
Evolution of Thermophilic Microbial Communities from a Deep-Sea Hydrothermal Chimney under Electrolithoautotrophic Conditions with Nitrate

 ,
,
Abstract
:1. Introduction
2. Materials and Methods
2.1. Sample Collection and Preparation
2.2. Electrotrophic Enrichment in a Microbial Electrochemical System
2.3. Nitrate/Nitrite Quantification
2.4. Identification and Quantification of Organic Compounds Production
2.5. Biodiversity Analysis
2.6. Microscopy Observation with Fluorescent In Situ Hybridization (FISH)
2.7. Quantitative PCR of Phylogenetic Orders
2.8. Principal Component Analysis
3. Results
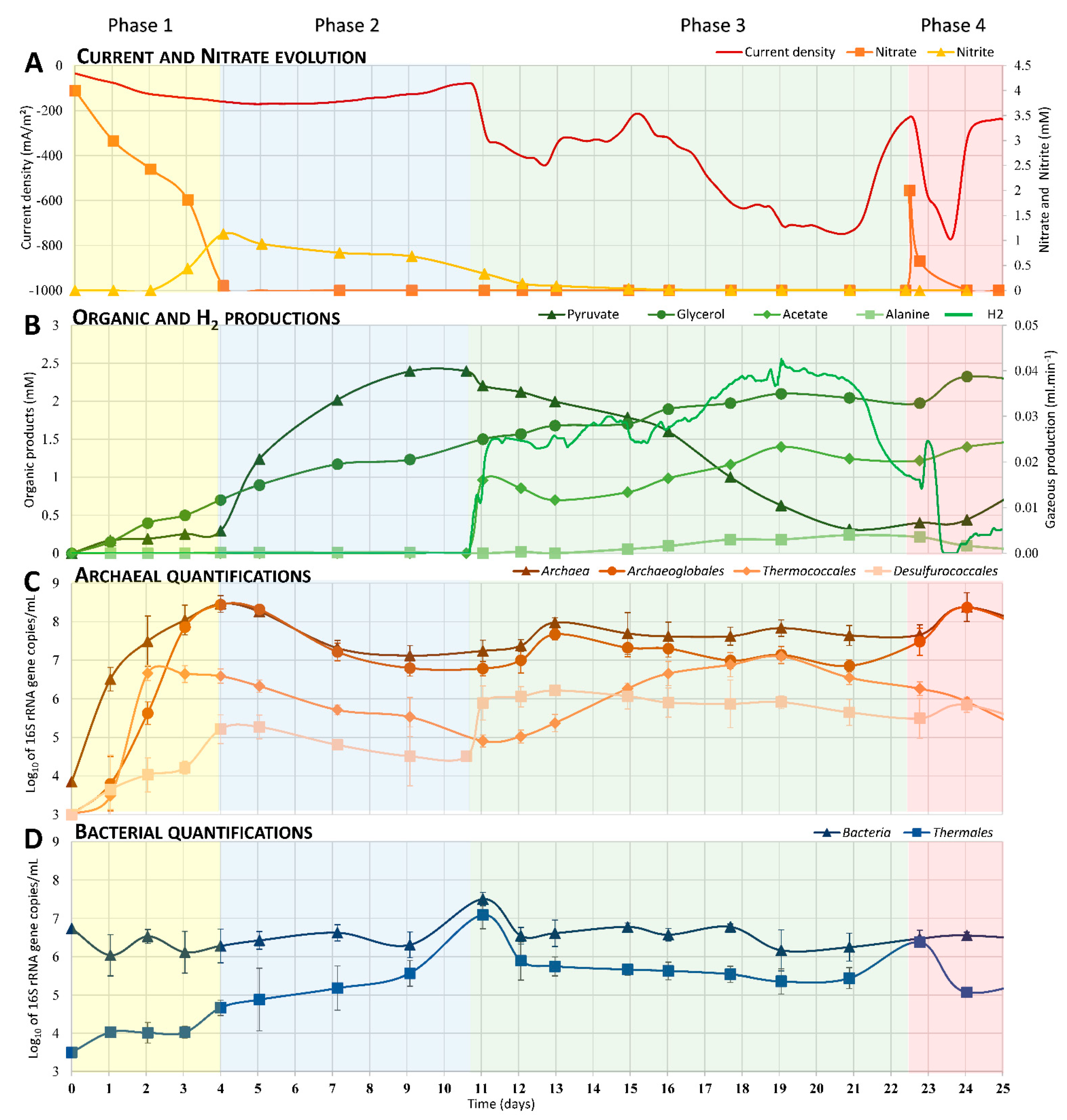
3.1. Current and Nitrate Consumptions
3.2. Electrosynthesis of Organic Compounds
3.3. Enrichment over Time of Dominant Phylogenetic Orders
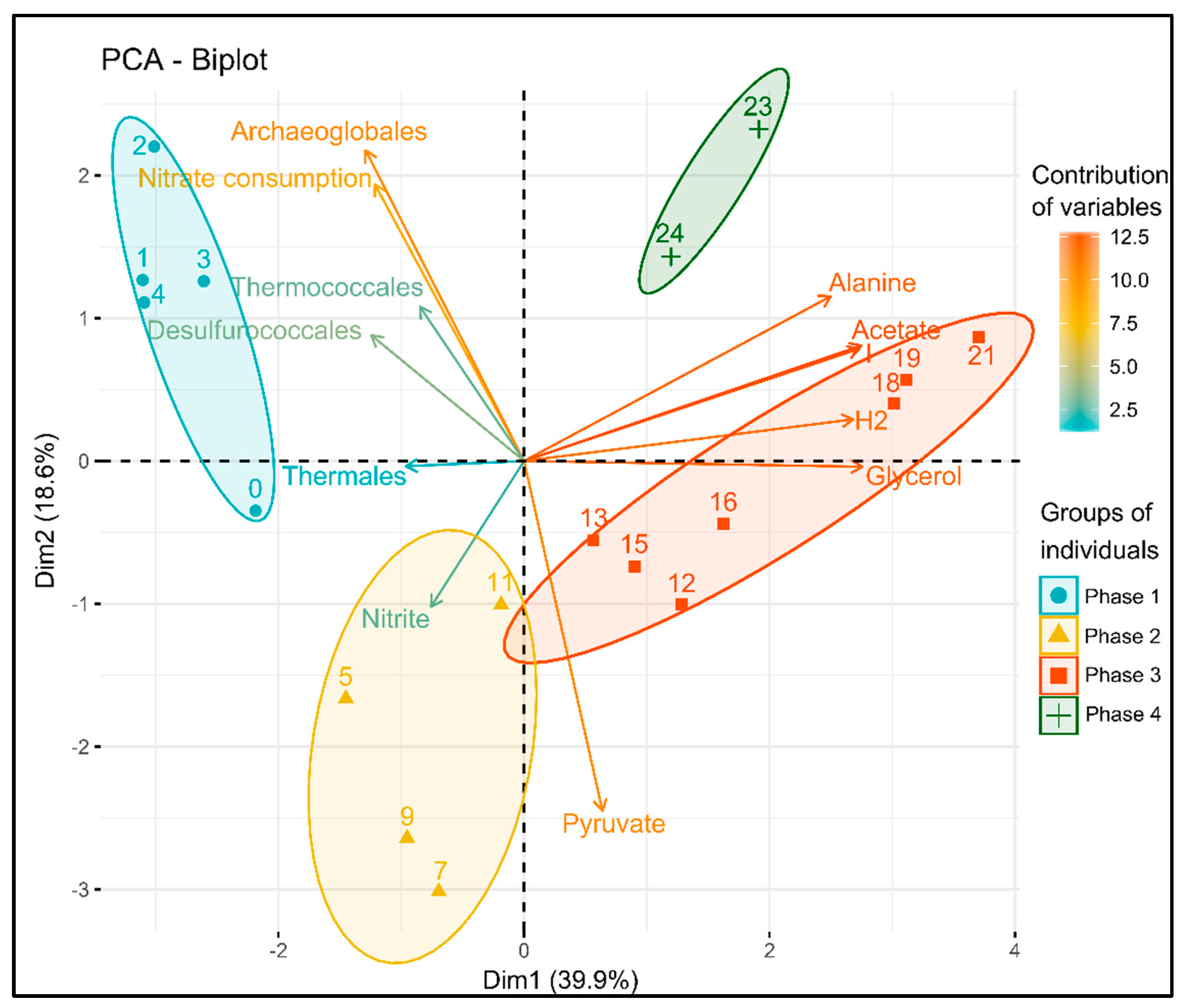
3.4. Statistical Analysis of the Correlation between Variables
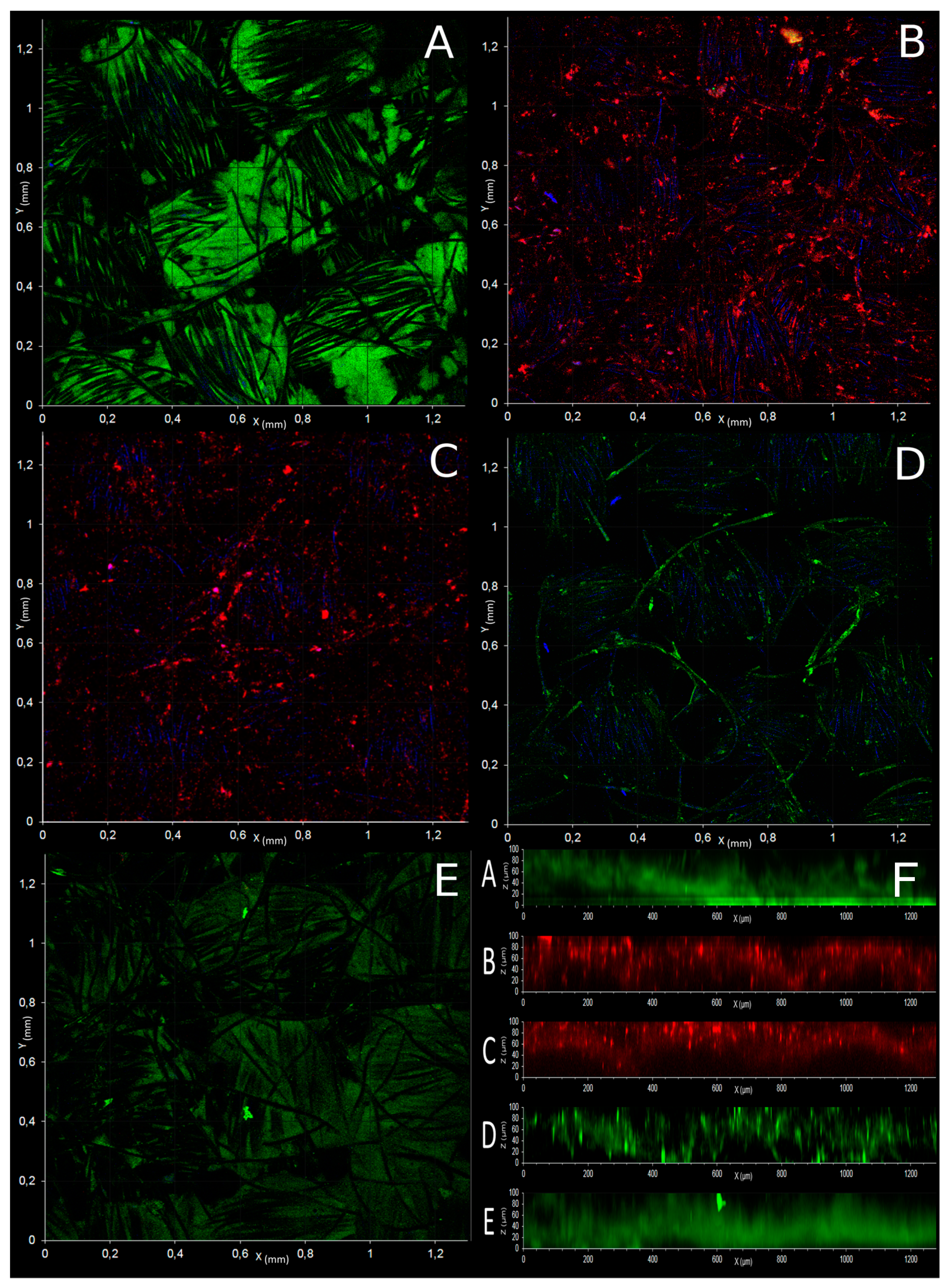
3.5. Microscopic Observation of Electrotrophic Biofilm
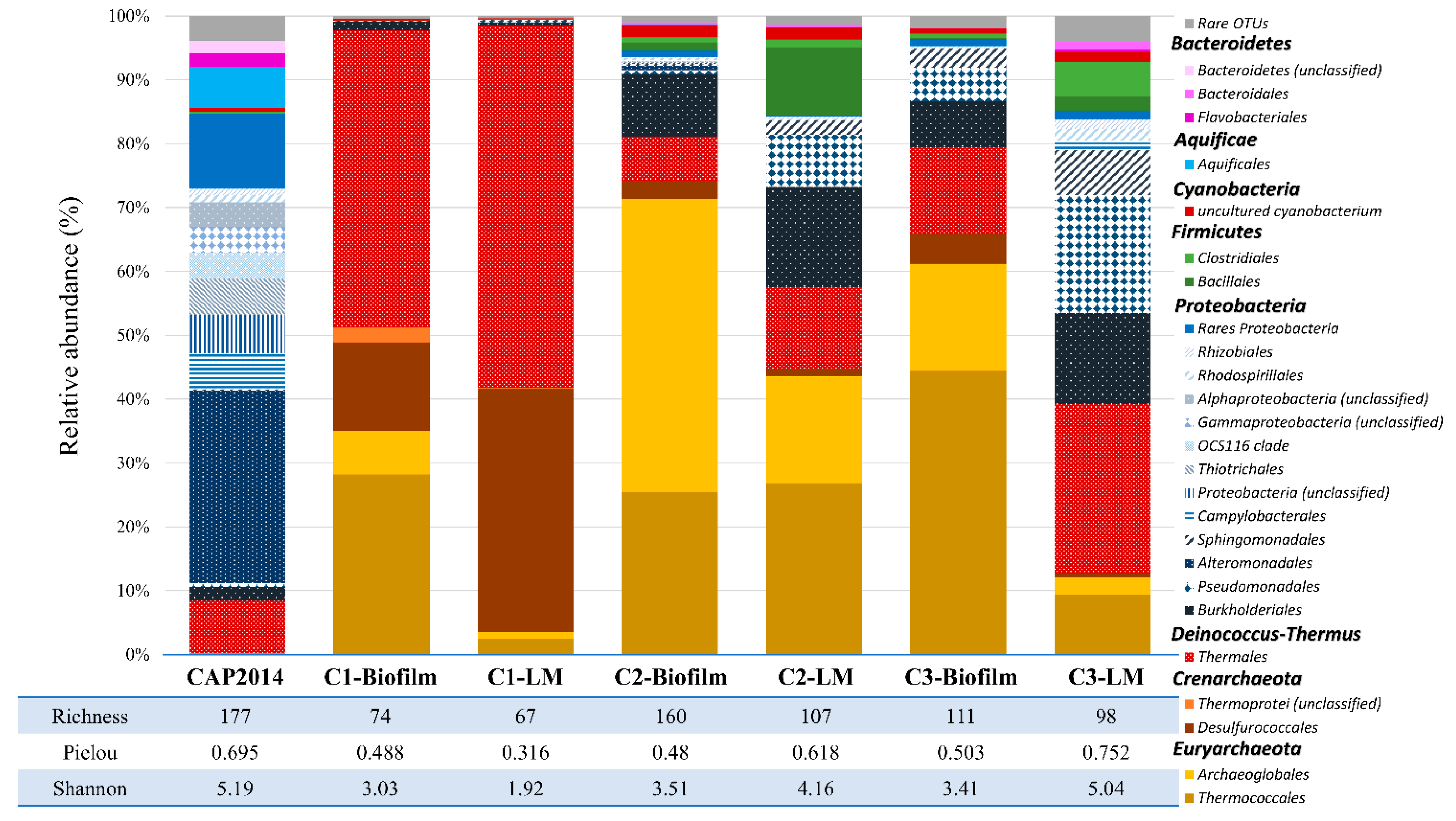
3.6. Biodiversity on Cathodes (Biofilms) and in Liquid Media (Planktonic Cells) of the Experiments
4. Discussion
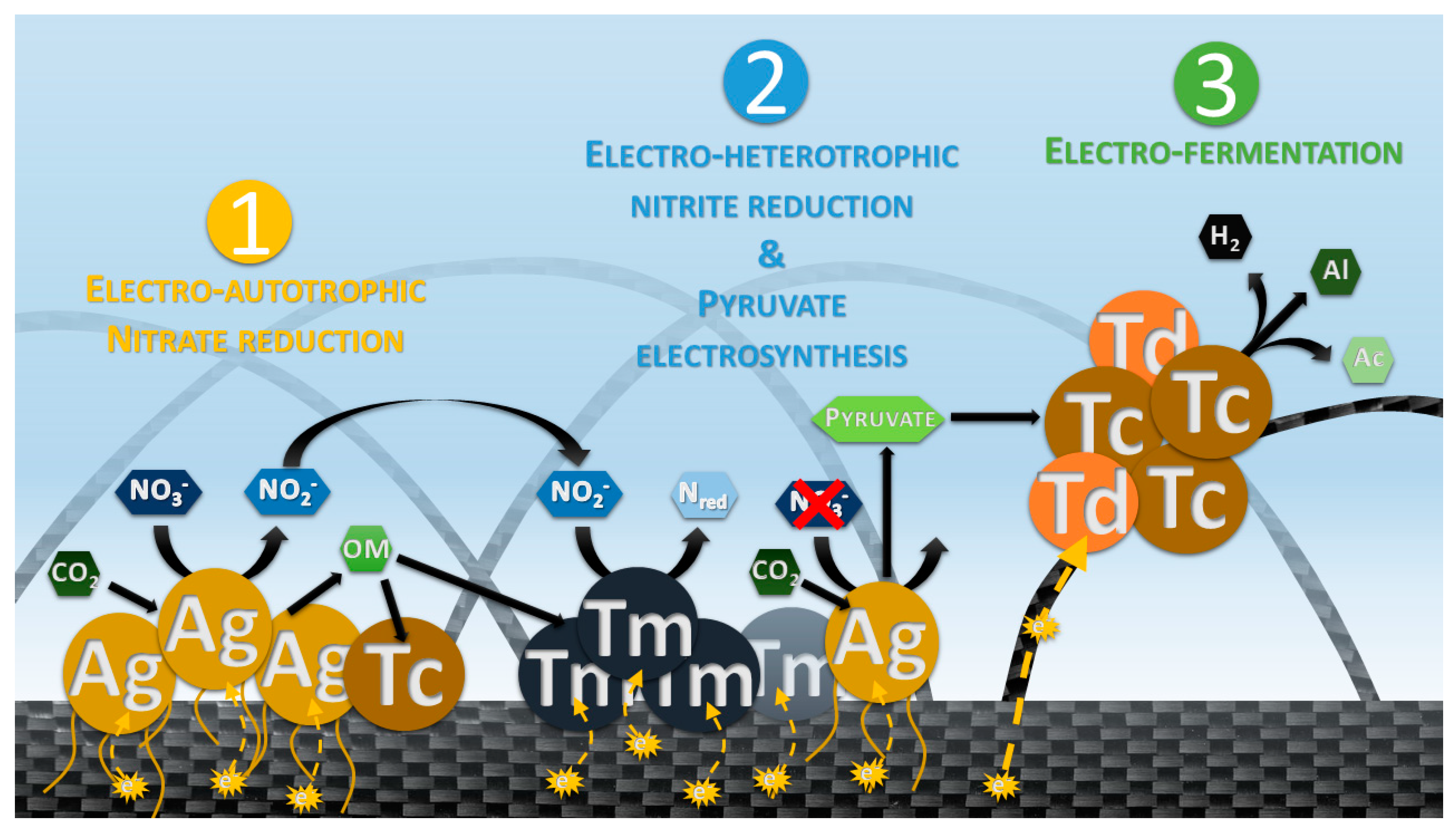
4.1. Phase 1: Electrotrophic Hyperthermophilic Nitrate Reduction
4.2. Phase 2: Electrosynthesis of Pyruvate
4.3. Phase 3: Electrofermentation of Pyruvate
4.4. Phase 4: The Trophic Chain from Electroautotrophic Nitrate Reduction to Heterotrophy
4.5. Thereafter: Development of More Complex Ecosystems over Subcultures
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Kristall, B.; Kelley, D.S.; Hannington, M.D.; Delaney, J.R. Growth History of a Diffusely Venting Sulfide Structure from the Juan de Fuca Ridge: A Petrological and Geochemical Study. Geochem. Geophys. Geosyst. 2006, 7. [Google Scholar] [CrossRef]
- Kelley, D.S.; Baross, J.A.; Delaney, J.R. Volcanoes, Fluids, and Life at Mid-Ocean Ridge Spreading Centers. Annu. Rev. Earth Planet. Sci. 2002, 30, 385–491. [Google Scholar] [CrossRef] [Green Version]
- Wirth, R. Colonization of Black Smokers by Hyperthermophilic Microorganisms. Trends Microbiol. 2017, 25, 92–99. [Google Scholar] [CrossRef]
- Pagé, A.; Margaret, T.K.; Debra, S.S.; Reysenbach, A. Temporal and Spatial Archaeal Colonization of Hydrothermal Vent Deposits. Environ. Microbiol. 2008, 10, 874–884. [Google Scholar] [CrossRef] [PubMed]
- McCliment, E.A.; Voglesonger, K.M.; O’Day, P.A.; Dunn, E.E.; Holloway, J.R.; Cary, S.C. Colonization of Nascent, Deep-sea Hydrothermal Vents by a Novel Archaeal and Nanoarchaeal Assemblage. Environ. Microbiol. 2005, 8, 114–125. [Google Scholar] [CrossRef] [PubMed]
- Reysenbach, A.-L.; Longnecker, K.; Kirshtein, J. Novel Bacterial and Archaeal Lineages from an in Situ Growth Chamber Deployed at a Mid-Atlantic Ridge Hydrothermal Vent. Appl. Environ. Microbiol. 2000, 66, 3798–3806. [Google Scholar] [CrossRef] [Green Version]
- Flores, G.E.; Campbell, J.H.; Kirshtein, J.D.; Meneghin, J.; Podar, M.; Steinberg, J.I.; Seewald, J.S.; Tivey, M.K.; Voytek, M.A.; Yang, Z.K.; et al. Microbial Community Structure of Hydrothermal Deposits from Geochemically Different Vent Fields along the Mid-Atlantic Ridge. Environ. Microbiol. 2011, 13, 2158–2171. [Google Scholar] [CrossRef] [PubMed]
- Wirth, R.; Luckner, M.; Wanner, G. Validation of a Hypothesis: Colonization of Black Smokers by Hyperthermophilic Microorganisms. Front. Microbiol. 2018, 9, 524. [Google Scholar] [CrossRef] [Green Version]
- Walker, D.J.; Adhikari, R.Y.; Holmes, D.E.; Ward, J.E.; Woodard, T.L.; Nevin, K.P.; Lovley, D.R. Electrically Conductive Pili from Pilin Genes of Phylogenetically Diverse Microorganisms. ISME J. 2017, 12, 48. [Google Scholar] [CrossRef] [Green Version]
- Yamamoto, M.; Nakamura, R.; Kasaya, T.; Kumagai, H.; Suzuki, K.; Takai, K. Spontaneous and Widespread Electricity Generation in Natural Deep-Sea Hydrothermal Fields. Angew. Chem. Int. Ed. 2017, 56, 5725–5728. [Google Scholar] [CrossRef]
- Lovley, D.R. Electromicrobiology. Annu. Rev. Microbiol. 2012, 66, 391–409. [Google Scholar] [CrossRef] [PubMed]
- He, Z.; Angenent, L.T. Application of Bacterial Biocathodes in Microbial Fuel Cells. Electroanalysis 2006, 18, 2009–2015. [Google Scholar] [CrossRef]
- Fu, Q.; Kobayashi, H.; Kuramochi, Y.; Xu, J.; Wakayama, T.; Maeda, H.; Sato, K. Bioelectrochemical Analyses of a Thermophilic Biocathode Catalyzing Sustainable Hydrogen Production. Int. J. Hydrog. Energy 2013, 38, 15638–15645. [Google Scholar] [CrossRef]
- Fu, Q.; Kuramochi, Y.; Fukushima, N.; Maeda, H.; Sato, K.; Kobayashi, H. Bioelectrochemical Analyses of the Development of a Thermophilic Biocathode Catalyzing Electromethanogenesis. Environ. Sci. Technol. 2015, 49, 1225–1232. [Google Scholar] [CrossRef]
- Luo, H.; Teng, W.; Liu, G.; Zhang, R.; Lu, Y. Sulfate Reduction and Microbial Community of Autotrophic Biocathode in Response to Acidity. Process Biochem. 2017, 54, 120–127. [Google Scholar] [CrossRef]
- Chaudhary, S.; Singh, R.; Yadav, S.; Patil, S.A. Electrochemical Enrichment of Haloalkaliphilic Nitrate-Reducing Microbial Biofilm at the Cathode of Bioelectrochemical Systems. iScience 2021, 24, 102682. [Google Scholar] [CrossRef] [PubMed]
- Aryal, N.; Tremblay, P.-L.; Lizak, D.M.; Zhang, T. Performance of Different Sporomusa Species for the Microbial Electrosynthesis of Acetate from Carbon Dioxide. Bioresour. Technol. 2017, 233, 184–190. [Google Scholar] [CrossRef]
- Beese-Vasbender, P.F.; Grote, J.-P.; Garrelfs, J.; Stratmann, M.; Mayrhofer, K.J.J. Selective Microbial Electrosynthesis of Methane by a Pure Culture of a Marine Lithoautotrophic Archaeon. Bioelectrochemistry 2015, 102, 50–55. [Google Scholar] [CrossRef]
- Pillot, G.; Amin Ali, O.; Davidson, S.; Shintu, L.; Godfroy, A.; Combet-Blanc, Y.; Bonin, P.; Liebgott, P.-P. Identification of Enriched Hyperthermophilic Microbial Communities from a Deep-Sea Hydrothermal Vent Chimney under Electrolithoautotrophic Culture Conditions. Sci. Rep. 2021, 11, 14782. [Google Scholar] [CrossRef]
- Wong, M.L.; Charnay, B.D.; Gao, P.; Yung, Y.L.; Russell, M.J. Nitrogen Oxides in Early Earth’s Atmosphere as Electron Acceptors for Life’s Emergence. Astrobiology 2017, 17, 975–983. [Google Scholar] [CrossRef] [Green Version]
- Vetriani, C.; Voordeckers, J.W.; Crespo-Medina, M.; O’Brien, C.E.; Giovannelli, D.; Lutz, R.A. Deep-Sea Hydrothermal Vent Epsilonproteobacteria Encode a Conserved and Widespread Nitrate Reduction Pathway (Nap). ISME J. 2014, 8, 1510–1521. [Google Scholar] [CrossRef] [PubMed]
- Sarradin, P.-M.; Cannat, M. MOMARSAT2014 Cruise, Pourquoi Pas ? R/V 2014. [Google Scholar] [CrossRef]
- Pillot, G.; Davidson, S.; Auria, R.; Combet-Blanc, Y.; Godfroy, A.; Liebgott, P.-P. Production of Current by Syntrophy Between Exoelectrogenic and Fermentative Hyperthermophilic Microorganisms in Heterotrophic Biofilm from a Deep-Sea Hydrothermal Chimney. Microb. Ecol. 2020, 79, 38–49. [Google Scholar] [CrossRef] [PubMed]
- Pillot, G.; Frouin, E.; Pasero, E.; Godfroy, A.; Combet-Blanc, Y.; Davidson, S.; Liebgott, P.-P. Specific Enrichment of Hyperthermophilic Electroactive Archaea from Deep-Sea Hydrothermal Vent on Electrically Conductive Support. Bioresour. Technol. 2018, 259, 304–311. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Aminot, A.; Kérouel, R. Dosage Automatique Des Nutriments Dans Les Eaux Marines, 2007th ed.; Méthodes d’analyse en milieu marin; Ifremer: Brest, France, 2007; ISBN 978-2-7592-0023-8. [Google Scholar]
- Bax, A.; Davis, D.G. MLEV-17-Based Two-Dimensional Homonuclear Magnetization Transfer Spectroscopy. J. Magn. Reson. 1985, 65, 355–360. [Google Scholar] [CrossRef]
- Schleucher, J.; Schwendinger, M.; Sattler, M.; Schmidt, P.; Schedletzky, O.; Glaser, S.J.; Sørensen, O.W.; Griesinger, C. A General Enhancement Scheme in Heteronuclear Multidimensional NMR Employing Pulsed Field Gradients. J. Biomol. NMR 1994, 4, 301–306. [Google Scholar] [CrossRef] [PubMed]
- Zhang, L.; Kang, M.; Xu, J.; Xu, J.; Shuai, Y.; Zhou, X.; Yang, Z.; Ma, K. Bacterial and Archaeal Communities in the Deep-Sea Sediments of Inactive Hydrothermal Vents in the Southwest India Ridge. Sci. Rep. 2016, 6. [Google Scholar] [CrossRef]
- Bates, S.T.; Berg-Lyons, D.; Caporaso, J.G.; Walters, W.A.; Knight, R.; Fierer, N. Examining the Global Distribution of Dominant Archaeal Populations in Soil. ISME J. 2011, 5, 908–917. [Google Scholar] [CrossRef] [Green Version]
- Kwak, T.J.; Peterson, J.T. Community Indices, Parameters, and Comparisons. In Analysis and Interpretation of Freshwater Fisheries Data; Guy, C.S., Brown, M.L., Eds.; American Fisheries Society: Bethesda, MD, USA, 2007; pp. 677–763. [Google Scholar]
- Rusch, A.; Amend, J.P. Order-Specific 16S RRNA-Targeted Oligonucleotide Probes for (Hyper)Thermophilic Archaea and Bacteria. Extremophiles 2004, 8, 357–366. [Google Scholar] [CrossRef]
- Teira, E.; Reinthaler, T.; Pernthaler, A.; Pernthaler, J.; Herndl, G.J. Combining Catalyzed Reporter Deposition-Fluorescence In Situ Hybridization and Microautoradiography To Detect Substrate Utilization by Bacteria and Archaea in the Deep Ocean. Appl. Environ. Microbiol. 2004, 70, 4411–4414. [Google Scholar] [CrossRef] [Green Version]
- Einen, J.; Thorseth, I.H.; Øvreås, L. Enumeration of Archaea and Bacteria in Seafloor Basalt Using Real-Time Quantitative PCR and Fluorescence Microscopy. FEMS Microbiol. Lett. 2008, 282, 182–187. [Google Scholar] [CrossRef] [Green Version]
- Michotey, V.; Guasco, S.; Boeuf, D.; Morezzi, N.; Durieux, B.; Charpy, L.; Bonin, P. Spatio-Temporal Diversity of Free-Living and Particle-Attached Prokaryotes in the Tropical Lagoon of Ahe Atoll (Tuamotu Archipelago) and Its Surrounding Oceanic Waters. Mar. Pollut. Bull. 2012, 65, 525–537. [Google Scholar] [CrossRef]
- Tamura, K.; Stecher, G.; Peterson, D.; Filipski, A.; Kumar, S. MEGA6: Molecular Evolutionary Genetics Analysis Version 6.0. Mol. Biol. Evol. 2013, 30, 2725–2729. [Google Scholar] [CrossRef] [Green Version]
- Deutzmann, J.S.; Sahin, M.; Spormann, A.M. Extracellular Enzymes Facilitate Electron Uptake in Biocorrosion and Bioelectrosynthesis. mBio 2015, 6, e00496-15. [Google Scholar] [CrossRef] [Green Version]
- Yilmazel, Y.D.; Zhu, X.; Kim, K.-Y.; Holmes, D.E.; Logan, B.E. Electrical Current Generation in Microbial Electrolysis Cells by Hyperthermophilic Archaea Ferroglobus Placidus and Geoglobus Ahangari. Bioelectrochemistry 2018, 119, 142–149. [Google Scholar] [CrossRef]
- Yee, M.O.; Rotaru, A.-E. Extracellular Electron Uptake in Methanosarcinales Is Independent of Multiheme c-Type Cytochromes. Sci. Rep. 2020, 10, 372. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ishii, T.; Kawaichi, S.; Nakagawa, H.; Hashimoto, K.; Nakamura, R. From Chemolithoautotrophs to Electrolithoautotrophs: CO2 Fixation by Fe(II)-Oxidizing Bacteria Coupled with Direct Uptake of Electrons from Solid Electron Sources. Front. Microbiol. 2015, 6. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Vorholt, J.A.; Hafenbradl, D.; Stetter, K.O.; Thauer, R.K. Pathways of Autotrophic CO2 Fixation and of Dissimilatory Nitrate Reduction to N2O in Ferroglobus Placidus. Arch. Microbiol. 1997, 167, 19–23. [Google Scholar] [CrossRef] [PubMed]
- Cabello, P.; Roldán, M.D.; Moreno-Vivián, C. Nitrate Reduction and the Nitrogen Cycle in Archaea. Microbiology 2004, 150, 3527–3546. [Google Scholar] [CrossRef]
- von Jan, M.; Lapidus, A.; Glavina Del Rio, T.; Copeland, A.; Tice, H.; Cheng, J.-F.; Lucas, S.; Chen, F.; Nolan, M.; Goodwin, L.; et al. Complete Genome Sequence of Archaeoglobus Profundus Type Strain (AV18T). Stand. Genom. Sci. 2010, 2, 327–346. [Google Scholar] [CrossRef] [Green Version]
- Rosenberg, E.; DeLong, E.F.; Lory, S.; Stackebrandt, E.; Thompson, F. (Eds.) The Prokaryotes: Other Major Lineages of Bacteria and The Archaea, 4th ed.; Springer: Berlin/Heidelberg, Germany, 2014; ISBN 978-3-642-38953-5. [Google Scholar]
- Schut, G.J.; Lipscomb, G.L.; Han, Y.; Notey, J.S.; Kelly, R.M.; Adams, M.M.W. The Order Thermococcales and the Family Thermococcaceae. In The Prokaryotes; Rosenberg, E., DeLong, E.F., Lory, S., Stackebrandt, E., Thompson, F., Eds.; Springer: Berlin/Heidelberg, Germany, 2014; pp. 363–383. ISBN 978-3-642-38953-5. [Google Scholar]
- Chou, C.-J.; Shockley, K.R.; Conners, S.B.; Lewis, D.L.; Comfort, D.A.; Adams, M.W.W.; Kelly, R.M. Impact of Substrate Glycoside Linkage and Elemental Sulfur on Bioenergetics of and Hydrogen Production by the Hyperthermophilic Archaeon Pyrococcus Furiosus. Appl. Environ. Microbiol. 2007, 73, 6842–6853. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sokolova, T.G.; Jeanthon, C.; Kostrikina, N.A.; Chernyh, N.A.; Lebedinsky, A.V.; Stackebrandt, E.; Bonch-Osmolovskaya, E.A. The First Evidence of Anaerobic CO Oxidation Coupled with H2 Production by a Hyperthermophilic Archaeon Isolated from a Deep-Sea Hydrothermal Vent. Extremophiles 2004, 8, 317–323. [Google Scholar] [CrossRef] [PubMed]
- Lee, H.S.; Kang, S.G.; Bae, S.S.; Lim, J.K.; Cho, Y.; Kim, Y.J.; Jeon, J.H.; Cha, S.-S.; Kwon, K.K.; Kim, H.-T.; et al. The Complete Genome Sequence of Thermococcus Onnurineus NA1 Reveals a Mixed Heterotrophic and Carboxydotrophic Metabolism. J. Bacteriol. 2008, 190, 7491–7499. [Google Scholar] [CrossRef] [Green Version]
- Oger, P.; Sokolova, T.G.; Kozhevnikova, D.A.; Chernyh, N.A.; Bartlett, D.H.; Bonch-Osmolovskaya, E.A.; Lebedinsky, A.V. Complete Genome Sequence of the Hyperthermophilic Archaeon Thermococcus Sp. Strain AM4, Capable of Organotrophic Growth and Growth at the Expense of Hydrogenogenic or Sulfidogenic Oxidation of Carbon Monoxide. J. Bacteriol. 2011, 193, 7019–7020. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bae, S.S.; Kim, T.W.; Lee, H.S.; Kwon, K.K.; Kim, Y.J.; Kim, M.-S.; Lee, J.-H.; Kang, S.G. H2 Production from CO, Formate or Starch Using the Hyperthermophilic Archaeon, Thermococcusonnurineus. Biotechnol. Lett. 2012, 34, 75–79. [Google Scholar] [CrossRef] [PubMed]
- Huber, H.; Stetter, K.O. Desulfurococcales Ord. Nov. In Bergey’s Manual of Systematics of Archaea and Bacteria; Whiteman, W.B., Ed.; John Wiley & Sons, Ltd.: New York, NY, USA, 2015; pp. 1–2. ISBN 978-1-118-96060-8. [Google Scholar]
- Albuquerque, L.; da Costa, M.S. The Family Thermaceae. In The Prokaryotes; Springer: Berlin/Heidelberg, Germany, 2014; pp. 955–987. ISBN 978-3-642-38953-5. [Google Scholar]
- Borrel, G.; Adam, P.S.; Gribaldo, S. Methanogenesis and the Wood–Ljungdahl Pathway: An Ancient, Versatile, and Fragile Association. Genome Biol. Evol. 2016, 8, 1706–1711. [Google Scholar] [CrossRef] [Green Version]
- Berg, I.A.; Kockelkorn, D.; Ramos-Vera, W.H.; Say, R.F.; Zarzycki, J.; Hügler, M.; Alber, B.E.; Fuchs, G. Autotrophic Carbon Fixation in Archaea. Nat. Rev. Microbiol. 2010, 8, 447–460. [Google Scholar] [CrossRef]
- Fuchs, G. Alternative Pathways of Carbon Dioxide Fixation: Insights into the Early Evolution of Life? Annu. Rev. Microbiol. 2011, 65, 631–658. [Google Scholar] [CrossRef]
- Utter, M.; Keech, D. Pyruvate Carboxylase. I. Nature of the Reaction. J. Biol. Chem. 1963, 238, 2603–2608. [Google Scholar] [CrossRef]
- Blamey, J.M.; Adams, M.W.W. Purification and Characterization of Pyruvate Ferredoxin Oxidoreductase from the Hyperthermophilic Archaeon Pyrococcus Furiosus. Biochim. Biophys. Acta (BBA) 1993, 1161, 19–27. [Google Scholar] [CrossRef]
- Huber, H.; Stetter, K.O. Desulfurococcales. In The prokaryotes; Springer: Berlin/Heidelberg, Germany, 2006; pp. 52–68. [Google Scholar]
- Ward, D.E.; Kengen, S.W.M.; van der Oost, J.; de Vos, W.M. Purification and Characterization of the Alanine Aminotransferase from the Hyperthermophilic Archaeon Pyrococcus Furiosus and Its Role in Alanine Production. J. Bacteriol. 2000, 182, 2559–2566. [Google Scholar] [CrossRef] [Green Version]
- Lovley, D.R. Syntrophy Goes Electric: Direct Interspecies Electron Transfer. Annu. Rev. Microbiol. 2017, 71, 643–664. [Google Scholar] [CrossRef]
- Rotaru, A.-E.; Shrestha, P.M.; Liu, F.; Markovaite, B.; Chen, S.; Nevin, K.P.; Lovley, D.R. Direct Interspecies Electron Transfer between Geobacter Metallireducens and Methanosarcina Barkeri. Appl. Environ. Microbiol. 2014, 80, 4599–4605. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yuan, J.; Lai, Q.; Zheng, T.; Shao, Z. Novosphingobium Indicum Sp. Nov., a Polycyclic Aromatic Hydrocarbon-Degrading Bacterium Isolated from a Deep-Sea Environment. Int. J. Syst. Evol. Microbiol. 2009, 59, 2084–2088. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Dong, C.; Chen, L.; Liao, Y.; Shao, Z. Phylogenetic and degrading genes analysis of a PAH-degrading bacterium TVG9-VII from deep-sea hydrythermal environment. Wei Sheng Wu Xue Bao 2011, 51, 1548–1554. [Google Scholar] [PubMed]
- Fortunato, C.S.; Huber, J.A. Coupled RNA-SIP and Metatranscriptomics of Active Chemolithoautotrophic Communities at a Deep-Sea Hydrothermal Vent. ISME J. 2016, 10, 1925–1938. [Google Scholar] [CrossRef]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Phylogenetic Groups | ||||
|---|---|---|---|---|
| Target | Name | Sequence | Thyb | Ref. |
| Bacteria | GML5F | GCCTACGGGAGGCAGCAG | 55 °C | [34] |
| Univ516 | GTDTTACCGCGGCKGCTGRCA | |||
| Archaea | Arc931F | AGGAATTGGCGGGGGAGCA | 62 °C | [33] |
| m1100R | BTGGGTCTCGCTCGTTRC C | |||
| Thermococcales | Tcoc_F959 | CGTGAGGCGTCCACTTAAGTGTGGT | 63 °C | This study |
| Tcoc_R1233 | GATGATGACRCGCGGGTACTAGGG | |||
| Archaeoglobales | Arglo_F1077 | CGGGCAACGGCAGGTCCGTATG | 62 °C | |
| Arglo_R1191 | GTTGCAGCCCTCGATCCCAGGT | |||
| Desulfurococcales | Univ516F | TGYCAGCMGCCGCGGTAAHAC | 61 °C | |
| DSC_748R | AACASYTAGCCCGCATCGTTTACAGCC | |||
| Thermales | Therma/Deino_341F | GGAGGCAGCAGTTAGGAATCTTC | 59 °C | |
| Univ516R | GTDTTACCGCGGCKGCTGRCA | |||
| Fish Probes | ||||
| Target | Name | Sequence | Label | Ref. |
| Bacteria | EUB338 | GCT GCC TCC CGT AGG AGT | FITC | [31,32] |
| Archaea | ARCH917 | GTG CTC CCC CGC CAA TTC | CY3 | |
| Euryarchaeota | Eury806 | CAC AGC GTT TAC ACC TAG | CY3 | |
| Crenarchaeota | Cren537 | TGA CCA CTT GAG GTG CTG | CY3 | |
| Thermococcales | Tcoc164 | CAV RCC TAT GGG GGA TTA GC | FITC | |
| Archaeoglobales | Arglo32 | TTA GTC CCA GCC GGA TAG CA | FITC | |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Pillot, G.; Amin Ali, O.; Davidson, S.; Shintu, L.; Combet-Blanc, Y.; Godfroy, A.; Bonin, P.; Liebgott, P.-P. Evolution of Thermophilic Microbial Communities from a Deep-Sea Hydrothermal Chimney under Electrolithoautotrophic Conditions with Nitrate. Microorganisms 2021, 9, 2475. https://doi.org/10.3390/microorganisms9122475
Pillot G, Amin Ali O, Davidson S, Shintu L, Combet-Blanc Y, Godfroy A, Bonin P, Liebgott P-P. Evolution of Thermophilic Microbial Communities from a Deep-Sea Hydrothermal Chimney under Electrolithoautotrophic Conditions with Nitrate. Microorganisms. 2021; 9(12):2475. https://doi.org/10.3390/microorganisms9122475
Chicago/Turabian StylePillot, Guillaume, Oulfat Amin Ali, Sylvain Davidson, Laetitia Shintu, Yannick Combet-Blanc, Anne Godfroy, Patricia Bonin, and Pierre-Pol Liebgott. 2021. "Evolution of Thermophilic Microbial Communities from a Deep-Sea Hydrothermal Chimney under Electrolithoautotrophic Conditions with Nitrate" Microorganisms 9, no. 12: 2475. https://doi.org/10.3390/microorganisms9122475