
Metabolic Characteristics in Meal of Black Rapeseed and Yellow-Seeded Progeny of Brassica napus–Sinapis alba Hybrids

 ,
,
Abstract
:
1. Introduction
2. Results and Discussion
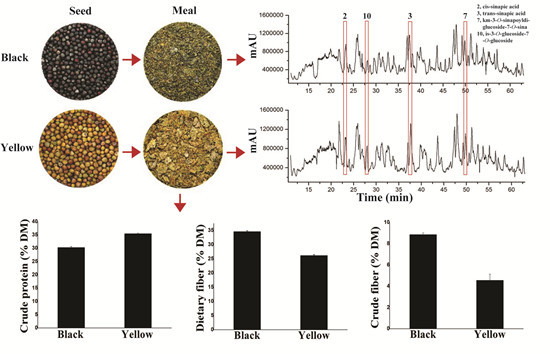
2.1. Differences in Chemical Composition between Yellow- and Black-Seeded Rapeseed Meals

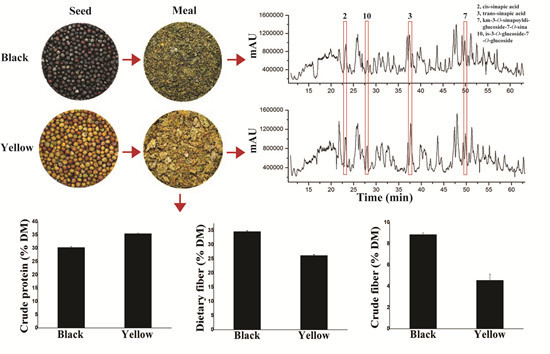{kind=link}
| Component | B. napus | B. napus |
|---|---|---|
| Black (% DM) | Yellow (% DM) | |
| Moisture | 4.02 ± 0.20 | 3.54 ± 0.08 |
| Ash | 4.55 ± 0.0 a | 3.65 ± 0.0 b |
| Crude protein | 30.29 ± 0.37 b | 35.46 ± 0.27 a |
| Sucrose | 7.29 ± 0.09 b | 7.85 ± 0.03 a |
| Starch | 1.60 ± 0.04 b | 2.44 ± 0.08 a |
| Dietary fiber | 34.63 ± 0.3 a | 26.19 ± 0.31 b |
| Crude fiber | 8.86 ± 0.15 a | 4.56 ± 0.59 b |
| Phytic acid | 5.60 ± 0.06 a | 4.98 ± 0.06 b |
| Total polyphenols | 2.82 ± 0.14 | 2.67 ± 0.04 |
| Total glucosinolates (μmol/g) | 28.19 ± 0.35 a | 22.18 ± 0.52 b |
| Amino Acid | Free Amino Acid | Total Amino Acid | ||
|---|---|---|---|---|
| B. napus | B. napus | B. napus | B. napus | |
| Yellow (mg/g) | Black (mg/g) | Yellow (mg/g) | Black (mg/g) | |
| Asparagine | 6.42 ± 0.11 | 6.67 ± 0.33 | 12.91 ± 0.28 a | 11.71 ± 0.08 b |
| Threonine | 0.44 ± 0.01 | 0.45 ± 0.01 | 9.37 ± 0.25 a | 8.29 ± 0.14 b |
| Serine | 5.86 ± 0.03 a | 2.65 ± 0.22 b | 8.27 ± 0.34 | 7.50 ± 0.22 |
| Glutamic | 15.5 ± 0.11 | 15.5 ± 0.55 | 23.75 ± 0.25 a | 21.18 ± 0.41 b |
| Methionine + Cysteine | 1.36 ± 0.18 a | 0.21 ± 0.01 b | 32.15 ± 0.47 a | 29.01 ± 0.33 b |
| Isoleucine | 0.33 ± 0.04 a | 0.20 ± 0.0 b | 13.39 ± 0.36 a | 11.97 ± 0.21 b |
| Leucine | 0.34 ± 0.04 | 0.28 ± 0.04 | 2.52 ± 0.04 | 2.23 ± 0.16 |
| Tyrosine | 1.22 ± 0.09 | 1.15 ± 0.04 | 15.44 ± 0.32 a | 13.80 ± 0.27 b |
| Glycine | 0.62 ± 0.02 a | 0.39 ± 0.02 b | 2.28 ± 0.10 a | 1.83 ± 0.04 b |
| Alanine | 2.95 ± 0.03 a | 1.06 ± 0.0 b | 6.99 ± 0.22 a | 6.04 ± 0.09 b |
| Valine | 0.70 ± 0.12 | 0.51 ± 0.03 | 4.25 ± 0.20 | 3.69 ± 0.02 |
| phenylalanine | 2.08 ± 0.01 a | 1.26 ± 0.05 b | 8.17 ± 0.23 a | 7.06 ± 0.00 b |
| Histidine | 0.98 ± 0.09 a | 0.50 ± 0.04 b | 6.73 ± 0.21 a | 5.88 ± 0.05 b |
| Lysine | 1.65 ± 0.01 a | 0.80 ± 0.01 b | 9.52 ± 0.19 | 9.02 ± 0.03 |
| Arginine | 1.59 ± 0.05 a | 1.17 ± 0.02 b | 12.13 ± 0.42 a | 10.44 ± 0.06 b |
| Component | B. napus | B. napus |
|---|---|---|
| Black (mg/kg) | Yellow (mg/kg) | |
| Cu | 4.67 ± 0.16 | 4.96 ± 0.24 |
| Fe | 87.48 ± 0.4 b | 91.02 ± 0.19 a |
| Mn | 37.63 ± 0.59 b | 48.12 ± 0.37 a |
| Zn | 54.82 ± 0.41 b | 74.89 ± 0.42 a |
| K | 12,157.70 ± 72.3 | 12,161.50 ± 176.9 |
| P | 11,966.29 ± 56.9 a | 10,999.92 ± 108.9 b |
| Ca | 5645.36 ± 38.42 a | 3990.40 ± 64.24 b |
| Mg | 3860.59 ± 26.6 a | 3767.20 ± 70.3 b |
2.2. Comparison of Polyphenolics between Yellow- and Black-Seeded Rapeseed Meals
3. Experimental Section
3.1. Plant Materials
| Peak | Compound | Content (μg/g) | Mass Spectrum (m/z) | MS2 [M − H]− (m/z) | MS2 [M + H]+ (m/z) | UV λmax (nm) | ||
|---|---|---|---|---|---|---|---|---|
| Black | Yellow | [M − H]− | [M + H]+ | |||||
| 1 | sinapine | 5469.73 ± 145.78 b | 8309.9 ± 125.54 a | - | 310 | - | 119, 175, 207, 251 | nd |
| 2 | cis-sinapic acid | 371.5 ± 35.2 b | 465.49 ± 43.32 a | 223 | - | 193, 164 | - | nd |
| 3 | trans-sinapic acid | 4439.24 ± 89.08 b | 8706.89 ± 154.39 a | 223 | - | 208, 179, 164 | - | nd |
| 4 | km-3-O-glucoside | 15.36 ± 0.34 a | 7.98 ± 0.04 b | 447 | - | 357, 285, 284 | - | 265, 345 |
| 5 | km-3-O-diglucoside | 14.53 ± 0.65 a | 6.2 ± 0.02 b | 609 | - | 447, 285 | - | 265, 335 |
| 6 | km-3-O-sinapoylsophoroside | 27.34 ± 0.78 a | 16.06 ± 0.97 b | 815 | - | 623, 609, 591 | - | nd |
| 7 | km-3-O-sinapoyldiglucoside-7-O-sinapoylglucoside | 57.71 ± 2.56 a | 63.96 ± 2.63 a | 1183 | - | 815, 609 | - | 270, 330 |
| 8 | is-3-O-glucoside | 34.18 ± 2.43 a | 17.44 ± 1.07 b | 477 | - | 315, 314 | - | 265, 355 |
| 9 | is-O-glucoside-sulfate | 4.29 ± 0.04 a | 2.67 ± 0.02 b | 557 | - | 477, 395, 315 | - | 265, 320 |
| 10 | is-3-O-glucoside-7-O-glucoside | 17.74 ± 0.56 a | 1.77 ± 0.00 b | 639 | - | 477, 315 | - | nd |
| 11 | is-3-O-glucoside-7-O-acetylglucoside | 16.58 ± 0.54 b | 21.89 ± 0.83 a | 681 | - | 476, 315 | - | nd |
| 12 | qn-3-O-glucoside | 596.84 ± 34.78 a | 492.51 ± 37.62 b | 463 | - | 300 | - | nd |
| 13 | qn-3-O-sophoroside | 21.11 ± 0.35 a | 22.8 ± 3.24 a | 625 | - | 463, 445, 300 | - | 265, 345 |
| 14 | qn-3-O-diglucoside-7-O-glucoside | 22.16 ± 2.02 a | 6.14 ± 0.38 b | 787 | - | 625 | - | nd |
| 15 | procyanidin B2 (DP [2]) | 15.56 ± 0.89 b | 68.1 ± 3.87 a | 577 | - | 425, 407, 289 | - | nd |
3.2. Chemical Analysis of Seed Meal
3.3. Analysis of Phenolics in Seed Meal
4. Conclusions
Acknowledgements
Author Contributions
Conflicts of Interest
References
- González-Vega, J.C.; Stein, H.H. Amino acid digestibility in canola, cotton seed, and sunflower products fed to finishing pigs. J. Anim. Sci. 2012, 90, 4391–4400. [Google Scholar] [CrossRef] [PubMed]
- Theodoridou, K.; Yu, P.Q. Metabolic characteristics of the proteins in yellow-seeded and brown-seeded canola meal and presscake in dairy cattle: Comparison of three systems (PDI, DVE, and NRC) in nutrient supply and feed milk value (FMV). J. Agric. Food Chem. 2013, 61, 2820–2830. [Google Scholar] [CrossRef] [PubMed]
- Theodoridou, K.; Yu, P.Q. Effect of processing conditions on the nutritive value of canola meal and presscake. Comparison of the yellow and brown-seeded canola meal with the brown-seeded canola presscake. J. Sci. Food Agric. 2013, 93, 1986–1995. [Google Scholar] [CrossRef] [PubMed]
- Khajali, F.; Slominski, B.A. Factors that affect the nutritive value of canola meal for poultry. Poult. Sci. 2012, 91, 2564–2575. [Google Scholar] [CrossRef] [PubMed]
- Wittkop, B.; Snowdon, R.J.; Friedt, W. Status and perspectives of breeding for enhanced yield and quality of oilseed crops for Europe. Euphytica 2009, 170, 131–140. [Google Scholar] [CrossRef]
- Wen, J.; Zhu, L.X.; Qi, L.P.; Ke, H.M.; Yi, B.; Shen, J.X.; Tu, J.X.; Ma, C.Z.; Fu, T.D. Charaterization of interploid hybrids from crosed between Brassica juncea and B. olercea and the production of yellow-seeded B. napus. Theor. Appl. Genet. 2012, 125, 19–32. [Google Scholar] [CrossRef] [PubMed]
- Jiang, J.J.; Shao, Y.L.; Li, A.M.; Lu, C.L.; Zhang, Y.T.; Wang, Y.P. Phenolic composition analysis and gene expression in developing seeds of yellow- and black-seeded Brassica napus. J. Integr. Plant Biol. 2013, 55, 537–551. [Google Scholar] [CrossRef] [PubMed]
- Theander, O.; Aman, P.; Miksche, G.E.; Yasunda, S. Carbohydrates, polyphenols and lignin in seed hulls of different colors from turnip rapeseed. J. Agric. Food Chem. 1977, 25, 270–273. [Google Scholar] [CrossRef]
- Daun, J.; KDeClercq, D.R. Quality of yellow and dark seeds in Brassica campestris canola varieties Candle and Tobin. J. Am. Oil Chem. Soc. 1988, 65, 122–126. [Google Scholar] [CrossRef]
- Slominski, B.A. Developments in the breeding of low fibre rapeseed/canola. J. Anim. Feed Sci. 1997, 6, 303–317. [Google Scholar]
- Bell, J.M. Factors affecting the nutritional value of canola meal: A review. Can. J. Anim. Sci. 1993, 73, 679–697. [Google Scholar] [CrossRef]
- Slominski, B.A.; Jia, W.; Rogiewicz, A.; Nyachoti, C.M.; Hickling, D. Low-Fiber Canola. Part 1. Chemical and nutritive composition of the meal. J. Agric. Food Chem. 2012, 60, 12225–12230. [Google Scholar] [CrossRef] [PubMed]
- Obied, H.K.; Song, Y.; Foley, S.; Loughlin, M.; Rehman, A.; Mailer, R.; Masud, T.; Agboola, S. Biophenols and antioxidant properties of Australian canola meal. J. Agric. Food Chem. 2013, 61, 9176–9184. [Google Scholar] [CrossRef] [PubMed]
- Landero, J.L.; Beltranena, E.; Zijlstra, R.T. Diet nutrient digestibility and growth performance of weaned pigs fed solvent-extracted Brassica juncea canola meal. Anim. Feed Sci. Technol. 2013, 180, 64–72. [Google Scholar] [CrossRef]
- Landero, J.L.; Beltranena, E.; Zijlstra, R.T. Growth performance and preference studies to evaluate solvent-extracted Brassica napus or Brassica juncea canola meal fed to weaned pigs. J. Anim. Sci. 2015, 90, 406–408. [Google Scholar] [CrossRef] [PubMed]
- Moreno, D.A.; Perez-Balibrea, S.; Ferreres, F.; Gil-lzquierdo, A.; Garcia-Viguera, C. Acylated anthocyanins in broccoli sprouts. Food Chem. 2010, 123, 358–363. [Google Scholar] [CrossRef]
- Simbaya, J.; Slominski, B.A.; Rakow, G. Quality characteristics of yellow-seeded Brassica seed meals: Protein, carbohydrate, and dietary fiber components. J. Agric. Food Chem. 1995, 43, 2062–2066. [Google Scholar] [CrossRef]
- Wallig, M.A.; Belyea, R.L.; Tumbleson, M.E. Effect of pelletingon glucosinolates content of Crambe meal. Anim. Feed Sci. Technol. 2002, 99, 205–214. [Google Scholar] [CrossRef]
- Lee, J.G.; Bonnema, G.; Zhang, N.; Kwak, J.H.; de vos, R.C.H.; Beekwilder, J. Evaluation of glucosinolate variation in a collection of turnip (Brassica rapa) germplasm by the analysis of intact and desulfo glucosinolates. J. Agric. Food Chem. 2013, 61, 3984–3993. [Google Scholar] [CrossRef] [PubMed]
- Vale, A.P.; Santos, J.; Brito, N.V.; Fernandes, D.; Rosa, E.; Oliverira, M. Evaluating the impact of sprouting conditions on the glucosinolate content of Brassica oleracea sprouts. Phytochemistry 2015, 115, 252–260. [Google Scholar] [CrossRef] [PubMed]
- Erdman, J.W.; Forbes, R.M. Mineral bioavailability from phytate-containing foods. Food Prod. Dev. 1977, 11, 46. [Google Scholar]
- Nwokolo, E.N.; Bragg, D.B. Influence of phytic acid and crude fibre on the availability of minerals from four protein supplements in growing chicks. Can. J. Anim. Sci. 1977, 57, 475–477. [Google Scholar] [CrossRef]
- Sigh, M.; Krikorian, A.D. Inhibition of trypsin activity in vitro by phyate. Agric. Food Chem. 1982, 30, 799–800. [Google Scholar]
- Cowieson, A.J.; Bedford, M.R.; Selle, P.H.; Ravindran, V. Phytate and microbial phytase: Implications for endogenous nitrogen losses and nutrient availability. World Poult. Sci. J. 2009, 65, 401–417. [Google Scholar] [CrossRef]
- Slominki, B.A. Recent advances in research on enzymes for poultry. Poult. Sci. 2011, 90, 2013–2023. [Google Scholar] [CrossRef] [PubMed]
- Fontaine, J.; Hörr, J.; Schirmer, B. Near-infrared reflectance spectroscopy enables the fast and accurate prediction of the essential amino acid contents in soy, rapeseed meal, sunflower meal, peas, fishmeal, meat meal products, and poultry meal. J. Agric. Food Chem. 2001, 49, 57–66. [Google Scholar] [CrossRef] [PubMed]
- Jia, W.; Mikulski, D.; Rogiewicz, A.; Zdunczyk, Z.; Jankowski, J.; Slominki, B.A. Low-fiber canola. Part 2. Nutritive value of the meal. J. Agric. Food Chem. 2012, 60, 12231–12237. [Google Scholar] [CrossRef] [PubMed]
- Tenore, G.C.; Troisi, J.; di Fiore, R. Chemical composition, antioxidant and antimicrobial properties of Rapa Catozza Napoletana (Brassica rapa L. var. rapa DC.) seed meal, a promising protein source of Campania region (southern Italy) horticultural germplasm. J. Sci. Food Agric. 2012, 92, 1716–1724. [Google Scholar] [CrossRef] [PubMed]
- Shao, Y.L.; Jiang, J.J.; Ran, L.P. Analysis of flavonoids and hydroxycinnamic acid derivatives in rapeseeds (Brassica napus L. var. napus) by HPLC-PDA–ESI (−)-MSn/HRMS. J. Agric. Food Chem. 2014, 62, 2935–2945. [Google Scholar] [PubMed]
- Jakobek, L. Interactions of polyphenols with carbohydrates, lipids and proteins. Food Chem. 2015, 175, 556–567. [Google Scholar] [CrossRef] [PubMed]
- Zhu, F. Interactions between starch and phenolic compound. Trends Food Sci. Technol. 2015, 43, 129–143. [Google Scholar] [CrossRef]
- Cartea, M.E.; Francisco, M.; Soengas, P.; Velasco, P. Phenolic compounds in Brassica vegetables. Molecules 2011, 16, 251–280. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Augar, B.; Marnet, N.; Gautier, V.; Maia-Grondard, A.; Leprince, F.; Renard, M.; Guyot, S.; Nesi, N.; Routaboul, J.M. A detailed survey of seed coat flavonoids in developing seeds of Brassica napus L. J. Agric. Food Chem. 2010, 58, 6246–6256. [Google Scholar] [CrossRef] [PubMed]
- Murota, K.; Terao, J. Antioxidative flavonoid quercetin: Implication of its intestinal absorption and metaboliam. Arch. Biochem. Biophys. 2003, 417, 12–17. [Google Scholar] [CrossRef]
- Li, A.M.; Jiang, J.J.; Zhang, Y.T.; Snowdon, R.J.; Liang, G.H.; Wang, Y.P. Molecular and cytological characterization of introgression lines in yellow seed derived from somatic hybrids between Brassica napus and Sinapis alba. Mol. Breed. 2012, 29, 209–219. [Google Scholar] [CrossRef]
- Jiang, J.J.; Shao, Y.L.; Li, A.M.; Zhang, Y.T.; Wei, C.X.; Wang, Y.P. FT-IR and NMR study of seed coat dissected from different colored progenies of Brassica napus–Sinapis alba hybrids. J. Sci. Food Agric. 2013, 93, 1898–1902. [Google Scholar] [CrossRef] [PubMed]
- Slominski, B.A.; Campbell, L.D.; Guenter, W. Carbohydrates and dietary fiber components of yellow- and brown-seeded canola. J. Sci. Food Agric. 1994, 42, 704–707. [Google Scholar] [CrossRef]
- Andrews, R.P.; Baldar, N.A. Amino acid analysis of feed constituents. Sci. Tools 1985, 32, 44–48. [Google Scholar]
- Association of official analytical chemicals. Official Methods of AOAC International, 18th ed.; AOAC International: Gaithersburg, MD, USA, 2005. [Google Scholar]
- Thompson, D.B.; Erdman, J.W. Phytic acid determination in soybeans. J. Food Sci. 1982, 47, 513–517. [Google Scholar] [CrossRef]
- Sample Availability: Samples of the seed meal, for research only, are available from the authors.
© 2015 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons by Attribution (CC-BY) license ( http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Jiang, J.; Wang, Y.; Xie, T.; Rong, H.; Li, A.; Fang, Y.; Wang, Y. Metabolic Characteristics in Meal of Black Rapeseed and Yellow-Seeded Progeny of Brassica napus–Sinapis alba Hybrids. Molecules 2015, 20, 21204-21213. https://doi.org/10.3390/molecules201219761
Jiang J, Wang Y, Xie T, Rong H, Li A, Fang Y, Wang Y. Metabolic Characteristics in Meal of Black Rapeseed and Yellow-Seeded Progeny of Brassica napus–Sinapis alba Hybrids. Molecules. 2015; 20(12):21204-21213. https://doi.org/10.3390/molecules201219761
Chicago/Turabian StyleJiang, Jinjin, Yue Wang, Tao Xie, Hao Rong, Aimin Li, Yujie Fang, and Youping Wang. 2015. "Metabolic Characteristics in Meal of Black Rapeseed and Yellow-Seeded Progeny of Brassica napus–Sinapis alba Hybrids" Molecules 20, no. 12: 21204-21213. https://doi.org/10.3390/molecules201219761