
Arginine in the FARM and SARM: A Role in Chain-Length Determination for Arginine in the Aspartate-Rich Motifs of Isoprenyl Diphosphate Synthases from Mycobacterium tuberculosis †

 ,
,
Abstract
:1. Introduction
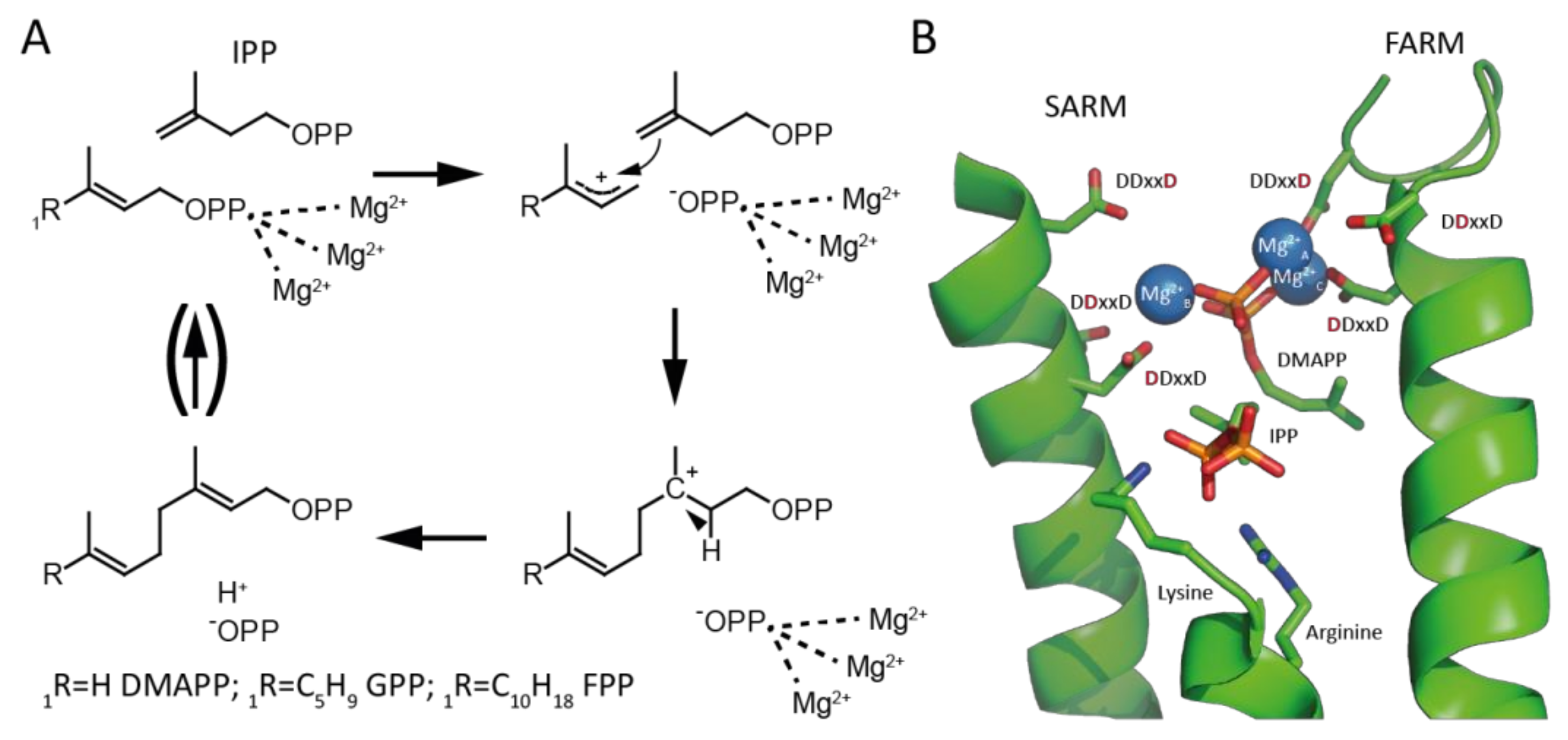
2. Results
3. Materials and Methods
3.1. General Reagents
3.2. Sequence Analysis and Alignment
3.3. Site-Directed Mutagenesis
3.4. Protein Expression and Assay
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Gershenzon, J.; Dudareva, N. The function of terpene natural products in the natural world. Nat. Chem. Biol. 2007, 3, 408–414. [Google Scholar] [CrossRef] [PubMed]
- Pichersky, E.; Raguso, R.A. Why do plants produce so many terpenoid compounds? New Phytol. 2016. [Google Scholar] [CrossRef] [PubMed]
- Buckingham, J. Dictionary of Natural Products (on-line web edition); Chapman & Hall/CRC Press: London, UK, 2007. [Google Scholar]
- Liang, P.-H.; Ko, T.-P.; Wang, A.H.-J. Structure, mechanism and function of prenyltransferases. Eur. J. Biochem. 2002, 269, 3339–3354. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chen, A.P.; Chang, S.Y.; Lin, Y.C.; Sun, Y.S.; Chen, C.T.; Wang, A.H.; Liang, P.H. Substrate and product specificities of cis-type undecaprenyl pyrophosphate synthase. Biochem. J. 2005, 386, 169–176. [Google Scholar] [CrossRef] [PubMed]
- Kaur, D.; Brennan, P.J.; Crick, D.C. Decaprenyl diphosphate synthesis in Mycobacterium tuberculosis. J. Bacteriol. 2004, 186, 7564–7570. [Google Scholar] [CrossRef] [PubMed]
- Swiezewska, E.; Danikiewicz, W. Polyisoprenoids: Structure, biosynthesis and function. Prog. Lipid Res. 2005, 44, 235–258. [Google Scholar] [CrossRef] [PubMed]
- Akhtar, T.A.; Matsuba, Y.; Schauvinhold, I.; Yu, G.; Lees, H.A.; Klein, S.E.; Pichersky, E. The tomato cisprenyltransferase gene family. Plant J. 2013, 73, 640–652. [Google Scholar] [CrossRef] [PubMed]
- Poulter, C.D.; Rilling, H.C. Prenyltransferase: the mechanism of the reaction. Biochemistry 1976, 15, 1079–1083. [Google Scholar] [CrossRef] [PubMed]
- Poulter, C.D.; Satterwhite, D.M.; Rilling, H.C. Prenyltransferase—Mechanism Of Reaction. J. Am. Chem. Soc. 1976, 98, 3376–3377. [Google Scholar] [CrossRef] [PubMed]
- Aaron, J.A.; Christianson, D.W. Trinuclear Metal Clusters in Catalysis by Terpenoid Synthases. Pure Appl. Chem. 2010, 82, 1585–1597. [Google Scholar] [CrossRef] [PubMed]
- Schulbach, M.C.; Brennan, P.J.; Crick, D.C. Identification of a Short (C15) ChainZ-Isoprenyl Diphosphate Synthase and a Homologous Long (C50) Chain Isoprenyl Diphosphate Synthase inMycobacterium tuberculosis. J. Biol. Chem. 2000, 275, 22876–22881. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Schulbach, M.C.; Mahapatra, S.; Macchia, M.; Barontini, S.; Papi, C.; Minutolo, F.; Bertini, S.; Brennan, P.J.; Crick, D.C. Purification, Enzymatic Characterization, and Inhibition of theZ-Farnesyl Diphosphate Synthase from Mycobacterium tuberculosis. J. Biol. Chem. 2001, 276, 11624–11630. [Google Scholar] [CrossRef] [PubMed]
- Mann, F.M.; Thomas, J.A.; Peters, R.J. Rv0989c encodes a novel (E)-geranyl diphosphate synthase facilitating decaprenyl diphosphate biosynthesis in Mycobacterium tuberculosis. FEBS Lett. 2011, 585, 549–554. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Young, D.; Moody, D. In vivo biosynthesis of terpene nucleosides provides unique chemical markers of Mycobacterium tuberculosis infection. Chem. Bio. 2015, 22, 516–526. [Google Scholar] [CrossRef] [PubMed]
- Mann, F.M.; Prisic, S.; Hu, H.; Xu, M.; Coates, R.M.; Peters, R.J. Characterization and inhibition of a class II diterpene cyclase from Mycobacterium tuberculosis: implications for tuberculosis. J. Biol. Chem. 2009, 284, 23574–23579. [Google Scholar] [CrossRef] [PubMed]
- Nakano, C.; Hoshino, T. Characterization of the Rv3377c gene product, a type-B diterpene cyclase, from the Mycobacterium tuberculosis H37 genome. Chembiochem 2009, 10, 2060–2071. [Google Scholar] [CrossRef] [PubMed]
- Layre, E.; Lee, H.J.; Young, D.C.; Martinot, A.J.; Buter, J.; Minnaard, A.J.; Annand, J.W.; Fortune, S.M.; Snider, B.B. Molecular profiling of Mycobacterium tuberculosis identifies tuberculosinyl nucleoside products of the virulence-associated enzyme Rv3378c. Proc. National Acad. Sci. 2014, 111, 2978–2983. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pethe, K.; Swenson, D.L.; Alonso, S.; Anderson, J.; Wang, C.; Russell, D.G. Isolation of Mycobacterium tuberculosis mutants defective in the arrest of phagosome maturation. Proc. National Acad. Sci. 2004, 101, 13642–13647. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mann, F.M.; Xu, M.; Chen, X.; Fulton, D.B.; Russell, D.G.; Peters, R.J. Edaxadiene: A new bioactive diterpene from Mycobacterium tuberculosis. J. Am. Chem. Soc. 2009, 131, 15726–15727. [Google Scholar] [CrossRef] [PubMed]
- Mann, F.M.; Xu, M.; Davenport, E.K.; Peters, R.J. Functional characterization and evolution of the isotuberculosinol operon in Mycobacterium tuberculosis and related Mycobacteria. Front Microbiol. 2012, 3, 368. [Google Scholar] [CrossRef] [PubMed]
- Ohnuma, S.; Narita, K.; Nakazawa, T.; Ishida, C.; Takeuchi, C.; Ohto, C.; Nishino, T. A role of the amino acid residue located on the fifth position before the first aspartate-rich motif of farnesyl diphosphate synthase on determination of the final product. J. Biol. Chem. 1996, 271, 30748–30754. [Google Scholar] [CrossRef] [PubMed]
- Joly, A.; Edwards, P.A. Effect of site-directed mutagenesis of conserved aspartate and arginine residues upon farnesyl diphosphate synthase activity. J. Biol. Chem. 1993, 268, 26983–26989. [Google Scholar] [PubMed]
- Dhiman, R.K.; Schulbach, M.C.; Mahapatra, S.; Baulard, A.R.; Vissa, V.; Brennan, P.J.; Crick, D.C. Identification of a novel class of omega,E,E-farnesyl diphosphate synthase from Mycobacterium tuberculosis. J. Lipid Res. 2004, 45, 1140–1147. [Google Scholar] [CrossRef] [PubMed]
- Kumar, S.; Stecher, G.; Tamura, K. MEGA7: Molecular Evolutionary Genetics Analysis Version 7.0 for Bigger Datasets. Mol. Biol. Evol. 2016, 33, 1870–1874. [Google Scholar] [CrossRef] [PubMed]
Sample Availability: Samples of the compounds (plasmids) are available from the authors, while the isoprenoids are commercially available. |



{kind=link}
{kind=link}
| Enzyme | GPP | FPP | GGPP |
|---|---|---|---|
| Rv0989c | 100 | 0 | 0 |
| Rv0989c:R92D | 76 | 24 | 0 |
| Rv0989c:R217D | 40 | 60 | 0 |
| Rv0989c:R92D/R217D | 30 | 70 | 0 |
| Rv0562 | 0 | 0 | 100 |
| Rv0562:D98R | 66 | 34 | 0 |
| Rv0562:D223R | 15 | 51 | 34 |
| Rv0562:D98R/D223R | 9 | 90 | 1 |
© 2018 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Nagel, R.; Thomas, J.A.; Adekunle, F.A.; Mann, F.M.; Peters, R.J. Arginine in the FARM and SARM: A Role in Chain-Length Determination for Arginine in the Aspartate-Rich Motifs of Isoprenyl Diphosphate Synthases from Mycobacterium tuberculosis. Molecules 2018, 23, 2546. https://doi.org/10.3390/molecules23102546
Nagel R, Thomas JA, Adekunle FA, Mann FM, Peters RJ. Arginine in the FARM and SARM: A Role in Chain-Length Determination for Arginine in the Aspartate-Rich Motifs of Isoprenyl Diphosphate Synthases from Mycobacterium tuberculosis. Molecules. 2018; 23(10):2546. https://doi.org/10.3390/molecules23102546
Chicago/Turabian StyleNagel, Raimund, Jill A. Thomas, Faith A. Adekunle, Francis M. Mann, and Reuben J. Peters. 2018. "Arginine in the FARM and SARM: A Role in Chain-Length Determination for Arginine in the Aspartate-Rich Motifs of Isoprenyl Diphosphate Synthases from Mycobacterium tuberculosis" Molecules 23, no. 10: 2546. https://doi.org/10.3390/molecules23102546