Protective Mechanism of Sulforaphane on Cadmium-Induced Sertoli Cell Injury in Mice Testis via Nrf2/ARE Signaling Pathway

 ,
,
Abstract
:1. Introduction
2. Experimental Design and Treatment
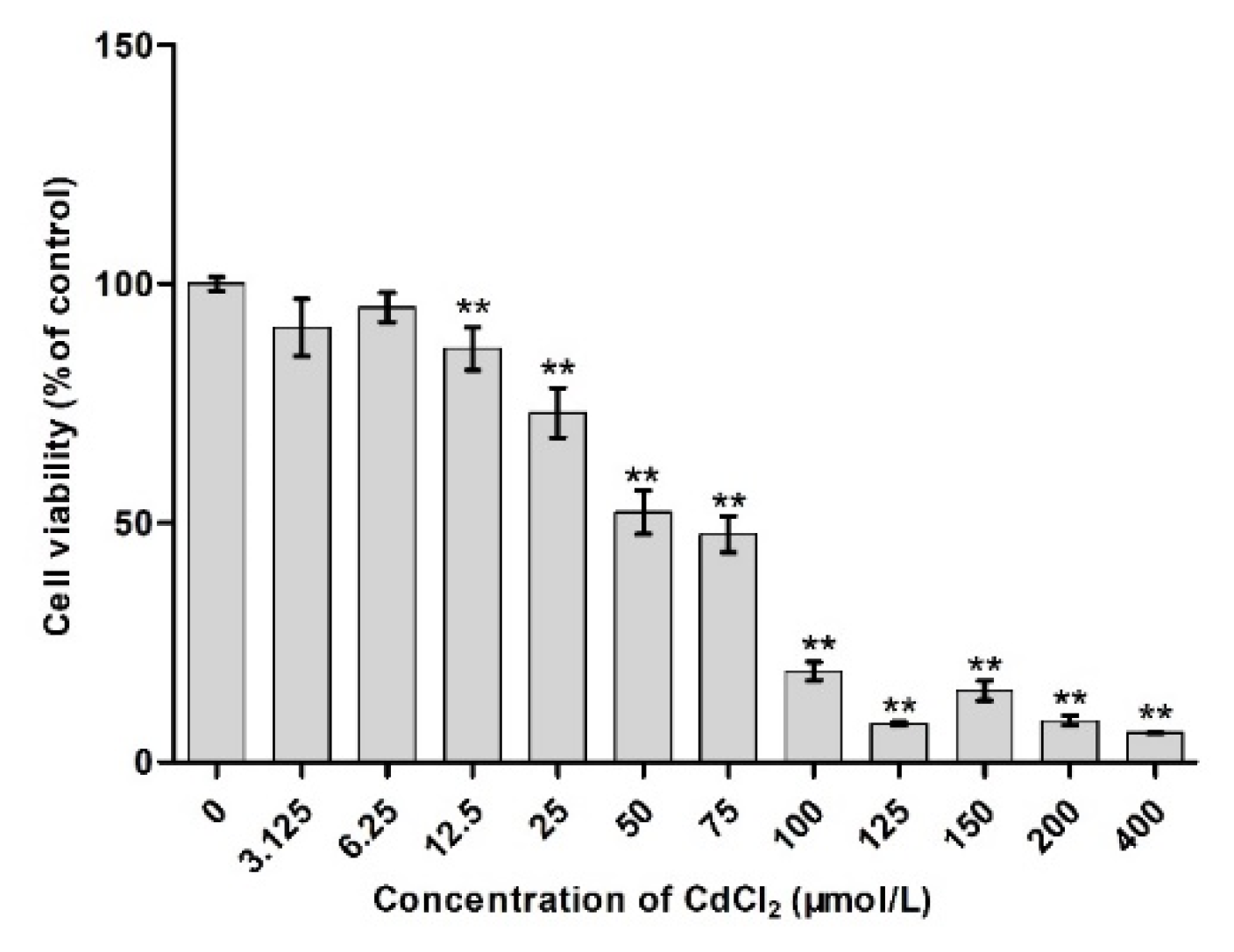
2.1. Effects of Different Concentrations of Cd on Cell Viability
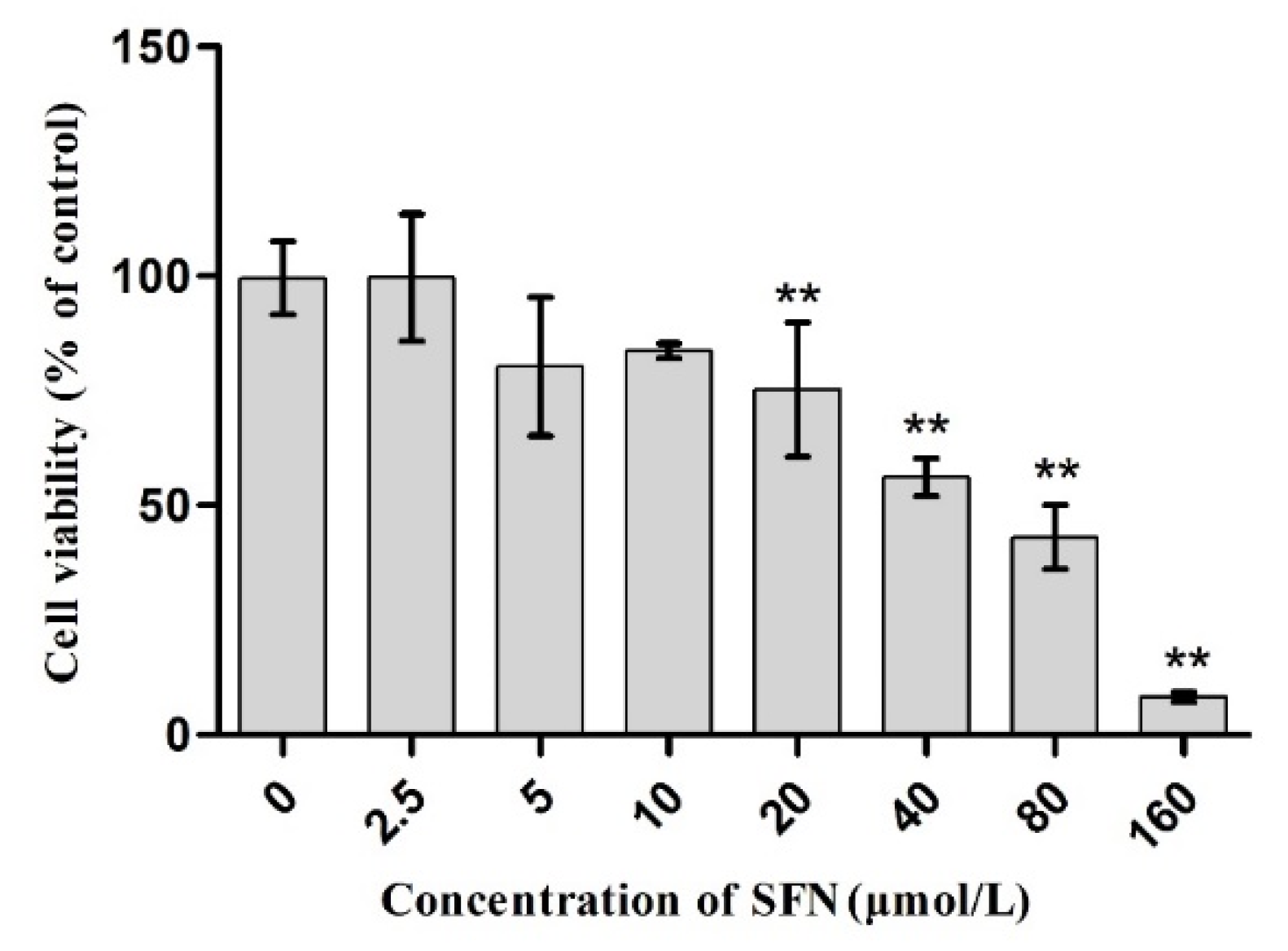
2.2. Effect of SFN on Cell Viability
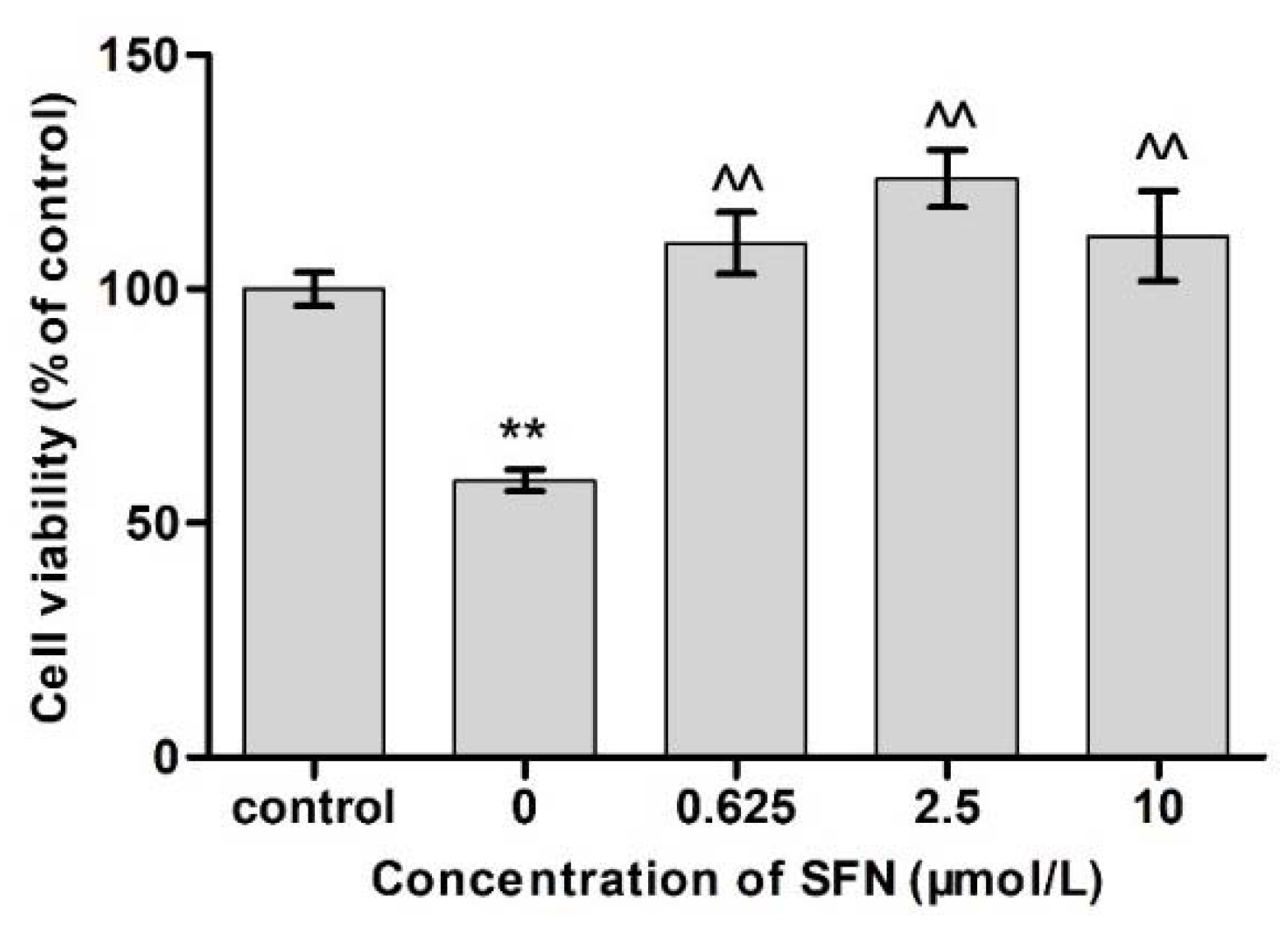
2.3. Effects of SFN and Cd on Cell Viability
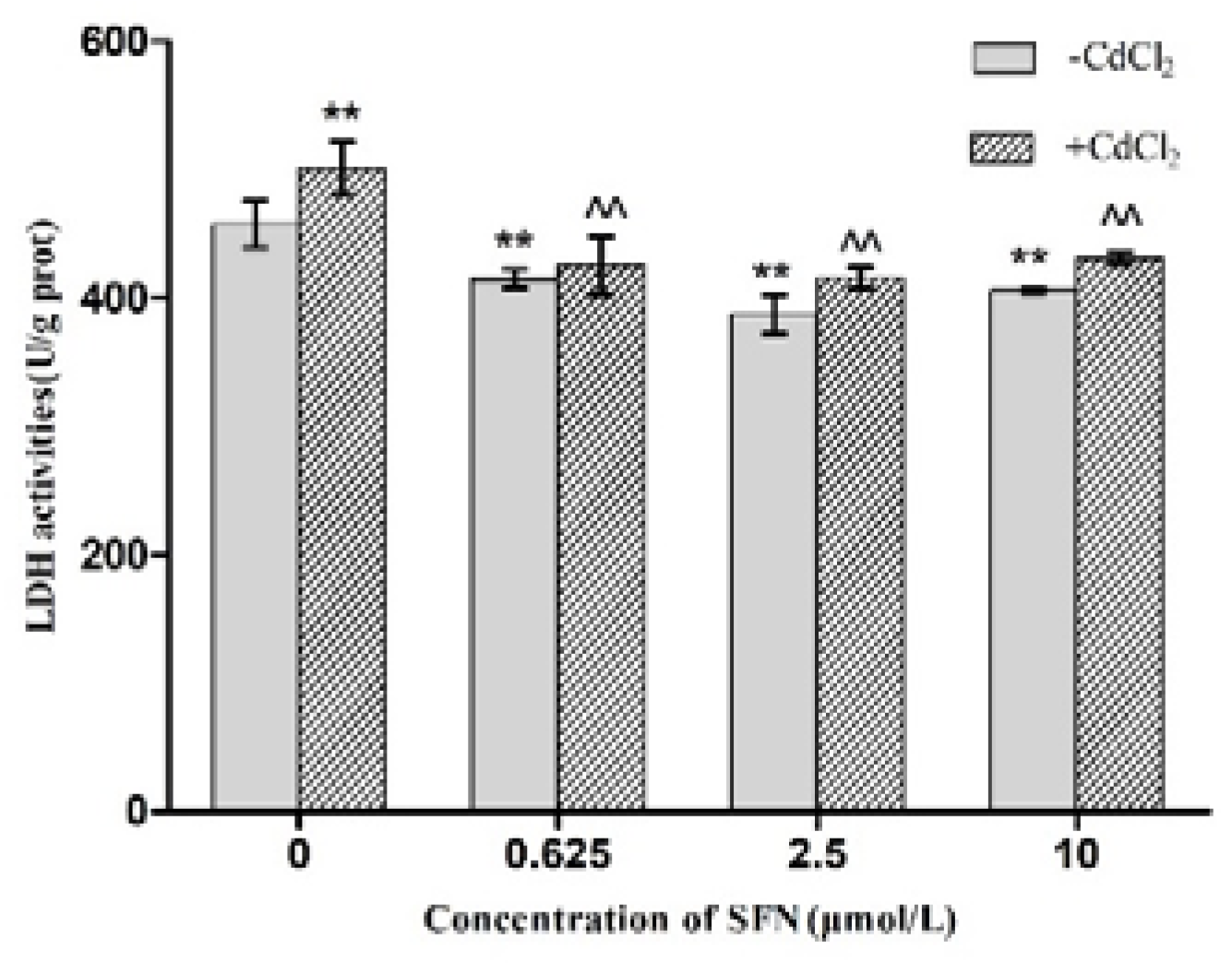
2.4. Effects of SFN and Cd on the Activity of LDH in Cells
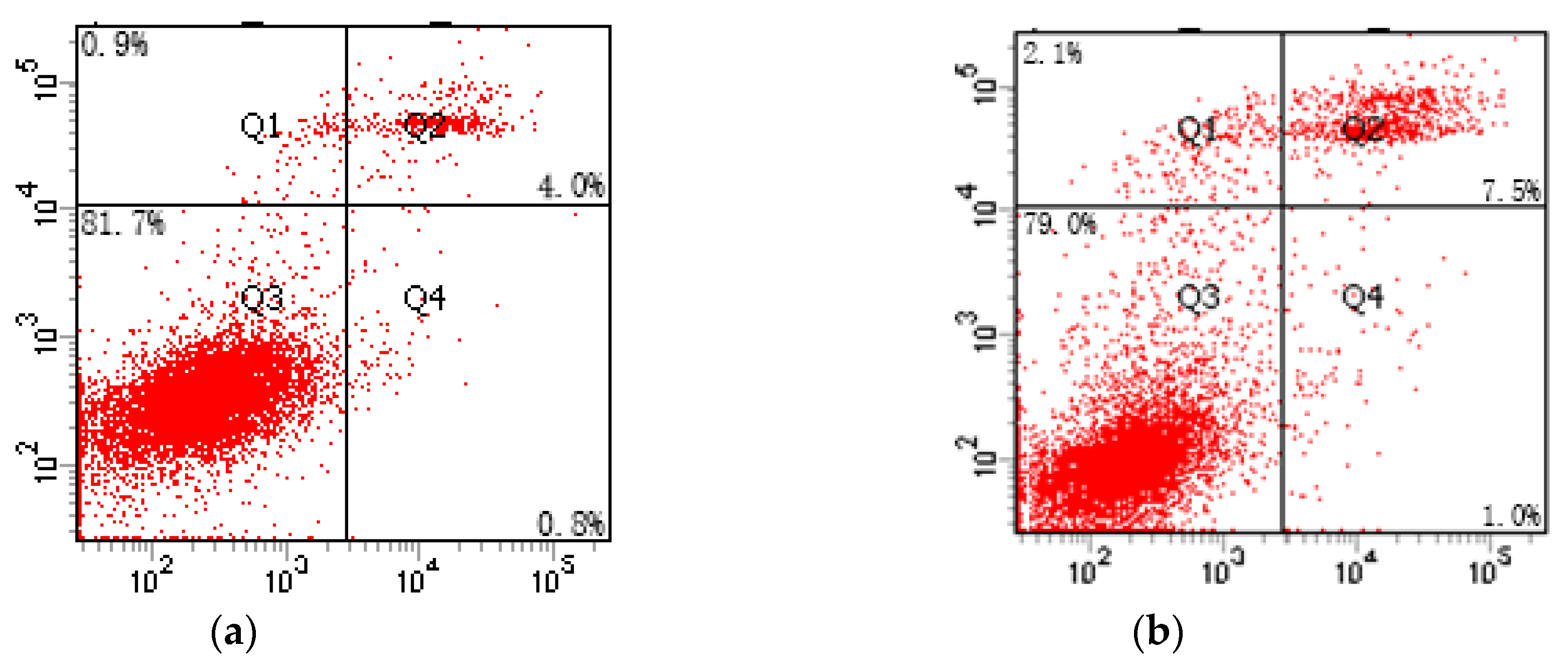
2.5. Apoptosis Rate Assessed by Flow Cytometry
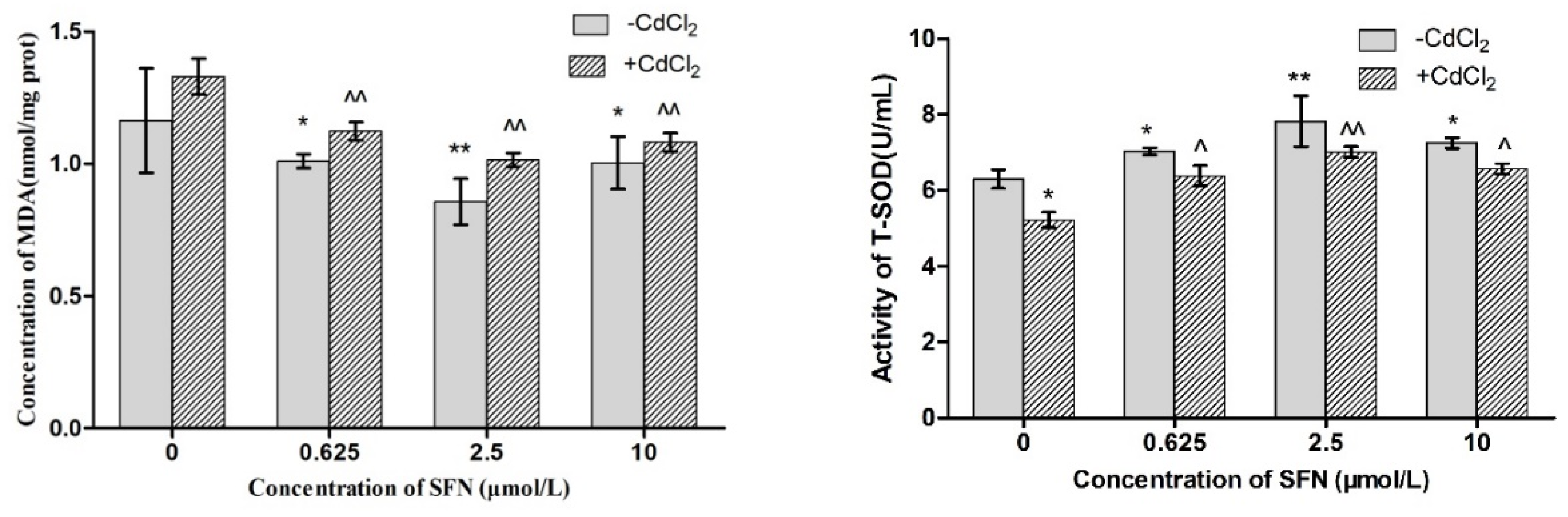
2.6. Detection of Oxidation Damage
2.7. Detection of mRNA Expression
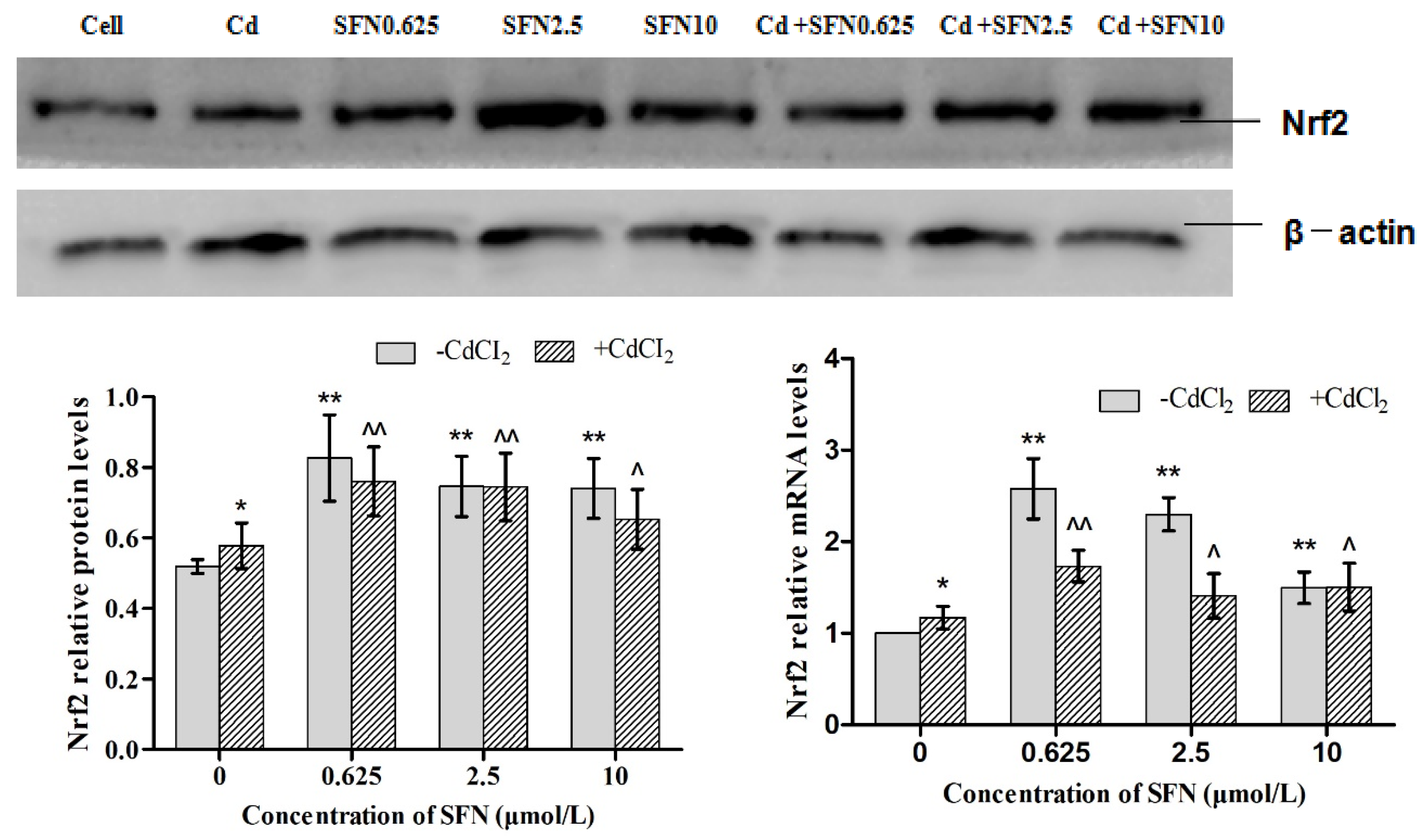
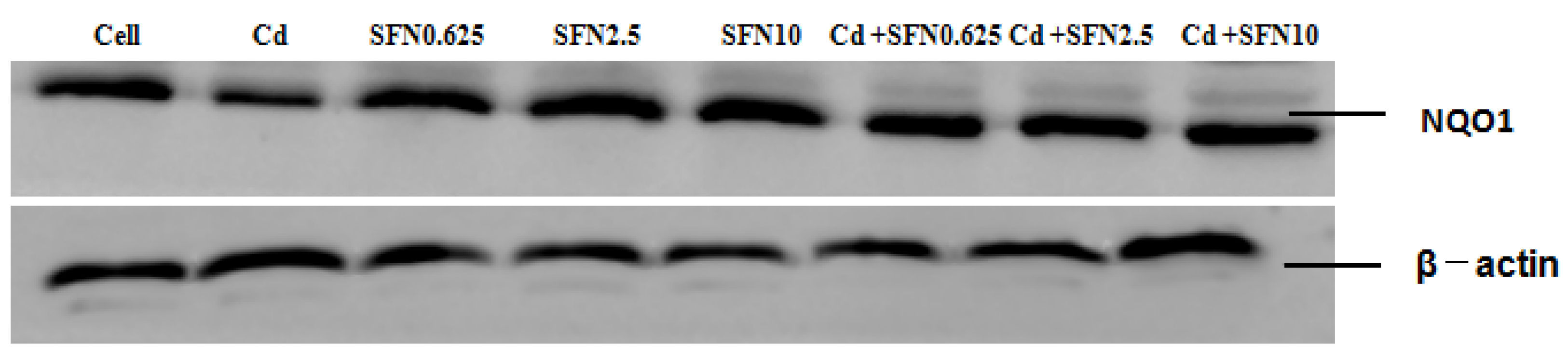
2.8. Western Blot
2.9. Statistical Analysis
3. Results
3.1. Survival Rate of Cd-Induced TM4 Cells
3.2. Effect of SFN on the Survival Rate of TM4 Cells
3.3. Effects of Different Concentrations of SFN on the Viability of Cd-Exposed TM4 Cells
3.4. Detection of TM4 Cell Activity by LDH
3.5. Effect of Cd-Induced SFN on the Apoptosis of TM4 Cells
3.6. Effects of SFN on Factors Related to Oxidative Stress in Cd-Induced TM4 Cells
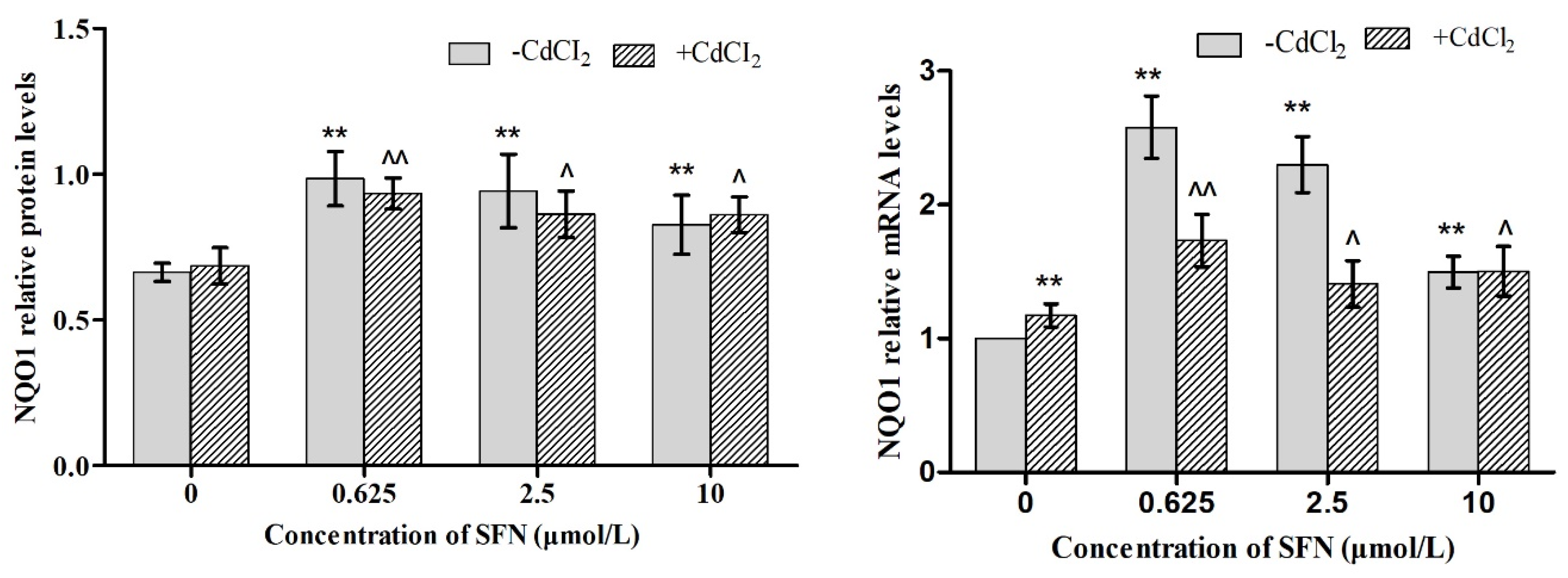
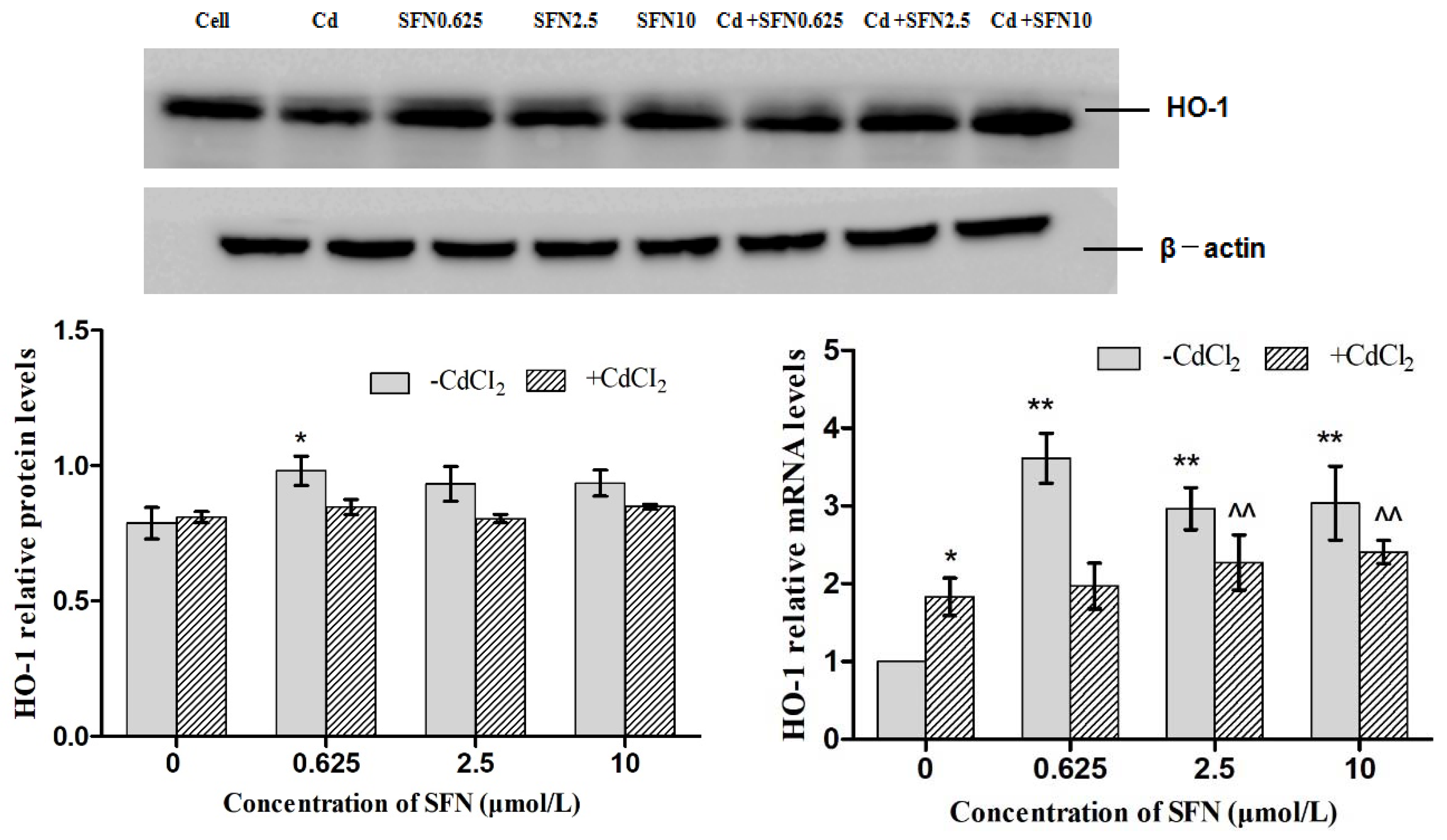
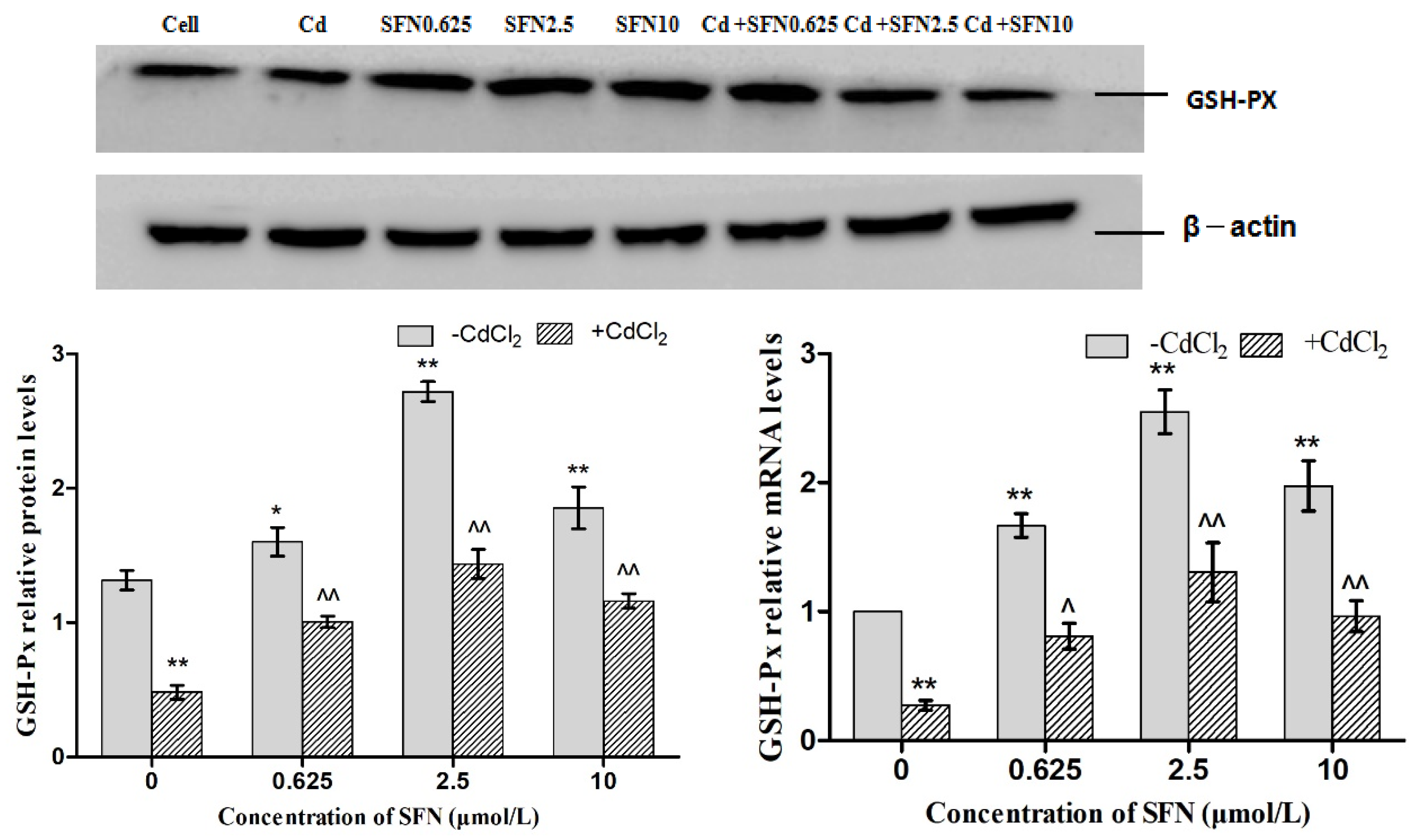
3.7. Effects of SFN and Cd on the mRNA and Relative Protein Expressions of Nrf2, HO-1, NQO1, GSH-Px, and γ-GCS in TM4 Cells
4. Discussion
Author Contributions
Funding
Conflicts of Interest
References
- E-Demerdash, F.M.; Yousef, M.I.; Kedwany, F.S.; Baghdadi, H.H. Cadmium-induced changes in lipid peroxidation, blood hematology, biochemical parameters and semen quality of male rats: Protective role of vitamin E and beta-carotene. Food Chem. Toxicol. 2004, 42, 1563–1571. [Google Scholar] [CrossRef] [PubMed]
- De Souza Predes, F.; Diamante, M.A.; Dolder, H. Testis response to low doses of cadmium in Wistar rats. Int. J. Exp. Pathol. 2010, 91, 125–131. [Google Scholar] [CrossRef] [PubMed]
- Thompson, J.; Bannigan, J. Cadmium: Toxic effects on the reproductive system and the embryo. Reprod. Toxicol. 2008, 25, 304–315. [Google Scholar] [CrossRef] [PubMed]
- Ognjanović, B.I.; Marković, S.D.; Ethordević, N.Z.; Trbojević, I.S.; Stajn, A.S.; Saicić, Z.S. Cadmium-induced lipid peroxidation and changes in antioxidant defense system in the rat testes: Protective role of coenzyme Q(10) and vitamin E. Reprod. Toxicol. 2010, 29, 191–197. [Google Scholar] [CrossRef] [PubMed]
- Ponnusamy, M.; Pari, L. Protective role of diallyl tetrasulfide on cadmium-induced testicular damage in adult rats: Abiochemical and histological study. Toxicol. Ind. Health 2011, 27, 407–416. [Google Scholar] [CrossRef] [PubMed]
- Kanter, M.; Aktoz, T.; Aktas, C.; Ozen, F.; Yarali, O.; Kanter, B. Role of quercetin in cadmium-induced oxidative stress, testicular damage, and apoptosis in rats. Anal. Quant. Cytopathol. Histpathol. 2016, 38, 45–51. [Google Scholar] [PubMed]
- Abarikwu, S.O.; Iserhienrhien, B.O.; Badejo, T.A. Rutin- and selenium-attenuated cadmium -induced testicular pathophysiology in rats. Hum. Exp. Toxicol. 2013, 32, 395–406. [Google Scholar] [CrossRef] [PubMed]
- Elbaghdady, H.; Alwaili, M.A.; El-Demerdash, R.S. Amelioration of cadmium-induced testes’ damage in rats by the bone marrow mesenchymalstem cells. Ecotoxicol. Environ. Saf. 2018, 148, 763–769. [Google Scholar] [CrossRef] [PubMed]
- Abdelrazek, H.M.; Helmy, S.A.; Elsayed, D.H.; Ebaid, H.M.; Mohamed, R.M. Ameliorating effects of green tea extract on cadmium induced reproductive injury in male Wistarrats with respect to androgen receptors and caspase-3. Reprod. Biol. 2016, 16, 300–308. [Google Scholar] [CrossRef] [PubMed]
- Jia, Y.; Lin, J.; Mi, Y.; Zhang, C. Quercetin attenuates cadmium-induced oxidative damage and apoptosis in granulosa cells from chicken ovarian follicles. Reprod. Toxicol. 2011, 31, 477–485. [Google Scholar] [CrossRef] [PubMed]
- Oguzturk, O.; Ciftci, M.; Aydin, N.; Imurkaan, A.; Beytur, F.; Yilmaz, H. Ameliorative effects of curcumin against acute cadmium toxicity on male reproductive system in rats. Andrologia 2012, 44, 243–249. [Google Scholar] [CrossRef]
- Bashir, N.; Manoharan, V.; Miltonprabu, S. Grape seed proanthocyanidins protects against cadmium induced oxidative pancreatitis in rats by attenuating oxidative stress, inflammation and apoptosis via Nrf-2/HO-1 signaling. J. Nutr. Biochem. 2016, 32, 128–141. [Google Scholar] [CrossRef] [PubMed]
- Yari, A.; Sarveazad, E.; Asadi, J.; Raouf Sarshoori, A.; Babahajian, N.; Amini, F.; Amidi, H.; Bahadoran, M.T.; Joghataei, M.H.; Asadi, A.; et al. Efficacy of Crocus sativus on reduction of cadmium-induced toxicity on spermatogenesis in adult rats. Andrologia 2016, 48, 1244–1252. [Google Scholar] [CrossRef]
- Lamas, C.; Cuquetto-Leite, L.; do Nascimento da Silva, E.; Thomazini, B.F.; Cordeiro, G.d.S.; Predes, F.d.S. Grape juice concentrate alleviates epididymis and sperm damage in cadmium-intoxicated rats. Int. J. Exp. Pathol. 2017, 98, 86–99. [Google Scholar] [CrossRef]
- Hongmei, W.; Ruili, Z.; Yangyang, S.; Tianqi, L.; Ming, G. Protective effect of ganoderma triterpenoids on cadmium-induced testicular toxicity in chickens. Biol. Trace Elem. Res. 2018, 3, 1–10. [Google Scholar]
- Jed, W.; Fahey, S.L.; Wehage, W.; David, H.; Thomas, W.K.; Patricia, A.E.; Theresa, A.S.; Paul, T. Protection of humans by plant glucosinolates: Efficiency of conversion of glucosinolates to isothiocyanates by the gastrointestinal microflora. Cancer Prev. Res. 2012, 5, 603–611. [Google Scholar]
- HayesJ, D.; Elleher, G.; Lestoni, M. The cancer actions of phytochemicals derived from chemopreventive glucosinolates. J. Eur. J. Nutr. 2008, 47, 73–88. [Google Scholar] [CrossRef] [PubMed]
- Angeloni, C.; Leoncini, E.; Malaguti, M. Modulation of phase II enzymes by sulforaphane: Implications for its cardioprotective potential. J. Agric. Food Chem. 2009, 57, 5615–5622. [Google Scholar] [CrossRef] [PubMed]
- Toyama, T.; Sumi, D.; Shinkai, Y. Cytoprotective role of Nrf2/Keap1 system in methylmercury toxicity. J. Biochem. Biophys. Res. Commun. 2007, 363, 645–650. [Google Scholar] [CrossRef]
- Li, J.-H.; Xv, Z.-F.; Li, S.-S.; Yang, T.-Y.; Liu, W.; Li, Y.-H.; Feng, S. Protective effect of sulforaphane on renal damage caused by cadmium in rats. Chin. J. Ind. Med. 2015, 28, 11–13. [Google Scholar]
- Shinkai, Y.; Sumi, D.; Fukami, I. Sulforaphane, an activator of Nrf2, suppresses cellular accumulation of arsenic and its cytotoxicity in primary mouse hepatocytes. J. FEBS Lett. 2006, 580, 1771–1774. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Alkharashi, N.A.O.; Periasamy, V.S.; Athinarayanan, J.; Alshatwi, A.A. Sulforaphane mitigatescadmium-induced toxicity pattern in human peripheral blood lymphocytes and monocytes. Environ. Toxicol. Pharmacol. 2017, 55, 223–239. [Google Scholar] [CrossRef]
- Jahan, S.; Ahmed, S. Comparative analysis of antioxidants against cadmium induced reproductive toxicity in adult male rats. J. Syst. Biol. Reprod. Med. 2014, 60, 28–34. [Google Scholar] [CrossRef] [PubMed]
- Yang, S.-H.; Miao, L.; Yu, L.-H.; Li, L.; Li, P.; Zhang, Y.; Guo, Y.; Gao, G.; Liu, M.-D.; He, J.-B. Sulforaphane prevents testicular damage in kunming mice exposed to cadmium via activation of Nrf2/ARE signaling pathways. Int. J. Mol. Sci. 2016, 17. [Google Scholar] [CrossRef] [PubMed]
- Zhai, Q.; Narbad, A.; Chen, W. Dietary strategies for the treatment of cadmium and lead toxicity. Nutrients 2015, 7, 552–571. [Google Scholar] [CrossRef]
- Porru, S.; Alessio, L. The use of chelating agents in occupational lead poisoning. Occup. Med. 1996, 46, 41–48. [Google Scholar] [CrossRef]
- Xue, M.; Qian, Q.; Adaikalakoteswari, A.; Rabbani, N.; Babaei-Jadidi, R.; Thornalley, P.J. Activation of NF-E2-related actor-2 reverses biochemical dysfunction of endothelial cellsinduced by hyperglycemia linked to vascular disease. Diabetes 2008, 57, 2809–2817. [Google Scholar] [CrossRef]
- Dinkova-Kostova, A.T.; Kostov, R.V. Glucosinolates and isothiocyanates in health and disease. Trends Mol. Med. 2012, 18, 337–347. [Google Scholar] [CrossRef]
- Lee, J.S.; Surh, Y.J. Nrf2 as a novel molecular target for chemoprevention. J. Cancer Lett. 2005, 224, 171–184. [Google Scholar] [CrossRef] [PubMed]
- Guillouzo, A.; Uguen-Guillouzo, C. In Vitro Methods in Toxicology; Academic Press: New York, NY, USA, 1992. [Google Scholar]
- Wang, L.; Cao, J.; Chen, D.; Liu, X.; Lu, H.; Liu, Z. Role of oxidative stress, apoptosis, and intracellular homeostasis in primary cultures of ratproximal tubular cells exposed to cadmium. Biol. Trace Elem. Res. 2009, 127, 53–68. [Google Scholar] [CrossRef]
- Angenard, G.; Muczynski, V.; Coffigny, H.; Pairault, C.; Duquenne, C.; Frydman, R.; Habert, R.; Rouiller-Fabre, V.; Livera, G. Cadmium increases human fetal germ cell apoptosis. Environ. Health Perspect. 2010, 118, 331–337. [Google Scholar] [CrossRef] [PubMed]
- Liu, W.; Zhao, H.Y.; Wang, Y.; Jiang, C.Y.; Xia, P.P.; Gu, J.H. Calcium-calmodulin signaling elicits mitochondrial dysfunction and the release of cytochrome c during cadmium-induced apoptosis in primary osteoblasts. Toxicol. Lett. 2014, 224, 1–6. [Google Scholar] [CrossRef] [PubMed]
- Alkharashi, N.A.O.; Periasamy, V.S.; Athinarayanan, J.; Alshatwi, A.A. Cadmium triggers mitochondrial oxidative stress in human peripheral blood lymphocytes and monocytes: Analysis using in vitro and system toxicology approaches. J. Trace Elem. Med. Biol. 2017, 42, 117–128. [Google Scholar] [CrossRef] [PubMed]
- Abdulkareem, N.; Alkharashi, O.; Periasamy, V.S.; Athinarayanan, J.; Shatwi, A.A.A. Assessment of sulforaphane-induced protective mechanisms against cadmium toxicity in human mesenchymal stem cells. Environ. Sci. Pollut. Res. 2018, 25, 10080–10089. [Google Scholar]
- Tao, S.; Justiniano, R.; Zhang, D.D.; Wondrak, G.T. The Nrf2-inducers tanshinone I and dihydrotanshinone protect human skin cells and reconstructed human skin against solar simulated UV. Redox Biol. 2013, 29, 532–541. [Google Scholar] [CrossRef] [PubMed]
- Jung, K.A.; Kwak, M.K. The Nrf2 system as a potential target for the development of indirect antioxidants. J. Mol. 2010, 15, 7266–7291. [Google Scholar] [CrossRef] [PubMed]
- Oh, C.J.; Kim, J.Y.; Min, A.K. Sulforaphane attenuates hepatic fibrosis via NF-E2-related factor 2-mediated inhibition of transforming growth factor-β/smad signaling. J. Free Radic. Biol. Med. 2012, 52, 671–682. [Google Scholar] [CrossRef] [PubMed]
- Lu, M.C.; Ji, J.A.; Jiang, Z.Y.; You, Q.D. The Keap1-Nrf2-ARE pathway as a potential preventive and therapeutic target: An update. Med. Res. Rev. 2016, 36, 924–963. [Google Scholar] [CrossRef] [PubMed]
- Furia, E.; Aiello, D.; di Donna, L.; Mazzotti, F.; Tagarelli, A.; Thangavel, H.; Napoli, A.; Sindona, G. Mass spectrometry and potentiometry studies of Pb(II)–, Cd(II)– and Zn(II)–cystine complexes. Dalton Trans. 2013, 43, 1055–1062. [Google Scholar] [CrossRef] [PubMed]
Sample Availability: Not Available |















{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Gene | Accession No. | Primer Sequence (5′–3′) | Product Length |
|---|---|---|---|
| Nrf2 | NM_010902.3 | Forward: TCCTATGCGTGAATCCCAAT | 103 bp |
| Reverse: GCGGCTTGAATGTTTGTCTT | |||
| GSH-Px | X03920.1 | Forward: GAAGTGCGAAGTGAATGG | 224 bp |
| Reverse: TGTCGATGGTACGAAAGC | |||
| HO-1 | NM_010442.2 | Forward: GGGCTGTGAACTCTGTCCAAT | 162 bp |
| Reverse: GGTGAGGGAACTGTGTCAGG | |||
| γ-GCS | U85414.1 | Forward: TGGATGATGCCAACGAGTC | 185 bp |
| Reverse: CCTAGTGAGCAGTACCACGAATA | |||
| NQO1 | NM_008706.5 | Forward: TTCTGTGGCTTCCAGGTCTT | 104 bp |
| Reverse: TCCAGACGTTTCTTCCATCC | |||
| β-actin | BC138614.1 | Forward: CTGTCCCTGTATGCCTCTG | 221 bp |
| Reverse: TTGATGTCACGCACGATT |
| Group | Apoptosis Rate (%) |
|---|---|
| Control (0 µmo/L) | 4.3 ± 0.37 |
| CdCl2 (12.5 µmo/L) | 6.17 ± 0.82 * |
| SFN (0.625 µmo/L) | 2.67 ± 0.62 ** |
| SFN (2.5 µmo/L) | 1.95 ± 0.14 ** |
| SFN (10 µmo/L) | 2.37 ± 0.08 ** |
| CdCl2 (12.5 µmo/L) + SFN0.625 µmo/L | 2.92 ± 0.18 ## |
| CdCl2 (12.5 µmo/L) + SFN2.5 µmo/L | 2.52 ± 0.25 ## |
| CdCl2 (12.5 µmo/L) + SFN10 µmo/L | 2.70 ± 0.35 ## |
© 2018 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Yang, S.-H.; Yu, L.-H.; Li, L.; Guo, Y.; Zhang, Y.; Long, M.; Li, P.; He, J.-B. Protective Mechanism of Sulforaphane on Cadmium-Induced Sertoli Cell Injury in Mice Testis via Nrf2/ARE Signaling Pathway. Molecules 2018, 23, 1774. https://doi.org/10.3390/molecules23071774
Yang S-H, Yu L-H, Li L, Guo Y, Zhang Y, Long M, Li P, He J-B. Protective Mechanism of Sulforaphane on Cadmium-Induced Sertoli Cell Injury in Mice Testis via Nrf2/ARE Signaling Pathway. Molecules. 2018; 23(7):1774. https://doi.org/10.3390/molecules23071774
Chicago/Turabian StyleYang, Shu-Hua, Li-Hui Yu, Lin Li, Yang Guo, Yi Zhang, Miao Long, Peng Li, and Jian-Bin He. 2018. "Protective Mechanism of Sulforaphane on Cadmium-Induced Sertoli Cell Injury in Mice Testis via Nrf2/ARE Signaling Pathway" Molecules 23, no. 7: 1774. https://doi.org/10.3390/molecules23071774