
Infection Heterogeneity and Microbiota Differences in Chicks Infected by Salmonella enteritidis

 , , ,
, , ,
Abstract
:1. Introduction
2. Materials and Methods
2.1. Ethical Statement
2.2. Animal Management
2.3. Challenge with S. enteritidis
2.4. Sample Collection
2.5. Salmonella Load Measurement and Sample Grouping
2.6. Liver and Spleen Histopathology and Intestinal Morphology Determination
2.7. RNA Isolation and Quantitative Real-Time PCR
2.8. DNA Extraction and Sequencing Library Construction
2.9. Quality Filtering and Sequence Analysis
2.10. Statistical Analysis
3. Results
3.1. Body Weight, Tissue Index, and Salmonella Loads of S. enteritidis-Susceptible and -Resistant Chicks
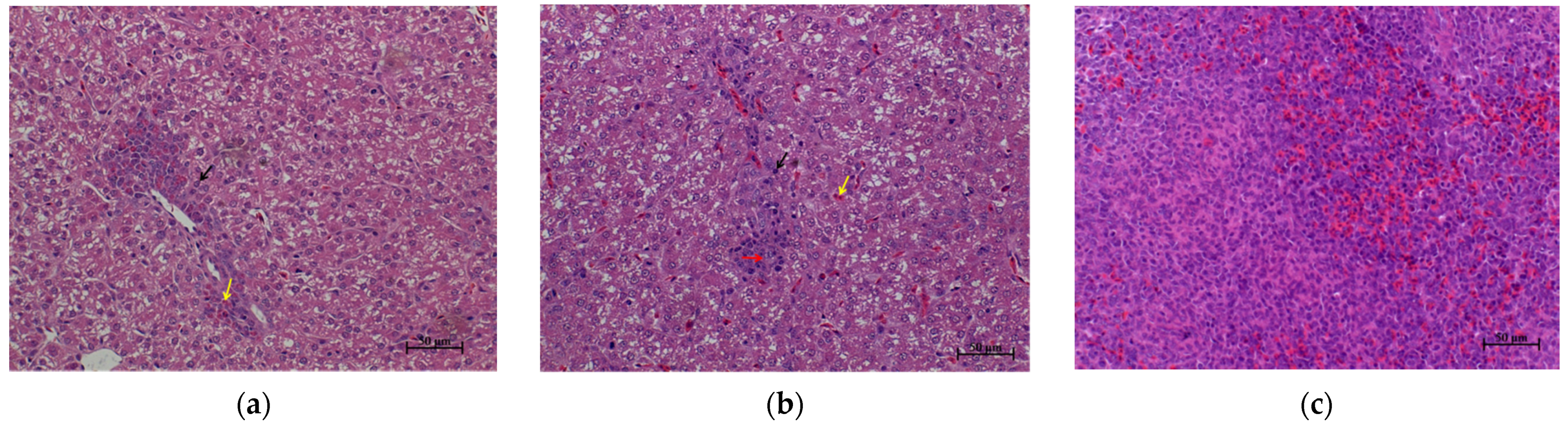
3.2. Liver and Spleen Histopathology of S. enteritidis-Susceptible and -Resistant Chicks
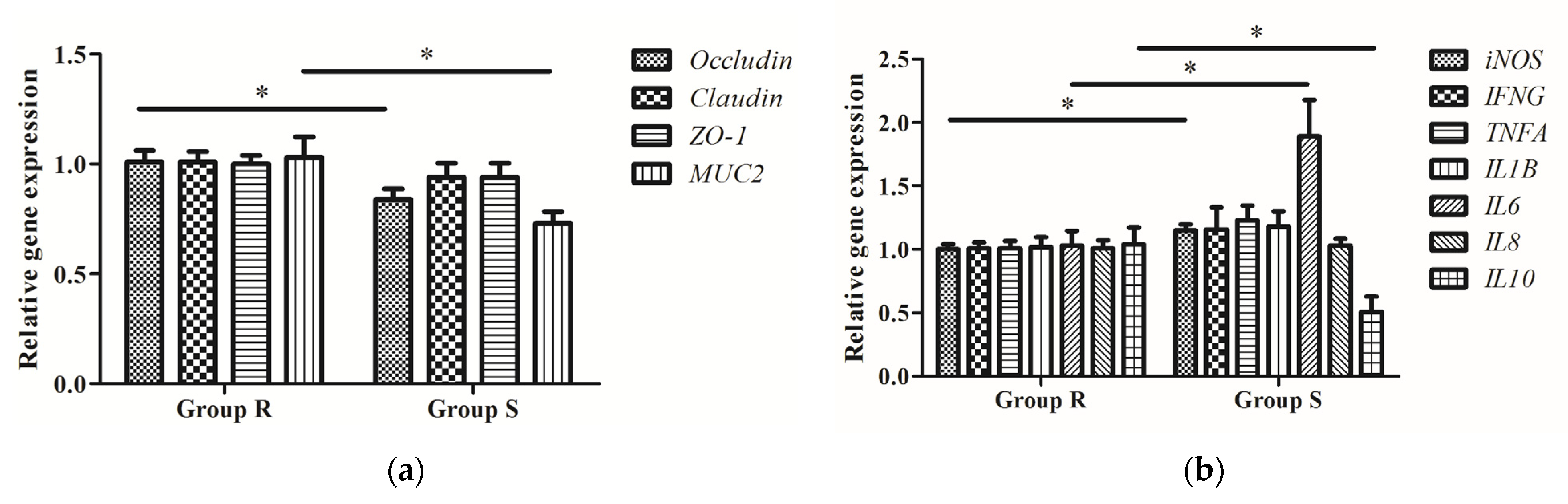
3.3. Intestinal Morphology and Barrier Function of S. enteritidis-Susceptible and -Resistant Chicks
3.4. Expression of Inflammatory Cytokine-Related Genes in S. enteritidis-Susceptible and -Resistant Chicks
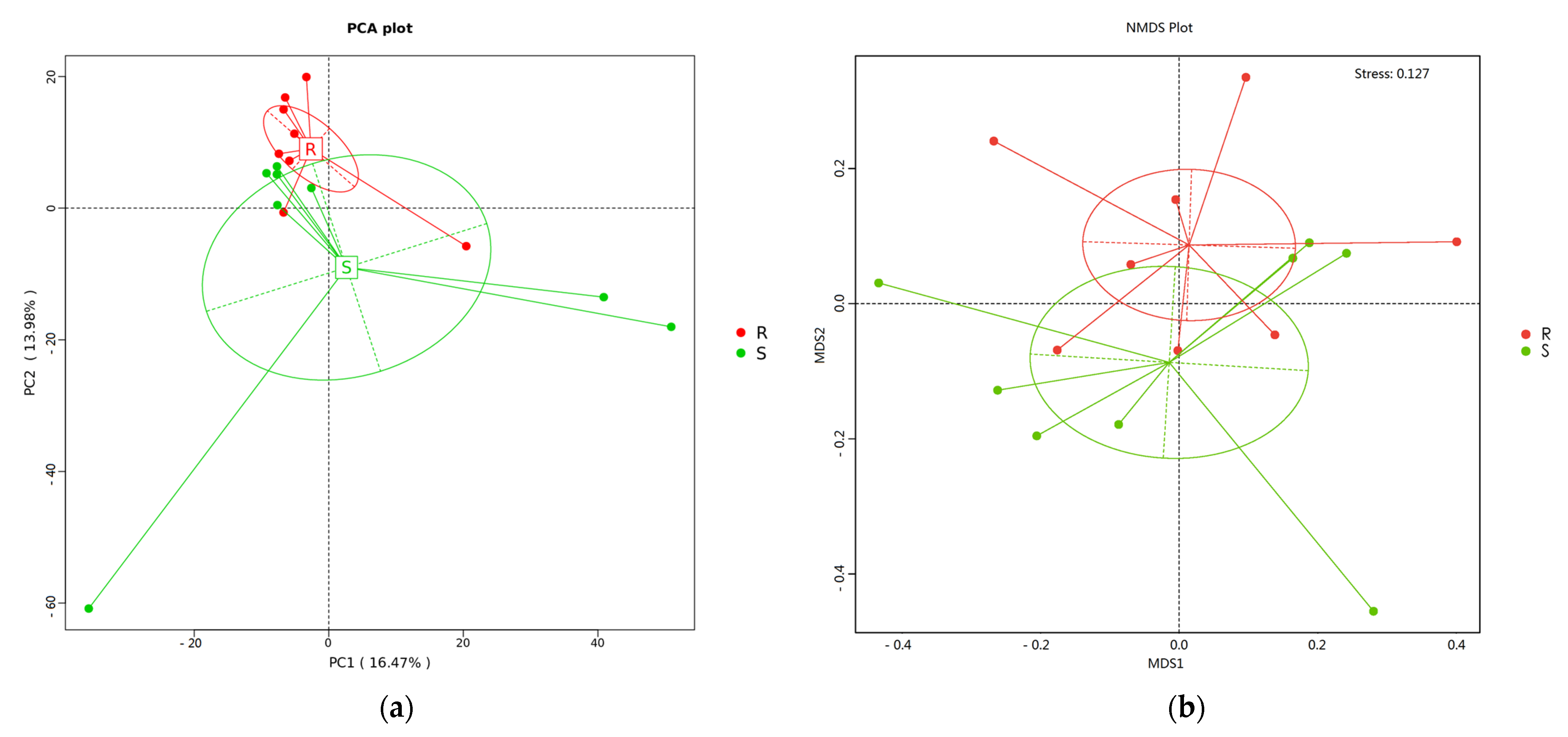
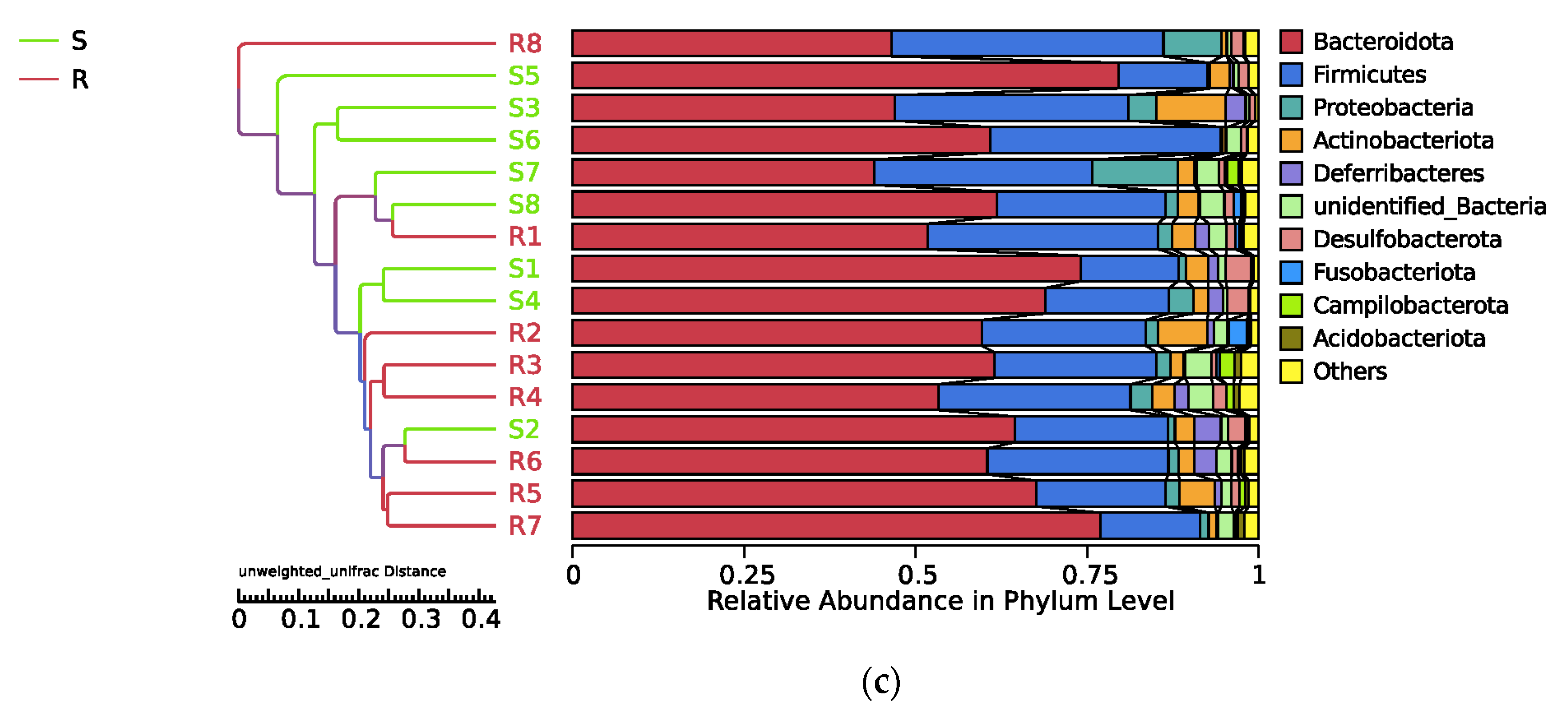
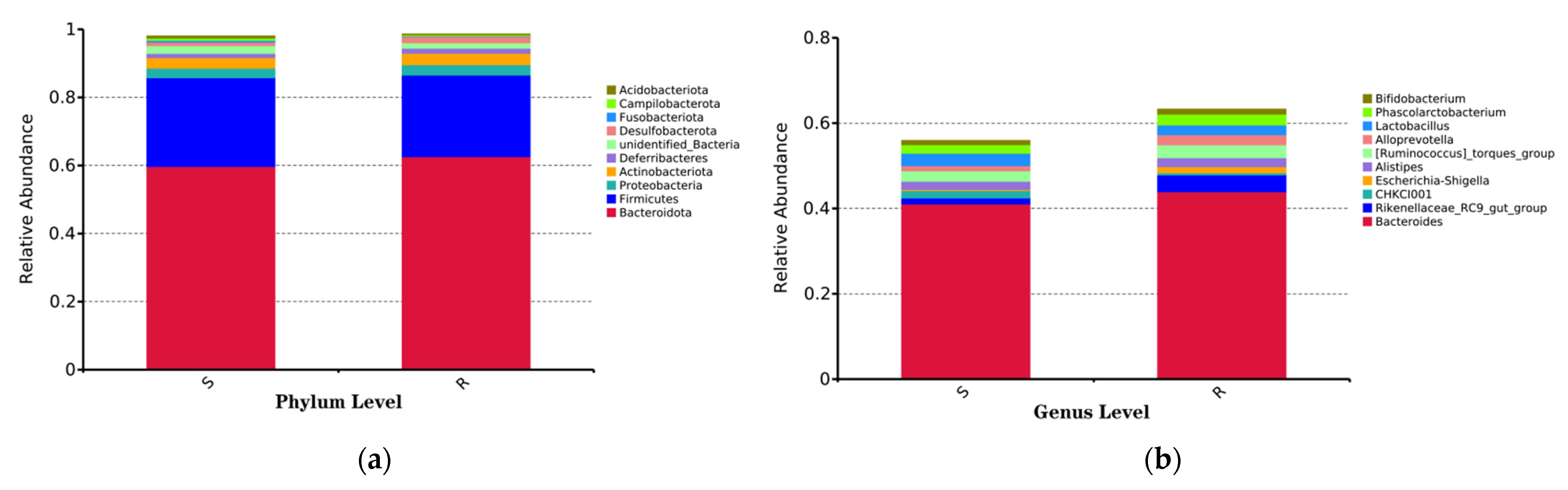
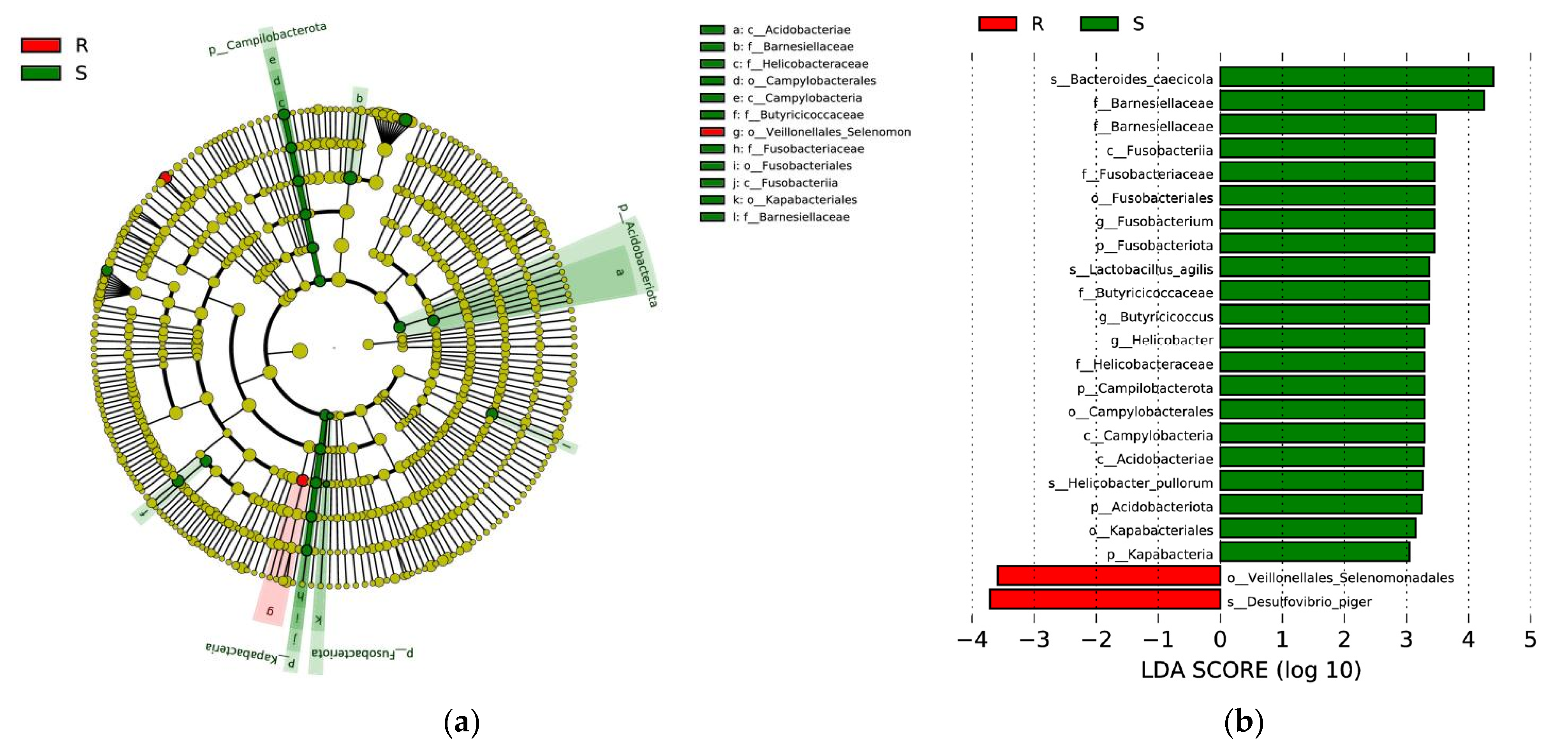
3.5. Composition and Diversity of Cecal Microorganisms in S. enteritidis-Susceptible and -Resistant Chicks
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Shi, S.; Wu, S.; Shen, Y.; Zhang, S.; Xiao, Y.; He, X.; Gong, J.; Farnell, Y.; Tang, Y.; Huang, Y.; et al. Iron oxide nanozyme suppresses intracellular Salmonella enteritidis growth and alleviates infection in vivo. Theranostics 2018, 8, 6149–6162. [Google Scholar] [CrossRef]
- WHO. Salmonella (Non-Typhoidal). 2018. Available online: http://www.who.int/mediacentre/factsheets/fs139/en/ (accessed on 20 February 2018).
- Jiang, M.; Zhu, F.; Yang, C.; Deng, Y.; Kwan, P.S.L.; Li, Y.; Lin, Y.; Qiu, Y.; Shi, X.; Chen, H.; et al. Whole-genome analysis of Salmonella enterica serovar enteritidis isolates in outbreak linked to online food delivery, Shenzhen, China, 2018. Emerg. Infect. Dis. 2020, 26, 789–792. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Smith, A.M.; Tau, N.P.; Ngomane, H.M.; Sekwadi, P.; Ramalwa, N.; Moodley, K.; Govind, C.; Khan, S.; Archary, M.; Thomas, J. Whole-genome sequencing to investigate two concurrent outbreaks of Salmonella enteritidis in South Africa, 2018. J. Med. Microbiol. 2020, 69, 1303–1307. [Google Scholar] [CrossRef]
- Sher, A.A.; Mustafa, B.E.; Grady, S.C.; Gardiner, J.C.; Saeed, A.M. Outbreaks of foodborne Salmonella enteritidis in the United States between 1990 and 2015: An analysis of epidemiological and spatial-temporal trends. Int. J. Infect. Dis. 2021, 105, 54–61. [Google Scholar] [CrossRef]
- Shah, D.H.; Paul, N.C.; Sischo, W.C.; Crespo, R.; Guard, J. Population dynamics and antimicrobial resistance of the most prevalent poultry-associated Salmonella serotypes. Poult. Sci. 2017, 96, 687–702. [Google Scholar] [CrossRef]
- Steed, A.L.; Christophi, G.P.; Kaiko, G.E.; Sun, L.; Goodwin, V.M.; Jain, U.; Esaulova, E.; Artyomov, M.N.; Morales, D.J.; Holtzman, M.J.; et al. The microbial metabolite desaminotyrosine protects from influenza through type I interferon. Science 2017, 357, 498–502. [Google Scholar] [CrossRef] [Green Version]
- Sanchez, K.K.; Chen, G.Y.; Schieber, A.M.P.; Redford, S.E.; Shokhirev, M.N.; Leblanc, M.; Lee, Y.M.; Ayres, J.S. Cooperative metabolic adaptations in the host can favor asymptomatic infection and select for attenuated virulence in an enteric pathogen. Cell 2018, 175, 146–158. [Google Scholar] [CrossRef] [Green Version]
- Velge, P.; Cloeckaert, A.; Barrow, P. Emergence of Salmonella epidemics: The problems related to Salmonella enterica serotype enteritidis and multiple antibiotic resistance in other major serotypes. Vet. Res. 2005, 36, 267–288. [Google Scholar] [CrossRef] [Green Version]
- Calenge, F.; Kaiser, P.; Vignal, A.; Beaumont, C. Genetic control of resistance to salmonellosis and to Salmonella carrier-state in fowl: A review. Genet. Sel. Evol. 2010, 42, 11. [Google Scholar] [CrossRef] [Green Version]
- Chaussé, A.M.; Grépinet, O.; Bottreau, E.; Vern, L.Y.; Menanteau, P.; Trotereau, J.; Robert, V.; Wu, Z.; Kerboeuf, D.; Beaumont, C.; et al. Expression of Toll-like receptor 4 and downstream effectors in selected cecal cell subpopulations of chicks resistant or susceptible to Salmonella carrier state. Infect. Immun. 2011, 79, 3445–3454. [Google Scholar] [CrossRef] [Green Version]
- Lawley, T.D.; Bouley, D.M.; Hoy, Y.E.; Gerke, C.; Relman, D.A.; Monack, D.M. Host transmission of Salmonella enterica serovar Typhimurium is controlled by virulence factors and indigenous intestinal microbiota. Infect. Immun. 2008, 76, 403–416. [Google Scholar] [CrossRef] [Green Version]
- Menanteau, P.; Kempf, F.; Trotereau, J.; Virlogeux-Payant, I.; Gitton, E.; Dalifard, J.; Gabriel, I.; Rychlik, I.; Velge, P. Role of systemic infection, cross contaminations and super-shedders in Salmonella carrier state in chicken. Environ. Microbiol. 2018, 20, 3246–3260. [Google Scholar] [CrossRef]
- Brown, R.L.; Clarke, T.B. The regulation of host defences to infection by the microbiota. Immunology 2017, 150, 1–6. [Google Scholar] [CrossRef] [Green Version]
- Flemer, B.; Warren, R.D.; Barrett, M.P.; Cisek, K.; Das, A.; Jeffery, I.B.; Hurley, E.; O’Riordain, M.; Shanahan, F.; O’Toole, P.W. The oral microbiota in colorectal cancer is distinctive and predictive. Gut 2018, 67, 1454–1463. [Google Scholar] [CrossRef] [Green Version]
- Masetti, G.; Moshkelgosha, S.; Köhling, H.L.; Covelli, D.; Banga, J.P.; Berchner-Pfannschmidt, U.; Horstmann, M.; Diaz-Cano, S.; Goertz, G.E.; Plummer, S.; et al. Gut microbiota in experimental murine model of Graves’ orbitopathy established in different environments may modulate clinical presentation of disease. Microbiome 2018, 6, 97. [Google Scholar] [CrossRef] [Green Version]
- Jie, Z.; Xia, H.; Zhong, S.L.; Feng, Q.; Li, S.; Liang, S.; Zhong, H.; Liu, Z.; Gao, Y.; Zhao, H.; et al. The gut microbiome in atherosclerotic cardiovascular disease. Nat. Commun. 2017, 8, 845. [Google Scholar] [CrossRef] [Green Version]
- Zhao, L.; Zhang, F.; Ding, X.; Wu, G.; Lam, Y.Y.; Wang, X.; Fu, H.; Xue, X.; Lu, C.; Ma, J.; et al. Gut bacteria selectively promoted by dietary fibers alleviate type 2 diabetes. Science 2018, 359, 1151–1156. [Google Scholar] [CrossRef] [Green Version]
- Cho, I.; Blaser, M.J. The human microbiome: At the interface of health and disease. Nat. Rev. Genet. 2012, 13, 260–270. [Google Scholar] [CrossRef] [Green Version]
- Juricova, H.; Videnska, P.; Lukac, M.; Faldynova, M.; Babak, V.; Havlickova, H.; Sisak, F.; Rychlik, I. Influence of Salmonella enterica serovar enteritidis infection on the development of the cecum microbiota in newly hatched chicks. Appl. Environ. Microbiol. 2013, 79, 745–747. [Google Scholar] [CrossRef] [Green Version]
- Borewicz, K.A.; Kim, H.B.; Singer, R.S.; Gebhart, C.J.; Sreevatsan, S.; Johnson, T.; Isaacson, R.E. Changes in the porcine intestinal microbiome in response to infection with Salmonella enterica and Lawsonia intracellularis. PLoS ONE 2015, 10, e0139106. [Google Scholar] [CrossRef]
- Pollock, J.; Hutchings, M.R.; Hutchings, K.E.K.; Gally, D.L.; Houdijk, J.G.M. Changes in the ileal, but not fecal, microbiome in response to increased dietary protein level and enterotoxigenic Escherichia coli exposure in pigs. Appl. Environ. Microbiol. 2019, 85, e01252-19. [Google Scholar] [CrossRef] [Green Version]
- Pickard, J.M.; Zeng, M.Y.; Caruso, R.; Núñez, G. Gut microbiota: Role in pathogen colonization, immune responses, and inflammatory disease. Immunol. Rev. 2017, 279, 70–89. [Google Scholar] [CrossRef]
- Litvak, Y.; Mon, K.K.Z.; Nguyen, H.; Chanthavixay, G.; Liou, M.; Velazquez, E.M.; Kutter, L.; Alcantara, M.A.; Byndloss, M.X.; Tiffany, C.R.; et al. Commensal Enterobacteriaceae protect against Salmonella colonization through oxygen competition. Cell Host. Microbe 2019, 25, 128–139. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sabry, M.A.; Abdel-Moein, K.A.; Abdel-Kader, F.; Hamza, E. Extended-spectrum β-lactamase-producing Salmonella serovars among healthy and diseased chickens and their public health implication. J. Glob. Antimicrob. Resist. 2020, 22, 742–748. [Google Scholar] [CrossRef] [PubMed]
- Zhang, S.; Shen, Y.; Wu, S.; Xiao, Y.; He, Q.; Shi, S. The dietary combination of essential oils and organic acids reduces Salmonella enteritidis in challenged chicks. Poult. Sci. 2019, 98, 6349–6355. [Google Scholar] [CrossRef] [PubMed]
- Shen, Y.; Xiao, Y.; Zhang, S.; Wu, S.; Gao, L.; Shi, S. Fe3O4 nanoparticles attenuated Salmonella infection in chicken liver through reactive oxygen and autophagy via PI3K/Akt/mTOR signaling. Front. Physiol. 2020, 10, 1580. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hu, Y.; Wang, L.; Shao, D.; Wang, Q.; Wu, Y.; Han, Y.; Shi, S. Selectived and reshaped early dominant microbial community in the cecum with similar proportions and better homogenization and species diversity due to organic acids as AGP alternatives mediate their effects on broilers growth. Front. Microbiol. 2020, 10, 2948. [Google Scholar] [CrossRef] [Green Version]
- Sousa, B.F.; Silva, A.F.B.D.; Lima-Filho, J.V.; Agostinho, A.G.; Oliveira, D.N.; De Alencar, N.M.N.; De Freitas, C.D.T.; Ramos, M.V. Latex proteins downregulate inflammation and restores blood-coagulation homeostasis in acute Salmonella infection. Mem. Inst. Oswaldo Cruz 2020, 115, e200458. [Google Scholar] [CrossRef]
- Guo, L.D.; Hyde, K.D.; Liew, E.C.Y. Identification of endophytic fungi from Livistona chinensis based on morphology and rDNA sequences. New Phytol. 2000, 147, 617–630. [Google Scholar] [CrossRef]
- Yu, X.; Wu, X.; Qiu, L.; Wang, D.; Gan, M.; Chen, X.; Wei, H.; Xu, F. Analysis of the intestinal microbial community structure of healthy and long-living elderly residents in Gaotian Village of Liuyang City. Appl. Microbiol. Biotechnol. 2015, 99, 9085–9095. [Google Scholar] [CrossRef]
- Caporaso, J.G.; Lauber, C.L.; Walters, W.A.; Berg-Lyons, D.; Lozupone, C.A.; Turnbaugh, P.J.; Fierer, N.; Knight, R. Global patterns of 16S rRNA diversity at a depth of millions of sequences per sample. Proc. Natl. Acad. Sci. USA 2011, 108 (Suppl. 1), 4516–4522. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gao, Y.; Wang, C.; Zhang, W.; Di, P.; Yi, N.; Chen, C. Vertical and horizontal assemblage patterns of bacterial communities in a eutrophic river receiving domestic wastewater in southeast China. Environ. Pollut. 2017, 230, 469–478. [Google Scholar] [CrossRef] [PubMed]
- Magoč, T.; Salzberg, S.L. FLASH: Fast length adjustment of short reads to improve genome assemblies. Bioinformatics 2011, 27, 2957–2963. [Google Scholar] [CrossRef] [PubMed]
- Caporaso, J.G.; Kuczynski, J.; Stombaugh, J.; Bittinger, K.; Bushman, F.D.; Costello, E.K.; Fierer, N.; Peña, A.G.; Goodrich, J.K.; Gordon, J.I.; et al. QIIME allows analysis of high-throughput community sequencing data. Nat. Methods 2010, 7, 335–336. [Google Scholar] [CrossRef] [Green Version]
- Haas, B.J.; Gevers, D.; Earl, A.M.; Feldgarden, M.; Ward, D.V.; Giannoukos, G.; Ciulla, D.; Tabbaa, D.; Highlander, S.K.; Sodergren, E.; et al. Chimeric 16S rRNA sequence formation and detection in Sanger and 454-pyrosequenced PCR amplicons. Genome Res. 2011, 21, 494–504. [Google Scholar] [CrossRef] [Green Version]
- Wang, Q.; Garrity, G.M.; Tiedje, J.M.; Cole, J.R. Naive Bayesian classifier for rapid assignment of rRNA sequences into the new bacterial taxonomy. Appl. Environ. Microbiol. 2007, 73, 5261–5267. [Google Scholar] [CrossRef] [Green Version]
- Zhang, L.; Wu, W.; Lee, Y.K.; Xie, J.; Zhang, H. Spatial heterogeneity and co-occurrence of mucosal and luminal microbiome across swine intestinal tract. Front. Microbiol. 2018, 9, 48. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lozupone, C.; Knight, R. UniFrac: A new phylogenetic method for comparing microbial communities. Appl. Environ. Microbiol. 2005, 71, 8228–8235. [Google Scholar] [CrossRef] [Green Version]
- Zhang, B.; Li, G.; Shahid, M.S.; Gan, L.; Fan, H.; Lv, Z.; Yan, S.; Guo, Y. Dietary l-arginine supplementation ameliorates inflammatory response and alters gut microbiota composition in broiler chickens infected with Salmonella enterica serovar Typhimurium. Poult. Sci. 2020, 99, 1862–1874. [Google Scholar] [CrossRef]
- Shah, D.H.; Zhou, X.; Kim, H.Y.; Call, D.R.; Guard, J. Transposon mutagenesis of Salmonella enterica serovar enteritidis identifies genes that contribute to invasiveness in human and chicken cells and survival in egg albumen. Infect. Immun. 2012, 80, 4203–4215. [Google Scholar] [CrossRef] [Green Version]
- Wang, C.L.; Fan, Y.C.; Wang, C.; Tsai, H.J.; Chou, C.H. The impact of Salmonella enteritidis on lipid accumulation in chicken hepatocytes. Avian. Pathol. 2016, 45, 450–457. [Google Scholar] [CrossRef] [Green Version]
- König, J.; Wells, J.; Cani, P.D.; García-Ródenas, C.L.; MacDonald, T.; Mercenier, A.; Whyte, J.; Troost, F.; Brummer, R.J. Human intestinal barrier function in health and disease. Clin. Transl. Gastroenterol. 2016, 7, e196. [Google Scholar] [CrossRef] [PubMed]
- Nathan, C.; Shiloh, M.U. Reactive oxygen and nitrogen intermediates in the relationship between mammalian hosts and microbial pathogens. Proc. Natl. Acad. Sci. USA 2000, 97, 8841–8848. [Google Scholar] [CrossRef] [Green Version]
- Fu, Y.; Galán, J.E. A Salmonella protein antagonizes Rac-1 and Cdc42 to mediate host-cell recovery after bacterial invasion. Nature 1999, 401, 293–297. [Google Scholar] [CrossRef] [PubMed]
- Khan, S.; Chousalkar, K.K. Salmonella Typhimurium infection disrupts but continuous feeding of Bacillus based probiotic restores gut microbiota in infected hens. J. Anim. Sci. Biotechnol. 2020, 11, 29. [Google Scholar] [CrossRef] [Green Version]
- Pourabedin, M.; Zhao, X. Prebiotics and gut microbiota in chickens. FEMS. Microbiol. Lett. 2015, 362, fnv122. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Li, B.; Zhang, X.; Guo, F.; Wu, W.; Zhang, T. Characterization of tetracycline resistant bacterial community in saline activated sludge using batch stress incubation with high-throughput sequencing analysis. Water. Res. 2013, 47, 4207–4216. [Google Scholar] [CrossRef]
- De Witte, C.; Demeyere, K.; De Bruyckere, S.; Taminiau, B.; Daube, G.; Ducatelle, R.; Meyer, E.; Haesebrouck, F. Characterization of the non-glandular gastric region microbiota in Helicobacter suis-infected versus non-infected pigs identifies a potential role for Fusobacterium gastrosuis in gastric ulceration. Vet. Res. 2019, 50, 39. [Google Scholar] [CrossRef] [Green Version]
- Mármol, I.; Sánchez-de-Diego, C.; Pradilla Dieste, A.; Cerrada, E.; Rodriguez Yoldi, M.J. Colorectal carcinoma: A general overview and future perspectives in colorectal cancer. Int. J. Mol. Sci. 2017, 18, 197. [Google Scholar] [CrossRef] [Green Version]
- Rubinstein, M.R.; Baik, J.E.; Lagana, S.M.; Han, R.P.; Raab, W.J.; Sahoo, D.; Dalerba, P.; Wang, T.C.; Han, Y.W. Fusobacterium nucleatum promotes colorectal cancer by inducing Wnt/β-catenin modulator Annexin A1. EMBO Rep. 2019, 20, e47638. [Google Scholar] [CrossRef]
- Kollarcikova, M.; Kubasova, T.; Karasova, D.; Crhanova, M.; Cejkova, D.; Sisak, F.; Rychlik, I. Use of 16S rRNA gene sequencing for prediction of new opportunistic pathogens in chicken ileal and cecal microbiota. Poult. Sci. 2019, 98, 2347–2353. [Google Scholar] [CrossRef] [PubMed]
- Javed, S.; Gul, F.; Javed, K.; Bokhari, H. Helicobacter pullorum: An emerging zoonotic pathogen. Front. Microbiol. 2017, 8, 604. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Qin, M.; Long, F.; Wu, W.; Yang, D.; Huang, M.; Xiao, C.; Chen, X.; Liu, X.; Zhu, Y.Z. Hydrogen sulfide protects against DSS-induced colitis by inhibiting NLRP3 inflammasome. Free Radic. Biol. Med. 2019, 137, 99–109. [Google Scholar] [CrossRef] [PubMed]
- Fiorucci, S.; Orlandi, S.; Mencarelli, A.; Caliendo, G.; Santagada, V.; Distrutti, E.; Santucci, L.; Cirino, G.; Wallace, J.L. Enhanced activity of a hydrogen sulphide-releasing derivative of mesalamine (ATB-429) in a mouse model of colitis. Br. J. Pharmacol. 2007, 150, 996–1002. [Google Scholar] [CrossRef] [PubMed] [Green Version]









{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Ingredient | % |
|---|---|
| Corn | 55.24 |
| Soybean meal (46%) | 36.92 |
| Soybean oil | 3.50 |
| Limestone | 1.12 |
| Calcium hydrogen phosphate | 2.10 |
| Methionine | 0.28 |
| Lysine (98%) | 0.22 |
| NaCl | 0.30 |
| Vitamin premix 1 | 0.03 |
| Mineral premix 2 | 0.20 |
| Choline chloride (70%) | 0.09 |
| Total | 100.00 |
| Nutrient levels (%) 3 | |
| ME (kcal/kg) | 2950 |
| CP | 21.00 |
| Ca | 1.00 |
| Total phosphorus | 0.67 |
| Nonphytate phosphorous | 0.45 |
| Digestible Lys | 1.20 |
| Digestible sulfur-containing amino acid | 0.85 |
| Digestible Thr | 0.66 |
| Digestible Trp | 0.22 |
| Gene | GenBank Accession No. | Primer Orientation | Primer Sequence (5′→3′) | Product Size (bp) |
|---|---|---|---|---|
| GAPDH | NM_204305.1 | Forward | GCCCAGAACATCATCCCA | 137 |
| Reverse | CGGCAGGTCAGGTCAACA | |||
| iNOS | NM_204961.1 | Forward | CCTGGAGGTCCTGGAAGAGT | 82 |
| Reverse | CCTGGGTTTCAGAAGTGGC | |||
| IFNG | NM_205149.1 | Forward | CAAGCTCCCGATGAACGACTT | 162 |
| Reverse | AGTTGAGCACAGGAGGTCAT | |||
| TNFA | NM_204267.1 | Forward | CAGGACAGCCTATGCCAACAAG | 114 |
| Reverse | GGTTACAGGAAGGGCAACTCATC | |||
| IL1B | NM_204524.1 | Forward | CCGAGGAGCAGGGACTTT | 133 |
| Reverse | AGGACTGTGAGCGGGTGT | |||
| IL6 | NM_204628.1 | Forward | TTTATGGAGAAGACCGTGAGG | 106 |
| Reverse | TGTGGCAGATTGGTAACAGAG | |||
| IL8 | NM_205498.1 | Forward | ATGAACGGCAAGCTTGGAGCTG | 233 |
| Reverse | TCCAAGCACACCTCTCTTCCATCC | |||
| IL10 | NM_001004414.2 | Forward | GCTGAGGGTGAAGTTTGAG | 272 |
| Reverse | CAGGTGAAGAAGCGGTGA | |||
| occludin | NM_205128.1 | Forward | TCATCGCCTCCATCGTCTAC | 141 |
| Reverse | TCTTACTGCGCGTCTTCTGG | |||
| claudin | NM_001013611 | Forward | CTGATTGCTTCCAACCAG | 140 |
| Reverse | CAGGTCAAACAGAGGTACAAG | |||
| ZO-1 | XM_413773 | Forward | CTTCAGGTGTTTCTCTTCCTCCTC | 131 |
| Reverse | CTGTGGTTTCATGGCTGGATC | |||
| MUC2 | NM_001318434.1 | Forward | GTGAAGACCCTGATGAAA | 219 |
| Reverse | GTGAACACTGGCGAGAAT |
| Items | Group S 3 | Group R 3 | p-Value |
|---|---|---|---|
| BW (g) | 229.40 ± 7.47 | 230.88 ± 5.47 | 0.875 |
| Liver Index (%) | 0.043 ± 0.001 a | 0.038 ± 0.001 b | 0.006 |
| Spleen Index (%) | 0.024 ± 0.002 | 0.020 ± 0.001 | 0.158 |
| Liver Salmonella loads (log10CFU/g) | 2.750 ± 0.405 a | 1.152 ± 0.435 b | 0.018 |
| Spleen Salmonella loads (log10CFU/g) | 4.784 ± 0.100 a | 2.491 ± 0.055 b | <0.001 |
| Items | Group S 2 | Group R 2 | p-Value |
|---|---|---|---|
| Villus height (μm) | 1084.62 ± 35.20 | 1125.93 ± 90.23 | 0.683 |
| Crypt depth (μm) | 149.56 ± 7.48 | 131.55 ± 16.28 | 0.348 |
| Ratio of villus height-to-crypt depth | 7.32 ± 0.34 | 8.94 ± 0.75 | 0.090 |
| Muscle thickness (μm) | 117.86 ± 7.09 | 118.02 ± 14.37 | 0.992 |
| Taxa | Group S (%) 2 | Group R (%) 2 | p-Value |
|---|---|---|---|
| Fusobacterium | 0.5168 ± 0.9028 a | 0.1782 ± 0.3837 b | 0.043 |
| Helicobacter | 0.6998 ± 0.6460 a | 0.3238 ± 0.5453 b | 0.028 |
| Butyricicoccus | 0.8835 ± 0.5888 a | 0.3974 ± 0.4655 b | 0.050 |
| Bryobacter | 0.1440 ± 0.1037 a | 0.0426 ± 0.0371 b | 0.034 |
| Acidothermus | 0.1222 ± 0.0886 a | 0.0224 ± 0.0251 b | 0.026 |
| unidentified_Chloroplast | 0.0637 ± 0.0814 a | 0.0256 ± 0.0469 b | 0.039 |
| NK4A214_group | 0.0560 ± 0.0504 a | 0.0214 ± 0.0243 b | 0.040 |
| Marvinbryantia | 0.0685 ± 0.0414 a | 0.0278 ± 0.0272 b | 0.046 |
| Burkholderia-Caballeronia-Paraburkholderia | 0.0525 ± 0.0394 a | 0.0112 ± 0.0166 b | 0.024 |
| Granulicella | 0.0458 ± 0.0352 a | 0.0096 ± 0.0130 b | 0.035 |
| Puia | 0.0218 ± 0.0228 a | 0.0035 ± 0.0056 b | 0.048 |
| Oribacterium | 0.0013 ± 0.0036 b | 0.0102 ± 0.0162 a | 0.028 |
| unidentified_IMCC26256 | 0.0182 ± 0.0134 a | 0.0042 ± 0.0067 b | 0.037 |
| Actinospica | 0.0102 ± 0.0092 a | 0.0013 ± 0.0024 b | 0.021 |
| Herbinix | 0 b | 0.0064 ± 0.0079 a | 0.013 |
| Dyella | 0.0074 ± 0.0079 a | 0.0010 ± 0.0019 b | 0.037 |
| Nocardia | 0.0061 ± 0.0056 a | 0.0006 ± 0.0018 b | 0.017 |
| Papillibacter | 0.0003 ± 0.0009 b | 0.0041 ± 0.0034 a | 0.034 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Wu, S.; Cong, G.; Zhang, Q.; Yao, H.; Wang, Z.; Kang, K.; He, X.; Shi, S. Infection Heterogeneity and Microbiota Differences in Chicks Infected by Salmonella enteritidis. Microorganisms 2021, 9, 1705. https://doi.org/10.3390/microorganisms9081705
Wu S, Cong G, Zhang Q, Yao H, Wang Z, Kang K, He X, Shi S. Infection Heterogeneity and Microbiota Differences in Chicks Infected by Salmonella enteritidis. Microorganisms. 2021; 9(8):1705. https://doi.org/10.3390/microorganisms9081705
Chicago/Turabian StyleWu, Shu, Guanglei Cong, Qianyun Zhang, Hong Yao, Zhenxin Wang, Kelang Kang, Xi He, and Shourong Shi. 2021. "Infection Heterogeneity and Microbiota Differences in Chicks Infected by Salmonella enteritidis" Microorganisms 9, no. 8: 1705. https://doi.org/10.3390/microorganisms9081705