
Characterization of Phosphate Solubilizing Bacterial Endophytes and Plant Growth Promotion In Vitro and in Greenhouse

 ,
,  , ,
, ,
Abstract
:1. Introduction
2. Materials and Methods
2.1. Isolation of Bacterial Endophytes
2.2. Screening Phosphate Solubilizing Bacteria by Quantifying Soluble P in Medium
2.3. Acid Phosphatase and Phytase Activity Assays
2.4. Other Plant Growth Promoting Traits of Phosphate Solubilizing Bacteria
2.4.1. Auxin Quantification
2.4.2. ACC Deaminase Activity Screening
2.4.3. Siderophore Assay
2.4.4. N Fixation Screening
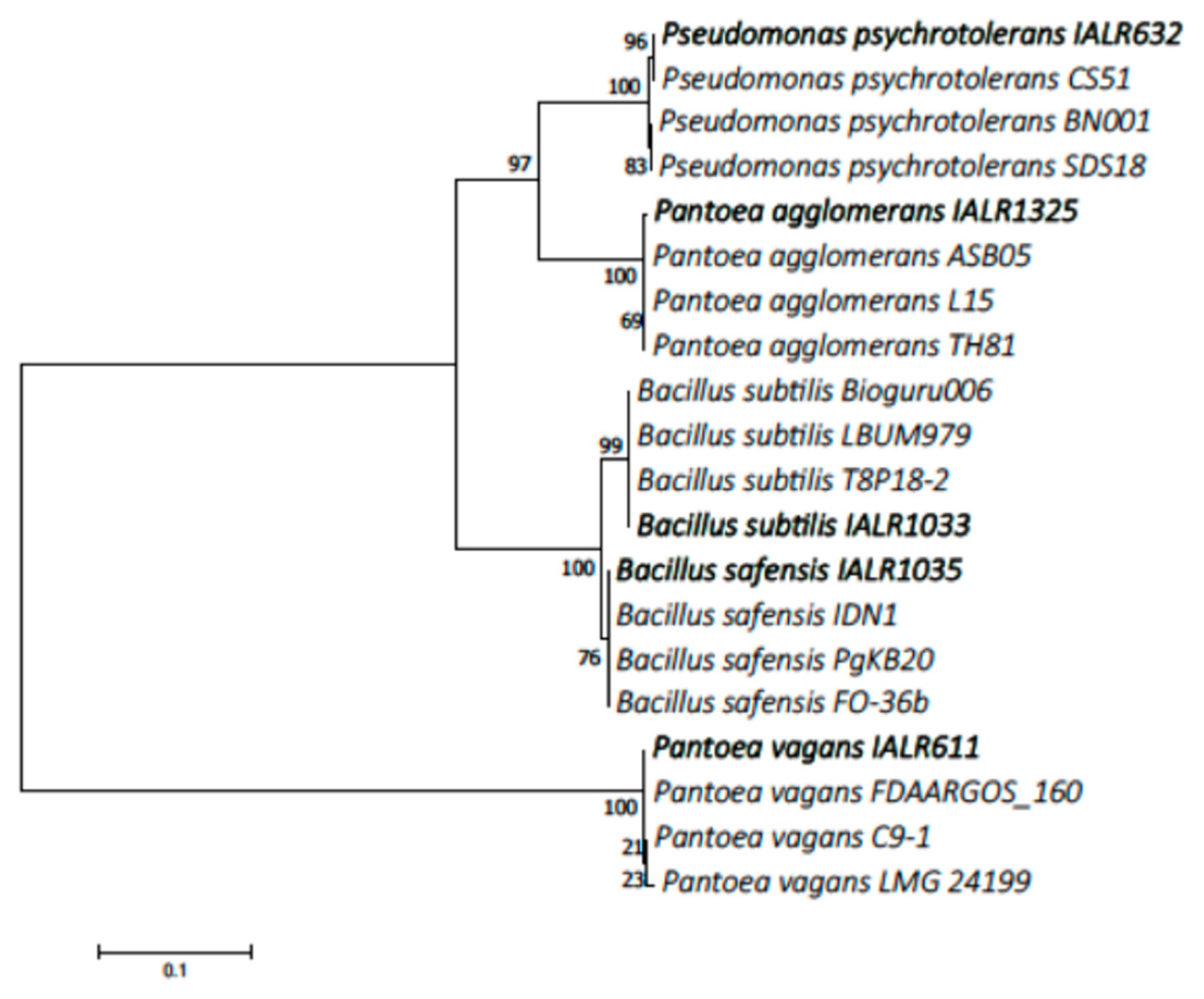
2.5. Identification of Phosphate Solubilizing Bacteria and Phylogenetic Tree Construction
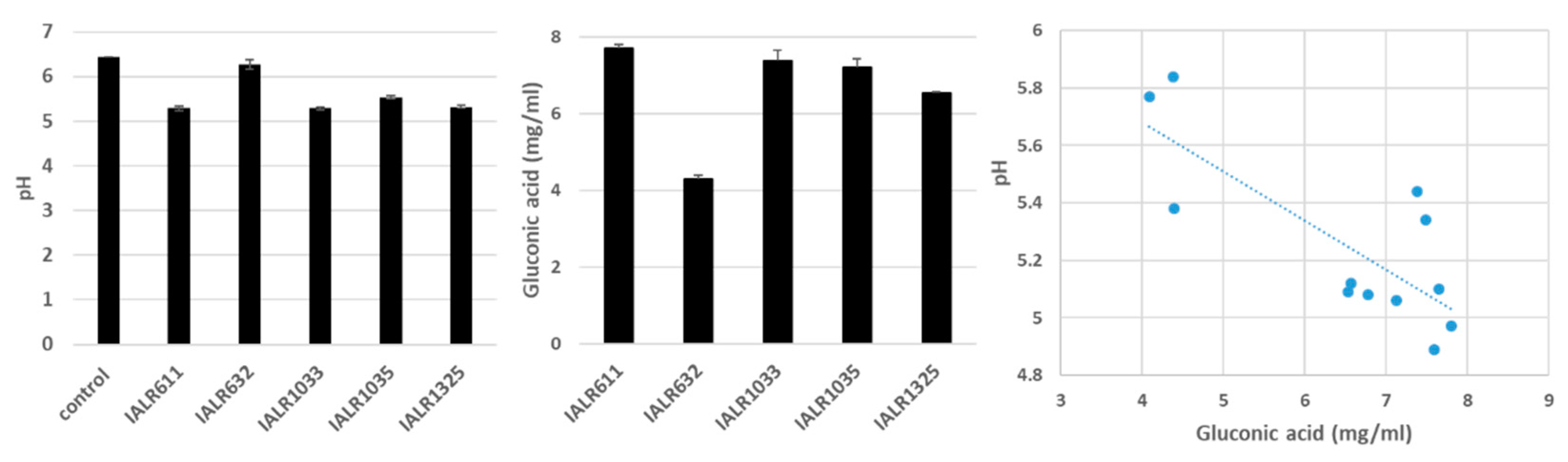
2.6. pH Measurement and Gluconic Acid Quantification
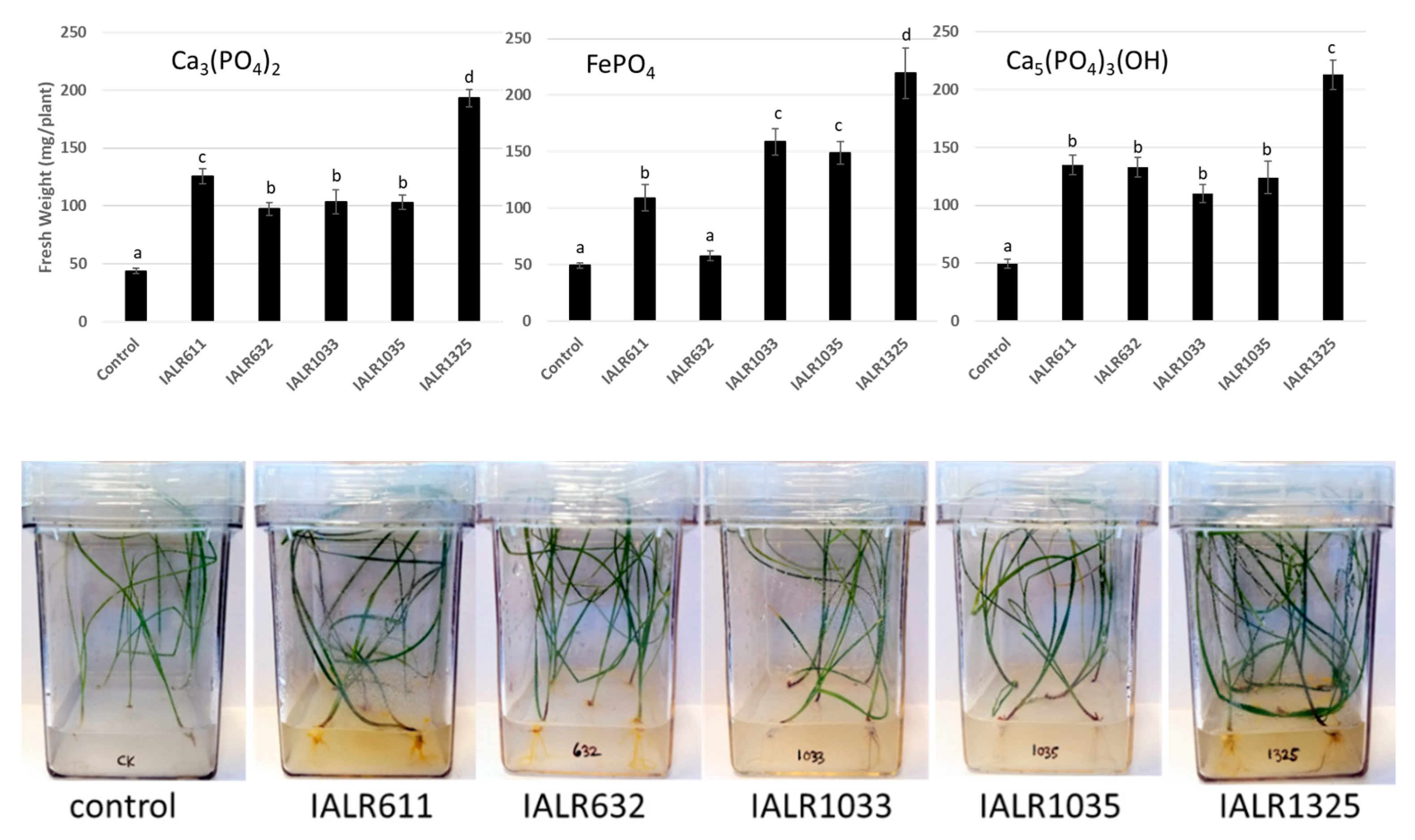
2.7. Plant Growth Promotion In Vitro
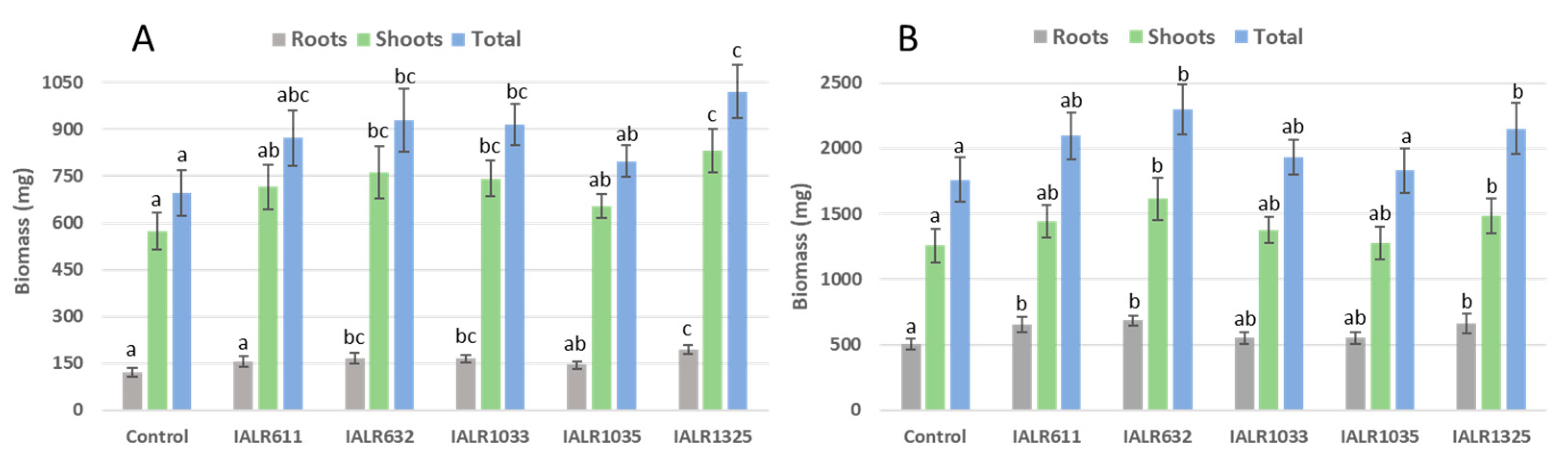
2.8. Plant Growth Promotion in Greenhouse
2.9. Statistical Analyses
3. Results
3.1. Identification of Phosphate Solubilizing Bacteria and Phylogenetic Tree
3.2. Phosphate Solubilizing Ability of Bacteria
3.3. Activities of Acid Phosphatase and Phytase of Phosphate Solubilizing Bacteria
3.4. Other Plant Growth Promoting Traits of the Phosphate Solubilizing Bacteria
3.5. pH Changes and Gluconic Acid Production
3.6. Plant Growth Promotion In Vitro
3.7. Growth Promotion by Phosphate Solubilizing Bacteria in Greenhouse Experiments
4. Discussion
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Conflicts of Interest
References
- Alori, E.T.; Glick, B.R.; Babalola, O.O. Microbial phosphorus solubilization and its potential for use in sustainable agriculture. Front. Microbiol. 2017, 8, 971. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sharma, S.B.; Sayyed, R.Z.; Trivedi, M.H.; Gobi, T.A. Phosphate solubilizing microbes: Sustainable approach for managing phosphorus deficiency in agricultural soils. SpringerPlus 2013, 2, 1–14. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Richardson, A.E. Prospects for using soil microorganisms to improve the acquisition of phosphorus by plants. Funct. Plant Biol. 2001, 28, 897–906. [Google Scholar] [CrossRef]
- Zhu, F.; Qu, L.; Hong, X.; Sun, X. Isolation and characterization of a phosphate-solubilizing halophilic bacterium Kushneria sp. YCWA18 from Daqiao Saltern on the coast of Yellow Sea of China. Evid. Based. Complement. Altern. Med. 2011. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ohio City Votes to Give Lake Erie Personhood Status over Algae Blooms. Available online: https://www.theguardian.com/us-news/2019/feb/28/toledo-lake-erie-personhood-status-bill-of-rights-algae-bloom (accessed on 3 April 2021).
- Malik, M.A.; Marschner, P.; Khan, K.S. Addition of organic and inorganic P sources to soil–Effects on P pools and microorganisms. Soil Biol. Biochem. 2012, 49, 106–113. [Google Scholar] [CrossRef]
- Zaidi, A.; Khan, M.; Ahemad, M.; Oves, M. Plant growth promotion by phosphate solubilizing bacteria. Acta. Microbiol. Immunol. Hung. 2009, 56, 263–284. [Google Scholar] [CrossRef]
- Song, O.R.; Lee, S.J.; Lee, Y.S.; Lee, S.C.; Kim, K.K.; Choi, Y.L. Solubilization of insoluble inorganic phosphate by Burkholderia cepacia DA23 isolated from cultivated soil. Braz. J. Microbiol. 2008, 39, 151–156. [Google Scholar] [CrossRef] [Green Version]
- Poonguzhali, S.; Madhaiyan, M.; Sa, T.M. Isolation and identification of phosphate solubilizing bacteria from Chinese cabbage and their effect on growth and phosphorus utilization of plants. J. Microbiol. Biotechnol. 2008, 18, 773–777. [Google Scholar]
- Sharon, J.A.; Hathwaik, L.T.; Glenn, G.M.; Imam, S.H.; Lee, C.C. Isolation of efficient phosphate solubilizing bacteria capable of enhancing tomato plant growth. J. Soil. Sci. Plant Nutr. 2016, 16, 525–536. [Google Scholar] [CrossRef] [Green Version]
- Lucero, C.T.; Lorda, G.S.; Anzuay, M.S.; Ludueña, L.M.; Taurian, T. Peanut endophytic phosphate solubilizing bacteria increase growth and P content of soybean and maize plants. Curr. Microbiol. 2021, 78, 1961–1972. [Google Scholar] [CrossRef] [PubMed]
- Behera, B.C.; Yadav, H.; Singh, S.K.; Mishra, R.R.; Sethi, B.K.; Dutta, S.K.; Thatoi, H.N. Phosphate solubilization and acid phosphatase activity of Serratia sp. isolated from mangrove soil of Mahanadi river delta, Odisha, India. J. Genet. Eng. Biotechnol. 2017, 15, 169–178. [Google Scholar] [CrossRef]
- Rasul, M.; Yasmin, S.; Suleman, M.; Zaheer, A.; Reitz, T.; Tarkka, M.T.; Islam, E.; Mirza, M.S. Glucose dehydrogenase gene containing phosphobacteria for biofortification of Phosphorus with growth promotion of rice. Microbiol. Res. 2019, 223, 1–12. [Google Scholar] [CrossRef] [PubMed]
- Richardson, A.E.; Lynch, J.P.; Ryan, P.R.; Delhaize, E.; Smith, F.A.; Smith, S.E.; Harvey, P.R.; Ryan, M.H.; Veneklaas, E.J.; Lambers, H.; et al. Plant and microbial strategies to improve the phosphorus efficiency of agriculture. Plant Soil. 2011, 349, 121–156. [Google Scholar] [CrossRef]
- Sashidhar, B.; Podile, A.R. Mineral phosphate solubilization by rhizosphere bacteria and scope for manipulation of the direct oxidation pathway involving glucose dehydrogenase. J. Appl. Microbiol. 2010, 109, 1–12. [Google Scholar] [CrossRef] [PubMed]
- Nautiyal, C.S. An efficient microbiological growth medium for screening phosphate solubilizing microorganisms. FEMS Microbiol. Lett. 1999, 170, 265–270. [Google Scholar] [CrossRef] [PubMed]
- Murphy, J.A.M.E.S.; Riley, J.P. A modified single solution method for the determination of phosphate in natural waters. Anal. Chim. Acta. 1962, 27, 31–36. [Google Scholar] [CrossRef]
- Rombola, T.H.; Pedrinho, E.A.N.; de Macedo Lemos, E.G.; Gonçalves, A.M.; dos Santos, L.F.J.; Pizauro, J.M. Identification and enzymatic characterization of acid phosphatase from Burkholderia gladioli. BMC Res. Notes 2014, 7, 1–7. [Google Scholar] [CrossRef] [Green Version]
- Parhamfar, M.; Badoei-Dalfard, A.; Parhamfar, M.; Fahimi Rad, S. Purification and characterization of an extracellular phosphatase enzyme from Bacillus spp. J. Cell Mol. Res. 2016, 8, 90–97. [Google Scholar]
- Patten, C.L.; Glick, B.R. Role of Pseudomonas putida indoleacetic acid in development of the host plant root system. Appl. Environ. Microbiol. 2002, 68, 3795–3801. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kim-Dura, S.; Lowman, S.; Zhang, S.; Mei, C. Growth promotion of switchgrass by bacterial endophyte Pantoea agglomerans strain PaKM isolated from seeds. J. Pathol. Microbiol. 2016, 1, 1–9. [Google Scholar]
- Sun, Y.; Cheng, Z.; Glick, B.R. The presence of a 1-aminocyclopropane-1-carboxylate (ACC) deaminase deletion mutation alters the physiology of the endophytic plant growth-promoting bacterium Burkholderia phytofirmans PsJN. FEMS Microbiol. Lett. 2009, 296, 131–136. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Payne, S.M. Detection, isolation, and characterization of siderophores. Methods Enzym. 1994, 235, 329–344. [Google Scholar]
- Pathak, L.; Kalekar, S. Exploring nitrogen fixing, chemo heterotrophic oligophiles from natural habitats for preparing biofertilizers. Sci. Rep. 2012, 1, 555. [Google Scholar] [CrossRef]
- Kumar, S.; Stecher, G.; Tamura, K. MEGA7: Molecular evolutionary genetics analysis version 7.0 for bigger datasets. Mol. Biol. Evol. 2016, 33, 1870–1874. [Google Scholar] [CrossRef] [Green Version]
- Bindraban, P.S.; Dimkpa, C.O.; Pandey, R. Exploring phosphorus fertilizers and fertilization strategies for improved human and environmental health. Biol. Fertil. Soils 2020, 56, 299–317. [Google Scholar] [CrossRef] [Green Version]
- Hayatsu, M. Utilization of phytic acid by cooperative interaction in rhizosphere. Microbes. Environ. 2013, 28, 1–2. [Google Scholar] [CrossRef] [Green Version]
- Unno, Y.; Shinano, T. Metagenomic analysis of the rhizospere soil microbiome with respect to phytic acid utilization. Microbes. Environ. 2013, 28, 120–127. [Google Scholar] [CrossRef] [Green Version]
- Otieno, N.; Lally, R.D.; Kiwanuka, S.; Lloyd, A.; Ryan, D.; Germaine, K.J.; Dowling, D.N. Plant growth promotion induced by phosphate solubilizing endophytic Pseudomonas isolates. Front. Microbiol. 2015, 6, 745. [Google Scholar] [CrossRef] [Green Version]
- Chen, J.; Zhao, G.; Wei, Y.; Dong, Y.; Hou, L.; Jiao, R. Isolation and screening of multifunctional phosphate solubilizing bacteria and its growth-promoting effect on Chinese fir seedlings. Sci. Rep. 2021, 11, 1–13. [Google Scholar]
- Walia, A.; Guleria, S.; Chauhan, A.; Mehta, P. Endophytic bacteria: Role in phosphate solubilization. In Endophytes: Crop Productivity and Protection; 2017; pp. 61–93. Available online: https://link.springer.com/chapter/10.1007/978-3-319-66544-3_4 (accessed on 8 September 2021).
- Varga, T.; Hixson, K.K.; Ahkami, A.H.; Sher, A.W.; Barnes, M.E.; Chu, R.K.; Battu, A.K.; Nicora, C.D.; Winkler, T.E.; Reno, L.R.; et al. Endophyte-promoted phosphorus solubilization in Populus. Front. Plant. Sci. 2020, 11, 1585. [Google Scholar] [CrossRef]
- Whitelaw, M.A.; Harden, T.J.; Helyar, K.R. Phosphate solubilisation in solution culture by the soil fungus Penicillium radicum. Soil. Biol. Biochem. 1999, 31, 655–665. [Google Scholar] [CrossRef]
- Vyas, P.; Gulati, A. Organic acid production in vitro and plant growth promotion in maize under controlled environment by phosphate-solubilizing fluorescent Pseudomonas. BMC Microbiol. 2009, 9, 1–15. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rodriguez, H.; Gonzalez, T.; Goire, I.; Bashan, Y. Gluconic acid production and phosphate solubilization by the plant growth-promoting bacterium Azospirillum spp. Naturwissenschaften 2004, 91, 552–555. [Google Scholar] [CrossRef] [PubMed]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Strains | Species | Accession Number | Tissues | Plants |
|---|---|---|---|---|
| IALR611 | Pantoea vagans | MZ519966 | Shoot | Taraxacum officinale |
| IALR632 | Pseudomonas psychrotolerans | MZ519967 | Leaf | Sorghum halepense |
| IALR1033 | Bacillus subtilis | MZ519968 | Leaf | Ambrosia trifida |
| IALR1035 | Bacillus sufensis | MZ519969 | Leaf | Viola sororia |
| IALR1325 | Pantoea agglomerans | MZ519970 | Seed | Festuca arundinacea |
| Phosphate Compounds | Pantoea vagans IALR611 | Pseudomonas psychrotolerans IALR632 | Bacillus subtilis IALR1033 | Bacillus safensis IALR1035 | Pantoea agglomerans IALR1325 |
|---|---|---|---|---|---|
| Ca3(PO4)2 | 284.5 ± 10.7 1 | 289.3 ± 23.7 | 349.7 ± 23.1 | 274.6 ± 15.2 | 372.8 ± 11.9 |
| Ca5(PO4)3OH | 118.2 ± 21.7 | 126.0 ± 5.5 | 105.2 ± 16.9 | 118.0 ± 14.4 | 146.2 ± 13.1 |
| AlPO4 | 4.6 ± 0.2 | 1.3 ± 0.6 | 6.0 ± 0.8 | 4.3 ± 1.3 | 9.1 ± 0.3 |
| FePO4 | 7.8 ± 0.1 | 6.5 ± 0.4 | 8.3 ± 0.1 | 8.1 ± 0.1 | 9.2 ± 0.4 |
| Enzyme Specific Activities | Pantoea vagans IALR611 | Pseudomonas psychrotolerans IALR632 | Bacillus subtilis IALR1033 | Bacillus safensis IALR1035 | Pantoea agglomerans IALR1325 |
|---|---|---|---|---|---|
| Acidphosphatase | 152.0 ± 7.1 | 721.9 ± 8.8 | 195.7 ± 3.9 | 198.8 ± 14.4 | 369.4 ± 16.3 |
| Phytase | 139.6 ± 21.9 | 64.7 ± 18.2 | 113.1 ± 27.4 | 211.3 ± 45.7 | 138.2 ± 29.1 |
| Plant Growth Promoting Traits | Pantoea vagans IALR611 | Pseudomonas psychrotolerans IALR632 | Bacillus subtilis IALR1033 | Bacillus safensis IALR1035 | Pantoea agglomerans IALR1325 |
|---|---|---|---|---|---|
| Auxins (µg/mL) | 106.3 ± 10.5 | 13.4 ± 1.8 | 1.9 ± 0.6 | 0 | 87.3 ± 6.3 |
| N Fixation | + | + | − | − | + |
| ACC deaminase | ++ | ++ | + | − | − |
| Siderophore (%) | 89 ± 1.6 | 97 ± 0.5 | 58 ± 2.2 | 50 ± 9.9 | 66 ± 5.5 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Mei, C.; Chretien, R.L.; Amaradasa, B.S.; He, Y.; Turner, A.; Lowman, S. Characterization of Phosphate Solubilizing Bacterial Endophytes and Plant Growth Promotion In Vitro and in Greenhouse. Microorganisms 2021, 9, 1935. https://doi.org/10.3390/microorganisms9091935
Mei C, Chretien RL, Amaradasa BS, He Y, Turner A, Lowman S. Characterization of Phosphate Solubilizing Bacterial Endophytes and Plant Growth Promotion In Vitro and in Greenhouse. Microorganisms. 2021; 9(9):1935. https://doi.org/10.3390/microorganisms9091935
Chicago/Turabian StyleMei, Chuansheng, Robert L. Chretien, B. Sajeewa Amaradasa, Yimeng He, Amy Turner, and Scott Lowman. 2021. "Characterization of Phosphate Solubilizing Bacterial Endophytes and Plant Growth Promotion In Vitro and in Greenhouse" Microorganisms 9, no. 9: 1935. https://doi.org/10.3390/microorganisms9091935