 2019; 24(9):1645. https://doi.org/10.3390/molecules24091645
Chicago/Turabian StyleTsai, Kai-Leun, Chen-Chung Liao, Yu-Sheng Chang, Ching-Wen Huang, Yu-Chu Huang, Jin-Hua Chen, Sheng-Hong Lin, Chih-Chun Tai, Yi-Fang Lin, and Ching-Yu Lin. 2019. "Low Levels of IgM and IgA Recognizing Acetylated C1-Inhibitor Peptides Are Associated with Systemic Lupus Erythematosus in Taiwanese Women" Molecules 24, no. 9: 1645. https://doi.org/10.3390/molecules24091645