
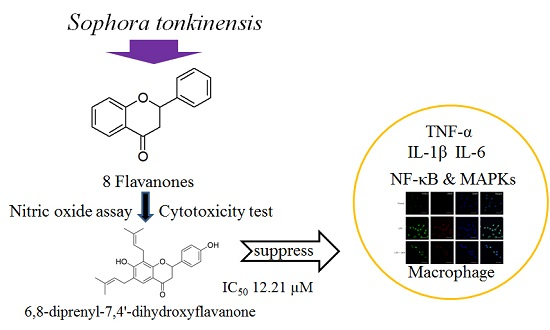
Anti-Inflammatory Effects of 6,8-Diprenyl-7,4′-dihydroxyflavanone from Sophora tonkinensis on Lipopolysaccharide-Stimulated RAW 264.7 Cells


 ,
, 
Abstract
:
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
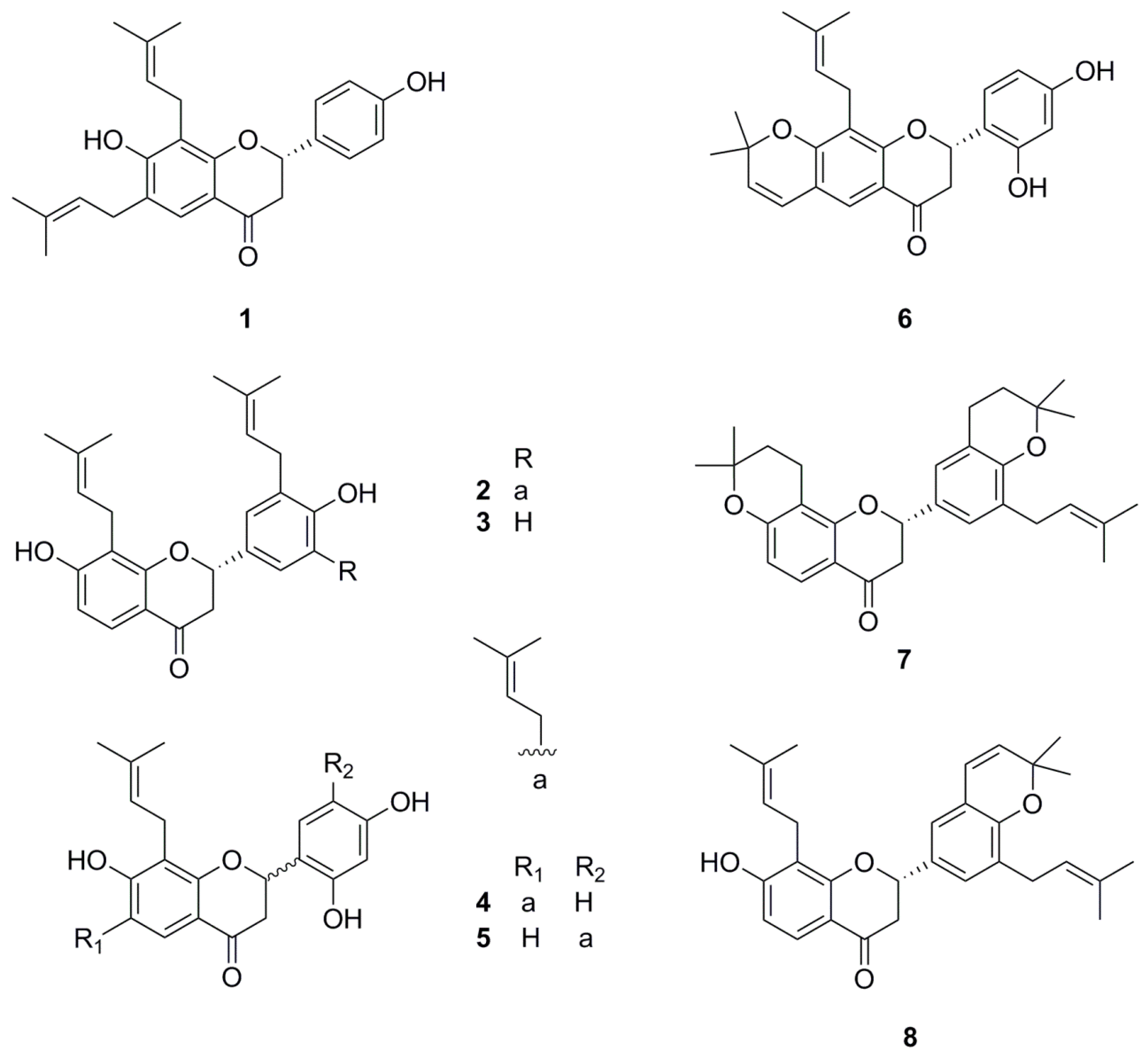
1. Introduction
2. Results
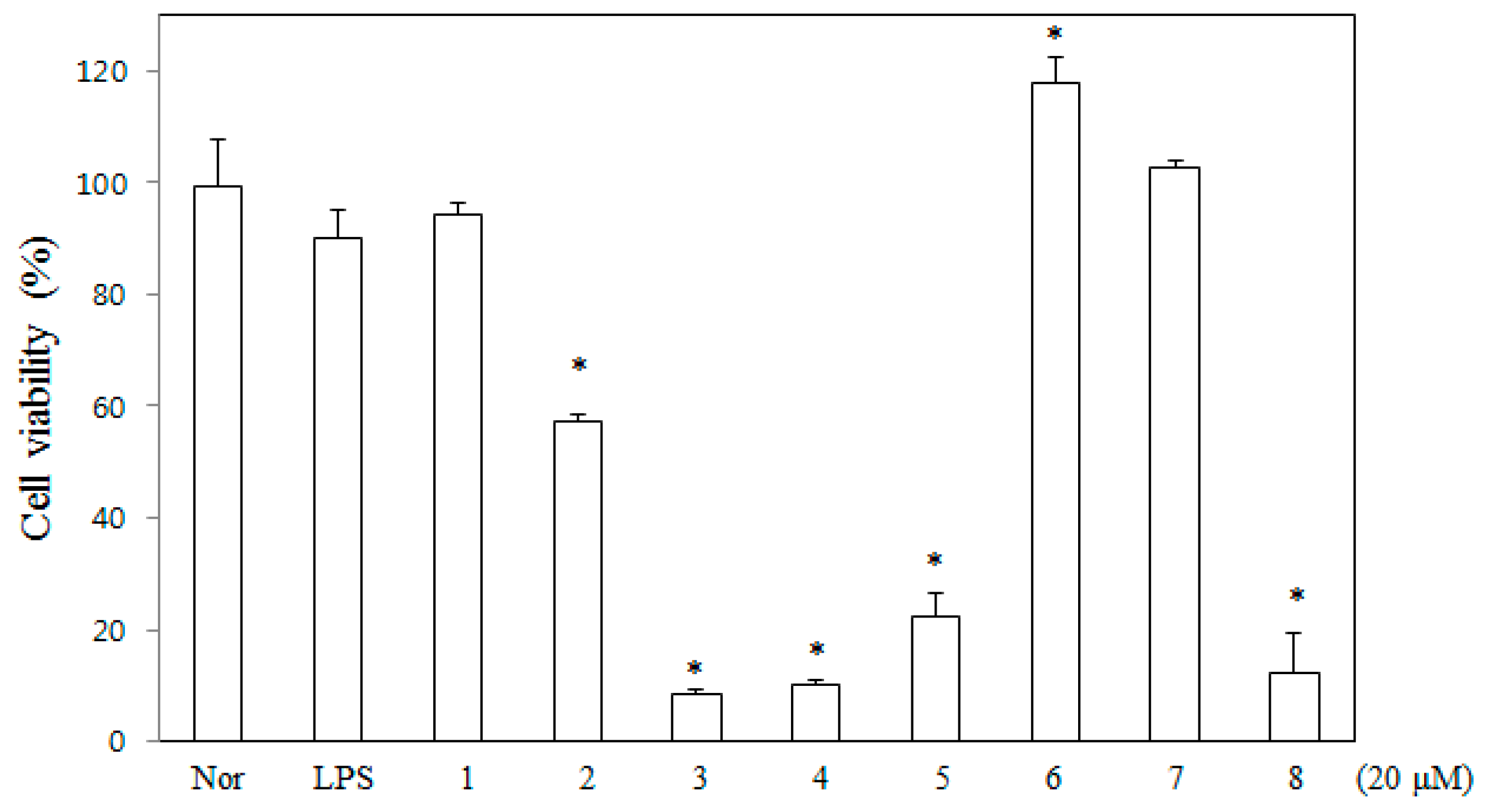
2.1. Toxicity of DDF on Macrophages
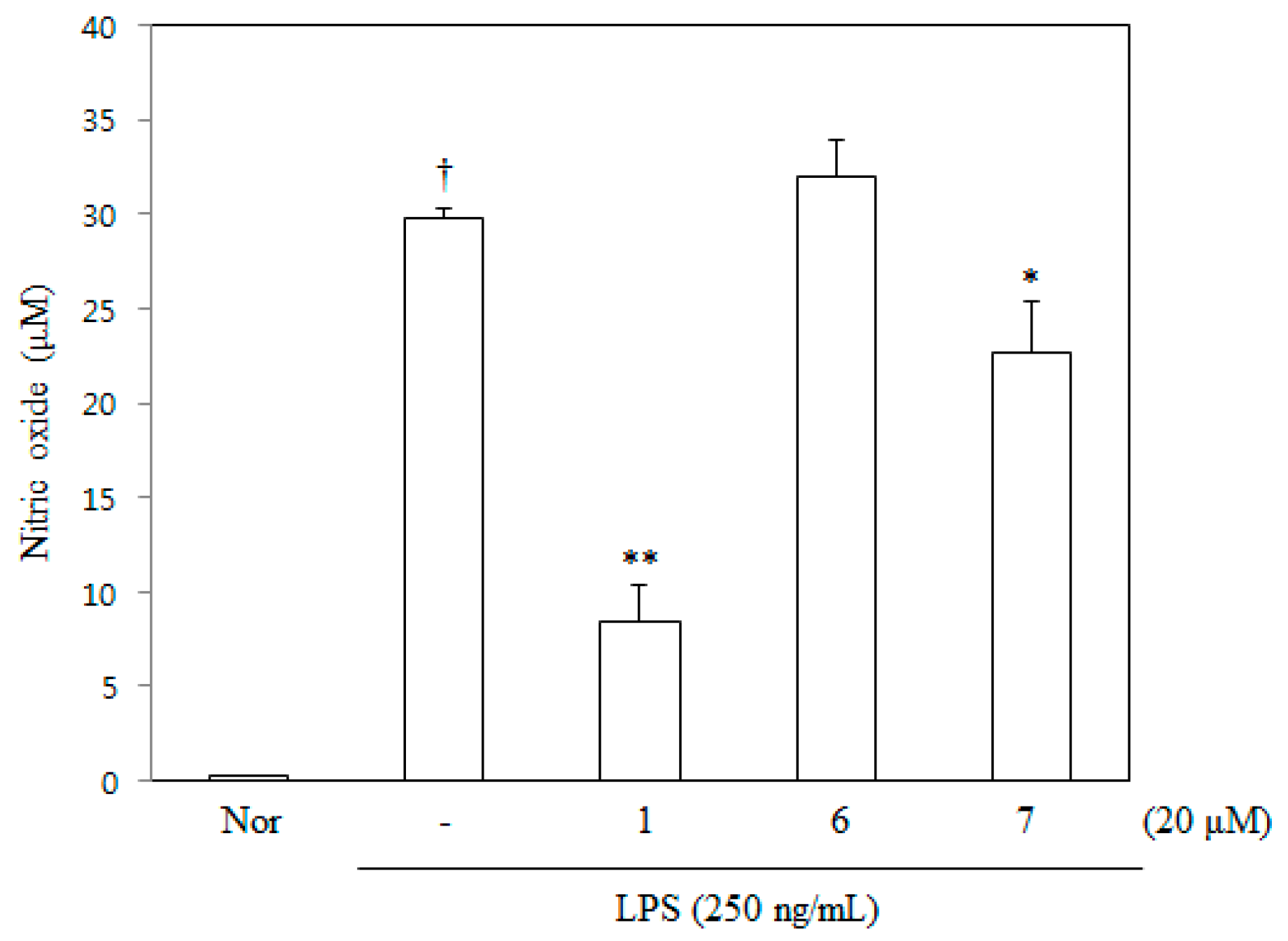
2.2. NO Inhibitory Activity of DDF on RAW 264.7 Cells
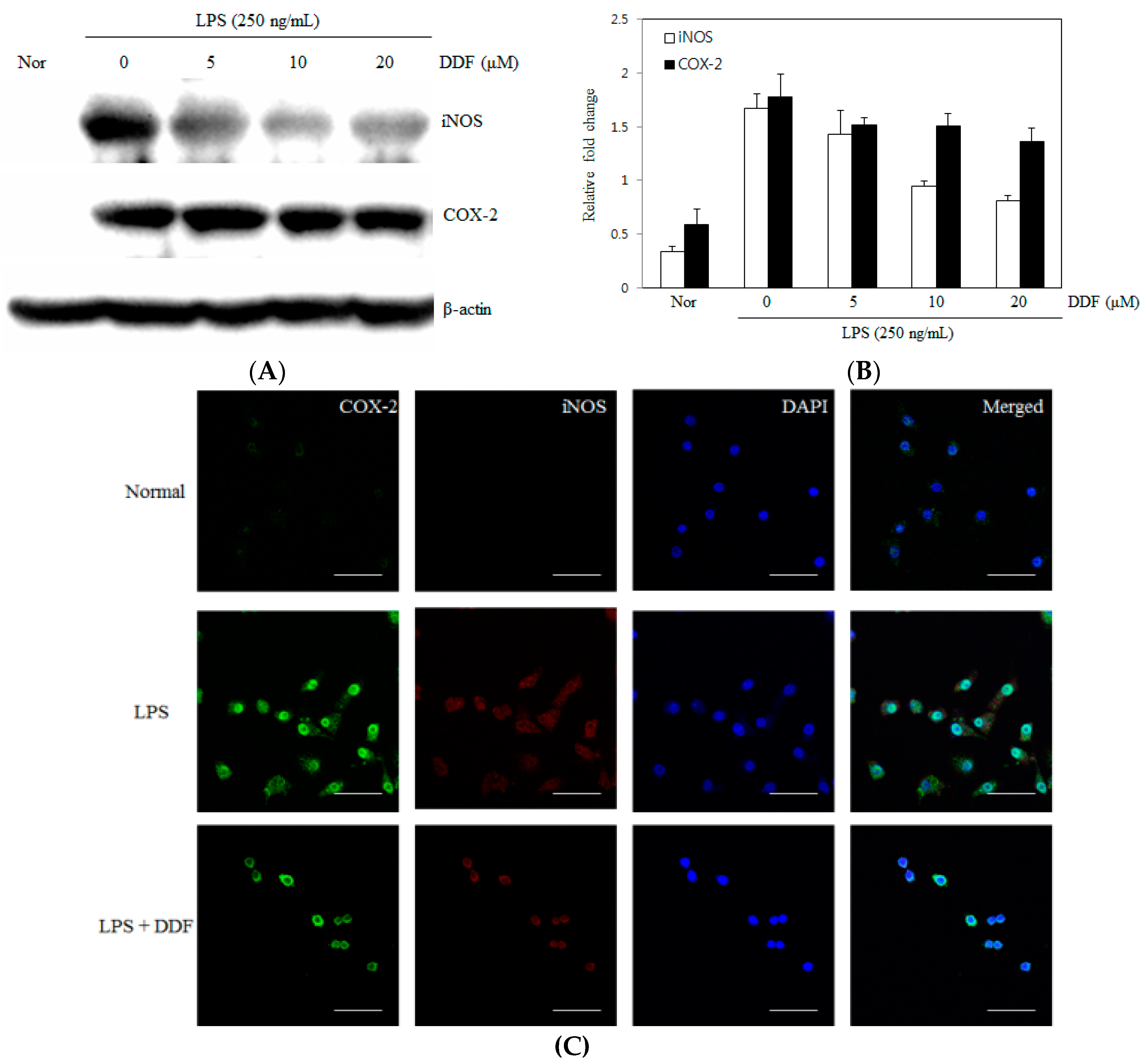
2.3. Effect of DDF on iNOS and COX-2 Expression Levels in RAW 264.7 Cells
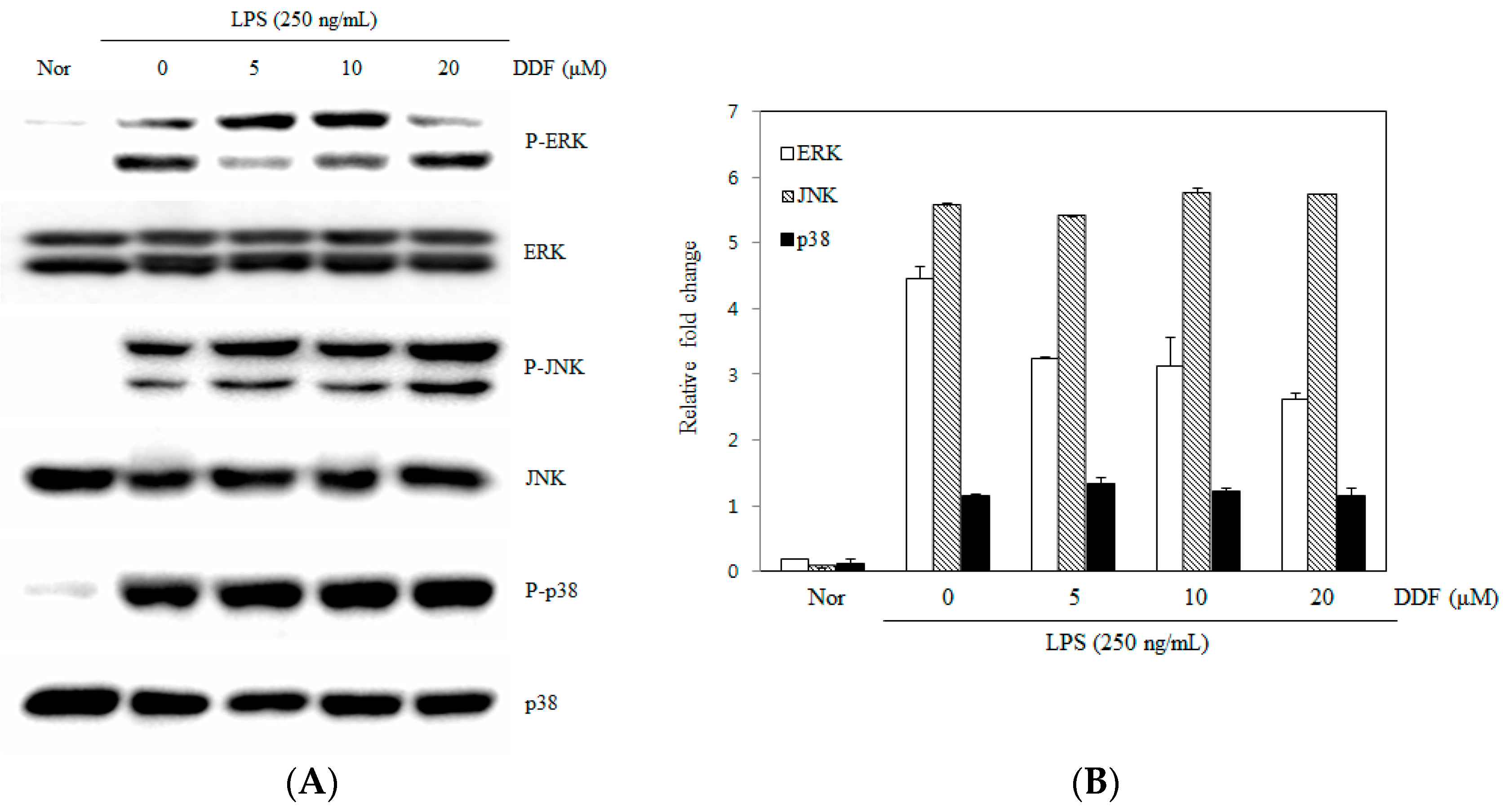
2.4. Activation of the MAPK Pathway by DDF
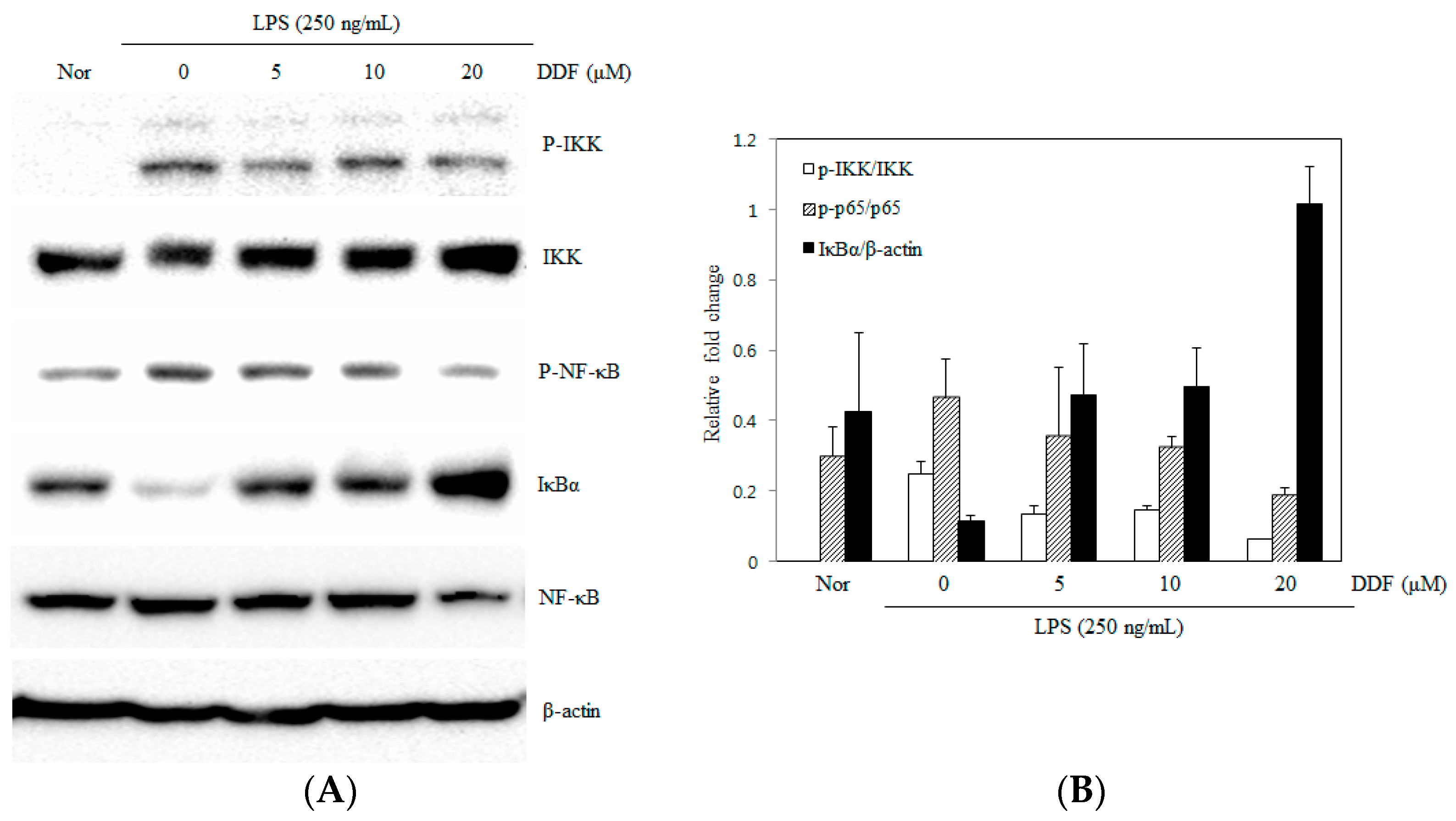
2.5. Modulation of the NF-κB Pathway by DDF
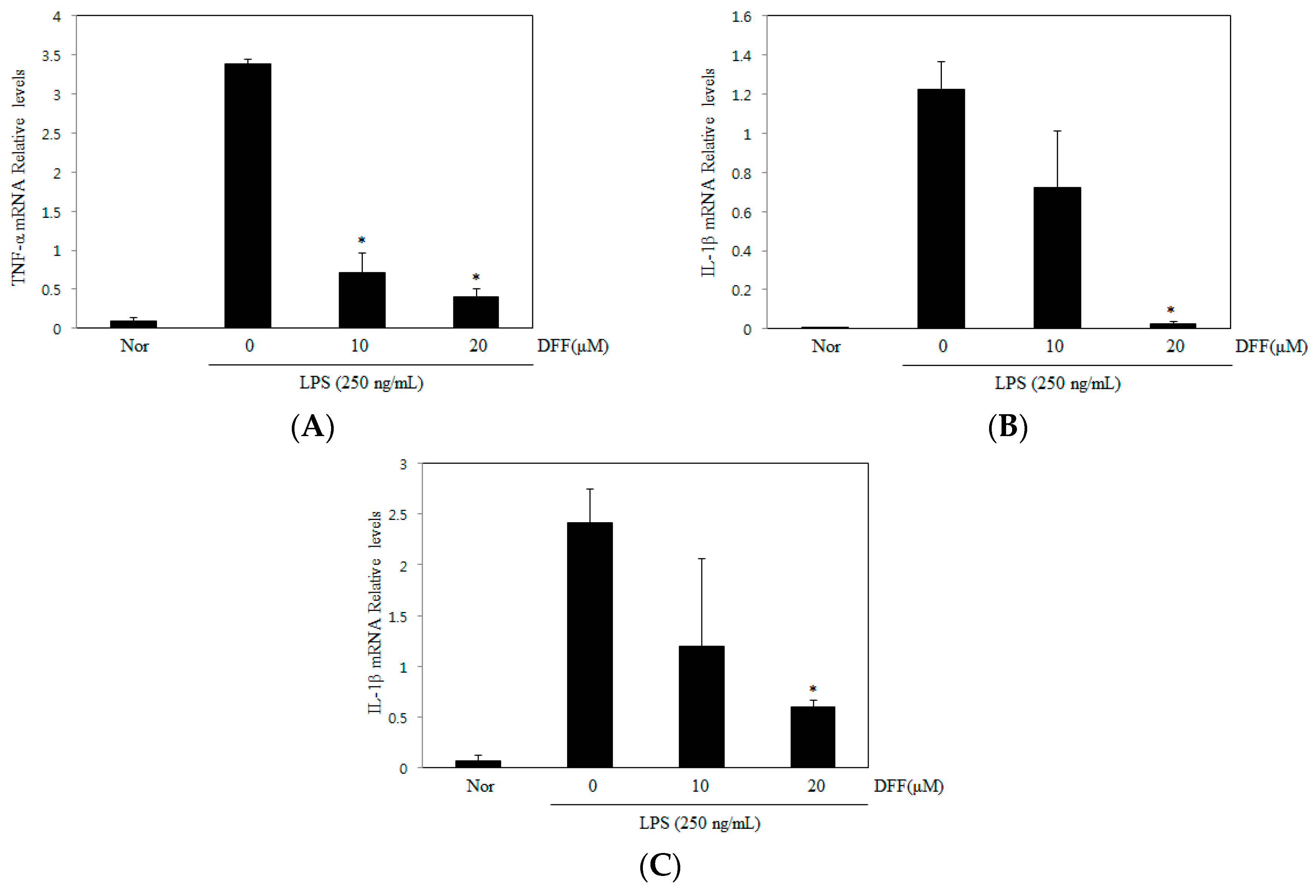
2.6. Effect of DDF on TNF-α, IL-1β, and IL-6
3. Discussion
4. Materials and Methods
4.1. Preparation of Compound 1
4.2. Cell Culture
4.3. Drugs and Chemicals
4.4. MTT Assay for Cell Viability
4.5. Measurement of NO Production
4.6. Immunoblot/Western Blot Analysis
4.7. Quantitative Real-Time RT-PCR
4.8. Immunofluorescence
4.9. Statistical Analysis
Supplementary Materials
Acknowledgments
Author Contributions
Conflicts of Interest
Abbreviations
| DDF | 6,8-diprenyl-7,4′-dihydroxyflavanone |
| So | sophoranone |
| Gl | glabrol |
| 5DTF | 8,5′-diprenyl-7,2′,4′-trihydroxyflavanone |
| Fl | flemichin D |
| To | tonkinochromane A |
| SC | sophoranochromene |
| iNOS | inducible nitric oxide synthases |
| MAPKs | mitogen-activated protein kinases |
| NF-κB | nuclear factor-kappa B |
References
- Ruocco, M.G.; Karin, M. IKKβ as a target for treatment of inflammation induced bone loss. Ann. Rheum. Dis. 2005, 64, 81–85. [Google Scholar] [CrossRef] [PubMed]
- Janssen, W.J.; Henson, P.M. Cellular regulation of the inflammatory response. Toxicol. Pathol. 2012, 40, 166–173. [Google Scholar] [CrossRef] [PubMed]
- Rennard, S.I. Inflammation and repair processes in chronic obstructive pulmonary disease. Am. J. Respir. Crit. Care Med. 1999, 160, S12–S16. [Google Scholar] [CrossRef] [PubMed]
- Henson, P.M.; Johnston, R.B., Jr. Tissue injury in inflammation. Oxidants, proteinases, and cationic proteins. J. Clin. Investig. 1987, 79, 669–674. [Google Scholar] [CrossRef] [PubMed]
- Martinez, F.O.; Sica, A.; Mantovani, A.; Locati, M. Macrophage activation and polarization. Front. Biosci. 2008, 13, 453–461. [Google Scholar] [CrossRef] [PubMed]
- Martinez, F.O.; Helming, L.; Gordon, S. Alternative activation of macrophages: An immunologic functional perspective. Annu. Rev. Immunol. 2009, 27, 451–483. [Google Scholar] [CrossRef] [PubMed]
- Gordon, S.; Taylor, P.R. Monocyte and macrophage heterogeneity. Nat. Rev. Immunol. 2005, 5, 953–964. [Google Scholar] [CrossRef] [PubMed]
- Surh, Y.J.; Chun, K.S.; Cha, H.H.; Han, S.S.; Keum, Y.S.; Park, K.K.; Lee, S.S. Molecular mechanisms underlying chemopreventive activities of anti-inflammatory phytochemicals: Down-regulation of COX-2 and iNOS through suppression of NF-kappa B activation. Mutat. Res. 2001, 480–481, 243–268. [Google Scholar] [CrossRef]
- Hwang, D.; Jang, B.C.; Yu, G.; Boudreau, M. Expression of mitogen-inducible cyclooxygenase induced by lipopolysaccharide: Mediation through both mitogen-activated protein kinase and NF-kappaB signaling pathways in macrophages. Biochem. Pharmacol. 1997, 54, 87–96. [Google Scholar] [CrossRef]
- Chen, F.; Castranova, V.; Shi, X.; Demers, L.M. New insights into the role of nuclear factor-kappaB, a ubiquitous transcription factor in the initiation of diseases. Clin. Chem. 1999, 45, 7–17. [Google Scholar] [PubMed]
- Lin, Y.; Bai, L.; Chen, W.; Xu, S. The NF-kappaB activation pathways, emerging molecular targets for cancer prevention and therapy. Expert Opin. Ther. Targets 2010, 14, 45–55. [Google Scholar] [CrossRef] [PubMed]
- Jung, M.; Choi, J.; Chae, H.S.; Cho, J.Y.; Kim, Y.D.; Htwe, K.M.; Lee, W.S.; Chin, Y.W.; Kim, J.; Yoon, K.D. Flavonoids from Symplocos racemose. Molecules 2014, 20, 358–365. [Google Scholar] [CrossRef] [PubMed]
- Kim, Y.M.; Chae, H.S.; Lee, E.J.; Yang, M.H.; Park, J.H.; Yoon, K.D.; Kim, J.; Ahn, H.C.; Choi, Y.H.; Chin, Y.W. A citrus flavonoid, 6-demethoxytangeretin, suppresses production and gene expression of interleukin-6 in human mast cell-1 via anaplastic lymphoma kinase and mitogen-activated protein kinase pathways. Biol. Pharm. Bull. 2014, 37, 871–876. [Google Scholar] [CrossRef] [PubMed]
- Lee, J.W.; Lee, J.H.; Lee, C.; Jin, Q.; Lee, D.; Kim, Y.; Hong, K.T.; Lee, M.K.; Hwang, B.Y. Inhibitory constituents of Sophora tonkinensis on nitric oxide production in RAW 264.7 macrophages. Bioorg. Med. Chem. Lett. 2015, 25, 960–962. [Google Scholar] [CrossRef] [PubMed]
- Inoue, M.; Tanabe, H.; Nakashima, K.; Ishida, Y.; Kotani, H. Rexinoids isolated from Sophora tonkinensis with a gene expression profile distinct from the synthetic rexinoid bexarotene. J. Nat. Prod. 2014, 77, 1670–1677. [Google Scholar] [CrossRef] [PubMed]
- Yoo, H.; Chae, H.S.; Kim, Y.M.; Kang, M.; Ryu, K.H.; Ahn, H.C.; Yoon, K.D.; Chin, Y.W.; Kim, J. Flavonoids and arylbenzofurans from the rhizomes and roots of Sophora tonkinensis with IL-6 production inhibitory activity. Bioorg. Med. Chem. Lett. 2014, 24, 5644–5647. [Google Scholar] [CrossRef] [PubMed]
- Azzolina, A.; Bongiovanni, A.; Lampiasi, N. Substance P induces TNF-alpha and IL-6 production through NF kappa B in peritoneal mast cells. Biochim. Biophys. Acta 2003, 1643, 75–83. [Google Scholar] [CrossRef] [PubMed]
- Lawrence, T. The nuclear factor NF-kappaB pathway in inflammation. Cold Spring Harb. Perspect. Biol. 2009, 1, a001651. [Google Scholar] [PubMed]
- Spitzer, J.A.; Zheng, M.; Kolls, J.K.; Vande Stouwe, C.; Spitzer, J.J. Ethanol and LPS modulate NF-kappaB activation, inducible NO synthase and COX-2 gene expression in rat liver cells in vivo. Front. Biosci. 2002, 7, a99–a108. [Google Scholar] [CrossRef] [PubMed]
- Mantovani, A.; Allavena, P.; Sica, A.; Balkwill, F. Cancer-related inflammation. Nature 2008, 454, 436–444. [Google Scholar] [CrossRef] [PubMed]
- Ho, F.M.; Lai, C.C.; Huang, L.J.; Kuo, T.C.; Chao, C.M.; Lin, W.W. The anti-inflammatory carbazole, LCY-2-CHO, inhibits lipopolysaccharide-induced inflammatory mediator expression through inhibition of the p38 mitogen-activated protein kinase signaling pathway in macrophages. Br. J. Pharmacol. 2004, 141, 1037–1047. [Google Scholar] [CrossRef] [PubMed]
- Nick, J.A.; Young, S.K.; Brown, K.K.; Avdi, N.J.; Arndt, P.G.; Suratt, B.T.; Janes, M.S.; Henson, P.M.; Worthen, G.S. Role of p38 mitogen-activated protein kinase in a murine model of pulmonary inflammation. J. Immunol. 2000, 164, 2151–2159. [Google Scholar] [CrossRef] [PubMed]
- Paul, A.; Wilson, S.; Belham, C.M.; Robinson, C.J.; Scott, P.H.; Gould, G.W.; Plevin, R. Stress-activated protein kinases: Activation, regulation and function. Cell Signal. 1997, 9, 403–410. [Google Scholar] [CrossRef]
- Baldwin, A.S., Jr. The NF-kappa B and I kappa B proteins: New discoveries and insights. Annu. Rev. Immunol. 1996, 14, 649–683. [Google Scholar] [CrossRef] [PubMed]
- Beg, A.A.; Baldwin, A.S., Jr. The I kappa B proteins: Multifunctional regulators of Rel/NF-kappa B transcription factors. Genes Dev. 1993, 7, 2064–2070. [Google Scholar] [CrossRef] [PubMed]
- Ohshima, H.; Bartsch, H. Chronic infections and inflammatory processes as cancer risk factors: Possible role of nitric oxide in carcinogenesis. Mutat. Res. 1994, 305, 253–264. [Google Scholar] [CrossRef]
- Alderton, W.K.; Cooper, C.E.; Knowles, R.G. Nitric oxide synthases: Structure, function and inhibition. Biochem. J. 2001, 357, 593–615. [Google Scholar] [CrossRef] [PubMed]
- Kyogoku, K.; Hatayama, K.; Suzuki, K.; Yokomori, S.; Maejima, K.; Komatsu, M. Studies on the Constituents of Guang-Dou-Gen (the Root of Sophora subprostrata CHUN et T.CHEN). (6). Isolation of Two New Flavanones. Chem. Pharm. Bull. 1973, 21, 1777–1782. [Google Scholar] [CrossRef]
- Green, L.C.; Wagner, D.A.; Glogowski, J.; Skipper, P.L.; Wishnok, J.S.; Tannenbaum, S.R. Analysis of nitrate, nitrite, and [15N]nitrate in biological fluids. Anal. Biochem. 1982, 126, 131–138. [Google Scholar] [CrossRef]
- Sample Availability: Samples of the compounds are available from the authors.







© 2016 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC-BY) license ( http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Chae, H.-S.; Yoo, H.; Kim, Y.-M.; Choi, Y.H.; Lee, C.H.; Chin, Y.-W. Anti-Inflammatory Effects of 6,8-Diprenyl-7,4′-dihydroxyflavanone from Sophora tonkinensis on Lipopolysaccharide-Stimulated RAW 264.7 Cells. Molecules 2016, 21, 1049. https://doi.org/10.3390/molecules21081049
Chae H-S, Yoo H, Kim Y-M, Choi YH, Lee CH, Chin Y-W. Anti-Inflammatory Effects of 6,8-Diprenyl-7,4′-dihydroxyflavanone from Sophora tonkinensis on Lipopolysaccharide-Stimulated RAW 264.7 Cells. Molecules. 2016; 21(8):1049. https://doi.org/10.3390/molecules21081049
Chicago/Turabian StyleChae, Hee-Sung, Hunseung Yoo, Young-Mi Kim, Young Hee Choi, Chang Hoon Lee, and Young-Won Chin. 2016. "Anti-Inflammatory Effects of 6,8-Diprenyl-7,4′-dihydroxyflavanone from Sophora tonkinensis on Lipopolysaccharide-Stimulated RAW 264.7 Cells" Molecules 21, no. 8: 1049. https://doi.org/10.3390/molecules21081049