The Possible Reduction Mechanism of Volatile Sulfur Compounds during Durian Wine Fermentation Verified in Modified Buffers


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Results and Discussion
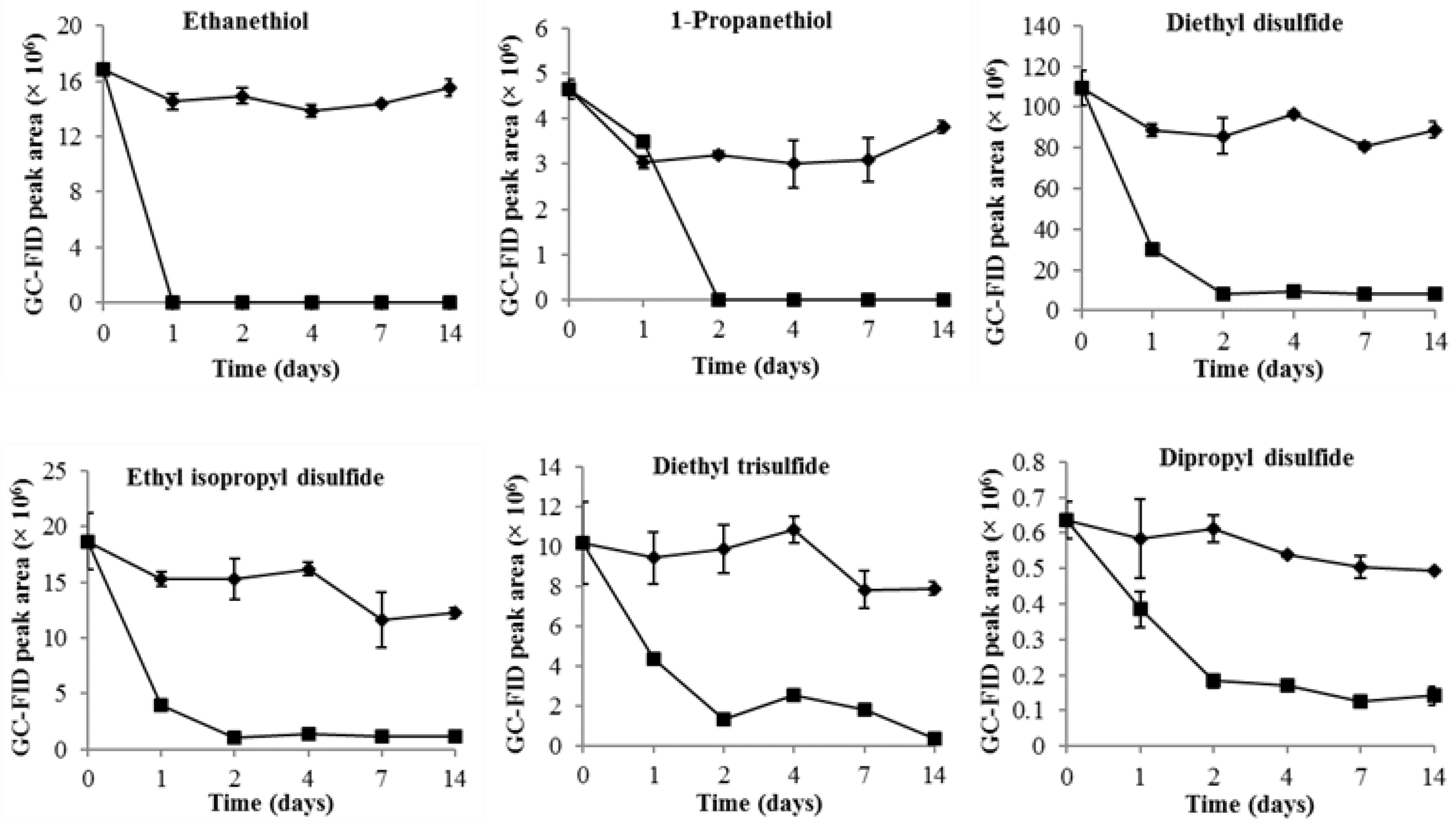
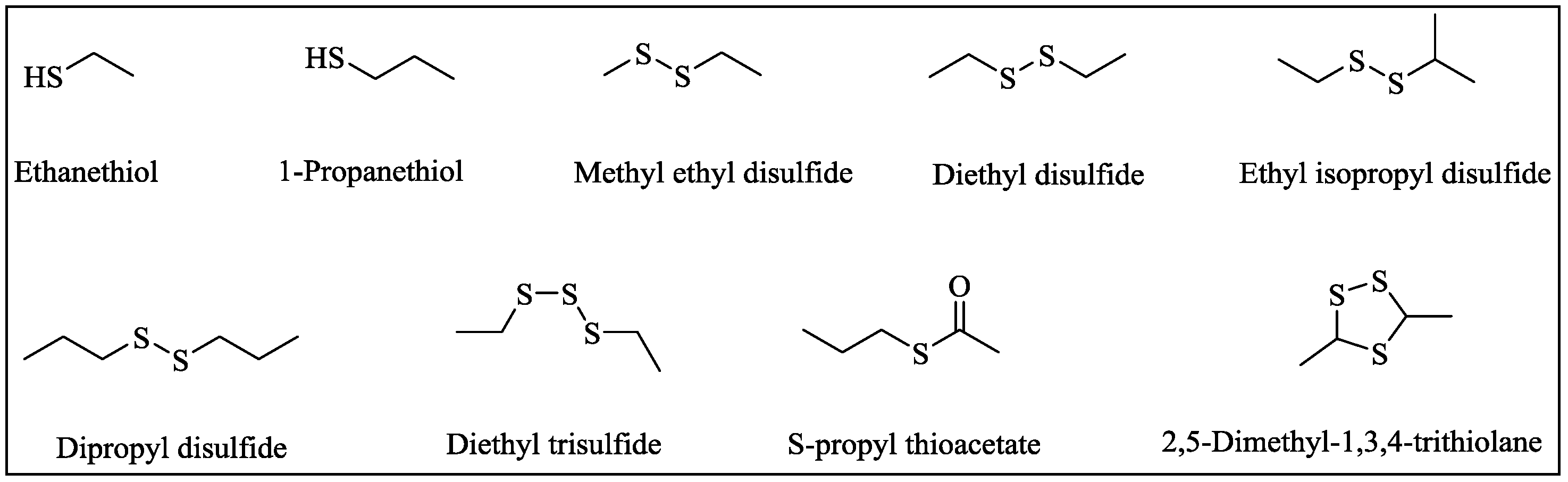
2.1. Changes of VSCs during Durian Wine Fermentation
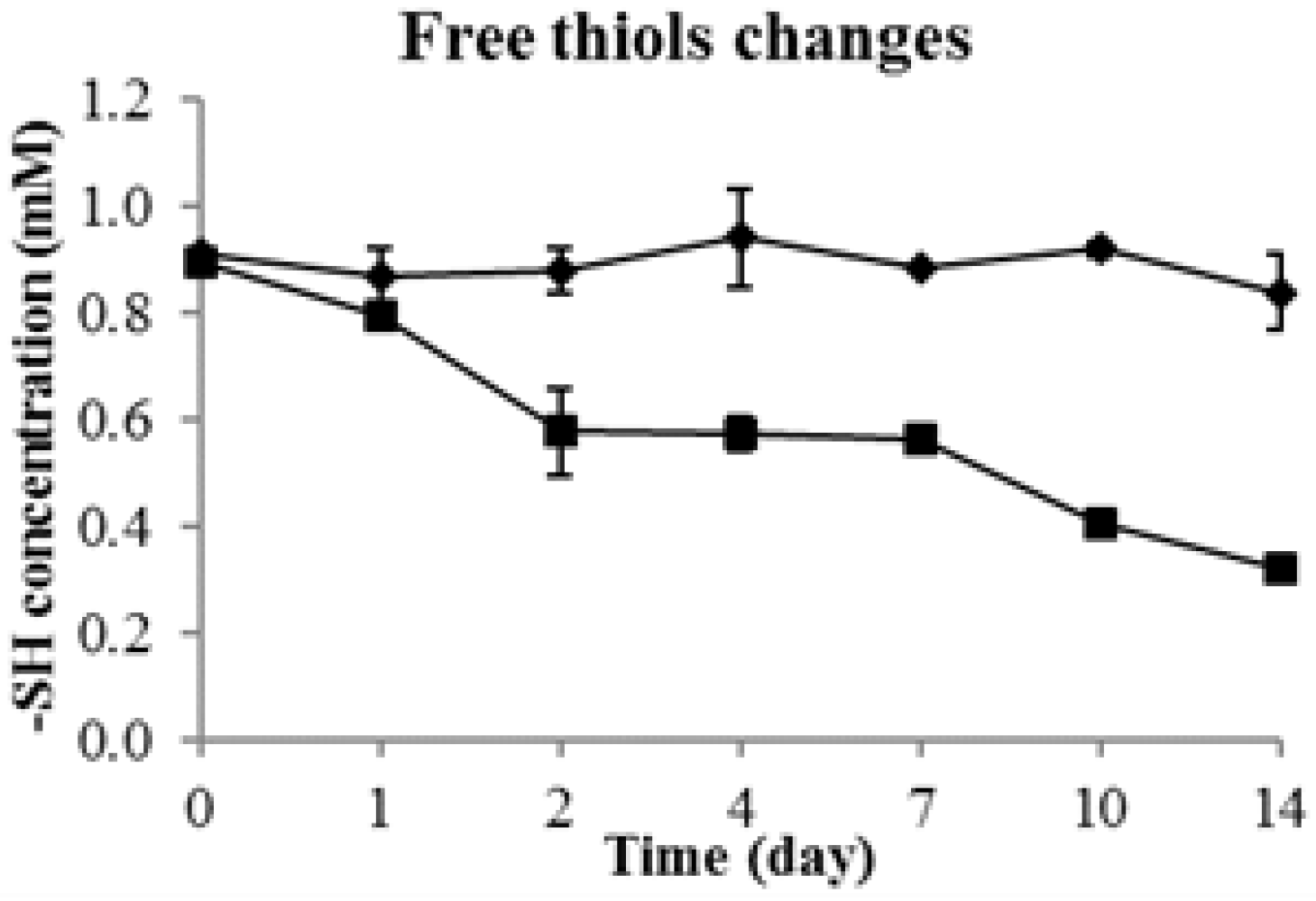
2.2. Changes of Total Thiols during Durian Wine Fermentation
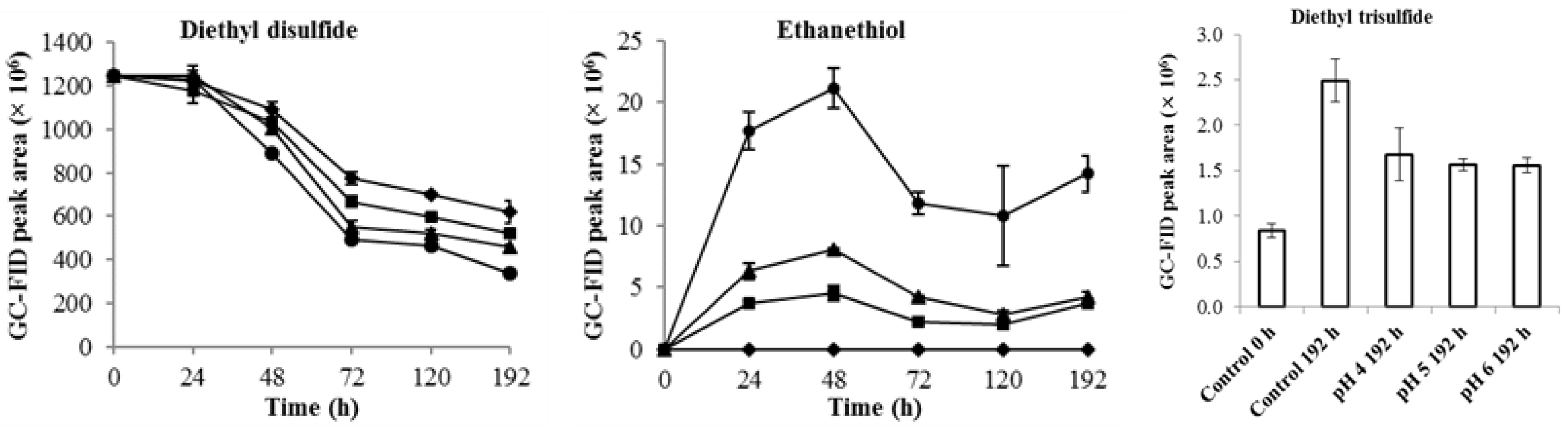
2.3. Interconversion of Diethyl Disulfide and Ethanethiol
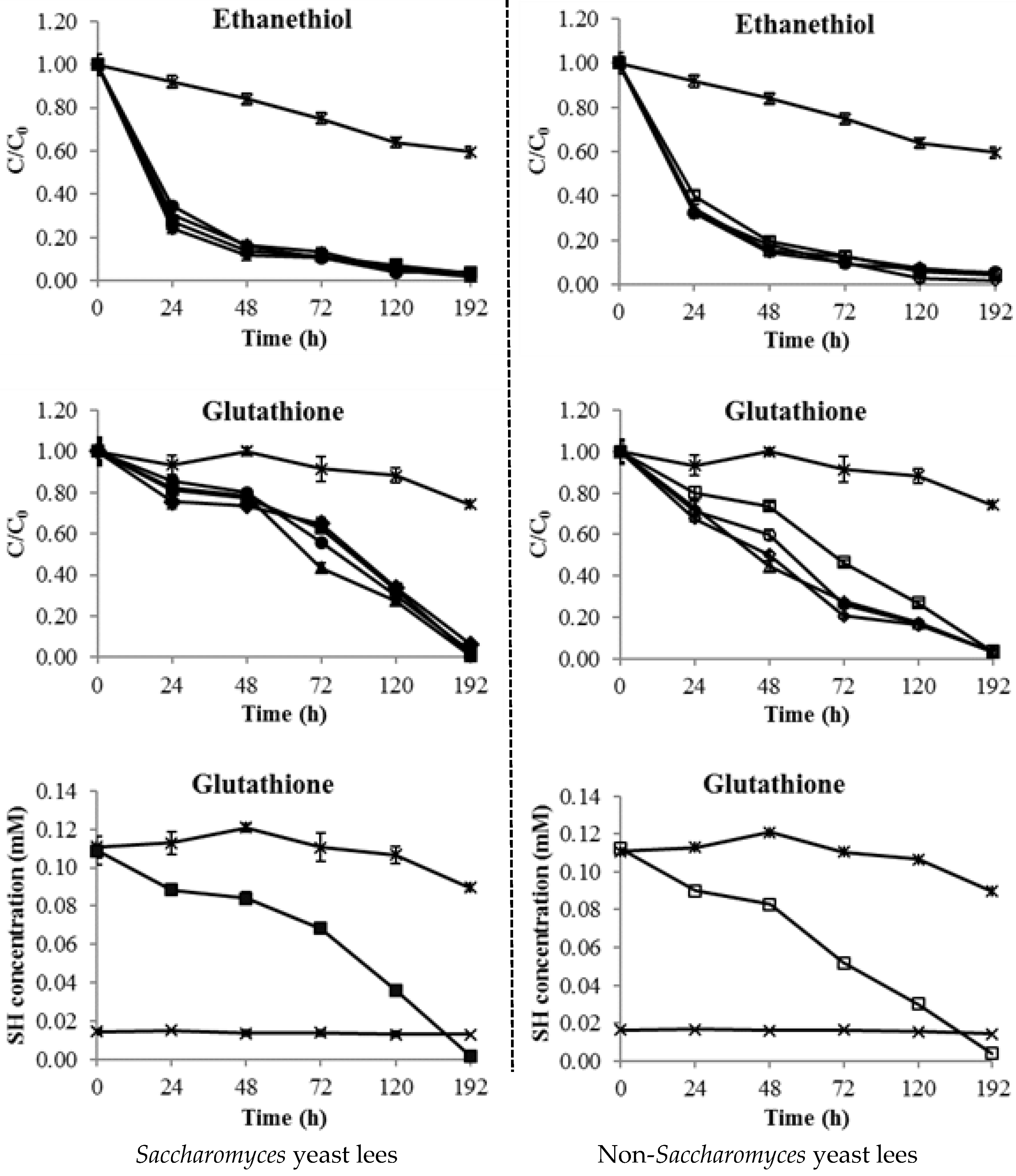
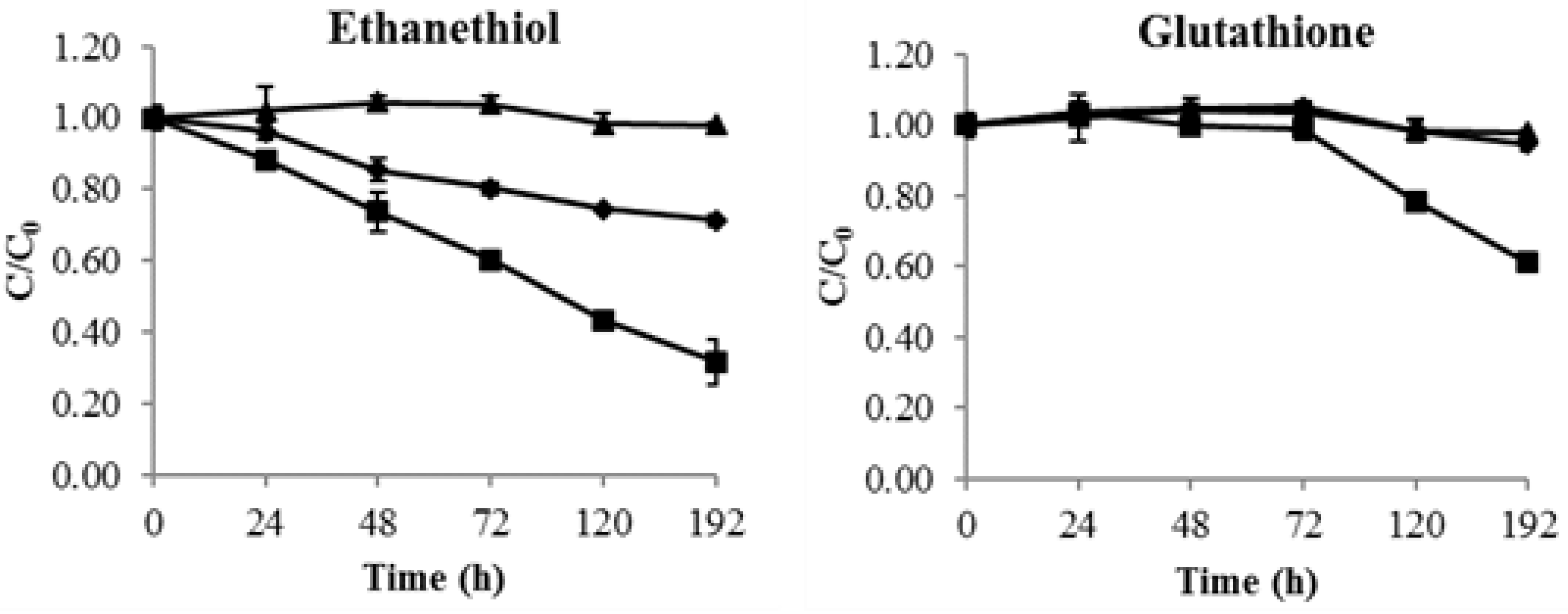
2.4. Consumption of Thiols by Yeast Lees
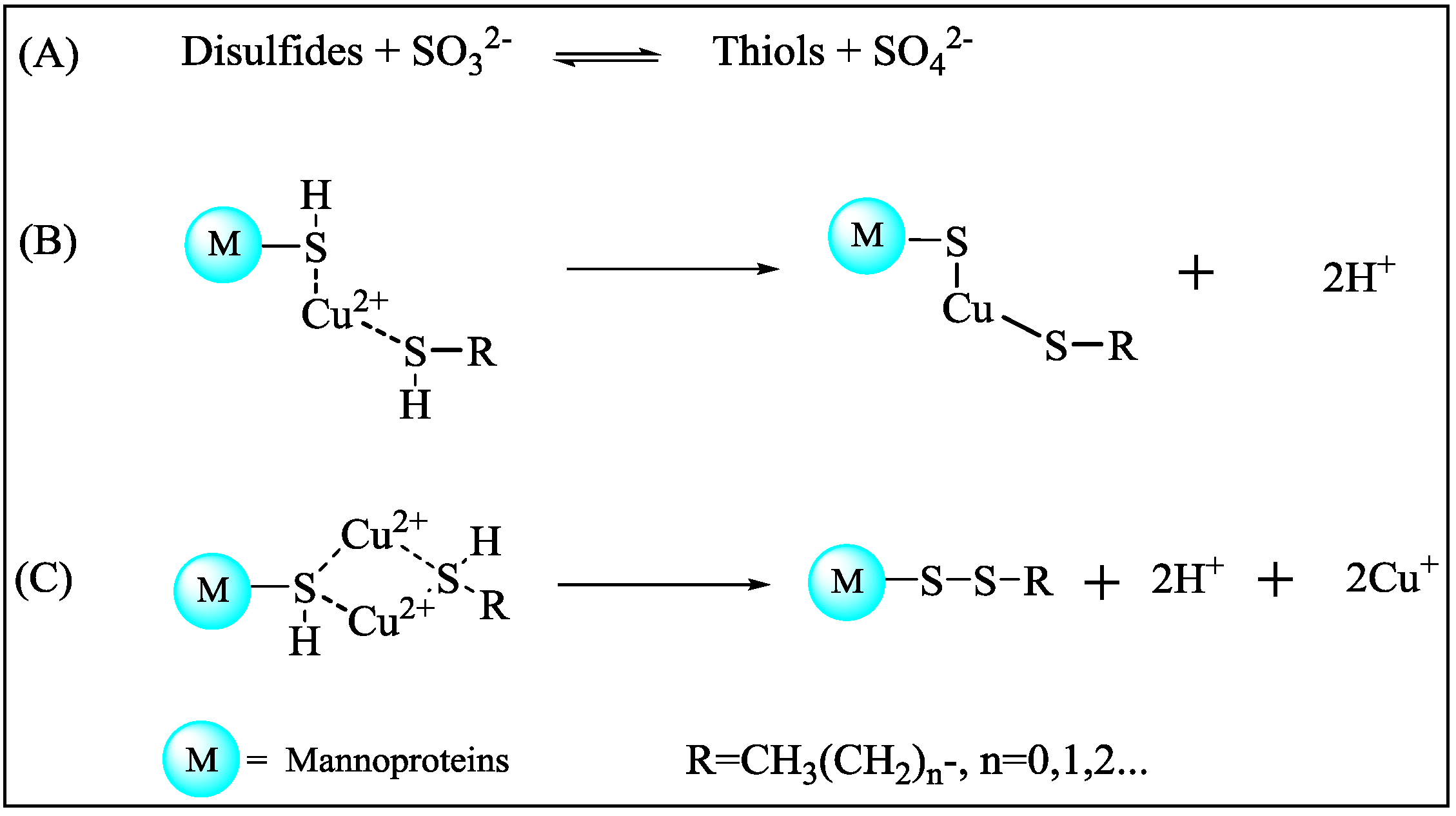
2.5. Consumption of Thiols by Mannoprotein
3. Materials and Methods
3.1. Durian Wine Fermentation with Saccharomyces Cerevisiae EC-1118
3.2. Interconversion of Diethyl Disulfide and Ethanethiol in Modified Buffer
3.3. Detection of Volatile Sulfur Compounds
3.4. Consumption of Thiols by Yeast Lees
3.5. Consumption of Thiols by Mannoproteins
3.6. Quantification of Free Thiol
3.7. Statistical Analysis
4. Conclusions
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Haruenkit, R.; Poovarodom, S.; Leontowicz, H.; Leontowicz, M.; Sajewicz, M.; Kowalska, T.; Delgado-Licon, E.; Rocha-Guzmán, N.E.; Gallegos-Infante, J.A.; Trakhtenberg, S. Comparative study of health properties and nutritional value of durian, mangosteen, and snake fruit: Experiments in vitro and in vivo. J. Agric. Food Chem. 2007, 55, 5842–5849. [Google Scholar] [CrossRef] [PubMed]
- Ho, L.H.; Bhat, R. Exploring the potential nutraceutical values of durian (Durio zibethinus L.)—An exotic tropical fruit. Food Chem. 2015, 168, 80–89. [Google Scholar] [CrossRef] [PubMed]
- Liang, D.; Wang, C.; Tocmo, R.; Wu, H.; Deng, L.W.; Huang, D. Hydrogen sulphide (H2S) releasing capacity of essential oils isolated from organosulphur rich fruits and vegetables. J. Funct. Foods 2015, 14, 634–640. [Google Scholar] [CrossRef]
- Arancibia-Avila, P.; Toledo, F.; Park, Y.S.; Jung, S.T.; Kang, S.G.; Heo, B.G.; Lee, S.H.; Sajewicz, M.; Kowalska, T.; Gorinstein, S. Antioxidant properties of durian fruit as influenced by ripening. LWT-Food Sci. Technol. 2008, 41, 2118–2125. [Google Scholar] [CrossRef]
- Leontowicz, M.; Leontowicz, H.; Jastrzebski, Z.; Jesion, I.; Haruenkit, R.; Poovarodom, S.; Katrich, E.; Tashma, Z.; Drzewiecki, J.; Trakhtenberg, S. The nutritional and metabolic indices in rats fed cholesterol-containing diets supplemented with durian at different stages of ripening. Biofactors 2007, 29, 123–136. [Google Scholar] [CrossRef] [PubMed]
- Berry, S.K. Fatty acid composition and organoleptic quality of four clones of durian (Durio zibethinus, Murr.). J. Am. Oil Chem. Soc. 1981, 58, 716–717. [Google Scholar] [CrossRef]
- Maninang, J.S.; Lizada, M.C.C.; Gemma, H. Inhibition of aldehyde dehydrogenase enzyme by Durian (Durio zibethinus Murray) fruit extract. Food Chem. 2009, 117, 352–355. [Google Scholar] [CrossRef] [Green Version]
- Mutschler, J.; Grosshans, M.; Soyka, M.; Rösner, S. Current findings and mechanisms of action of disulfiram in the treatment of alcohol dependence. Pharmacopsychiatry 2016, 49, 137–141. [Google Scholar] [CrossRef] [PubMed]
- Seitz, H.K.; Becker, P. Alcohol metabolism and cancer risk. Alcohol Res. Health 2007, 30, 38–47. [Google Scholar] [PubMed]
- Lee, P.R.; Saputra, A.; Yu, B.; Curran, P.; Liu, S.Q. Biotransformation of durian pulp by mono-and mixed-cultures of Saccharomyces cerevisiae and Williopsis saturnus. LWT-Food Sci. Technol. 2012, 46, 84–90. [Google Scholar] [CrossRef]
- Swiegers, J.; Pretorius, I. Modulation of volatile sulfur compounds by wine yeast. Appl. Microbiol. Biotechnol. 2007, 74, 954–960. [Google Scholar] [CrossRef] [PubMed]
- Suárez-Lepe, J.A.; Morata, A. New trends in yeast selection for winemaking. Trends Food Sci. Technol. 2012, 23, 39–50. [Google Scholar] [CrossRef]
- Moreira, N.; De Pinho, P.G.; Santos, C.; Vasconcelos, I. Relationship between nitrogen content in grapes and volatiles, namely heavy sulphur compounds, in wines. Food Chem. 2011, 126, 1599–1607. [Google Scholar] [CrossRef] [PubMed]
- Kinzurik, M.I.; Herbst-Johnstone, M.; Gardner, R.C.; Fedrizzi, B. Hydrogen sulfide production during yeast fermentation causes the accumulation of ethanethiol, S-ethyl thioacetate and diethyl disulfide. Food Chem. 2016, 209, 341–347. [Google Scholar] [CrossRef] [PubMed]
- Sedighi, M.; Vahabzadeh, F.; Zamir, S.M.; Naderifar, A. Ethanethiol degradation by Ralstonia eutropha. Biotechnol. Bioprocess E 2013, 18, 827–833. [Google Scholar] [CrossRef]
- Kreitman, G.Y.; Danilewicz, J.C.; Jeffery, D.W.; Elias, R.J. Reaction mechanisms of metals with hydrogen sulfide and thiols in model wine. Part 1: Copper-catalyzed oxidation. J. Agric. Food Chem. 2016, 64, 4095–4104. [Google Scholar] [CrossRef] [PubMed]
- Kreitman, G.Y.; Danilewicz, J.C.; Jeffery, D.W.; Elias, R.J. Reaction mechanisms of metals with hydrogen sulfide and thiols in model wine. Part 2: Iron-and copper-catalyzed oxidation. J. Agric. Food Chem. 2016, 64, 4105–4113. [Google Scholar] [CrossRef] [PubMed]
- Nikolantonaki, M.; Chichuc, I.; Teissedre, P.L.; Darriet, P. Reactivity of volatile thiols with polyphenols in a wine-model medium: Impact of oxygen, iron, and sulfur dioxide. Anal. Chim. Acta 2010, 660, 102–109. [Google Scholar] [CrossRef] [PubMed]
- Bassil, D.; Makris, D.P.; Kefalas, P. Oxidation of caffeic acid in the presence of l-cysteine: Isolation of 2-S-cysteinylcaffeic acid and evaluation of its antioxidant properties. Food Res. Int. 2005, 38, 395–402. [Google Scholar] [CrossRef]
- Lavigne, V.; Dubourdieu, D. Demonstration and interpretation of the yeast lees ability to absorb certain volatile thiols contained in wine. J. Int. Sci. Vigne Vin. 1996, 30, 201–206. [Google Scholar]
- Pérez-Serradilla, J.; De Castro, M.L. Role of lees in wine production: A review. Food Chem. 2008, 111, 447–456. [Google Scholar] [CrossRef] [PubMed]
- Tirelli, A.; Fracassetti, D.; De Noni, I. Determination of reduced cysteine in oenological cell wall fractions of Saccharomyces cerevisiae. J. Agric. Food Chem. 2010, 58, 4565–4570. [Google Scholar] [CrossRef] [PubMed]
- Vasserot, Y.; Steinmetz, V.; Jeandet, P. Study of thiol consumption by yeast lees. Anton Leeuw. 2003, 83, 201–207. [Google Scholar] [CrossRef]
- Palacios, S.; Vasserot, Y.; Maujean, A. Evidence for sulfur volatile products adsorption by yeast lees. Am. J. Enol. Vitic. 1997, 48, 525–526. [Google Scholar]
- Gómez-Plaza, E.; Cano-López, M. A review on micro-oxygenation of red wines: Claims, benefits and the underlying chemistry. Food Chem. 2011, 125, 1131–1140. [Google Scholar] [CrossRef]
- Bobet, R.A.; Noble, A.C.; Boulton, R.B. Kinetics of the ethanethiol and diethyl disulfide interconversion in wine-like solutions. J. Agric. Food Chem. 1990, 38, 449–452. [Google Scholar] [CrossRef]
- Lu, Y.; Voon, M.K.W.; Huang, D.; Lee, P.R.; Liu, S.Q. Combined effects of fermentation temperature and pH on kinetic changes of chemical constituents of durian wine fermented with Saccharomyces cerevisiae. Appl. Microbiol. Biotechnol. 2017, 101, 3005–3014. [Google Scholar] [CrossRef] [PubMed]
- Lu, Y.; Huang, D.; Lee, P.R.; Liu, S.Q. Effects of cofermentation and sequential inoculation of Saccharomyces bayanus and Torulaspora delbruckii on durian wine composition. Int. J. Food Sci. Technol. 2015, 50, 2653–2663. [Google Scholar] [CrossRef]
- Lu, Y.; Huang, D.; Lee, P.R.; Liu, S.Q. Assessment of volatile and non-volatile compounds in durian wines fermented with four commercial non-Saccharomyces yeasts. J. Sci. Food Agric. 2016, 96, 1511–1521. [Google Scholar] [CrossRef] [PubMed]
- Yuliana, N.; Dizon, E.I. Phenotypic identification of lactic acid bacteria isolated from Tempoyak (fermented durian) made in the Philippines. Int. J. Biol. 2011, 3, 145. [Google Scholar] [CrossRef]
- Devalaraja, S.; Jain, S.; Yadav, H. Exotic fruits as therapeutic complements for diabetes, obesity and metabolic syndrome. Food Res. Int. 2011, 44, 1856–1865. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Posada, J.O.G.; Hall, P.J. Towards the development of safe and commercially viable nickel-iron batteries: Improvements to Coulombic efficiency at high iron sulphide electrode formulations. J. Appl. Electrochem. 2016, 46, 451–458. [Google Scholar] [CrossRef]
- Asghar, H.; Hussain, S.; Sattar, H.; Brown, N.; Roberts, E. Mercaptan’s removal from aqueous solution using modified graphite-based adsorbent through batch-wise adsorption-regeneration. Chem. Eng. Commun. 2015, 202, 1155–1160. [Google Scholar] [CrossRef]
- Chithrani, B.D.; Ghazani, A.A.; Chan, W.C. Determining the size and shape dependence of gold nanoparticle uptake into mammalian cells. Nano Lett. 2006, 6, 662–668. [Google Scholar] [CrossRef] [PubMed]
- Jaehrig, S.C.; Rohn, S.; Kroh, L.W.; Fleischer, L.G.; Kurz, T. In vitro potential antioxidant activity of (1→3), (1→6)-β-d-glucan and protein fractions from Saccharomyces cerevisiae cell walls. J. Agric. Food Chem. 2007, 55, 4710–4716. [Google Scholar] [CrossRef] [PubMed]
Sample Availability: Samples of the compounds are not available from the authors. |







© 2018 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Lu, Y.; Fong, A.S.Y.L.; Chua, J.-Y.; Huang, D.; Lee, P.-R.; Liu, S.-Q. The Possible Reduction Mechanism of Volatile Sulfur Compounds during Durian Wine Fermentation Verified in Modified Buffers. Molecules 2018, 23, 1456. https://doi.org/10.3390/molecules23061456
Lu Y, Fong ASYL, Chua J-Y, Huang D, Lee P-R, Liu S-Q. The Possible Reduction Mechanism of Volatile Sulfur Compounds during Durian Wine Fermentation Verified in Modified Buffers. Molecules. 2018; 23(6):1456. https://doi.org/10.3390/molecules23061456
Chicago/Turabian StyleLu, Yuyun, Alicia Sarah Yoke Ling Fong, Jian-Yong Chua, Dejian Huang, Pin-Rou Lee, and Shao-Quan Liu. 2018. "The Possible Reduction Mechanism of Volatile Sulfur Compounds during Durian Wine Fermentation Verified in Modified Buffers" Molecules 23, no. 6: 1456. https://doi.org/10.3390/molecules23061456