Inhibitory Activity of Allergic Contact Dermatitis and Atopic Dermatitis-Like Skin in BALB/c Mouse through Oral Administration of Fermented Barks of Alnus sibirica

Abstract
:1. Introduction
2. Results and Discussions
2.1. Anti-Oxidative Activity
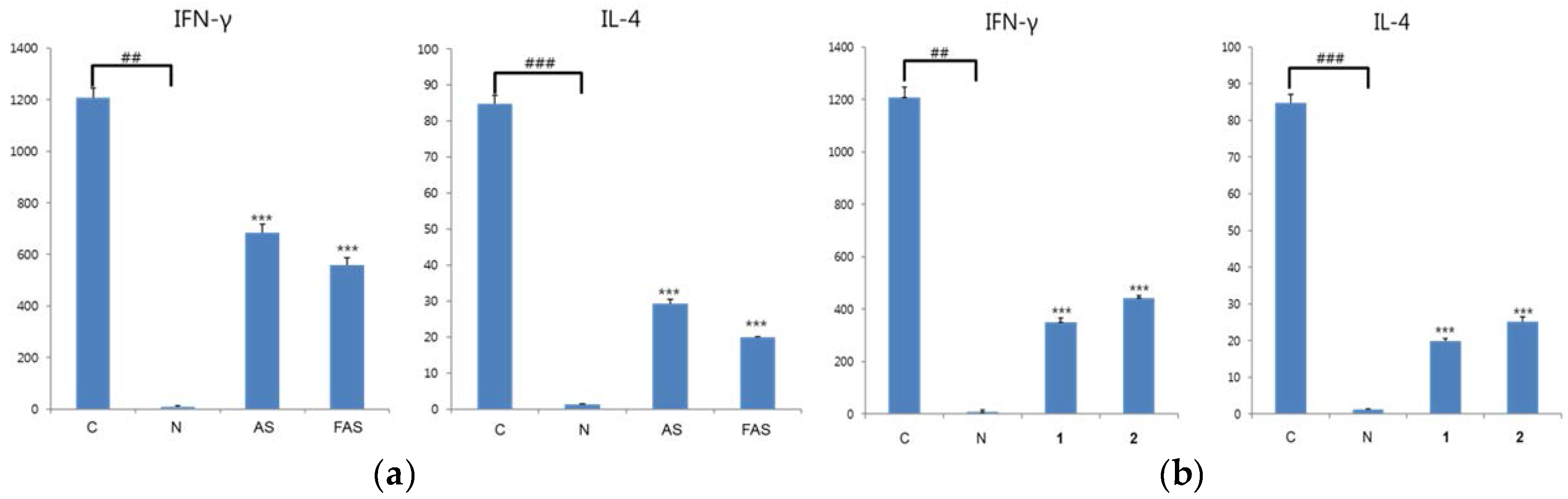
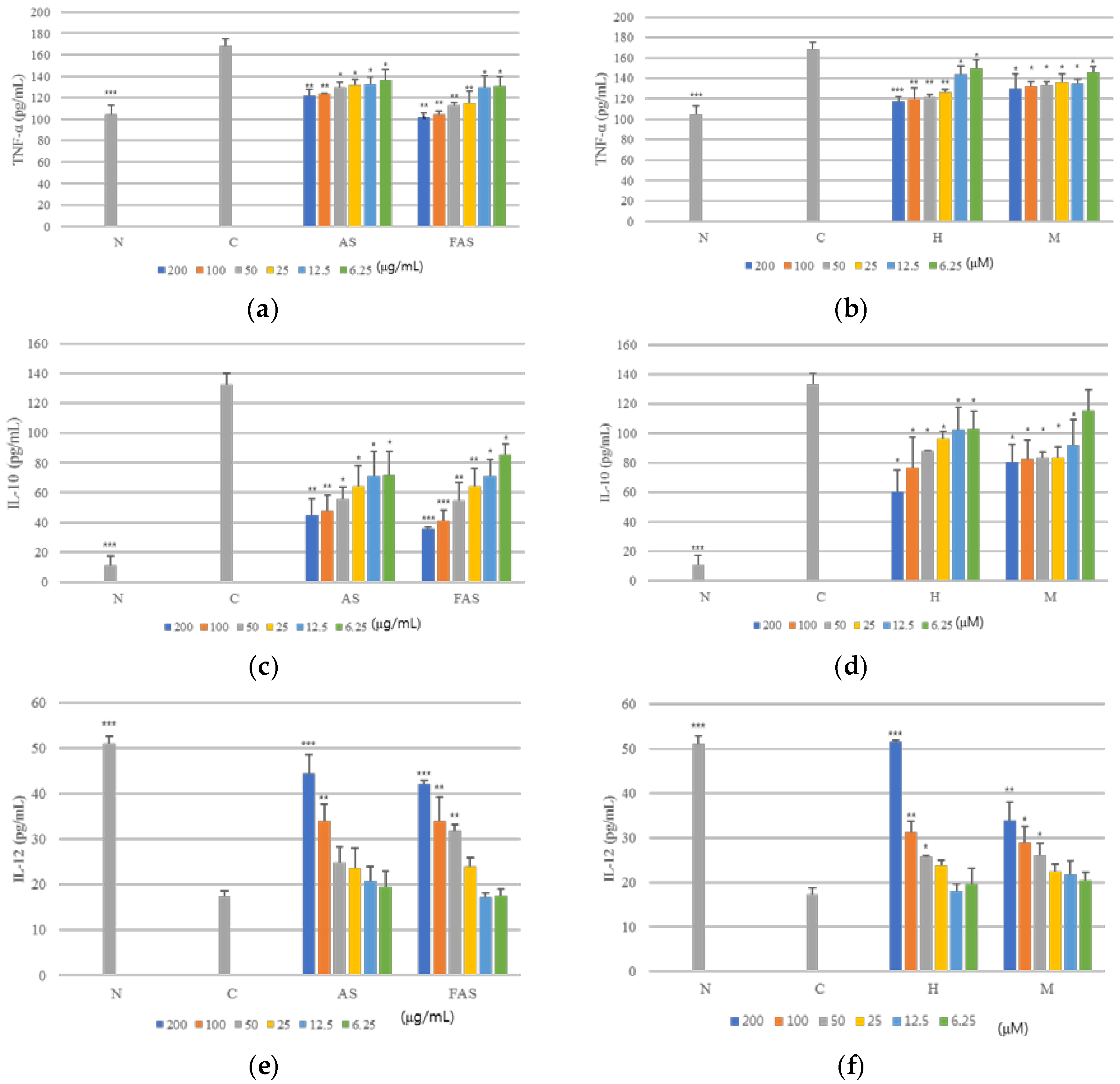
2.2. Anti-Inflammatory Activity
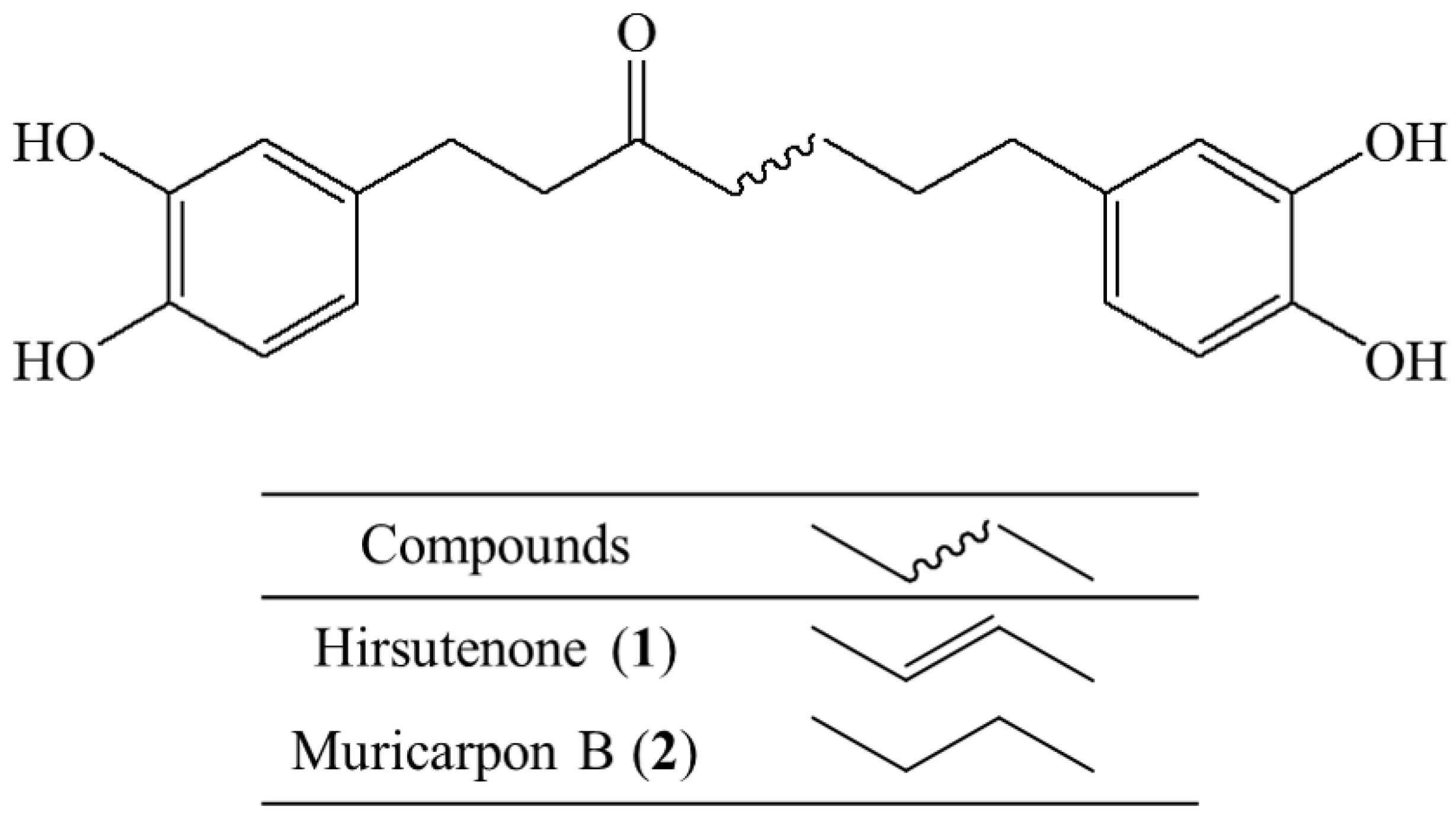
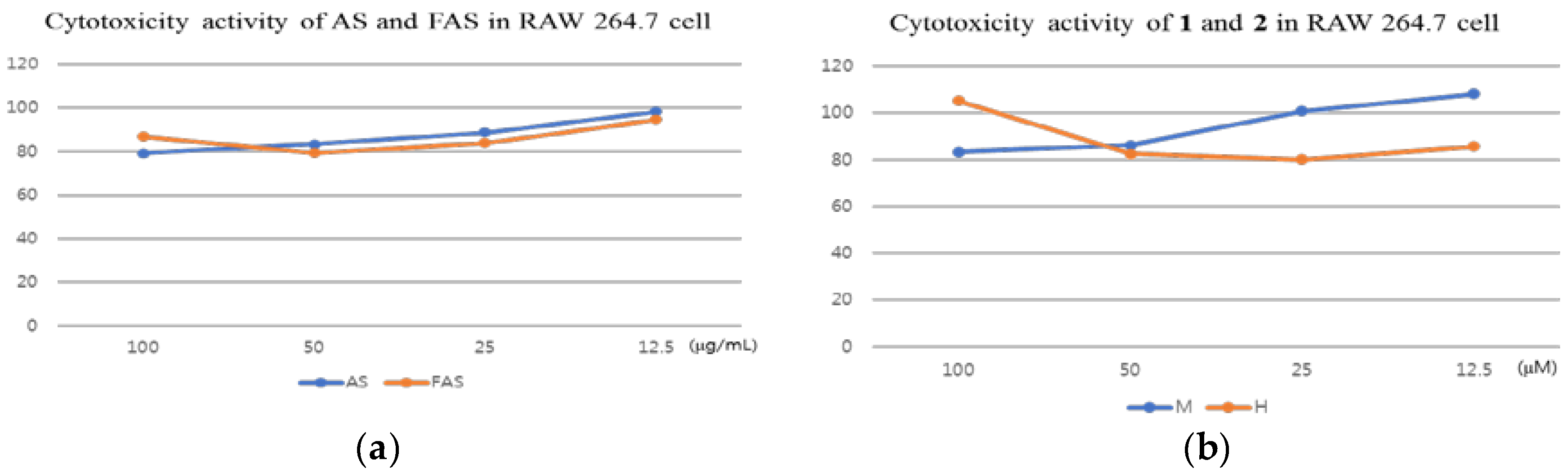
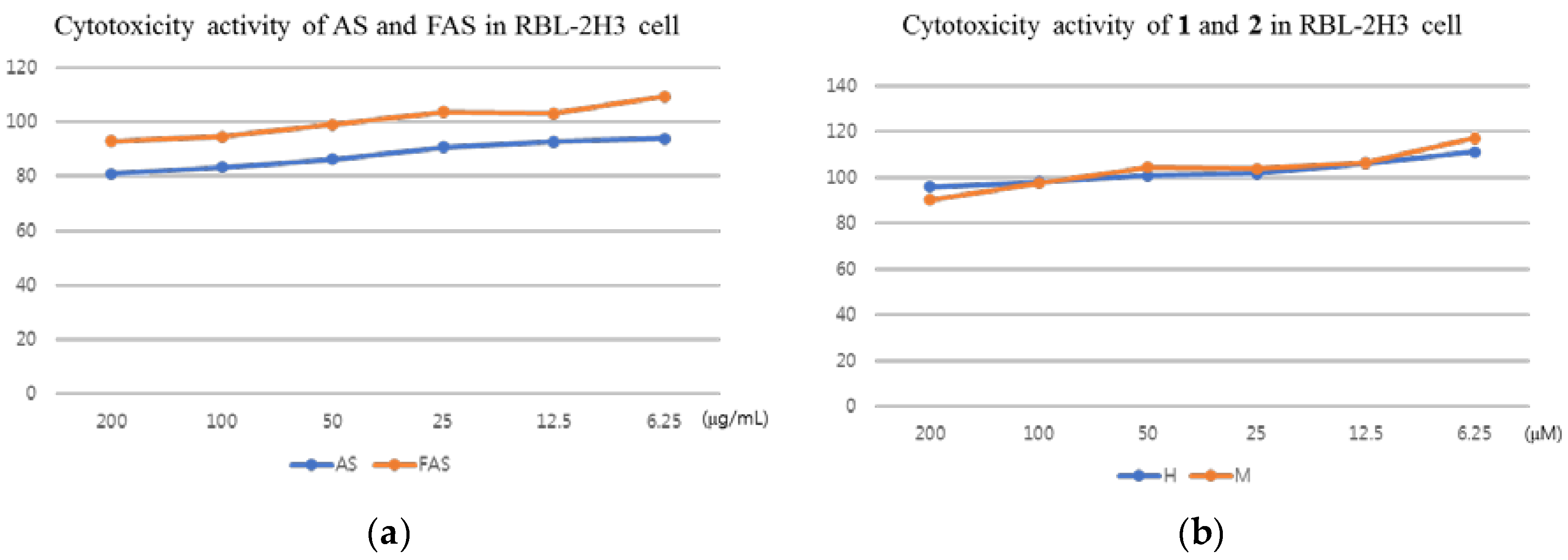
2.3. Anti-ACD and Anti-AD Activity in Vitro
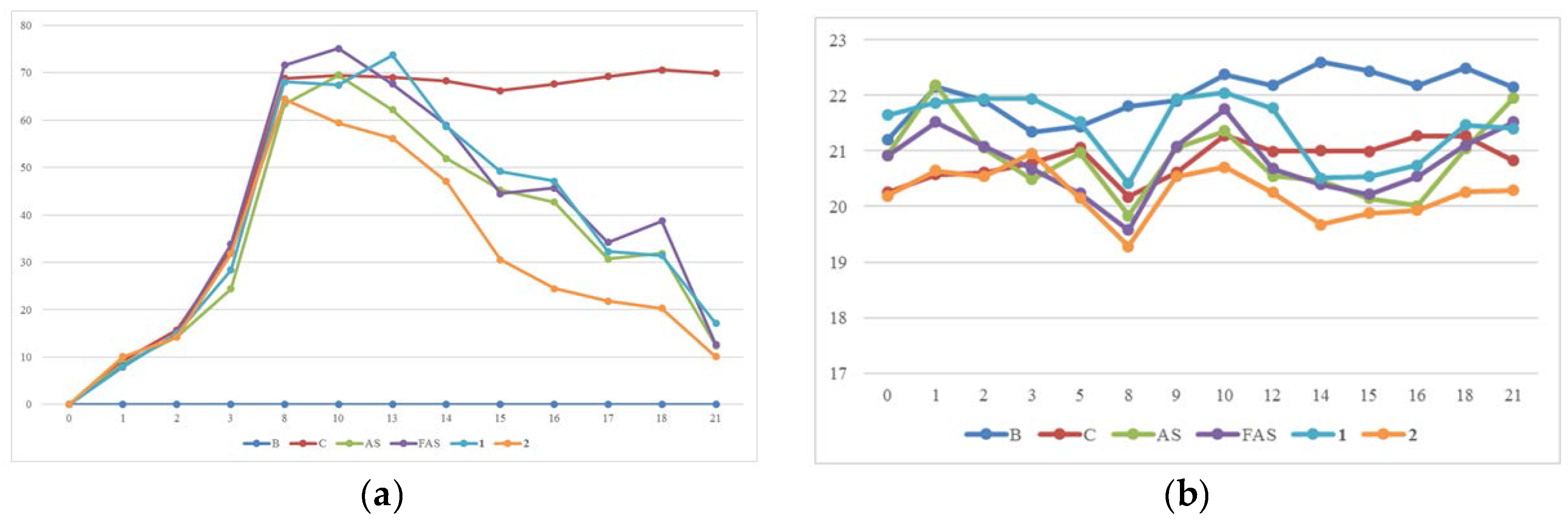
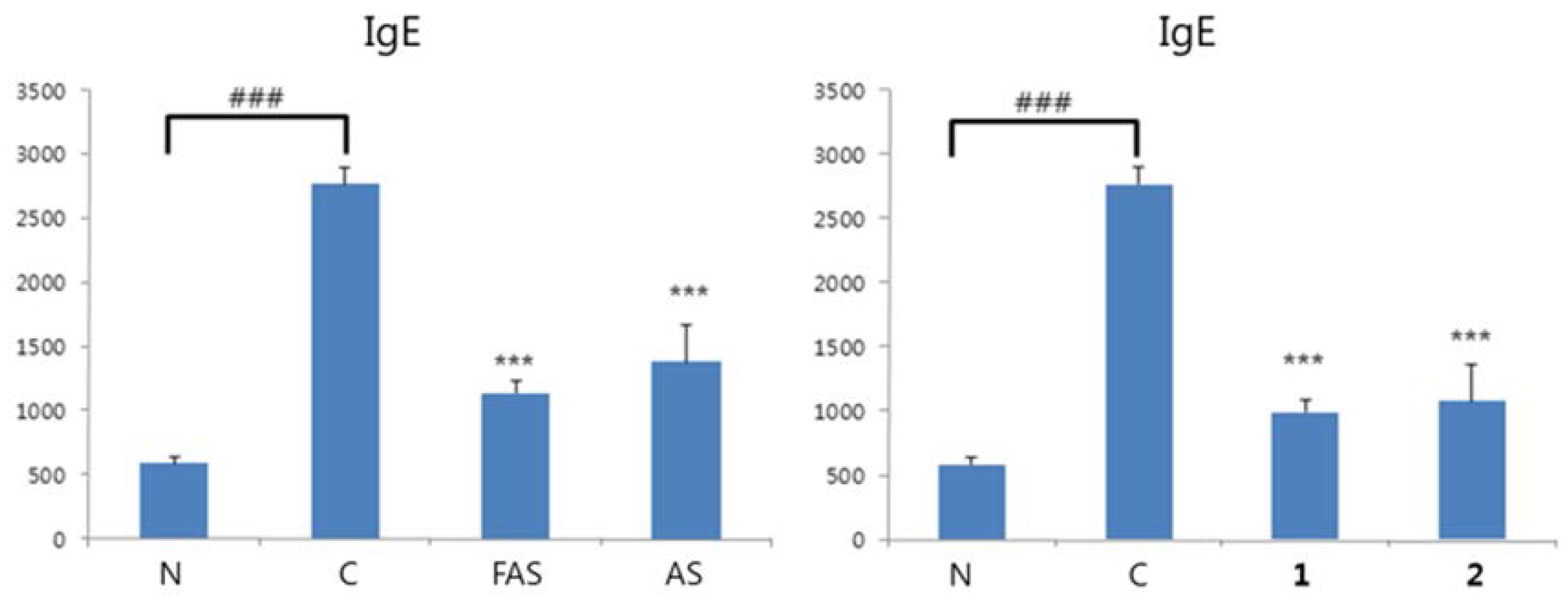
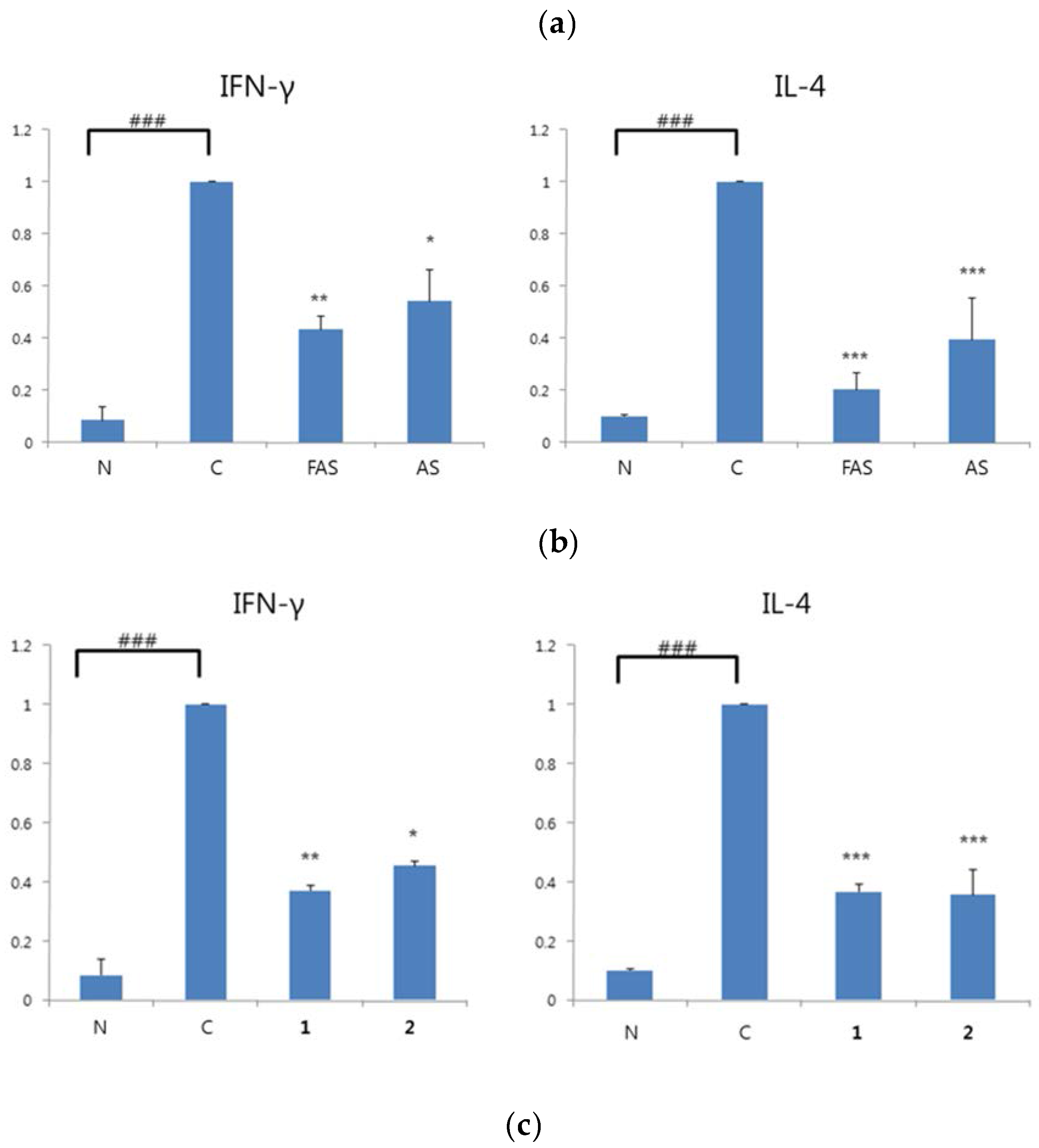
2.4. Anti-ACD/Anti-AD Evaluation on BALB/c Mice
3. Materials and Methods
3.1. Plant Material
3.2. Extraction, Fermentation and Isolation
3.3. Measuring DPPH Radical Scavenging Activity
3.4. Measuring NBT Superoxide Radical Scavenging Activity
3.5. Cell Culture
3.6. Viability Assay
3.7. Measuring NO Production Inhibition
3.8. Measuring Cytokine Production Inhibition
3.9. Animal Experiment Model
3.10. Preparing the Spleen Microsomes
3.11. Reverse Transcription RT-PCR
3.12. Statistical Analysis
4. Conclusions
Acknowledgments
Author Contributions
Conflicts of Interest
References
- Asher, M.I.; Montefort, S.; Bjorksten, B.; Lai, C.K.; Strachan, D.P.; Weiland, S.K.; Williams, H. ISAAC Phase Three Study Group. Worldwide Time Trends in the Prevalence of Symptoms of Asthma, Allergic Rhinoconjunctivitis, and Eczema in Childhood: ISAAC Phases One and Three Repeat Multicountry Cross-Sectional Surveys. Lancet 2006, 368, 733–743. [Google Scholar] [CrossRef]
- Barnetson, R.S.; Rogers, M. Childhood Atopic Eczema. BMJ 2002, 324, 1376–1379. [Google Scholar] [CrossRef] [PubMed]
- Grey, K.; Maguiness, S. Atopic Dermatitis: Update for Pediatricians. Pediatr. Ann. 2016, 45, e280–e286. [Google Scholar] [CrossRef] [PubMed]
- Simon, D.; Braathen, L.R.; Simon, H.U. Eosinophils and Atopic Dermatitis. Allergy 2004, 59, 561–570. [Google Scholar] [CrossRef] [PubMed]
- Hamid, Q.; Boguniewicz, M.; Leung, D.Y. Differential in Situ Cytokine Gene Expression in Acute Versus Chronic Atopic Dermatitis. J. Clin. Investig. 1994, 94, 870–876. [Google Scholar] [CrossRef] [PubMed]
- Leung, D.Y. Role of IgE in Atopic Dermatitis. Curr. Opin. Immunol. 1993, 5, 956–962. [Google Scholar] [CrossRef]
- Grewe, M.; Gyufko, K.; Schopf, E.; Krutmann, J. Lesional Expression of Interferon-Gamma in Atopic Eczema. Lancet 1994, 343, 25–26. [Google Scholar] [CrossRef]
- Chen, L.; Martinez, O.; Overbergh, L.; Mathieu, C.; Prabhakar, B.S.; Chan, L.S. Early Up-Regulation of Th2 Cytokines and Late Surge of Th1 Cytokines in an Atopic Dermatitis Model. Clin. Exp. Immunol. 2004, 138, 375–387. [Google Scholar] [CrossRef] [PubMed]
- Park, S.; Lee, J.B.; Kang, S. Topical Application of Chrysanthemum Indicum L. Attenuates the Development of Atopic Dermatitis-Like Skin Lesions by Suppressing Serum IgE Levels, IFN-γ, and IL-4 in Nc/Nga Mice. Evid Based. Complement. Alternat Med. 2012, 2012. [Google Scholar] [CrossRef]
- Usatine, R.P.; Riojas, M. Diagnosis and Management of Contact Dermatitis. Am. Fam. Physician 2010, 82, 249–255. [Google Scholar] [PubMed]
- Berke, R.; Singh, A.; Guralnick, M. Atopic Dermatitis: An Overview. Am. Fam. Physician 2012, 86, 35–42. [Google Scholar] [PubMed]
- Tabassum, N.; Hamdani, M. Plants used to Treat Skin Diseases. Pharmacogn. Rev. 2014, 8, 52–60. [Google Scholar] [CrossRef] [PubMed]
- Puupponen-Pimia, R.; Nohynek, L.; Juvonen, R.; Kosso, T.; Truchado, P.; Westerlund-Wikstrom, B.; Leppanen, T.; Moilanen, E.; Oksman-Caldentey, K.M. Fermentation and Dry Fractionation Increase Bioactivity of Cloudberry (Rubus Chamaemorus). Food Chem. 2016, 197, 950–958. [Google Scholar] [CrossRef] [PubMed]
- Zhang, L.X.; Gao, W.Y.; Wang, H.Y. Review of Traditional Chinese Medicine Processed by Fermentation. Zhongguo Zhong Yao Za Zhi 2012, 37, 3695–3700. [Google Scholar] [PubMed]
- Barbagallo, M.; Marotta, F.; Dominguez, L.J. Oxidative Stress in Patients with Alzheimer's Disease: Effect of Extracts of Fermented Papaya Powder. Mediators Inflamm. 2015, 2015, 624801. [Google Scholar] [CrossRef] [PubMed]
- Lee, S. (Ed.) Korea Folk Medicine; Seoul National University Publishing Center Press: Seoul, Korea, 1966. [Google Scholar]
- Pui-hay, B.P.; Sung, C.K. International Collation of Traditional and Folk Medicine; Kimura Takeatsu; World Scientific Publishing Company: Singapore, 2001. [Google Scholar]
- Kim, M.H.; Park, K.H.; Kim, S.R.; Park, K.J.; Oh, M.H.; Heo, J.H.; Yoon, K.H.; Yin, J.; Yoon, K.H.; Lee, M.W. Two New Phenolic Compounds from the Leaves of Alnus Sibirica Fisch. Ex Turcz. Nat. Prod. Res. 2016, 30, 206–213. [Google Scholar] [CrossRef] [PubMed]
- Choi, S.E.; Park, K.H.; Jeong, M.S.; Kim, H.H.; Lee, D.I.; Joo, S.S.; Lee, C.S.; Bang, H.; Choi, Y.W.; Lee, M.K.; et al. Effect of Alnus Japonica Extract on a Model of Atopic Dermatitis in NC/Nga Mice. J. Ethnopharmacol. 2011, 136, 406–413. [Google Scholar] [CrossRef] [PubMed]
- Jeong, M.S.; Choi, S.E.; Kim, J.Y.; Kim, J.S.; Kim, E.J.; Park, K.H.; Lee, D.I.; Joo, S.S.; Lee, C.S.; Bang, H.; et al. Atopic Dermatitis-Like Skin Lesions Reduced by Topical Application and Intraperitoneal Injection of Hirsutenone in NC/Nga Mice. Clin. Dev. Immunol. 2010, 2010. [Google Scholar] [CrossRef] [PubMed]
- Choi, S.E.; Jeong, M.S.; Kang, M.J.; Lee, D.I.; Joo, S.S.; Lee, C.S.; Bang, H.; Lee, M.K.; Myung, S.C.; Choi, Y.W.; et al. Effect of Topical Application and Intraperitoneal Injection of Oregonin on Atopic Dermatitis in NC/Nga Mice. Exp. Dermatol. 2010, 19, e37–e43. [Google Scholar] [CrossRef] [PubMed]
- Hwang, O. Role of Oxidative Stress in Parkinson's Disease. Exp. Neurobiol. 2013, 22, 11–17. [Google Scholar] [CrossRef] [PubMed]
- Halliwell, B. Oxidative Stress and Cancer: Have we Moved Forward? Biochem. J. 2007, 401, 1–11. [Google Scholar] [CrossRef] [PubMed]
- Valko, M.; Leibfritz, D.; Moncol, J.; Cronin, M.T.; Mazur, M.; Telser, J. Free Radicals and Antioxidants in Normal Physiological Functions and Human Disease. Int. J. Biochem. Cell Biol. 2007, 39, 44–84. [Google Scholar] [CrossRef] [PubMed]
- Sivaranjani, N.; Rao, S.V.; Rajeev, G. Role of Reactive Oxygen Species and Antioxidants in Atopic Dermatitis. J. Clin. Diagn. Res. 2013, 7, 2683–2685. [Google Scholar] [CrossRef] [PubMed]
- Lee, J.H.; Yeom, S.H.; Kim, M.K.; Kim, H.J.; Shim, J.G.; Lee, M.W. Antioxidative Activities of Diarylheptanoids from Alnus Japonica and their Structural Relationship. Korean J. Pharm. 2003, 34, 190–192. [Google Scholar]
- Hou, Y.C.; Janczuk, A.; Wang, P.G. Current Trends in the Development of Nitric Oxide Donors. Curr. Pharm. Des. 1999, 5, 417–441. [Google Scholar] [PubMed]
- Coleman, J.W. Nitric Oxide in Immunity and Inflammation. Int. Immunopharmacol. 2001, 1, 1397–1406. [Google Scholar] [CrossRef]
- Ryan, J.J. Interleukin-4 and its Receptor: Essential Mediators of the Allergic Response. J. Allergy Clin. Immunol. 1997, 99, 1–5. [Google Scholar] [CrossRef]
- Bacharier, L.B.; Geha, R.S. Molecular Mechanisms of IgE Regulation. J. Allergy Clin. Immunol. 2000, 105, S547–S558. [Google Scholar] [CrossRef]
- Jabara, H.H.; Ahern, D.J.; Vercelli, D.; Geha, R.S. Hydrocortisone and IL-4 Induce IgE Isotype Switching in Human B Cells. J. Immunol. 1991, 147, 1557–1560. [Google Scholar]
- Purkerson, J.M.; Isakson, P.C. Interleukin 5 (IL-5) Provides a Signal that is Required in Addition to IL-4 for Isotype Switching to Immunoglobulin (Ig) G1 and IgE. J. Exp. Med. 1992, 175, 973. [Google Scholar] [CrossRef] [PubMed]
- Kiniwa, M.; Gately, M.; Gubler, U.; Chizzonite, R.; Fargeas, C.; Delespesse, G. Recombinant Interleukin-12 Suppresses the Synthesis of Immunoglobulin E by Interleukin-4 Stimulated Human Lymphocytes. J. Clin. Investig. 1992, 90, 262–266. [Google Scholar] [CrossRef] [PubMed]
- Waage, A.; Brandtzaeg, P.; Halstensen, A.; Kierulf, P.; Espevik, T. The Complex Pattern of Cytokines in Serum from Patients with Meningococcal Septic Shock. Association between Interleukin 6, Interleukin 1, and Fatal Outcome. J. Exp. Med. 1989, 169, 333–338. [Google Scholar] [CrossRef] [PubMed]
- Slordal, L.; Warren, D.J.; Moore, M.A. Effect of Recombinant Murine Tumor Necrosis Factor on Hemopoietic Reconstitution in Sublethally Irradiated Mice. J. Immunol. 1989, 142, 833–835. [Google Scholar] [PubMed]
- Grau, G.E.; Piguet, P.; Vassalli, P.; Lambert, P. Involvement of Tumour Necrosis Factor and Other Cytokines in Immune-Mediated Vascular Pathology. Int. Arch. Allergy Immunol. 1989, 88, 34–39. [Google Scholar] [CrossRef]
- Scheurich, P.; Thoma, B.; Ucer, U.; Pfizenmaier, K. Immunoregulatory Activity of Recombinant Human Tumor Necrosis Factor (TNF)-Alpha: Induction of TNF Receptors on Human T Cells and TNF-Alpha-Mediated Enhancement of T Cell Responses. J. Immunol. 1987, 138, 1786. [Google Scholar] [PubMed]
- Behniafard, N.; Gharagozlou, M.; Farhadi, E.; Khaledi, M.; Sotoudeh, S.; Darabi, B.; Mohammad Fathi, S.; Gholizadeh Moghaddam, Z.; Mahmoudi, M.; Aghamohammadi, A.; et al. TNF-Alpha Single Nucleotide Polymorphisms in Atopic Dermatitis. Eur. Cytokine Netw. 2013, 23, 163–165. [Google Scholar]
- Sumimoto, S.; Kawai, M.; Kasailma, Y.; Hamamoto, T. Increased Plasma Tumour Necrosis Factor-Ut Concentration in Atopic Dermatitis. Arch. Dis. Child. 1992, 67, 277–279. [Google Scholar] [CrossRef]
- De Becker, G.; Moulin, V.; Tielemans, F.; De Mattia, F.; Urbain, J.; Leo, O.; Moser, M. Regulation of T Helper Cell Differentiation in Vivo by Soluble and Membrane Proteins Provided by Antigen-Presenting Cells. Eur. J. Immunol. 1998, 28, 3161–3171. [Google Scholar] [CrossRef]
- Blaher, B.; McCluskey, J.; Puy, R.; Czarny, D.; Rolland, J.M. T Cell Response to Grass Pollen Allergens: Correlation with Skin Test Reactivity and Serum IgE Levels. Immunol. Cell Biol. 1995, 73, 17–22. [Google Scholar] [CrossRef] [PubMed]
- Gober, M.D.; Fishelevich, R.; Zhao, Y.; Unutmaz, D.; Gaspari, A.A. Human Natural Killer T Cells Infiltrate into the Skin at Elicitation Sites of Allergic Contact Dermatitis. J. Investig. Dermatol. 2008, 128, 1460–1469. [Google Scholar] [CrossRef] [PubMed]
- Simon, D.; Kozlowski, E.; Simon, H. Natural Killer T Cells Expressing IFN-Gamma and IL-4 in Lesional Skin of Atopic Eczema. Allergy 2009, 64, 1681–1684. [Google Scholar] [CrossRef] [PubMed]
- Stalder, J.F.; Taïeb, A.; Atherton, D.J.; Bieber, P.; Bonifazi, E.; Broberg, A.; Calza, A.; Coleman, R.; De Prost, Y.; Stalder, J.F.; et al. Severity Scoring of Atopic Dermatitis: The SCORAD Index. Consensus Report of the European Task Force on Atopic Dermatitis. Dermatology 1993, 186, 23–31. [Google Scholar]
Sample Availability: Samples of the compounds and the extracts are available from the authors. |









{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Samples | DPPH Radical Scavenging Activity | NBT Radical Scavenging Activity | ||
|---|---|---|---|---|
| IC50 (μg/mL) | IC50 (μM) | IC50 (μg/mL) | IC50 (μM) | |
| FAS | 24.34 ± 0.9 | 1.11 ± 0.36 | ||
| AS | 31.41 ± 1.14 | 6.57 ± 0.44 | ||
| 1 | 14.08 ± 0.13 | 17.56 ± 0.29 | ||
| 2 | 24.08 ± 1.08 | 30.16 ± 2.85 | ||
| l-Ascorbic acid | 14.68 ± 0.36 | 24.81 ± 0.59 | ||
| Allopurinol | 0.95 ± 0.2 | 7.99 ± 0.66 | ||
| Samples | Inhibition of NO Production | |
|---|---|---|
| IC50 (μg/mL) | IC50 (μM) | |
| FAS | 13.00 ± 0.38 | |
| AS | 19.31 ± 1.22 | |
| 1 | 14.10 ± 0.20 | |
| 2 | 22.66 ± 1.18 | |
| L-NMMA | 2.22 ± 0.11 | 37.05 ± 1.21 |
© 2018 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Yin, J.; Yoon, S.H.; Ahn, H.S.; Lee, M.W. Inhibitory Activity of Allergic Contact Dermatitis and Atopic Dermatitis-Like Skin in BALB/c Mouse through Oral Administration of Fermented Barks of Alnus sibirica. Molecules 2018, 23, 450. https://doi.org/10.3390/molecules23020450
Yin J, Yoon SH, Ahn HS, Lee MW. Inhibitory Activity of Allergic Contact Dermatitis and Atopic Dermatitis-Like Skin in BALB/c Mouse through Oral Administration of Fermented Barks of Alnus sibirica. Molecules. 2018; 23(2):450. https://doi.org/10.3390/molecules23020450
Chicago/Turabian StyleYin, Jun, Seong Hye Yoon, Hye Shin Ahn, and Min Won Lee. 2018. "Inhibitory Activity of Allergic Contact Dermatitis and Atopic Dermatitis-Like Skin in BALB/c Mouse through Oral Administration of Fermented Barks of Alnus sibirica" Molecules 23, no. 2: 450. https://doi.org/10.3390/molecules23020450