
The Hammerhead Ribozyme: A Long History for a Short RNA



{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. The Discovery of the HHR in Infectious circRNAs of Plants
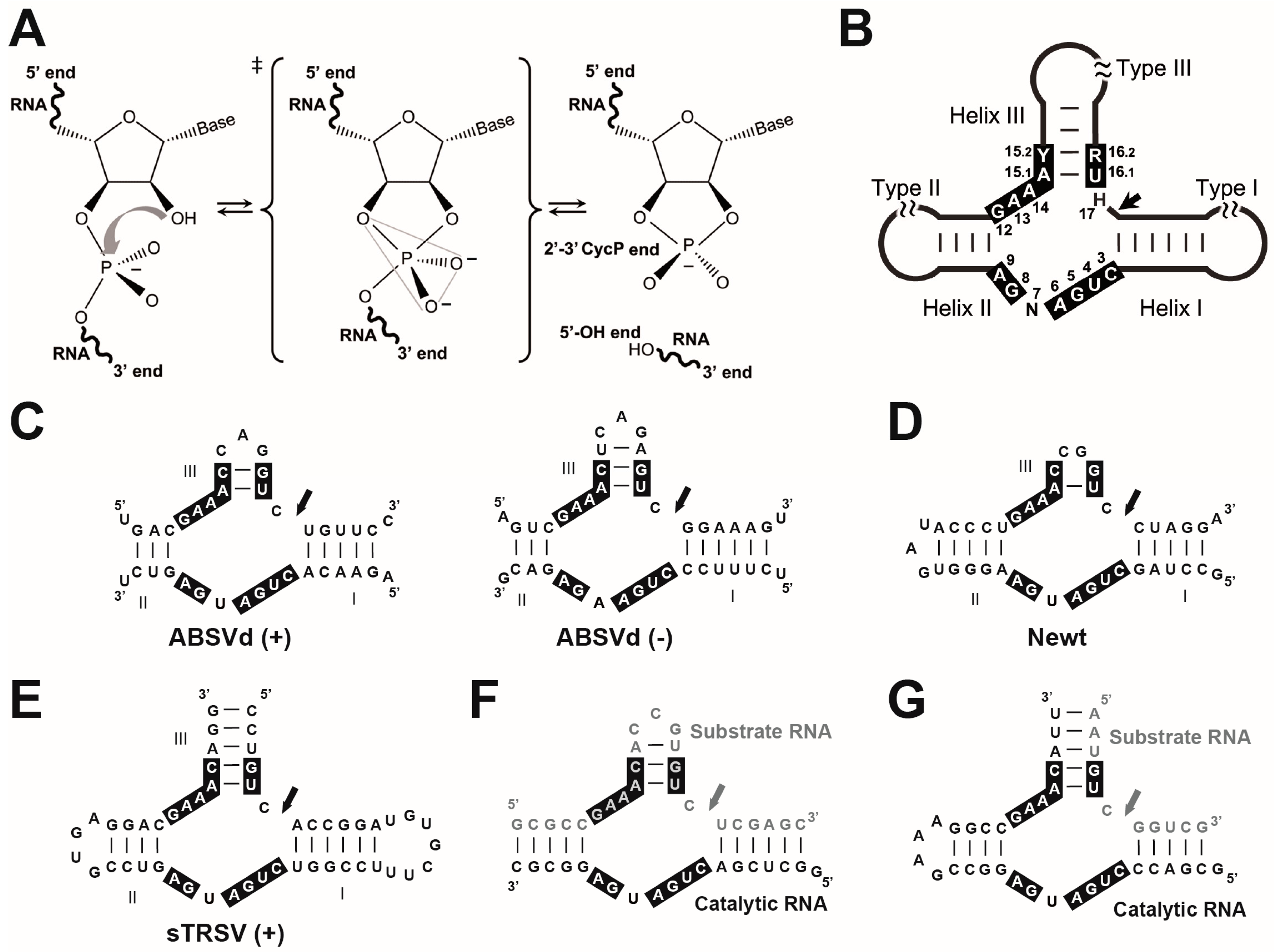
3. trans RNA Cleavage Using a Minimal HHR Motif: Too Minimal as a Model
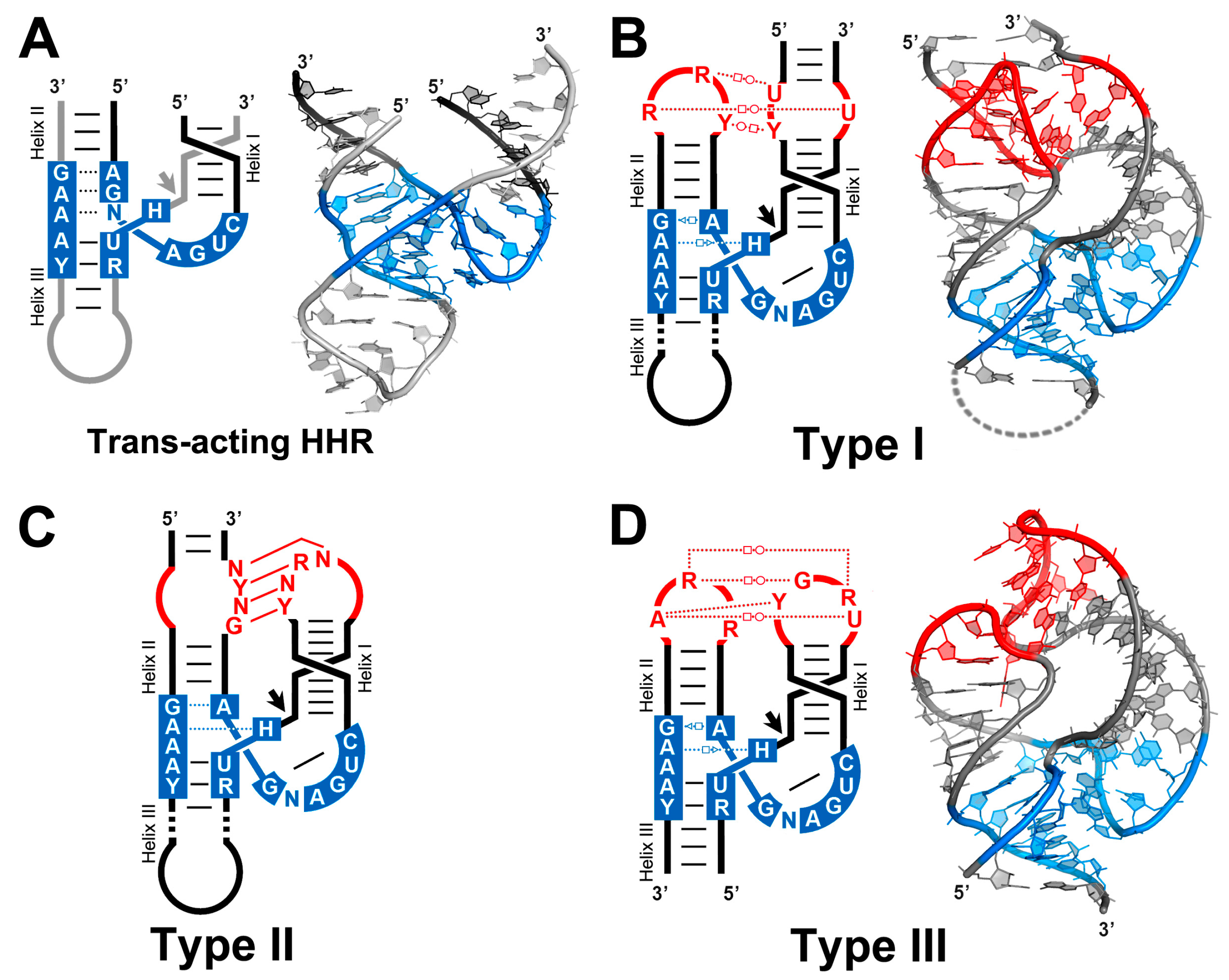
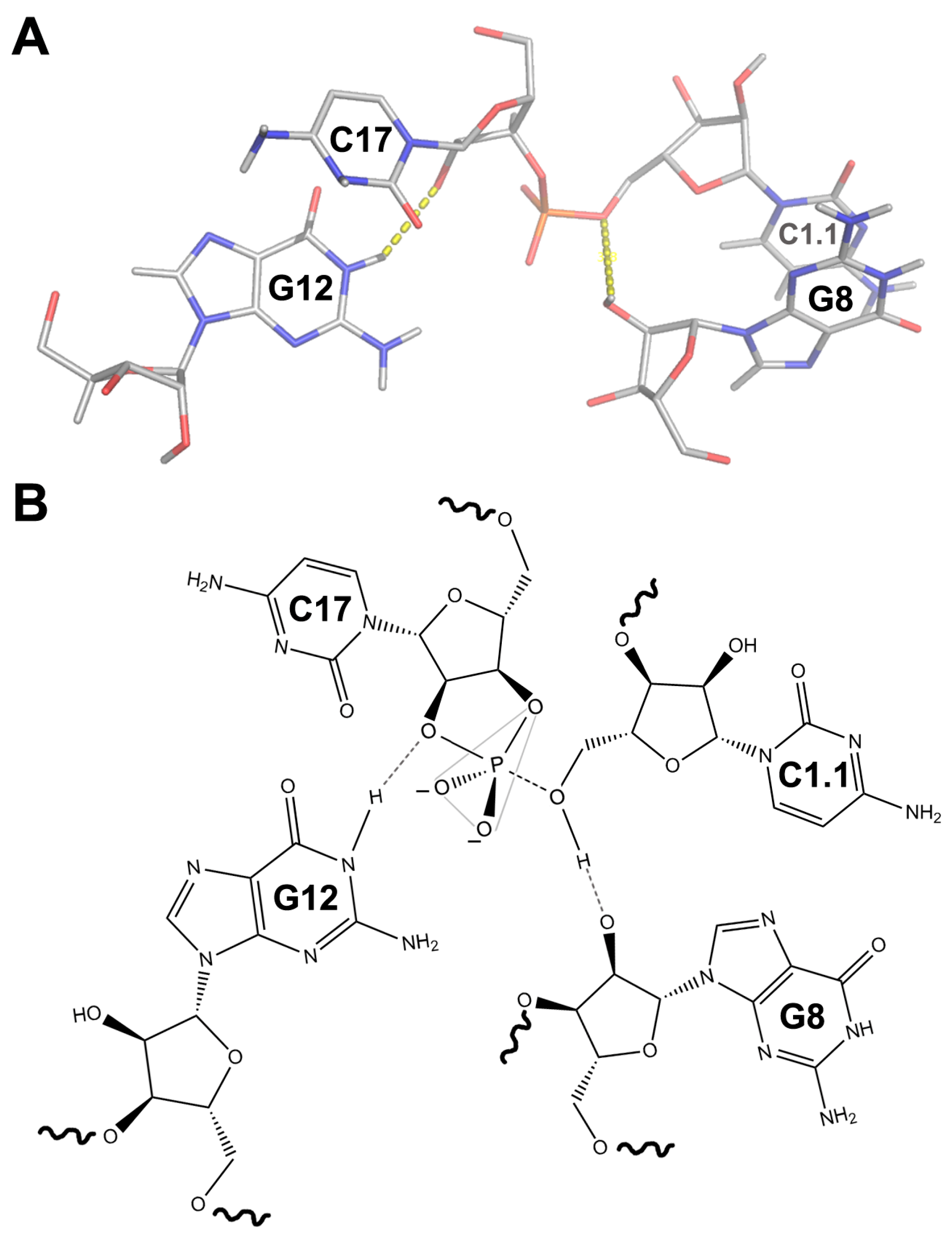
4. Tertiary Interactions of the HHR Allow for a More Efficient Self-Cleavage In Vivo
5. The Structure of a Full HHR
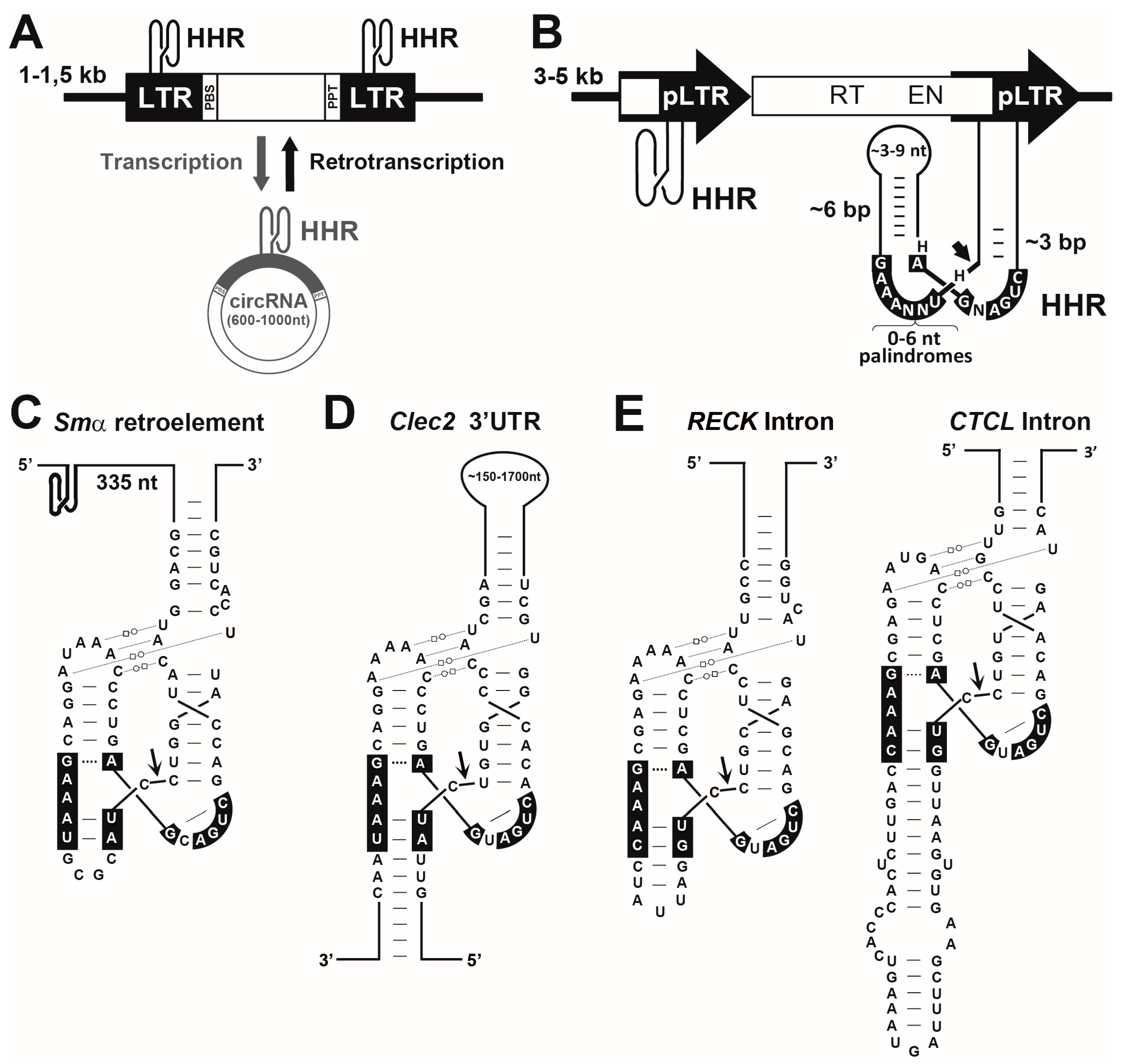
6. Widespread Occurrence of the HHR along the Tree of Life
7. RNA Self-Cleavage Catalysis in Biology: From Mobile Genetic Elements to Domesticated New Functions
8. Conclusions
Acknowledgments
Conflicts of Interest
References
- Sumner, J.B. The isolation and crystallization of the enzyme urease. J. Biol. Chem. 1926, 69, 435–441. [Google Scholar]
- Kruger, K.; Grabowski, P.J.; Zaug, A.J.; Sands, J.; Gottschling, D.E.; Cech, T.R. Self-splicing RNA: Autoexcision and autocyclization of the ribosomal RNA intervening sequence of Tetrahymena. Cell 1982, 31, 147–157. [Google Scholar] [CrossRef]
- Guerrier-Takada, C.; Gardiner, K.; Marsh, T.; Pace, N.; Altman, S. The RNA moiety of ribonuclease P is the catalytic subunit of the enzyme. Cell 1983, 35, 849–857. [Google Scholar] [CrossRef]
- Thomas, B.C.; Li, X.; Gegenheimer, P. Chloroplast ribonuclease P does not utilize the ribozyme-type pre-tRNA cleavage mechanism. RNA 2000, 6, 545–553. [Google Scholar] [CrossRef] [PubMed]
- Holzmann, J.; Frank, P.; Löffler, E.; Bennett, K.L.; Gerner, C.; Rossmanith, W. RNase P without RNA: Identification and functional reconstitution of the human mitochondrial tRNA processing enzyme. Cell 2008, 135, 462–474. [Google Scholar] [CrossRef] [PubMed]
- Prody, G.A.; Bakos, J.T.; Buzayan, J.M.; Schneider, I.R.; Bruening, G. Autolytic processing of dimeric plant virus satellite RNA. Science 1986, 231, 1577–1580. [Google Scholar] [CrossRef] [PubMed]
- Hutchins, C.J.; Rathjen, P.D.; Forster, A.C.; Symons, R.H. Self-cleavage of plus and minus RNA transcripts of avocado sunblotch viroid. Nucleic Acids Res. 1986, 14, 3627–3640. [Google Scholar] [CrossRef] [PubMed]
- Cech, T.R. Structural biology. The ribosome is a ribozyme. Science 2000, 289, 878–879. [Google Scholar] [CrossRef] [PubMed]
- Gilbert, W. The RNA World. Nature 1986, 319, 618. [Google Scholar] [CrossRef]
- Emilsson, G.M.; Nakamura, S.; Roth, A.; Breaker, R.R. Ribozyme speed limits. RNA 2003, 9, 907–918. [Google Scholar] [CrossRef] [PubMed]
- Canny, M.D.; Jucker, F.M.; Kellogg, E.; Khvorova, A.; Jayasena, S.D.; Pardi, A. Fast cleavage kinetics of a natural hammerhead ribozyme. J. Am. Chem. Soc. 2004, 126, 10848–10849. [Google Scholar] [CrossRef] [PubMed]
- Zamel, R.; Poon, A.; Jaikaran, D.; Andersen, A.; Olive, J.; De Abreu, D.; Collins, R.A. Exceptionally fast self-cleavage by a Neurospora Varkud satellite ribozyme. Proc. Natl. Acad. Sci. USA 2004, 101, 1467–1472. [Google Scholar] [CrossRef] [PubMed]
- Cochrane, J.C.; Strobel, S.A. Catalytic strategies of self-cleaving ribozymes. Acc. Chem. Res. 2008, 41, 1027–1035. [Google Scholar] [CrossRef] [PubMed]
- Thompson, J.E.; Kutateladze, T.G.; Schuster, M.C.; Venegas, F.D.; Messmore, J.M.; Raines, R.T. Limits to Catalysis by Ribonuclease A. Bioorg. Chem. 1995, 23, 471–481. [Google Scholar] [CrossRef] [PubMed]
- Buzayan, J.M.; Gerlach, W.L.; Bruening, G. Non-enzymatic cleavage and ligation of RNAs complementary to a plant virus satellite RNA. Nature 1986, 323, 349–353. [Google Scholar] [CrossRef]
- Kuo, M.Y.; Sharmeen, L.; Dinter-Gottlieb, G.; Taylor, J. Characterization of self-cleaving RNA sequences on the genome and antigenome of human hepatitis delta virus. J. Virol. 1988, 62, 4439–4444. [Google Scholar] [PubMed]
- Saville, B.J.; Collins, R.A. A site-specific self-cleavage reaction performed by a novel RNA in Neurospora mitochondria. Cell 1990, 61, 685–696. [Google Scholar] [CrossRef]
- Winkler, W.C.; Nahvi, A.; Roth, A.; Collins, J.A.; Breaker, R.R. Control of gene expression by a natural metabolite-responsive ribozyme. Nature 2004, 428, 281–286. [Google Scholar] [CrossRef] [PubMed]
- Roth, A.; Weinberg, Z.; Chen, A.G.; Kim, P.B.; Ames, T.D.; Breaker, R.R. A widespread self-cleaving ribozyme class is revealed by bioinformatics. Nat. Chem. Biol. 2014, 10, 56–60. [Google Scholar] [CrossRef] [PubMed]
- Weinberg, Z.; Kim, P.B.; Chen, T.H.; Li, S.; Harris, K.A.; Lunse, C.E.; Breaker, R.R. New classes of self-cleaving ribozymes revealed by comparative genomics analysis. Nat. Chem. Biol. 2015, 11, 606–610. [Google Scholar] [CrossRef] [PubMed]
- Ferré-D’Amaré, A.R.; Scott, W.G. Small self-cleaving ribozymes. Cold Spring Harb. Perspect. Biol. 2010, 2, a003574. [Google Scholar] [CrossRef]
- Diener, T.O. Potato spindle tuber “virus”. IV. A replicating, low molecular weight RNA. Virology 1971, 45, 411–428. [Google Scholar] [CrossRef]
- Linthorst, H.J.; Kaper, J.M. Circular satellite-RNA molecules in satellite of tobacco ringspot virus-infected tissue. Virology 1984, 137, 206–210. [Google Scholar] [CrossRef]
- Epstein, L.M.; Gall, J.G. Self-cleaving transcripts of satellite DNA from the newt. Cell 1987, 48, 535–543. [Google Scholar] [CrossRef]
- Forster, A.C.; Davies, C.; Sheldon, C.C.; Jeffries, A.C.; Symons, R.H. Self-cleaving viroid and newt RNAs may only be active as dimers. Nature 1988, 334, 265–267. [Google Scholar] [CrossRef] [PubMed]
- Forster, A.C.; Symons, R.H. Self-cleavage of virusoid RNA is performed by the proposed 55-nucleotide active site. Cell 1987, 50, 9–16. [Google Scholar] [CrossRef]
- Forster, A.C.; Symons, R.H. Self-cleavage of plus and minus RNAs of a virusoid and a structural model for the active sites. Cell 1987, 49, 211–220. [Google Scholar] [CrossRef]
- De la Peña, M.; Flores, R. An extra nucleotide in the consensus catalytic core of a viroid hammerhead ribozyme: Implications for the design of more efficient ribozymes. J. Biol. Chem. 2001, 276, 34586–34593. [Google Scholar] [CrossRef] [PubMed]
- Shepotinovskaya, I.V.; Uhlenbeck, O.C. Catalytic diversity of extended hammerhead ribozymes. Biochemistry 2008, 47, 7034–7042. [Google Scholar] [CrossRef] [PubMed]
- Carbonell, A.; De la Peña, M.; Flores, R.; Gago, S. Effects of the trinucleotide preceding the self-cleavage site on eggplant latent viroid hammerheads: Differences in co- and post-transcriptional self-cleavage may explain the lack of trinucleotide AUC in most natural hammerheads. Nucleic Acids Res. 2006, 34, 5613–5622. [Google Scholar] [CrossRef] [PubMed]
- Uhlenbeck, O.C. A small catalytic oligoribonucleotide. Nature 1987, 328, 596–600. [Google Scholar] [CrossRef] [PubMed]
- Kim, R.; Holbrook, E.L.; Jancarik, J.; Pandit, J.; Weng, X.; Bohm, A.; Kim, S.H. High-resolution crystals and preliminary X-ray diffraction studies of a catalytic RNA. Acta Crystallogr. D Biol. Crystallogr. 1994, 50, 290–292. [Google Scholar] [CrossRef] [PubMed]
- Pley, H.W.; Flaherty, K.M.; McKay, D.B. Three-dimensional structure of a hammerhead ribozyme. Nature 1994, 372, 68–74. [Google Scholar] [CrossRef] [PubMed]
- Scott, W.G.; Finch, J.T.; Klug, A. The crystal structure of an all-RNA hammerhead ribozyme: A proposed mechanism for RNA catalytic cleavage. Cell 1995, 81, 991–1002. [Google Scholar] [CrossRef]
- Doudna, J.A. Hammerhead ribozyme structure: U-turn for RNA structural biology. Structure 1995, 3, 747–750. [Google Scholar] [CrossRef]
- Robertus, J.D.; Ladner, J.E.; Finch, J.T.; Rhodes, D.; Brown, R.S.; Clark, B.F.; Klug, A. Structure of yeast phenylalanine tRNA at 3 A resolution. Nature 1974, 250, 546–551. [Google Scholar] [CrossRef] [PubMed]
- Murray, J.B.; Seyhan, A.A.; Walter, N.G.; Burke, J.M.; Scott, W.G. The hammerhead, hairpin and VS ribozymes are catalytically proficient in monovalent cations alone. Chem. Biol. 1998, 5, 587–595. [Google Scholar] [CrossRef]
- Yang, H.; Jossinet, F.; Leontis, N.; Chen, L.; Westbrook, J.; Berman, H.; Westhof, E. Tools for the automatic identification and classification of RNA base pairs. Nucleic Acids Res. 2003, 31, 3450–3460. [Google Scholar] [CrossRef] [PubMed]
- McKay, D.B. Structure and function of the hammerhead ribozyme: An unfinished story. RNA 1996, 2, 395–403. [Google Scholar] [PubMed]
- Epstein, L.M.; Pabón-Peña, L.M. Alternative modes of self-cleavage by newt satellite 2 transcripts. Nucleic Acids Res. 1991, 19, 1699–1705. [Google Scholar] [CrossRef] [PubMed]
- Pabón-Peña, L.M.; Zhang, Y.; Epstein, L.M. Newt satellite 2 transcripts self-cleave by using an extended hammerhead structure. Mol. Cell. Biol. 1991, 11, 6109–6115. [Google Scholar] [CrossRef] [PubMed]
- Garrett, T.A.; Pabón-Peña, L.M.; Gokaldas, N.; Epstein, L.M. Novel requirements in peripheral structures of the extended satellite 2 hammerhead. RNA 1996, 2, 699–706. [Google Scholar] [PubMed]
- Miller, W.A.; Silver, S.L. Alternative tertiary structure attenuates self-cleavage of the ribozyme in the satellite RNA of barley yellow dwarf virus. Nucleic Acids Res. 1991, 19, 5313–5320. [Google Scholar] [CrossRef] [PubMed]
- De la Peña, M.; Gago, S.; Flores, R. Peripheral regions of natural hammerhead ribozymes greatly increase their self-cleavage activity. EMBO J. 2003, 22, 5561–5570. [Google Scholar] [CrossRef] [PubMed]
- Khvorova, A.; Lescoute, A.; Westhof, E.; Jayasena, S.D. Sequence elements outside the hammerhead ribozyme catalytic core enable intracellular activity. Nat. Struct. Biol. 2003, 10, 708–712. [Google Scholar] [CrossRef] [PubMed]
- Ferbeyre, G.; Smith, J.M.; Cedergren, R. Schistosome satellite DNA encodes active hammerhead ribozymes. Mol. Cell. Biol. 1998, 18, 3880–3888. [Google Scholar] [CrossRef] [PubMed]
- Canny, M.D.; Jucker, F.M.; Pardi, A. Efficient ligation of the Schistosoma hammerhead ribozyme. Biochemistry 2007, 46, 3826–3834. [Google Scholar] [CrossRef] [PubMed]
- Martick, M.; Scott, W.G. Tertiary contacts distant from the active site prime a ribozyme for catalysis. Cell 2006, 126, 309–320. [Google Scholar] [CrossRef] [PubMed]
- Pauling, L. Molecular architecture and biological reactions. Chem. Eng. News 1946, 24, 1375–1377. [Google Scholar] [CrossRef]
- Narlikar, G.J.; Herschlag, D. Mechanistic aspects of enzymatic catalysis: Lessons from comparison of RNA and protein enzymes. Annu. Rev. Biochem. 1997, 66, 19–59. [Google Scholar] [CrossRef] [PubMed]
- Chi, Y.I.; Martick, M.; Lares, M.; Kim, R.; Scott, W.G.; Kim, S.H. Capturing hammerhead ribozyme structures in action by modulating general base catalysis. PLoS Biol. 2008, 6, e234. [Google Scholar] [CrossRef] [PubMed]
- Dufour, D.; De la Peña, M.; Gago, S.; Flores, R.; Gallego, J. Structure-function analysis of the ribozymes of chrysanthemum chlorotic mottle viroid: A loop-loop interaction motif conserved in most natural hammerheads. Nucleic Acids Res. 2009, 37, 368–381. [Google Scholar] [CrossRef] [PubMed]
- De la Peña, M.; Dufour, D.; Gallego, J. Three-way RNA junctions with remote tertiary contacts: A recurrent and highly versatile fold. RNA 2009, 15, 1949–1964. [Google Scholar] [CrossRef] [PubMed]
- Symons, R.H.; Hutchins, C.J.; Forster, A.C.; Rathjen, P.D.; Keese, P.; Visvader, J.E. Self-cleavage of RNA in the replication of viroids and virusoids. J. Cell Sci. 1987, 1987 (Suppl. 7), 303–318. [Google Scholar] [CrossRef]
- Zhang, Y.; Epstein, L.M. Cloning and characterization of extended hammerheads from a diverse set of caudate amphibians. Gene 1996, 172, 183–190. [Google Scholar] [CrossRef]
- Cremisi, F.; Scarabino, D.; Carluccio, M.A.; Salvadori, P.; Barsacchi, G. A newt ribozyme: A catalytic activity in search of a function. Proc. Natl. Acad. Sci. USA 1992, 89, 1651–1655. [Google Scholar] [CrossRef] [PubMed]
- Daròs, J.A.; Flores, R. Identification of a retroviroid-like element from plants. Proc. Natl. Acad. Sci. USA 1995, 92, 6856–6860. [Google Scholar] [CrossRef] [PubMed]
- Przybilski, R.; Graf, S.; Lescoute, A.; Nellen, W.; Westhof, E.; Steger, G.; Hammann, C. Functional hammerhead ribozymes naturally encoded in the genome of Arabidopsis thaliana. Plant. Cell 2005, 17, 1877–1885. [Google Scholar] [CrossRef] [PubMed]
- Rojas, A.A.; Vazquez-Tello, A.; Ferbeyre, G.; Venanzetti, F.; Bachmann, L.; Paquin, B.; Sbordoni, V.; Cedergren, R. Hammerhead-mediated processing of satellite pDo500 family transcripts from Dolichopoda cave crickets. Nucleic Acids Res. 2000, 28, 4037–4043. [Google Scholar] [CrossRef] [PubMed]
- Bourdeau, V.; Ferbeyre, G.; Pageau, M.; Paquin, B.; Cedergren, R. The distribution of RNA motifs in natural sequences. Nucleic Acids Res. 1999, 27, 4457–4467. [Google Scholar] [CrossRef] [PubMed]
- Hammann, C.; Westhof, E. Searching genomes for ribozymes and riboswitches. Genome Biol. 2007, 8, 210. [Google Scholar] [CrossRef] [PubMed]
- De la Peña, M.; García-Robles, I. Ubiquitous presence of the hammerhead ribozyme motif along the tree of life. RNA 2010, 16, 1943–1950. [Google Scholar] [CrossRef] [PubMed]
- Jimenez, R.M.; Delwart, E.; Lupták, A. Structure-based search reveals hammerhead ribozymes in the human microbiome. J. Biol. Chem. 2011, 286, 7737–7743. [Google Scholar] [CrossRef] [PubMed]
- Perreault, J.; Weinberg, Z.; Roth, A.; Popescu, O.; Chartrand, P.; Ferbeyre, G.; Breaker, R.R. Identification of hammerhead ribozymes in all domains of life reveals novel structural variations. PLoS Comput. Biol. 2011, 7, e1002031. [Google Scholar] [CrossRef] [PubMed]
- Seehafer, C.; Kalweit, A.; Steger, G.; Gräf, S.; Hammann, C. From alpaca to zebrafish: Hammerhead ribozymes wherever you look. RNA 2011, 17, 21–26. [Google Scholar] [CrossRef] [PubMed]
- De la Peña, M.; García-Robles, I. Intronic hammerhead ribozymes are ultraconserved in the human genome. EMBO Rep. 2010, 11, 711–716. [Google Scholar] [CrossRef] [PubMed]
- Cervera, A.; Urbina, D.; De la Peña, M. Retrozymes are a unique family of non-autonomous retrotransposons with hammerhead ribozymes that propagate in plants through circular RNAs. Genome Biol. 2016, 17, 135. [Google Scholar] [CrossRef] [PubMed]
- Evgen’ev, M.B.; Zelentsova, H.; Shostak, N.; Kozitsina, M.; Barskyi, V.; Lankenau, D.H.; Corces, V.G. Penelope, a new family of transposable elements and its possible role in hybrid dysgenesis in Drosophila virilis. Proc. Natl. Acad. Sci. USA 1997, 94, 196–201. [Google Scholar] [CrossRef] [PubMed]
- Gladyshev, E.A.; Arkhipova, I.R. Telomere-associated endonuclease-deficient Penelope-like retroelements in diverse eukaryotes. Proc. Natl. Acad. Sci. USA 2007, 104, 9352–9357. [Google Scholar] [CrossRef] [PubMed]
- Ruminski, D.J.; Webb, C.H.; Riccitelli, N.J.; Lupták, A. Processing and translation initiation of non-long terminal repeat retrotransposons by hepatitis delta virus (HDV)-like self-cleaving ribozymes. J. Biol. Chem. 2011, 286, 41286–41295. [Google Scholar] [CrossRef] [PubMed]
- Eickbush, D.G.; Eickbush, T.H. R2 retrotransposons encode a self-cleaving ribozyme for processing from an rRNA cotranscript. Mol. Cell. Biol. 2010, 30, 3142–3150. [Google Scholar] [CrossRef] [PubMed]
- Kennell, J.C.; Saville, B.J.; Mohr, S.; Kuiper, M.T.; Sabourin, J.R.; Collins, R.A.; Lambowitz, A.M. The VS catalytic RNA replicates by reverse transcription as a satellite of a retroplasmid. Genes Dev. 1995, 9, 294–303. [Google Scholar] [CrossRef] [PubMed]
- Martick, M.; Horan, L.H.; Noller, H.F.; Scott, W.G. A discontinuous hammerhead ribozyme embedded in a mammalian messenger RNA. Nature 2008, 454, 899–902. [Google Scholar] [CrossRef] [PubMed]
- Scott, W.G.; Horan, L.H.; Martick, M. The hammerhead ribozyme: Structure, catalysis, and gene regulation. Prog. Mol. Biol. Transl. Sci. 2013, 120, 1–23. [Google Scholar] [PubMed]
- García-Robles, I.; Sánchez-Navarro, J.; De la Peña, M. Intronic hammerhead ribozymes in mRNA biogenesis. Biol. Chem. 2012, 393, 1317–1326. [Google Scholar] [CrossRef] [PubMed]




© 2017 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC-BY) license ( http://creativecommons.org/licenses/by/4.0/).
Share and Cite
De la Peña, M.; García-Robles, I.; Cervera, A. The Hammerhead Ribozyme: A Long History for a Short RNA. Molecules 2017, 22, 78. https://doi.org/10.3390/molecules22010078
De la Peña M, García-Robles I, Cervera A. The Hammerhead Ribozyme: A Long History for a Short RNA. Molecules. 2017; 22(1):78. https://doi.org/10.3390/molecules22010078
Chicago/Turabian StyleDe la Peña, Marcos, Inmaculada García-Robles, and Amelia Cervera. 2017. "The Hammerhead Ribozyme: A Long History for a Short RNA" Molecules 22, no. 1: 78. https://doi.org/10.3390/molecules22010078