
An Uninvited Seat at the Dinner Table: How Apicomplexan Parasites Scavenge Nutrients from the Host


Abstract
:1. Introduction
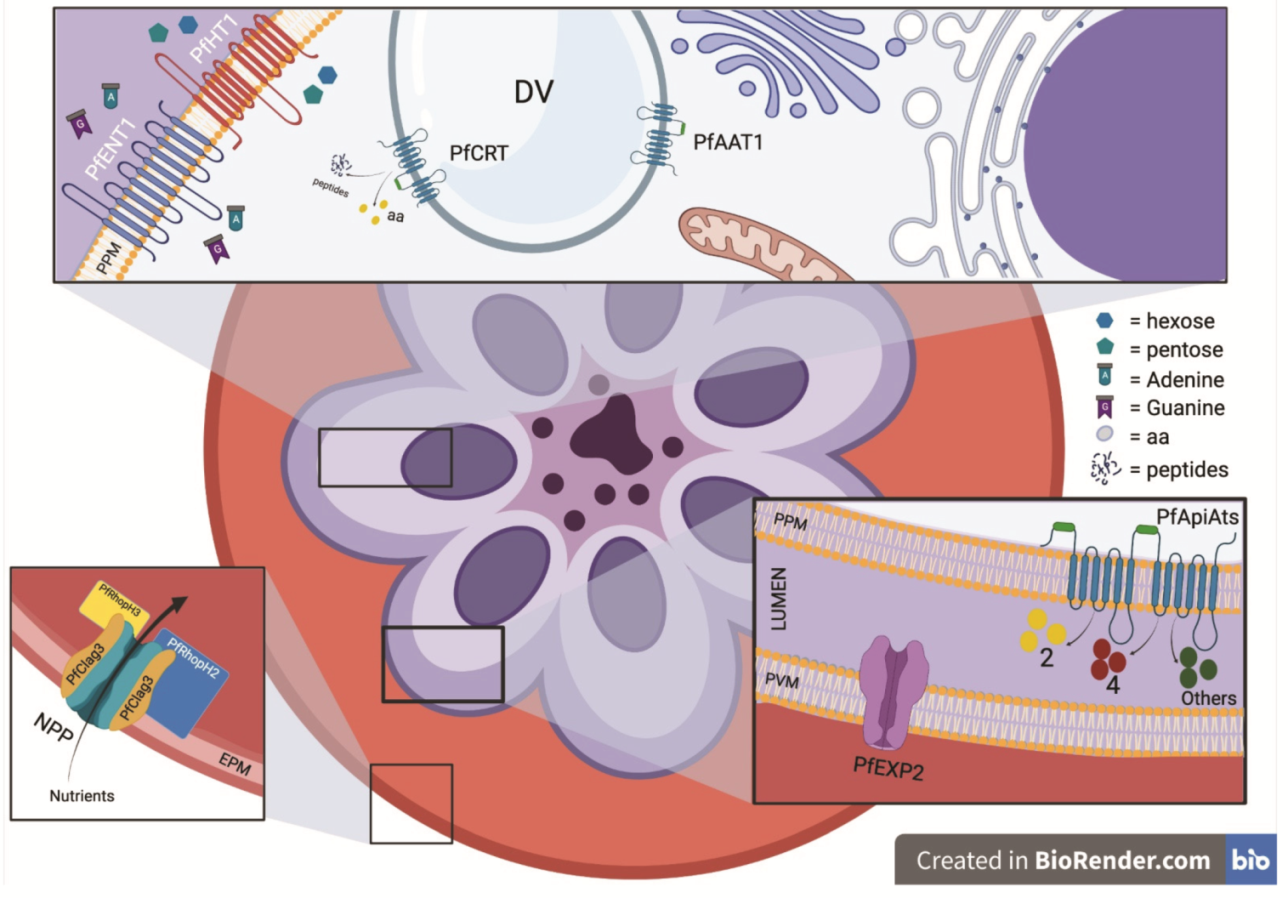
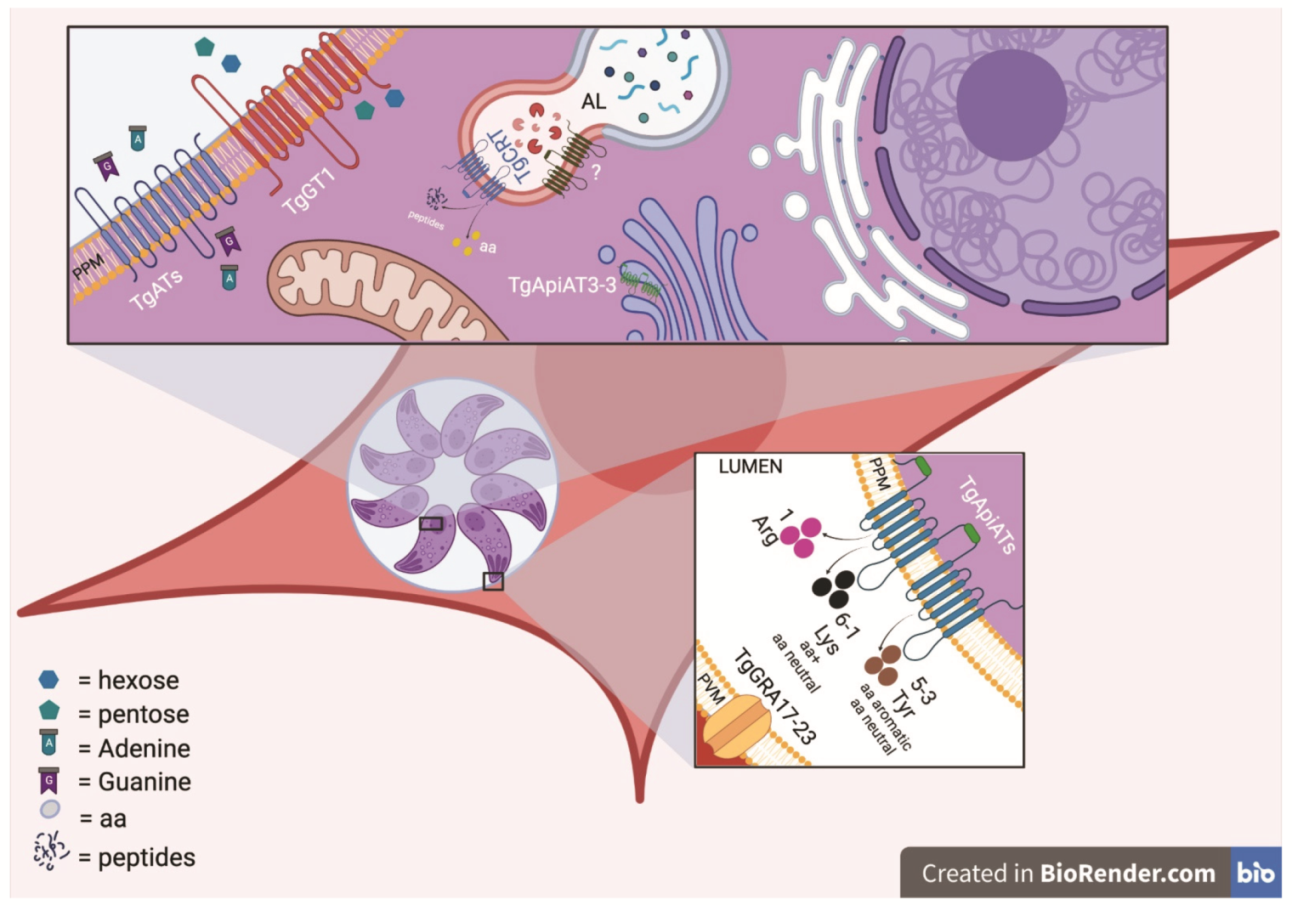
2. Apicomplexa Nutrient Uptake
2.1. Scavenging Nutrients across a Physical Barrier—The PVM
2.2. Transport across the Parasite Plasma Membrane
2.2.1. Sugar Transporters
2.2.2. Nucleotide Transporters
2.2.3. Amino Acid Transporters
3. Conclusions
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
Abbreviations
| ABC | ATP-Binding Cassette proteins |
| ABCG | ADP-binding cassette subfamily G |
| ApiATs | Apicomplexan Amino acid Transporters |
| αPFTs | α-pore-forming toxins |
| CRT | Chloroquine Resistance Transporter |
| DV | Digestive vacuole |
| ELC | endosome-like compartment |
| ENT | Equilibrative Nucleoside Transporter |
| ER | endoplasmic reticulum |
| GRA | Dense granule protein |
| HOST | host organelle-sequestering tubulo-structures |
| IFA | Indirect immunofluorescent assay |
| LDL | Low Density Lipoproteins |
| MFS | Major Facilitator Superfamily |
| MTOC | microtubule organizing centre |
| NPP | New Permeability Pathways |
| PSAC | Plasmodium Surface Anion Channel |
| PPM | Parasite plasma membrane |
| PTEX | Plasmodium translocon of exported proteins |
| PV | Parasitophorous vacuole |
| PVM | Parasitophorous vacuole membrane |
| SLC | Solute Carrier proteins |
| TM | Transmembrane |
| VAC | Vacuolar compartment |
References
- Adl, S.M.; Leander, B.S.; Simpson, A.G.B.; Archibald, J.M.; Anderson, O.R.; Bass, D.; Bowser, S.S.; Brugerolle, G.; Farmer, M.A.; Karpov, S.; et al. Diversity, Nomenclature, and Taxonomy of Protists. Syst. Biol. 2007, 56, 684–689. [Google Scholar] [CrossRef] [Green Version]
- White, M.W.; Suvorova, E.S. Apicomplexa Cell Cycles: Something Old, Borrowed, Lost, and New. Trends Parasitol. 2018, 34, 759–771. [Google Scholar] [CrossRef] [Green Version]
- Striepen, B.; Jordan, C.N.; Reiff, S.; van Dooren, G.G. Building the Perfect Parasite: Cell Division in Apicomplexa. PLoS Pathog. 2007, 3, e78. [Google Scholar] [CrossRef] [Green Version]
- Spielmann, T.; Gras, S.; Sabitzki, R.; Meissner, M. Endocytosis in Plasmodium and Toxoplasma Parasites. Trends Parasitol. 2020, 36, 520–532. [Google Scholar] [CrossRef] [PubMed]
- Guérin, A.; Striepen, B. The Biology of the Intestinal Intracellular Parasite Cryptosporidium. Cell Host Microbe 2020, 28, 509–515. [Google Scholar] [CrossRef] [PubMed]
- Attias, M.; Teixeira, D.E.; Benchimol, M.; Vommaro, R.C.; Crepaldi, P.H.; De Souza, W. The Life-Cycle of Toxoplasma gondii Reviewed Using Animations. Parasit Vectors 2020, 13, 588. [Google Scholar] [CrossRef]
- McCracken, A.N.; Edinger, A.L. Nutrient Transporters: The Achilles’ Heel of Anabolism. Trends Endocrinol. Metab. 2013, 24, 200–208. [Google Scholar] [CrossRef] [Green Version]
- Hediger, M.A. Structure, Function and Evolution of Solute Transporters in Prokaryotes and Eukaryotes. J. Exp. Biol. 1994, 196, 15–49. [Google Scholar] [CrossRef]
- Alexander, S.P.H.; Kelly, E.; Mathie, A.; Peters, J.A.; Veale, E.L.; Armstrong, J.F.; Faccenda, E.; Harding, S.D.; Pawson, A.J.; Sharman, J.L.; et al. THE CONCISE GUIDE TO PHARMACOLOGY 2019/20: Transporters. Br. J. Pharm. 2019, 176 (Suppl. S1), S397–S493. [Google Scholar] [CrossRef]
- Babcock, J.J.; Li, M. Deorphanizing the Human Transmembrane Genome: A Landscape of Uncharacterized Membrane Proteins. Acta Pharm. Sin. 2014, 35, 11–23. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Colas, C.; Ung, P.M.-U.; Schlessinger, A. SLC Transporters: Structure, Function, and Drug Discovery. Medchemcomm 2016, 7, 1069–1081. [Google Scholar] [CrossRef] [Green Version]
- Dean, M.; Hamon, Y.; Chimini, G. The Human ATP-Binding Cassette (ABC) Transporter Superfamily. J. Lipid Res. 2001, 42, 1007–1017. [Google Scholar] [CrossRef]
- Hediger, M.A.; Romero, M.F.; Peng, J.-B.; Rolfs, A.; Takanaga, H.; Bruford, E.A. The ABCs of Solute Carriers: Physiological, Pathological and Therapeutic Implications of Human Membrane Transport Proteins Introduction. Pflug. Arch. 2004, 447, 465–468. [Google Scholar] [CrossRef]
- Busch, W.; Saier, M.H. The Transporter Classification (TC) System, 2002. Crit. Rev. Biochem. Mol. Biol. 2002, 37, 287–337. [Google Scholar] [CrossRef]
- Dahl, S.G.; Sylte, I.; Ravna, A.W. Structures and Models of Transporter Proteins. J. Pharm. Exp. 2004, 309, 853–860. [Google Scholar] [CrossRef] [Green Version]
- Hediger, M.A.; Clémençon, B.; Burrier, R.E.; Bruford, E.A. The ABCs of Membrane Transporters in Health and Disease (SLC Series): Introduction. Mol. Asp. Med. 2013, 34, 95–107. [Google Scholar] [CrossRef]
- Rees, D.C.; Johnson, E.; Lewinson, O. ABC Transporters: The Power to Change. Nat. Rev. Mol. Cell Biol. 2009, 10, 218–227. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Thomas, C.; Tampé, R. Structural and Mechanistic Principles of ABC Transporters. Annu. Rev. Biochem. 2020, 89, 605–636. [Google Scholar] [CrossRef]
- Futai, M.; Sun-Wada, G.-H.; Wada, Y.; Matsumoto, N.; Nakanishi-Matsui, M. Vacuolar-Type ATPase: A Proton Pump to Lysosomal Trafficking. Proc. Jpn. Acad. Ser. B Phys. Biol. Sci. 2019, 95, 261–277. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gadsby, D.C. Structural Biology: Ion Pumps Made Crystal Clear. Nature 2007, 450, 957–959. [Google Scholar] [CrossRef] [PubMed]
- Lippe, G.; Coluccino, G.; Zancani, M.; Baratta, W.; Crusiz, P. Mitochondrial F-ATP Synthase and Its Transition into an Energy-Dissipating Molecular Machine. Oxid. Med. Cell. Longev. 2019, 2019, 8743257. [Google Scholar] [CrossRef]
- Palmgren, M.G.; Nissen, P. P-Type ATPases. Annu. Rev. Biophys. 2011, 40, 243–266. [Google Scholar] [CrossRef] [PubMed]
- Skou, J.C. The Fourth Datta Lecture. The Energy Coupled Exchange of Na+ for K+ across the Cell Membrane. The Na+, K+-Pump. FEBS Lett. 1990, 268, 314–324. [Google Scholar] [CrossRef] [Green Version]
- Bai, X.; Moraes, T.F.; Reithmeier, R.A.F. Structural Biology of Solute Carrier (SLC) Membrane Transport Proteins. Mol. Membr. Biol. 2017, 34, 1–32. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Shi, Y. Common Folds and Transport Mechanisms of Secondary Active Transporters. Annu. Rev. Biophys. 2013, 42, 51–72. [Google Scholar] [CrossRef]
- Zhang, Y.; Zhang, Y.; Sun, K.; Meng, Z.; Chen, L. The SLC Transporter in Nutrient and Metabolic Sensing, Regulation, and Drug Development. J. Mol. Cell Biol. 2019, 11, 1–13. [Google Scholar] [CrossRef] [Green Version]
- Perland, E.; Fredriksson, R. Classification Systems of Secondary Active Transporters. Trends Pharm. Sci. 2017, 38, 305–315. [Google Scholar] [CrossRef]
- Bröer, S. Adaptation of Plasma Membrane Amino Acid Transport Mechanisms to Physiological Demands. Pflug. Arch. 2002, 444, 457–466. [Google Scholar] [CrossRef]
- Povey, S.; Lovering, R.; Bruford, E.; Wright, M.; Lush, M.; Wain, H. The HUGO Gene Nomenclature Committee (HGNC). Hum. Genet. 2001, 109, 678–680. [Google Scholar] [CrossRef]
- Forrest, L.R. Structural Symmetry in Membrane Proteins. Annu. Rev. Biophys. 2015, 44, 311–337. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Drew, D.; North, R.A.; Nagarathinam, K.; Tanabe, M. Structures and General Transport Mechanisms by the Major Facilitator Superfamily (MFS). Chem. Rev. 2021, 121, 5289–5335. [Google Scholar] [CrossRef]
- Screpanti, E.; Hunte, C. Discontinuous Membrane Helices in Transport Proteins and Their Correlation with Function. J. Struct. Biol. 2007, 159, 261–267. [Google Scholar] [CrossRef]
- Yan, N. Structural Biology of the Major Facilitator Superfamily Transporters. Annu. Rev. Biophys. 2015, 44, 257–283. [Google Scholar] [CrossRef] [PubMed]
- UniProt Consortium UniProt: A Worldwide Hub of Protein Knowledge. Nucleic Acids Res. 2019, 47, D506–D515. [CrossRef] [Green Version]
- Kakuda, D.K.; MacLeod, C.L. Na+-Independent Transport (Uniport) of Amino Acids and Glucose in Mammalian Cells. J. Exp. Biol. 1994, 196, 93–108. [Google Scholar] [CrossRef] [PubMed]
- Clough, B.; Frickel, E.-M. The Toxoplasma Parasitophorous Vacuole: An Evolving Host-Parasite Frontier. Trends Parasitol. 2017, 33, 473–488. [Google Scholar] [CrossRef]
- Goldberg, D.E.; Zimmerberg, J. Hardly Vacuous: The Parasitophorous Vacuolar Membrane of Malaria Parasites. Trends Parasitol. 2020, 36, 138–146. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jalovecka, M.; Hajdusek, O.; Sojka, D.; Kopacek, P.; Malandrin, L. The Complexity of Piroplasms Life Cycles. Front. Cell Infect. Microbiol. 2018, 8, 248. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Desai, S.A. Ion and Nutrient Uptake by Malaria Parasite-Infected Erythrocytes. Cell Microbiol. 2012, 14, 1003–1009. [Google Scholar] [CrossRef] [Green Version]
- Desai, S.A.; Bezrukov, S.M.; Zimmerberg, J. A Voltage-Dependent Channel Involved in Nutrient Uptake by Red Blood Cells Infected with the Malaria Parasite. Nature 2000, 406, 1001–1005. [Google Scholar] [CrossRef]
- Kutner, S.; Ginsburg, H.; Cabantchik, Z.I. Permselectivity Changes in Malaria (Plasmodium falciparum) Infected Human Red Blood Cell Membranes. J. Cell Physiol. 1983, 114, 245–251. [Google Scholar] [CrossRef] [PubMed]
- Nguitragool, W.; Bokhari, A.A.B.; Pillai, A.D.; Rayavara, K.; Sharma, P.; Turpin, B.; Aravind, L.; Desai, S.A. Malaria Parasite Clag3 Genes Determine Channel-Mediated Nutrient Uptake by Infected Red Blood Cells. Cell 2011, 145, 665–677. [Google Scholar] [CrossRef] [Green Version]
- Counihan, N.A.; Modak, J.K.; de Koning-Ward, T.F. How Malaria Parasites Acquire Nutrients from Their Host. Front. Cell Dev. Biol. 2021, 9, 649184. [Google Scholar] [CrossRef]
- Delling, C.; Daugschies, A.; Bangoura, B.; Dengler, F. Cryptosporidium Parvum Alters Glucose Transport Mechanisms in Infected Enterocytes. Parasitol. Res. 2019, 118, 3429–3441. [Google Scholar] [CrossRef] [PubMed]
- Garten, M.; Nasamu, A.S.; Niles, J.C.; Zimmerberg, J.; Goldberg, D.E.; Beck, J.R. EXP2 Is a Nutrient-Permeable Channel in the Vacuolar Membrane of Plasmodium and Is Essential for Protein Export via PTEX. Nat. Microbiol. 2018, 3, 1090–1098. [Google Scholar] [CrossRef] [Green Version]
- Gold, D.A.; Kaplan, A.D.; Lis, A.; Bett, G.C.L.; Rosowski, E.E.; Cirelli, K.M.; Bougdour, A.; Sidik, S.M.; Beck, J.R.; Lourido, S.; et al. The Toxoplasma Dense Granule Proteins GRA17 and GRA23 Mediate the Movement of Small Molecules between the Host and the Parasitophorous Vacuole. Cell Host Microbe 2015, 17, 642–652. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mesén-Ramírez, P.; Bergmann, B.; Tran, T.T.; Garten, M.; Stäcker, J.; Naranjo-Prado, I.; Höhn, K.; Zimmerberg, J.; Spielmann, T. EXP1 Is Critical for Nutrient Uptake across the Parasitophorous Vacuole Membrane of Malaria Parasites. PLoS Biol. 2019, 17, e3000473. [Google Scholar] [CrossRef] [Green Version]
- Mesén-Ramírez, P.; Reinsch, F.; Blancke Soares, A.; Bergmann, B.; Ullrich, A.-K.; Tenzer, S.; Spielmann, T. Stable Translocation Intermediates Jam Global Protein Export in Plasmodium falciparum Parasites and Link the PTEX Component EXP2 with Translocation Activity. PLoS Pathog. 2016, 12, e1005618. [Google Scholar] [CrossRef]
- Collins, C.R.; Das, S.; Wong, E.H.; Andenmatten, N.; Stallmach, R.; Hackett, F.; Herman, J.-P.; Müller, S.; Meissner, M.; Blackman, M.J. Robust Inducible Cre Recombinase Activity in the Human Malaria Parasite Plasmodium falciparum Enables Efficient Gene Deletion within a Single Asexual Erythrocytic Growth Cycle. Mol. Microbiol. 2013, 88, 687–701. [Google Scholar] [CrossRef] [Green Version]
- Lisewski, A.M.; Quiros, J.P.; Ng, C.L.; Adikesavan, A.K.; Miura, K.; Putluri, N.; Eastman, R.T.; Scanfeld, D.; Regenbogen, S.J.; Altenhofen, L.; et al. Supergenomic Network Compression and the Discovery of EXP1 as a Glutathione Transferase Inhibited by Artesunate. Cell 2014, 158, 916–928. [Google Scholar] [CrossRef] [Green Version]
- Bano, N.; Romano, J.D.; Jayabalasingham, B.; Coppens, I. Cellular Interactions of Plasmodium Liver Stage with Its Host Mammalian Cell. Int. J. Parasitol. 2007, 37, 1329–1341. [Google Scholar] [CrossRef]
- Vaughan, A.M.; Mikolajczak, S.A.; Wilson, E.M.; Grompe, M.; Kaushansky, A.; Camargo, N.; Bial, J.; Ploss, A.; Kappe, S.H.I. Complete Plasmodium falciparum Liver-Stage Development in Liver-Chimeric Mice. J. Clin. Investig. 2012, 122, 3618–3628. [Google Scholar] [CrossRef] [PubMed]
- Wang, Y.; Sangaré, L.O.; Paredes-Santos, T.C.; Saeij, J.P.J. Toxoplasma Mechanisms for Delivery of Proteins and Uptake of Nutrients Across the Host-Pathogen Interface. Annu. Rev. Microbiol. 2020, 74, 567–586. [Google Scholar] [CrossRef] [PubMed]
- Paredes-Santos, T.; Wang, Y.; Waldman, B.; Lourido, S.; Saeij, J.P. The GRA17 Parasitophorous Vacuole Membrane Permeability Pore Contributes to Bradyzoite Viability. Front. Cell Infect. Microbiol. 2019, 9, 321. [Google Scholar] [CrossRef] [PubMed]
- Gonzalez, M.R.; Bischofberger, M.; Pernot, L.; van der Goot, F.G.; Frêche, B. Bacterial Pore-Forming Toxins: The (w)Hole Story? Cell Mol. Life Sci. 2008, 65, 493–507. [Google Scholar] [CrossRef] [Green Version]
- Wunderlich, J.; Rohrbach, P.; Dalton, J.P. The Malaria Digestive Vacuole. Front. Biosci. 2012, 4, 1424–1448. [Google Scholar] [CrossRef] [Green Version]
- Cobbold, S.A.; Martin, R.E.; Kirk, K. Methionine Transport in the Malaria Parasite Plasmodium falciparum. Int. J. Parasitol. 2011, 41, 125–135. [Google Scholar] [CrossRef] [PubMed]
- Martin, R.E.; Kirk, K. Transport of the Essential Nutrient Isoleucine in Human Erythrocytes Infected with the Malaria Parasite Plasmodium falciparum. Blood 2007, 109, 2217–2224. [Google Scholar] [CrossRef]
- McGovern, O.L.; Rivera-Cuevas, Y.; Carruthers, V.B. Emerging Mechanisms of Endocytosis in Toxoplasma gondii. Life 2021, 11, 84. [Google Scholar] [CrossRef]
- Coppens, I.; Dunn, J.D.; Romano, J.D.; Pypaert, M.; Zhang, H.; Boothroyd, J.C.; Joiner, K.A. Toxoplasma gondii Sequesters Lysosomes from Mammalian Hosts in the Vacuolar Space. Cell 2006, 125, 261–274. [Google Scholar] [CrossRef] [Green Version]
- Ehrenman, K.; Sehgal, A.; Lige, B.; Stedman, T.T.; Joiner, K.A.; Coppens, I. Novel Roles for ATP-Binding Cassette G Transporters in Lipid Redistribution in Toxoplasma. Mol. Microbiol. 2010, 76, 1232–1249. [Google Scholar] [CrossRef] [Green Version]
- Blume, M.; Rodriguez-Contreras, D.; Landfear, S.; Fleige, T.; Soldati-Favre, D.; Lucius, R.; Gupta, N. Host-Derived Glucose and Its Transporter in the Obligate Intracellular Pathogen Toxoplasma gondii Are Dispensable by Glutaminolysis. Proc. Natl. Acad. Sci. USA 2009, 106, 12998–13003. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Woodrow, C.J.; Penny, J.I.; Krishna, S. Intraerythrocytic Plasmodium falciparum Expresses a High Affinity Facilitative Hexose Transporter. J. Biol. Chem. 1999, 274, 7272–7277. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Carter, N.S.; Ben Mamoun, C.; Liu, W.; Silva, E.O.; Landfear, S.M.; Goldberg, D.E.; Ullman, B. Isolation and Functional Characterization of the PfNT1 Nucleoside Transporter Gene from Plasmodium falciparum. J. Biol. Chem. 2000, 275, 10683–10691. [Google Scholar] [CrossRef]
- Parker, M.D.; Hyde, R.J.; Yao, S.Y.; McRobert, L.; Cass, C.E.; Young, J.D.; McConkey, G.A.; Baldwin, S.A. Identification of a Nucleoside/Nucleobase Transporter from Plasmodium falciparum, a Novel Target for Anti-Malarial Chemotherapy. Biochem. J. 2000, 349, 67–75. [Google Scholar] [CrossRef]
- Downie, M.J.; El Bissati, K.; Bobenchik, A.M.; Nic Lochlainn, L.; Amerik, A.; Zufferey, R.; Kirk, K.; Ben Mamoun, C. PfNT2, a Permease of the Equilibrative Nucleoside Transporter Family in the Endoplasmic Reticulum of Plasmodium falciparum. J. Biol. Chem. 2010, 285, 20827–20833. [Google Scholar] [CrossRef] [Green Version]
- Downie, M.J.; Saliba, K.J.; Bröer, S.; Howitt, S.M.; Kirk, K. Purine Nucleobase Transport in the Intraerythrocytic Malaria Parasite. Int. J. Parasitol. 2008, 38, 203–209. [Google Scholar] [CrossRef] [PubMed]
- Downie, M.J.; Saliba, K.J.; Howitt, S.M.; Bröer, S.; Kirk, K. Transport of Nucleosides across the Plasmodium falciparum Parasite Plasma Membrane Has Characteristics of PfENT1. Mol. Microbiol. 2006, 60, 738–748. [Google Scholar] [CrossRef]
- Chiang, C.W.; Carter, N.; Sullivan, W.J., Jr.; Donald, R.G.; Roos, D.S.; Naguib, F.N.; el Kouni, M.H.; Ullman, B.; Wilson, C.M. The Adenosine Transporter of Toxoplasma gondii Identification by Insertional Mutagenesis, Cloning, and Recombinant Expression. J. Biol. Chem. 1999, 274, 35255–35261. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wichers, J.S.; van Gelder, C.; Fuchs, G.; Ruge, J.M.; Pietsch, E.; Ferreira, J.L.; Safavi, S.; von Thien, H.; Burda, P.-C.; Mesén-Ramirez, P.; et al. Characterization of Apicomplexan Amino Acid Transporters (ApiATs) in the Malaria Parasite Plasmodium falciparum. mSphere 2021, 6, e0074321. [Google Scholar] [CrossRef]
- Rajendran, E.; Hapuarachchi, S.V.; Miller, C.M.; Fairweather, S.J.; Cai, Y.; Smith, N.C.; Cockburn, I.A.; Bröer, S.; Kirk, K.; van Dooren, G.G. Cationic Amino Acid Transporters Play Key Roles in the Survival and Transmission of Apicomplexan Parasites. Nat. Commun. 2017, 8, 14455. [Google Scholar] [CrossRef] [Green Version]
- Boisson, B.; Lacroix, C.; Bischoff, E.; Gueirard, P.; Bargieri, D.Y.; Franke-Fayard, B.; Janse, C.J.; Ménard, R.; Baldacci, P. The Novel Putative Transporter NPT1 Plays a Critical Role in Early Stages of Plasmodium berghei Sexual Development. Mol. Microbiol. 2011, 81, 1343–1357. [Google Scholar] [CrossRef]
- Parker, K.E.R.; Fairweather, S.J.; Rajendran, E.; Blume, M.; McConville, M.J.; Bröer, S.; Kirk, K.; van Dooren, G.G. The Tyrosine Transporter of Toxoplasma gondii Is a Member of the Newly Defined Apicomplexan Amino Acid Transporter (ApiAT) Family. PLoS Pathog. 2019, 15, e1007577. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rajendran, E.; Clark, M.; Goulart, C.; Steinhöfel, B.; Tjhin, E.T.; Gross, S.; Smith, N.C.; Kirk, K.; van Dooren, G.G. Substrate-Mediated Regulation of the Arginine Transporter of Toxoplasma gondii. PLoS Pathog. 2021, 17, e1009816. [Google Scholar] [CrossRef] [PubMed]
- Fairweather, S.J.; Rajendran, E.; Blume, M.; Javed, K.; Steinhöfel, B.; McConville, M.J.; Kirk, K.; Bröer, S.; van Dooren, G.G. Coordinated Action of Multiple Transporters in the Acquisition of Essential Cationic Amino Acids by the Intracellular Parasite Toxoplasma gondii. PLoS Pathog. 2021, 17, e1009835. [Google Scholar] [CrossRef]
- Kloehn, J.; Lunghi, M.; Varesio, E.; Dubois, D.; Soldati-Favre, D. Untargeted Metabolomics Uncovers the Essential Lysine Transporter in Toxoplasma gondii. Metabolites 2021, 11, 476. [Google Scholar] [CrossRef] [PubMed]
- Martin, R.E. The Transportome of the Malaria Parasite. Biol. Camb. Philos. 2020, 95, 305–332. [Google Scholar] [CrossRef] [PubMed]
- Fidock, D.A.; Nomura, T.; Talley, A.K.; Cooper, R.A.; Dzekunov, S.M.; Ferdig, M.T.; Ursos, L.M.; Sidhu, A.B.; Naudé, B.; Deitsch, K.W.; et al. Mutations in the P. Falciparum Digestive Vacuole Transmembrane Protein PfCRT and Evidence for Their Role in Chloroquine Resistance. Cell 2000, 6, 861–871. [Google Scholar] [CrossRef]
- Martin, R.E.; Marchetti, R.V.; Cowan, A.I.; Howitt, S.M.; Bröer, S.; Kirk, K. Chloroquine Transport via the Malaria Parasite’s Chloroquine Resistance Transporter. Science 2009, 325, 1680–1682. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Thornton, L.B.; Teehan, P.; Floyd, K.; Cochrane, C.; Bergmann, A.; Riegel, B.; Stasic, A.J.; Di Cristina, M.; Moreno, S.N.J.; Roepe, P.D.; et al. An Ortholog of Plasmodium falciparum Chloroquine Resistance Transporter (PfCRT) Plays a Key Role in Maintaining the Integrity of the Endolysosomal System in Toxoplasma gondii to Facilitate Host Invasion. PLoS Pathog. 2019, 15, e1007775. [Google Scholar] [CrossRef] [Green Version]
- Kannan, G.; Di Cristina, M.; Schultz, A.J.; Huynh, M.-H.; Wang, F.; Schultz, T.L.; Lunghi, M.; Coppens, I.; Carruthers, V.B. Role of Toxoplasma gondii Chloroquine Resistance Transporter in Bradyzoite Viability and Digestive Vacuole Maintenance. mBio 2019, 10, e01324-19. [Google Scholar] [CrossRef] [Green Version]
- Warring, S.D.; Dou, Z.; Carruthers, V.B.; McFadden, G.I.; van Dooren, G.G. Characterization of the Chloroquine Resistance Transporter Homologue in Toxoplasma gondii. Eukaryot. Cell 2014, 13, 1360–1370. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Al-Anouti, F.; Tomavo, S.; Parmley, S.; Ananvoranich, S. The Expression of Lactate Dehydrogenase Is Important for the Cell Cycle of Toxoplasma gondii. J. Biol. Chem. 2004, 279, 52300–52311. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- MacRae, J.I.; Sheiner, L.; Nahid, A.; Tonkin, C.; Striepen, B.; McConville, M.J. Mitochondrial Metabolism of Glucose and Glutamine Is Required for Intracellular Growth of Toxoplasma gondii. Cell Host Microbe 2012, 12, 682–692. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Saliba, K.J.; Kirk, K. Nutrient Acquisition by Intracellular Apicomplexan Parasites: Staying in for Dinner. Int. J. Parasitol. 2001, 31, 1321–1330. [Google Scholar] [CrossRef]
- Shen, B.; Sibley, L.D. Toxoplasma Aldolase Is Required for Metabolism but Dispensable for Host-Cell Invasion. Proc. Natl. Acad. Sci. USA 2014, 111, 3567–3572. [Google Scholar] [CrossRef] [Green Version]
- Pomel, S.; Luk, F.C.Y.; Beckers, C.J.M. Host Cell Egress and Invasion Induce Marked Relocations of Glycolytic Enzymes in Toxoplasma gondii Tachyzoites. PLoS Pathog. 2008, 4, e1000188. [Google Scholar] [CrossRef] [Green Version]
- Shukla, A.; Olszewski, K.L.; Llinás, M.; Rommereim, L.M.; Fox, B.A.; Bzik, D.J.; Xia, D.; Wastling, J.; Beiting, D.; Roos, D.S.; et al. Glycolysis Is Important for Optimal Asexual Growth and Formation of Mature Tissue Cysts by Toxoplasma gondii. Int. J. Parasitol. 2018, 48, 955–968. [Google Scholar] [CrossRef]
- Kirk, K.; Horner, H.A.; Kirk, J. Glucose Uptake in Plasmodium falciparum-Infected Erythrocytes Is an Equilibrative Not an Active Process. Mol. Biochem. Parasitol. 1996, 82, 195–205. [Google Scholar] [CrossRef]
- Slavic, K.; Straschil, U.; Reininger, L.; Doerig, C.; Morin, C.; Tewari, R.; Krishna, S. Life Cycle Studies of the Hexose Transporter of Plasmodium Species and Genetic Validation of Their Essentiality. Mol. Microbiol. 2010, 75, 1402–1413. [Google Scholar] [CrossRef] [Green Version]
- Roth, E. Plasmodium falciparum Carbohydrate Metabolism: A Connection between Host Cell and Parasite. Blood Cells 1990, 16, 453–460, discussion 461–466. [Google Scholar] [PubMed]
- Qureshi, A.A.; Suades, A.; Matsuoka, R.; Brock, J.; McComas, S.E.; Nji, E.; Orellana, L.; Claesson, M.; Delemotte, L.; Drew, D. The Molecular Basis for Sugar Import in Malaria Parasites. Nature 2020, 578, 321–325. [Google Scholar] [CrossRef] [PubMed]
- Woodrow, C.J.; Burchmore, R.J.; Krishna, S. Hexose Permeation Pathways in Plasmodium falciparum-Infected Erythrocytes. Proc. Natl. Acad. Sci. USA 2000, 97, 9931–9936. [Google Scholar] [CrossRef] [Green Version]
- Slavic, K.; Delves, M.J.; Prudêncio, M.; Talman, A.M.; Straschil, U.; Derbyshire, E.T.; Xu, Z.; Sinden, R.E.; Mota, M.M.; Morin, C.; et al. Use of a Selective Inhibitor to Define the Chemotherapeutic Potential of the Plasmodial Hexose Transporter in Different Stages of the Parasite’s Life Cycle. Antimicrob. Agents Chemother. 2011, 55, 2824–2830. [Google Scholar] [CrossRef] [Green Version]
- Mahmud, O.; Kissinger, J.C. Evolution of the Apicomplexan Sugar Transporter Gene Family Repertoire. Int. J. Genom. 2017, 2017, 1707231. [Google Scholar] [CrossRef] [Green Version]
- Sherman, I.W. Malaria: Parasite Biology, Pathogenesis, and Protection; Amer Society for Microbiology: Washington, DC, USA, 1998; ISBN 978-1-55581-131-0. [Google Scholar]
- Berens, R.L.; Krug, E.C.; Marr, J.J. 6—Purine and Pyrimidine Metabolism. In Biochemistry and Molecular Biology of Parasites; Marr, J.J., Müller, M., Eds.; Academic Press: San Diego, CA, USA, 1995; pp. 89–117. ISBN 978-0-12-473345-9. [Google Scholar]
- Martin, R.E.; Henry, R.I.; Abbey, J.L.; Clements, J.D.; Kirk, K. The “permeome” of the Malaria Parasite: An Overview of the Membrane Transport Proteins of Plasmodium falciparum. Genome Biol. 2005, 6, R26. [Google Scholar] [CrossRef] [Green Version]
- Riegelhaupt, P.M.; Frame, I.J.; Akabas, M.H. Transmembrane Segment 11 Appears to Line the Purine Permeation Pathway of the Plasmodium falciparum Equilibrative Nucleoside Transporter 1 (PfENT1). J. Biol. Chem. 2010, 285, 17001–17010. [Google Scholar] [CrossRef] [Green Version]
- Riegelhaupt, P.M.; Cassera, M.B.; Fröhlich, R.F.G.; Hazleton, K.Z.; Hefter, J.J.; Schramm, V.L.; Akabas, M.H. Transport of Purines and Purine Salvage Pathway Inhibitors by the Plasmodium falciparum Equilibrative Nucleoside Transporter PfENT1. Mol. Biochem. Parasitol. 2010, 169, 40–49. [Google Scholar] [CrossRef] [Green Version]
- El Bissati, K.; Zufferey, R.; Witola, W.H.; Carter, N.S.; Ullman, B.; Ben Mamoun, C. The Plasma Membrane Permease PfNT1 Is Essential for Purine Salvage in the Human Malaria Parasite Plasmodium falciparum. Proc. Natl. Acad. Sci. USA 2006, 103, 9286–9291. [Google Scholar] [CrossRef] [Green Version]
- El Bissati, K.; Downie, M.J.; Kim, S.-K.; Horowitz, M.; Carter, N.; Ullman, B.; Ben Mamoun, C. Genetic Evidence for the Essential Role of PfNT1 in the Transport and Utilization of Xanthine, Guanine, Guanosine and Adenine by Plasmodium falciparum. Mol. Biochem. Parasitol. 2008, 161, 130–139. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Quashie, N.B.; Dorin-Semblat, D.; Bray, P.G.; Biagini, G.A.; Doerig, C.; Ranford-Cartwright, L.C.; De Koning, H.P. A Comprehensive Model of Purine Uptake by the Malaria Parasite Plasmodium falciparum: Identification of Four Purine Transport Activities in Intraerythrocytic Parasites. Biochem. J. 2008, 411, 287–295. [Google Scholar] [CrossRef]
- Frame, I.J.; Merino, E.F.; Schramm, V.L.; Cassera, M.B.; Akabas, M.H. Malaria Parasite Type 4 Equilibrative Nucleoside Transporters (ENT4) Are Purine Transporters with Distinct Substrate Specificity. Biochem. J. 2012, 446, 179–190. [Google Scholar] [CrossRef] [Green Version]
- Cassera, M.B.; Hazleton, K.Z.; Merino, E.F.; Obaldia, N.; Ho, M.-C.; Murkin, A.S.; DePinto, R.; Gutierrez, J.A.; Almo, S.C.; Evans, G.B.; et al. Plasmodium falciparum Parasites Are Killed by a Transition State Analogue of Purine Nucleoside Phosphorylase in a Primate Animal Model. PLoS ONE 2011, 6, e26916. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Frame, I.J.; Deniskin, R.; Arora, A.; Akabas, M.H. Purine Import into Malaria Parasites as a Target for Antimalarial Drug Development. Ann. N. Y. Acad. Sci. 2015, 1342, 19–28. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hyde, J.E. Targeting Purine and Pyrimidine Metabolism in Human Apicomplexan Parasites. Curr. Drug Targets 2007, 8, 31–47. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- De Koning, H.P.; Al-Salabi, M.I.; Cohen, A.M.; Coombs, G.H.; Wastling, J.M. Identification and Characterisation of High Affinity Nucleoside and Nucleobase Transporters in Toxoplasma gondii. Int. J. Parasitol. 2003, 33, 821–831. [Google Scholar] [CrossRef]
- Sangaré, L.O.; Alayi, T.D.; Westermann, B.; Hovasse, A.; Sindikubwabo, F.; Callebaut, I.; Werkmeister, E.; Lafont, F.; Slomianny, C.; Hakimi, M.-A.; et al. Unconventional Endosome-like Compartment and Retromer Complex in Toxoplasma gondii Govern Parasite Integrity and Host Infection. Nat. Commun. 2016, 7, 11191. [Google Scholar] [CrossRef] [Green Version]
- Sidik, S.M.; Huet, D.; Ganesan, S.M.; Huynh, M.-H.; Wang, T.; Nasamu, A.S.; Thiru, P.; Saeij, J.P.J.; Carruthers, V.B.; Niles, J.C.; et al. A Genome-Wide CRISPR Screen in Toxoplasma Identifies Essential Apicomplexan Genes. Cell 2016, 166, 1423–1435.e12. [Google Scholar] [CrossRef] [Green Version]
- Augusto, L.; Amin, P.H.; Wek, R.C.; Sullivan, W.J. Regulation of Arginine Transport by GCN2 EIF2 Kinase Is Important for Replication of the Intracellular Parasite Toxoplasma gondii. PLoS Pathog. 2019, 15, e1007746. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wallbank, B.A.; Dominicus, C.S.; Broncel, M.; Legrave, N.; Kelly, G.; MacRae, J.I.; Staines, H.M.; Treeck, M. Characterisation of the Toxoplasma gondii Tyrosine Transporter and Its Phosphorylation by the Calcium-Dependent Protein Kinase 3. Mol. Microbiol. 2019, 111, 1167–1181. [Google Scholar] [CrossRef] [Green Version]
- Divo, A.A.; Geary, T.G.; Davis, N.L.; Jensen, J.B. Nutritional Requirements of Plasmodium falciparum in Culture. I. Exogenously Supplied Dialyzable Components Necessary for Continuous Growth. J. Protozool. 1985, 32, 59–64. [Google Scholar] [CrossRef]
- Liu, J.; Istvan, E.S.; Gluzman, I.Y.; Gross, J.; Goldberg, D.E. Plasmodium falciparum Ensures Its Amino Acid Supply with Multiple Acquisition Pathways and Redundant Proteolytic Enzyme Systems. Proc. Natl. Acad. Sci. USA 2006, 103, 8840–8845. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hill, R.J.; Konigsberg, W.; Guidotti, G.; Craig, L.C. The Structure of Human Hemoglobin. I. The Separation of the Alpha and Beta Chains and Their Amino Acid Composition. J. Biol. Chem. 1962, 237, 1549–1554. [Google Scholar] [CrossRef]
- Kenthirapalan, S.; Waters, A.P.; Matuschewski, K.; Kooij, T.W.A. Functional Profiles of Orphan Membrane Transporters in the Life Cycle of the Malaria Parasite. Nat. Commun. 2016, 7, 10519. [Google Scholar] [CrossRef]
- Martin, R.E.; Ginsburg, H.; Kirk, K. Membrane Transport Proteins of the Malaria Parasite. Mol. Microbiol. 2009, 74, 519–528. [Google Scholar] [CrossRef]
- Zhang, M.; Wang, C.; Otto, T.D.; Oberstaller, J.; Liao, X.; Adapa, S.R.; Udenze, K.; Bronner, I.F.; Casandra, D.; Mayho, M.; et al. Uncovering the Essential Genes of the Human Malaria Parasite Plasmodium falciparum by Saturation Mutagenesis. Science 2018, 360, eaap7847. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ngotho, P.; Soares, A.B.; Hentzschel, F.; Achcar, F.; Bertuccini, L.; Marti, M. Revisiting Gametocyte Biology in Malaria Parasites. FEMS Microbiol. 2019, 43, 401–414. [Google Scholar] [CrossRef]
- Bártfai, R.; Hoeijmakers, W.A.M.; Salcedo-Amaya, A.M.; Smits, A.H.; Janssen-Megens, E.; Kaan, A.; Treeck, M.; Gilberger, T.-W.; Françoijs, K.-J.; Stunnenberg, H.G. H2A.Z Demarcates Intergenic Regions of the Plasmodium falciparum Epigenome That Are Dynamically Marked by H3K9ac and H3K4me3. PLoS Pathog. 2010, 6, e1001223. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cowell, A.N.; Istvan, E.S.; Lukens, A.K.; Gomez-Lorenzo, M.G.; Vanaerschot, M.; Sakata-Kato, T.; Flannery, E.L.; Magistrado, P.; Owen, E.; Abraham, M.; et al. Mapping the Malaria Parasite Druggable Genome by Using In Vitro Evolution and Chemogenomics. Science 2018, 359, 191–199. [Google Scholar] [CrossRef] [Green Version]
- Otto, T.D.; Wilinski, D.; Assefa, S.; Keane, T.M.; Sarry, L.R.; Böhme, U.; Lemieux, J.; Barrell, B.; Pain, A.; Berriman, M.; et al. New Insights into the Blood-Stage Transcriptome of Plasmodium falciparum Using RNA-Seq. Microbiology 2010, 76, 12–24. [Google Scholar] [CrossRef] [Green Version]
- Juge, N.; Moriyama, S.; Miyaji, T.; Kawakami, M.; Iwai, H.; Fukui, T.; Nelson, N.; Omote, H.; Moriyama, Y. Plasmodium falciparum Chloroquine Resistance Transporter Is a H+-Coupled Polyspecific Nutrient and Drug Exporter. Proc. Natl. Acad. Sci. USA 2015, 112, 3356–3361. [Google Scholar] [CrossRef] [Green Version]
- Shafik, S.H.; Cobbold, S.A.; Barkat, K.; Richards, S.N.; Lancaster, N.S.; Llinás, M.; Hogg, S.J.; Summers, R.L.; McConville, M.J.; Martin, R.E. The Natural Function of the Malaria Parasite’s Chloroquine Resistance Transporter. Nat. Commun. 2020, 11, 3922. [Google Scholar] [CrossRef] [PubMed]
- Lewis, I.A.; Wacker, M.; Olszewski, K.L.; Cobbold, S.A.; Baska, K.S.; Tan, A.; Ferdig, M.T.; Llinás, M. Metabolic QTL Analysis Links Chloroquine Resistance in Plasmodium Falciparum to Impaired Hemoglobin Catabolism. PLoS Genet. 2014, 10, e1004085. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Patzewitz, E.-M.; Salcedo-Sora, J.E.; Wong, E.H.; Sethia, S.; Stocks, P.A.; Maughan, S.C.; Murray, J.A.H.; Krishna, S.; Bray, P.G.; Ward, S.A.; et al. Glutathione Transport: A New Role for PfCRT in Chloroquine Resistance. Antioxid. Redox Signal. 2013, 19, 683–695. [Google Scholar] [CrossRef] [Green Version]
- Dou, Z.; McGovern, O.L.; Di Cristina, M.; Carruthers, V.B. Toxoplasma gondii Ingests and Digests Host Cytosolic Proteins. mBio 2014, 5, e01188-14. [Google Scholar] [CrossRef] [Green Version]
- Di Cristina, M.; Dou, Z.; Lunghi, M.; Kannan, G.; Huynh, M.-H.; McGovern, O.L.; Schultz, T.L.; Schultz, A.J.; Miller, A.J.; Hayes, B.M.; et al. Toxoplasma Depends on Lysosomal Consumption of Autophagosomes for Persistent Infection. Nat. Microbiol. 2017, 2, 17096. [Google Scholar] [CrossRef] [Green Version]
- McDonald, C.; Smith, D.; Di Cristina, M.; Kannan, G.; Dou, Z.; Carruthers, V.B. Toxoplasma Cathepsin Protease B and Aspartyl Protease 1 Are Dispensable for Endolysosomal Protein Digestion. mSphere 2020, 5, e00869-19. [Google Scholar] [CrossRef] [Green Version]
- Smith, D.; Kannan, G.; Coppens, I.; Wang, F.; Nguyen, H.M.; Cerutti, A.; Olafsson, E.B.; Rimple, P.A.; Schultz, T.L.; Mercado Soto, N.M.; et al. Toxoplasma TgATG9 Is Critical for Autophagy and Long-Term Persistence in Tissue Cysts. eLife 2021, 10, e59384. [Google Scholar] [CrossRef] [PubMed]
- Di Cristina, M.; Carruthers, V.B. New and Emerging Uses of CRISPR/Cas9 to Genetically Manipulate Apicomplexan Parasites. Parasitology 2018, 145, 1119–1126. [Google Scholar] [CrossRef] [Green Version]



{kind=link}
{kind=link}
| Class of Transporters | Sub-Class of Transporters | Porter Type | Energy-Dependent Transport | Substance Transport Direction | Typical Molecules Using Pathway | Refs. |
|---|---|---|---|---|---|---|
| Channels |
| Potential-dependent channel proteins—activated by a change in the membrane potential Ligand-dependent channel proteins—activated by binding to a ligand-mediator Mechanically dependent channel proteins—activated by mechanical deformation of the cell membrane | NO | Down concentration gradients | Ions: Na+, K+, Ca2+ | [14,15] |
| Active transporters |
| Importers, exporters and extruders | ATP hydrolysis | Against concentration gradients | Ions: Na+, K+, Ca2+, H+ | [12,16,17,18,19,20,21,22,23] |
| Secondary active transporters | SLCs:
| Symporter (cotransporter) and antiporter (exchanger) | Ion electrochemical gradient (cotransport of Na+, H+ or Cl− and/or the counter-transport of K+) | Against concentration gradients | Polar: amino acids, glucose, some ions | [24,25,26,27,28,29,30,31,32,33,34] |
| Facilitated transporters | SLCs:
| Uniporter | NO | Down concentration gradients | Polar: glucose | [25,26,27,35] |
| Toxoplasma gondii | Plasmodium falciparum | ID | Class | Putative Funtion | TM | Nutriens | Size aa | Refs. | |
|---|---|---|---|---|---|---|---|---|---|
| Host Cells | PfCLAG3.1 | PF3D7_0302500 | PSAC | Dimer/oligomer channel form | NO | Purine, sugar, amino acid, inorganic cations. | 1417 | [43] | |
| PfCLAG3.2 | PF3D7_0302200 | PSAC | Dimer/oligomer channel form | NO | Purine, sugar, amino acid, inorganic cations. | 1416 | [43] | ||
| PfRhopH2 | PF3D7_0929400 | PSAC | Dimer/oligomer channel form | NO | Purine, sugar, amino acid, inorganic cations. | 1378 | [43] | ||
| PfRhopH3 | PF3D7_0905400 | PSAC | Channel form | NO | Purine, sugar, amino acid, inorganic cations. | 897 | [43] | ||
| PVM | PfEXP1 | PF3D7_1121600 | PTEX | Pore exporting proteins and macromolecules | 1 | Host nutrients | 162 | [49,50] | |
| PfEXP2 | PF3D7_1471100 | PTEX | Pore exporting proteins and macromolecules | NO | Host nutrients | 287 | [49,50] | ||
| TgGRA17 | TGME49_222170 | PTEX Orthologus | Nutrient Transporters | NO | Host nutrients | 300 | [46,53,54] | ||
| TgGRA23 | TGME49_297880 | PTEX Orthologus | Nutrient Transporters | 1 | Host nutrients | 219 | [46,53] | ||
| TgABCG107 | TGME49_247540 | ATP-binding cassette subfamily G | Lipid import | 5 | Lipids | 981 | [61] | ||
| PPM | PfHT1 | PF3D7_0204700 | MSF | Sugar Transporters | 12 | Sugar | 504 | [62,63] | |
| TgGT1 | TGME49_214320 | MSF | Sugar Transporters | 12 | Sugar | 568 | [62,63] | ||
| TgST1 | TGME49_257120 | MSF | Sugar Transporters | 12 | Sugar | 601 | [62] | ||
| TgST2 | TGME49_272500 | MSF | Sugar Transporters | 12 | Sugar | 689 aa | [62] | ||
| TgST3 | TGME49_201260 | MSF | Sugar Transporters | 10 | Sugar | 693 | [62] | ||
| PfNT1 | PF3D7_1347200 | PfENT | Nucleotide Transporters | 9 | Purines | 422 | [64,65] | ||
| PfNT2 | PF3D7_0824400 | PfENT | Nucleotide Transporters | 10 | Purines | 585 | [66] | ||
| PfNT3 | PF3D7_1469400 | PfENT | Nucleotide Transporters | 11 | Purines | 437 | [65,67,68] | ||
| PfNT4 | PF3D7_0103200 | PfENT | Nucleotide Transporters | 11 | Purines | 434 | [65,67,68] | ||
| TgAT1 | TGME49_244440 | TgAT Family protein | Nucleotide Transporters | 10 | Purines | 462 | [69] | ||
| PfApiAT2 | PF3D7_0914700 | ApiAT | Amino Acid Transporters | 12 | ? | 516 | [70] | ||
| PfApiAT4 | PF3D7_1129900 | ApiAT | Amino Acid Transporters | 12 | ? | 609 | [70] | ||
| PfApiAT8 | PF3D7_0104800 | ApiAT | Amino Acid Transporters | 12 | Catonic Amino Acid | 577 | [70,71,72] | ||
| PfApiAT10 | PF3D7_0312500 | ApiAT | Amino Acid Transporters | 12 | ? | 579 | [70] | ||
| TgApiAT1 | TGME49_215490 | ApiAT | Amino Acid Transporters | 12 | Arginine | 534 | [73,74] | ||
| TgApiAT5-3 | TGME49_257530 | ApiAT | Amino Acid Transporters | 12 | Tyrosine, Phenilalanine, Triptofane, Leucine | 504 | [73] | ||
| TgApiAT6-1 | TGME49_240810 | ApiAT | Amino Acid Transporters | 10 | Lysine, Arginine | 566 | [75,76] | ||
| DV/VAC | PfAAT1 | PF3D7_0629500 | ApiAT | Amino Acid Transporters | 9 | Aromatic amino acid | 606 | [77] | |
| PfCRT | PF3D7_0709000 | DMT | Amino Acid Transporters | 10 | Arginine, Lysine, Histidine, Glutathione | 424 | [78,79] | ||
| TgCRT | TGME49_313930 | DMT | Amino Acid Transporters | 9 | Amino Acids | 881 | [80,81,82] |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Piro, F.; Focaia, R.; Dou, Z.; Masci, S.; Smith, D.; Di Cristina, M. An Uninvited Seat at the Dinner Table: How Apicomplexan Parasites Scavenge Nutrients from the Host. Microorganisms 2021, 9, 2592. https://doi.org/10.3390/microorganisms9122592
Piro F, Focaia R, Dou Z, Masci S, Smith D, Di Cristina M. An Uninvited Seat at the Dinner Table: How Apicomplexan Parasites Scavenge Nutrients from the Host. Microorganisms. 2021; 9(12):2592. https://doi.org/10.3390/microorganisms9122592
Chicago/Turabian StylePiro, Federica, Riccardo Focaia, Zhicheng Dou, Silvia Masci, David Smith, and Manlio Di Cristina. 2021. "An Uninvited Seat at the Dinner Table: How Apicomplexan Parasites Scavenge Nutrients from the Host" Microorganisms 9, no. 12: 2592. https://doi.org/10.3390/microorganisms9122592