
Reviewing Interspecies Interactions as a Driving Force Affecting the Community Structure in Lakes via Cyanotoxins

 , , and
, , and 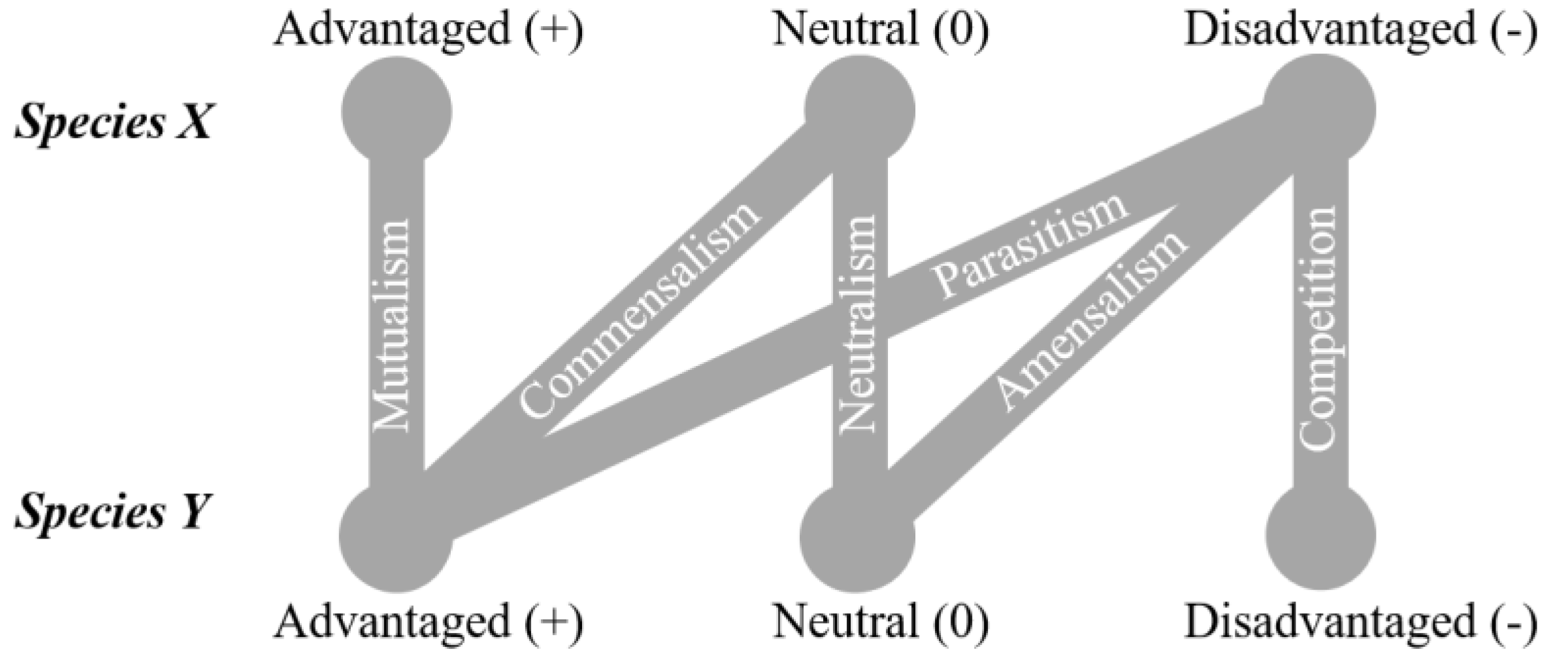{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Cyanotoxins and Interspecies Interplay
2.1. Microcystins (MCs)
2.1.1. MCs and Heterotrophic Bacteria
2.1.2. MCs and Phytoplankton, Including Other Cyanobacteria
2.1.3. MCs and Zooplankton
2.1.4. MCs and Aquatic Macrophytes
2.2. Anatoxins (ATXs)
2.2.1. ATXs and Heterotrophic Bacteria
2.2.2. ATXs and Phytoplankton
2.2.3. ATXs and Zooplankton
2.2.4. ATXs and Aquatic Macrophytes
2.3. Saxitoxins (STXs)
2.3.1. STXs and Heterotrophic Bacteria
2.3.2. STXs and Phytoplankton
2.3.3. STXs and Zooplankton
2.4. Cylindrospermopsin (CYN)
2.4.1. CYN and Heterotrophic Bacteria
2.4.2. CYN and Phytoplankton
2.4.3. CYN and Zooplankton
2.4.4. CYN and Macrophytes
2.5. BMAA
2.5.1. BMAA and Heterotrophic Bacteria
2.5.2. BMAA and Phytoplankton
2.5.3. BMAA and Zooplankton
2.5.4. BMAA and Macrophytes and Macroalgae
3. Concluding Remarks
Author Contributions
Funding
Data Availability Statement
Conflicts of Interest
References
- Van Goethem, M.W.; Makhalanyane, T.P.; Cowan, D.A.; Valverde, A. Cyanobacteria and alphaproteobacteria may facilitate cooperative interactions in niche communities. Front. Microbiol. 2017, 8, 2099. [Google Scholar] [CrossRef] [PubMed]
- Erhard, D. Allelopathy in aquatic environments. In Allelopathy: A Physiological Process with Ecological Implications; Reigosa, M.J., Nuria, P., González, L., Eds.; Springer: Berlin/Heidelberg, Germany, 2006; pp. 433–450. [Google Scholar]
- Protasov, A.; Barinova, S.; Novoselova, T.; Sylaieva, A. The aquatic organisms diversity, community structure, and environmental conditions. Diversity 2019, 11, 190. [Google Scholar] [CrossRef] [Green Version]
- Teittinen, A.; Virta, L.; Li, M.; Wang, J. Factors influencing the biodiversity of three microbial groups within and among islands of the Baltic Sea. FEMS Microbiol. Ecol. 2021, 97, fiab049. [Google Scholar] [CrossRef]
- Yang, J.R.; Yu, X.; Chen, H.; Kuo, Y.-M.; Yang, J. Structural and functional variations of phytoplankton communities in the face of multiple disturbances. J. Environ. Sci. 2021, 100, 287–297. [Google Scholar] [CrossRef]
- Zastepa, A.; Miller, T.R.; Watson, L.C.; Kling, H.; Watson, S.B. Toxins and other bioactive metabolites in deep chlorophyll layers containing the cyanobacteria Planktothrix cf. isothrix in two Georgian Bay Embayments, Lake Huron. Toxins 2021, 13, 445. [Google Scholar] [CrossRef] [PubMed]
- Lawrence, D.; Fiegna, F.; Behrends, V.; Bundy, J.G.; Phillimore, A.B.; Bell, T.; Barraclough, T.G. Species interactions alter evolutionary responses to a novel environment. PLoS Biol. 2012, 10, e1001330. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Odum, E.P. Fundamentals of Ecology, 2nd ed.; Saunders: Philadelphia, PA, USA, 1959; p. 546. [Google Scholar]
- Lang, J.M.; Benbow, M.E. Species interactions and competition. Nat. Educ. Knowl. 2013, 4, 8. [Google Scholar]
- Kordas, R.L.; Dudgeon, S. Dynamics of species interaction strength in space, time and with developmental stage. Proc. R. Soc. B 2011, 278, 1804–1813. [Google Scholar] [CrossRef]
- Zhu, C.-M.; Zhang, J.-Y.; Guan, R.; Hale, L.; Chen, N.; Li, M.; Lu, Z.-H.; Ge, Q.-Y.; Yang, Y.-F.; Zhou, J.-Z. Alternate succession of aggregate-forming cyanobacterial genera correlated with their attached bacteria by co-pathways. Sci. Total Environ. 2019, 688, 867–879. [Google Scholar] [CrossRef]
- Omidi, A.; Esterhuizen-Londt, M.; Pflugmacher, S. Still challenging: The ecological function of the cyanobacterial toxin microcystin—What we know so far. Toxin Rev. 2017, 37, 87–105. [Google Scholar] [CrossRef] [Green Version]
- Sukenik, A.; Quesada, A.; Salmaso, N. Global expansion of toxic and non-toxic cyanobacteria: Effect on ecosystem functioning. Biodivers. Conserv. 2015, 24, 889–908. [Google Scholar] [CrossRef]
- Scholz, S.N.; Esterhuizen-Londt, M.; Pflugmacher, S. Rise of toxic cyanobacterial blooms in temperate freshwater lakes: Causes, correlations and possible countermeasures. Toxicol. Environ. Chem. 2017, 99, 543–577. [Google Scholar] [CrossRef]
- Havens, K.E. Cyanobacteria blooms: Effects on aquatic ecosystems. In Cyanobacterial Harmful Algal Blooms: State of the Science and Research Needs; Hudnell, H.K., Ed.; Springer: New York, NY, USA, 2008; pp. 733–747. [Google Scholar]
- Osman, O.A.; Beier, S.; Grabherr, M.; Bertilsson, S. Interactions of freshwater cyanobacteria with bacterial antagonists. Appl. Environ. Microbiol. 2017, 83, e02634-16. [Google Scholar] [CrossRef] [Green Version]
- Sellner, K.; Olson, M.; Kononen, K. Copepod grazing in a summer cyanobacteria bloom in the Gulf of Finland. Hydrobiologia 1994, 292, 249–254. [Google Scholar] [CrossRef]
- Jiang, X.; Yang, W.; Zhao, S.; Liang, H.; Zhao, Y.; Chen, L.; Li, R. Maternal effects of inducible tolerance against the toxic cyanobacterium Microcystis aeruginosa in the grazer Daphnia carinata. Environ. Pollut. 2013, 178, 142–146. [Google Scholar] [CrossRef] [PubMed]
- Svirčev, Z.; Lalić, D.; Savić, G.B.; Tokodi, N.; Backović, D.D.; Chen, L.; Meriluoto, J.; Codd, G.A. Global geographical and historical overview of cyanotoxin distribution and cyanobacterial poisonings. Arch. Toxicol. 2019, 93, 2429–2481. [Google Scholar] [CrossRef] [PubMed]
- Sommer, U.; Gliwicz, Z.M.; Lampert, W.; Duncan, A. The PEG-model of seasonal succession of planktonic events in fresh waters. Arch. Hydrobiol. 1986, 106, 433–471. [Google Scholar]
- Wang, X.-l.; Lu, Y.-l.; He, G.-z.; Han, J.-y.; Wang, T.-y. Exploration of relationships between phytoplankton biomass and related environmental variables using multivariate statistic analysis in a eutrophic shallow lake: A 5-year study. J. Environ. Sci. 2007, 19, 920–927. [Google Scholar] [CrossRef]
- Wei, J.; Wang, M.; Chen, C.; Wu, H.; Lin, L.; Li, M. Seasonal succession of phytoplankton in two temperate artificial lakes with different water sources. Environ. Sci. Pollut. Res. 2020, 27, 42324–42334. [Google Scholar] [CrossRef] [PubMed]
- Jones, M.R.; Pinto, E.; Torres, M.A.; Dörr, F.; Mazur-Marzec, H.; Szubert, K.; Tartaglione, L.; Dell’Aversano, C.; Miles, C.O.; Beach, D.G. Comprehensive database of secondary metabolites from cyanobacteria. BioRxiv 2020. [Google Scholar] [CrossRef] [Green Version]
- Hernández-Zamora, M.; Santiago-Martínez, E.; Martínez-Jerónimo, F. Toxigenic Microcystis aeruginosa (Cyanobacteria) affects the population growth of two common green microalgae. Evidence of other allelopathic metabolites different to cyanotoxins. J. Phycol. 2021. [Google Scholar] [CrossRef]
- Pawlik-Skowrońska, B.; Toporowska, M.; Mazur-Marzec, H. Effects of secondary metabolites produced by different cyanobacterial populations on the freshwater zooplankters Brachionus calyciflorus and Daphnia pulex. Environ. Sci. Pollut. Res. 2019, 26, 11793–11804. [Google Scholar] [CrossRef] [Green Version]
- Gross, E.M.; Wolk, C.P.; Jüttner, F. Fischerellin, a new allelochemical from the freshwater cyanobacterium Fischerella muscicola. J. Phycol. 1991, 27, 686–692. [Google Scholar] [CrossRef] [Green Version]
- Mason, C.; Edwards, K.; Carlson, R.; Pignatello, J.; Gleason, F.; Wood, J. Isolation of chlorine-containing antibiotic from the freshwater cyanobacterium Scytonema hofmanni. Science 1982, 215, 400–402. [Google Scholar] [CrossRef]
- Song, H.; Lavoie, M.; Fan, X.; Tan, H.; Liu, G.; Xu, P.; Fu, Z.; Paerl, H.W.; Qian, H. Allelopathic interactions of linoleic acid and nitric oxide increase the competitive ability of Microcystis aeruginosa. ISME J. 2017, 11, 1865–1876. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Qian, H.; Xu, J.; Lu, T.; Zhang, Q.; Qu, Q.; Yang, Z.; Pan, X. Responses of unicellular alga Chlorella pyrenoidosa to allelochemical linoleic acid. Sci. Total Environ. 2018, 625, 1415–1422. [Google Scholar] [CrossRef]
- Rzymski, P.; Poniedziałek, B.; Kokociński, M.; Jurczak, T.; Lipski, D.; Wiktorowicz, K. Interspecific allelopathy in cyanobacteria: Cylindrospermopsin and Cylindrospermopsis raciborskii effect on the growth and metabolism of Microcystis aeruginosa. Harmful Algae 2014, 35, 1–8. [Google Scholar] [CrossRef]
- Pflugmacher, S. Possible allelopathic effects of cyanotoxins, with reference to microcystin-LR, in aquatic ecosystems. Environ. Toxicol. 2002, 17, 407–413. [Google Scholar] [CrossRef]
- Chia, M.A.; Jankowiak, J.G.; Kramer, B.J.; Goleski, J.A.; Huang, I.-S.; Zimba, P.V.; do Carmo Bittencourt-Oliveira, M.; Gobler, C.J. Succession and toxicity of Microcystis and Anabaena (Dolichospermum) blooms are controlled by nutrient-dependent allelopathic interactions. Harmful Algae 2018, 74, 67–77. [Google Scholar] [CrossRef] [PubMed]
- Wiegand, C.; Pflugmacher, S. Ecotoxicological effects of selected cyanobacterial secondary metabolites: A short review. Toxicol. Appl. Pharmacol. 2005, 203, 201–218. [Google Scholar] [CrossRef] [PubMed]
- Leão, P.N.; Vasconcelos, M.T.; Vasconcelos, V.M. Allelopathy in freshwater cyanobacteria. Crit. Rev. Microbiol. 2009, 35, 271–282. [Google Scholar] [CrossRef]
- Kaplan, A.; Harel, M.; Kaplan-Levy, R.N.; Hadas, O.; Sukenik, A.; Dittmann, E. The languages spoken in the water body (or the biological role of cyanobacterial toxins). Front. Microbiol. 2012, 3, 138. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Keating, K.I. Allelopathic influence on blue-green bloom sequence in a eutrophic lake. Science 1977, 196, 885–887. [Google Scholar] [CrossRef] [PubMed]
- Keating, K.I. Blue-green algal inhibition of diatom growth: Transition from mesotrophic to eutrophic community structure. Science 1978, 199, 971–973. [Google Scholar] [CrossRef] [PubMed]
- Suikkanen, S.; Fistarol, G.O.; Granéli, E. Allelopathic effects of the Baltic cyanobacteria Nodularia spumdigena, Aphanizomenon flos-aquae and Anabaena lemmermannii on algal monocultures. J. Exp. Mar. Biol. Ecol. 2004, 308, 85–101. [Google Scholar] [CrossRef]
- Suikkanen, S.; Fistarol, G.O.; Granéli, E. Effects of cyanobacterial allelochemicals on a natural plankton community. Mar. Ecol. Prog. Ser. 2005, 287, 1–9. [Google Scholar] [CrossRef]
- Lirås, V.; Lindberg, M.; Nyström, P.; Annadotter, H.; Lawton, L.A.; Graf, B. Can ingested cyanobacteria be harmful to the signal crayfish (Pacifastacus leniusculus)? Freshw. Biol. 1998, 39, 233–242. [Google Scholar] [CrossRef]
- Lehtiniemi, M.; Engström-Öst, J.; Karjalainen, M.; Kozlowsky-Suzuki, B.; Viitasalo, M. Fate of cyanobacterial toxins in the pelagic food web: Transfer to copepods or to faecal pellets? Mar. Ecol. Prog. Ser. 2002, 241, 13–21. [Google Scholar] [CrossRef]
- Andersen, R.J.; Luu, H.A.; Chen, D.Z.; Holmes, C.F.; Kent, M.L.; Le Blanc, M.; Williams, D.E. Chemical and biological evidence links microcystins to salmon ‘netpen liver disease’. Toxicon 1993, 31, 1315–1323. [Google Scholar] [CrossRef]
- Gilbert, J.J. Differential effects of Anabaena affinis on cladocerans and rotifers: Mechanisms and implications. Ecology 1990, 71, 1727–1740. [Google Scholar] [CrossRef]
- Ferrão-Filho, A.d.S.; Azevedo, S.M.; DeMott, W.R. Effects of toxic and non-toxic cyanobacteria on the life history of tropical and temperate cladocerans. Freshwat. Biol. 2000, 45, 1–19. [Google Scholar] [CrossRef]
- Ghadouani, A.; Pinel-Alloul, B.; Prepas, E.E. Effects of experimentally induced cyanobacterial blooms on crustacean zooplankton communities. Freshwat. Biol. 2003, 48, 363–381. [Google Scholar] [CrossRef]
- Miles, C.; Stirling, D. Toxin Mass List, Version 16. 2020. Available online: https://www.researchgate.net/publication/337258461_Toxin_mass_list_COM_v160_microcystin_and_nodularin_lists_and_mass_calculators_for_mass_spectrometry_of_microcystins_nodularins_saxitoxins_and_anatoxins (accessed on 14 April 2021).
- Janssen, E.M.-L. Cyanobacterial peptides beyond microcystins–A review on co-occurrence, toxicity, and challenges for risk assessment. Water Res. 2019, 151, 488–499. [Google Scholar] [CrossRef] [PubMed]
- Jones, M.R.; Pinto, E.; Torres, M.A.; Dörr, F.; Mazur-Marzec, H.; Szubert, K.; Tartaglione, L.; Dell’Aversano, C.; Miles, C.O.; Beach, D.G. CyanoMetDB, a comprehensive public database of secondary metabolites from cyanobacteria. Water Res. 2021, 196, 117017. [Google Scholar] [CrossRef] [PubMed]
- Schatz, D.; Keren, Y.; Vardi, A.; Sukenik, A.; Carmeli, S.; Borner, T.; Dittmann, E.; Kaplan, A. Towards clarification of the biological role of microcystins, a family of cyanobacterial toxins. Environ. Microbiol. 2007, 9, 965–970. [Google Scholar] [CrossRef]
- Zilliges, Y.; Kehr, J.C.; Meissner, S.; Ishida, K.; Mikkat, S.; Hagemann, M.; Kaplan, A.; Borner, T.; Dittmann, E. The cyanobacterial hepatotoxin microcystin binds to proteins and increases the fitness of Microcystis under oxidative stress conditions. PLoS ONE 2011, 6, e17615. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Meissner, S.; Steinhauser, D.; Dittmann, E. Metabolomic analysis indicates a pivotal role of the hepatotoxin microcystin in high light adaptation of Microcystis. Environ. Microbiol. 2015, 17, 1497–1509. [Google Scholar] [CrossRef] [PubMed]
- Tomescu, A.; Honegger, R.; Rothwell, G. Earliest fossil record of bacterial–cyanobacterial mat consortia: The early Silurian Passage Creek biota (440 Ma, Virginia, USA). Geobiology 2008, 6, 120–124. [Google Scholar] [CrossRef]
- Paerl, H.W. Role of heterotrophic bacteria in promoting N 2 fixation by Anabaena in aquatic habitats. Microb. Ecol. 1977, 4, 215–231. [Google Scholar] [CrossRef]
- Woodhouse, J.N.; Kinsela, A.S.; Collins, R.N.; Bowling, L.C.; Honeyman, G.L.; Holliday, J.K.; Neilan, B.A. Microbial communities reflect temporal changes in cyanobacterial composition in a shallow ephemeral freshwater lake. ISME J. 2016, 10, 1337–1351. [Google Scholar] [CrossRef]
- Weiss, G.; Kovalerchick, D.; Lieman-Hurwitz, J.; Murik, O.; De Philippis, R.; Carmeli, S.; Sukenik, A.; Kaplan, A. Increased algicidal activity of Aeromonas veronii in response to Microcystis aeruginosa: Interspecies crosstalk and secondary metabolites synergism. Environ. Microbiol. 2019, 21, 1140–1150. [Google Scholar] [CrossRef]
- Jankowiak, J.G.; Gobler, C.J. The composition and function of microbiomes within Microcystis colonies are significantly different than native bacterial assemblages in two North American Lakes. Front. Microbiol. 2020, 11, 1016. [Google Scholar] [CrossRef]
- Weiss, G.; Kovalerchick, D.; Murik, O.; Sukenik, A.; Kaplan, A.; Carmeli, S. Secondary metabolites of Aeromonas veronii strain A134 isolated from a Microcystis aeruginosa bloom. Metabolites 2019, 9, 110. [Google Scholar] [CrossRef] [Green Version]
- Louati, I.; Pascault, N.; Debroas, D.; Bernard, C.; Humbert, J.-F.; Leloup, J. Structural diversity of bacterial communities associated with bloom-forming freshwater cyanobacteria differs according to the cyanobacterial genus. PLoS ONE 2015, 10, e0140614. [Google Scholar] [CrossRef] [Green Version]
- Yuan, L.; Zhu, W.; Xiao, L.; Yang, L. Phosphorus cycling between the colonial cyanobacterium Microcystis aeruginosa and attached bacteria, Pseudomonas. Aquat. Ecol. 2009, 43, 859–866. [Google Scholar] [CrossRef]
- Zhao, G.; Du, J.; Jia, Y.; Lv, Y.; Han, G.; Tian, X. The importance of bacteria in promoting algal growth in eutrophic lakes with limited available phosphorus. Ecol. Eng. 2012, 42, 107–111. [Google Scholar] [CrossRef]
- Liu, M.; Liu, L.; Chen, H.; Yu, Z.; Yang, J.R.; Xue, Y.; Huang, B.; Yang, J. Community dynamics of free-living and particle-attached bacteria following a reservoir Microcystis bloom. Sci. Total Environ. 2019, 660, 501–511. [Google Scholar] [CrossRef] [PubMed]
- Liu, L.; Chen, H.; Liu, M.; Yang, J.R.; Xiao, P.; Wilkinson, D.M.; Yang, J. Response of the eukaryotic plankton community to the cyanobacterial biomass cycle over 6 years in two subtropical reservoirs. ISME J. 2019, 13, 2196–2208. [Google Scholar] [CrossRef] [Green Version]
- Chun, S.-J.; Cui, Y.; Lee, C.S.; Cho, A.R.; Baek, K.; Choi, A.; Ko, S.-R.; Lee, H.-G.; Hwang, S.; Oh, H.-M. Characterization of distinct cyanoHABs-related modules in microbial recurrent association network. Front. Microbiol. 2019, 10, 1637. [Google Scholar] [CrossRef] [Green Version]
- Chun, S.-J.; Cui, Y.; Lee, J.J.; Choi, I.-C.; Oh, H.-M.; Ahn, C.-Y. Network analysis reveals succession of Microcystis genotypes accompanying distinctive microbial modules with recurrent patterns. Water Res. 2020, 170, 115326. [Google Scholar] [CrossRef]
- Wang, S.; Zhao, D.; Zeng, J.; Xu, H.; Huang, R.; Jiao, C.; Guo, L. Variations of bacterial community during the decomposition of Microcystis under different temperatures and biomass. BMC Microbiol. 2019, 19, 207. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhu, L.; Zancarini, A.; Louati, I.; De Cesare, S.; Duval, C.; Tambosco, K.; Bernard, C.; Debroas, D.; Song, L.; Leloup, J. Bacterial communities associated with four cyanobacterial genera display structural and functional differences: Evidence from an experimental approach. Front. Microbiol. 2016, 7, 1662. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lezcano, M.Á.; Velázquez, D.; Quesada, A.; El-Shehawy, R. Diversity and temporal shifts of the bacterial community associated with a toxic cyanobacterial bloom: An interplay between microcystin producers and degraders. Water Res. 2017, 125, 52–61. [Google Scholar] [CrossRef]
- Li, J.; Li, R.; Li, J. Current research scenario for microcystins biodegradation–a review on fundamental knowledge, application prospects and challenges. Sci. Total Environ. 2017, 595, 615–632. [Google Scholar] [CrossRef]
- Lezcano, M.Á.; Morón-López, J.; Agha, R.; López-Heras, I.; Nozal, L.; Quesada, A.; El-Shehawy, R. Presence or absence of mlr genes and nutrient concentrations co-determine the microcystin biodegradation efficiency of a natural bacterial community. Toxins 2016, 8, 318. [Google Scholar] [CrossRef]
- Li, J.; Shimizu, K.; Akasako, H.; Lu, Z.; Akiyama, S.; Goto, M.; Utsumi, M.; Sugiura, N. Assessment of the factors contributing to the variations in microcystins biodegradability of the biofilms on a practical biological treatment facility. Bioresour. Technol. 2015, 175, 463–472. [Google Scholar] [CrossRef]
- Ding, Q.; Liu, K.; Song, Z.; Sun, R.; Zhang, J.; Yin, L.; Pu, Y. Effects of microcystin-LR on metabolic functions and structure succession of sediment bacterial community under anaerobic conditions. Toxins 2020, 12, 183. [Google Scholar] [CrossRef] [Green Version]
- Yang, C.; Xia, C.; Zhou, S.; Liu, Y. The permeability effect of microcystin-RR on Escherichia coli and Bacillus subtilis. Chin. Sci. Bull. 2010, 55, 1894–1898. [Google Scholar] [CrossRef]
- Miguéns, D.; Valério, E. The impact of some microcystins on the growth of heterotrophic bacteria from Portuguese freshwater reservoir. Limnetica 2015, 34, 215–226. [Google Scholar] [CrossRef]
- Tang, X.; Gao, G.; Chao, J.; Wang, X.; Zhu, G.; Qin, B. Dynamics of organic-aggregate-associated bacterial communities and related environmental factors in Lake Taihu, a large eutrophic shallow lake in China. Limnol. Oceanogr. 2010, 55, 469–480. [Google Scholar] [CrossRef]
- Wilhelm, S.W.; Farnsley, S.E.; LeCleir, G.R.; Layton, A.C.; Satchwell, M.F.; DeBruyn, J.M.; Boyer, G.L.; Zhu, G.; Paerl, H.W. The relationships between nutrients, cyanobacterial toxins and the microbial community in Taihu (Lake Tai), China. Harmful Algae 2011, 10, 207–215. [Google Scholar] [CrossRef]
- Su, X.; Steinman, A.D.; Tang, X.; Xue, Q.; Zhao, Y.; Xie, L. Response of bacterial communities to cyanobacterial harmful algal blooms in Lake Taihu, China. Harmful Algae 2017, 68, 168–177. [Google Scholar] [CrossRef] [PubMed]
- Cole, J.J. Interactions between bacteria and algae in aquatic ecosystems. Annu. Rev. Ecol. Syst. 1982, 13, 291–314. [Google Scholar] [CrossRef]
- Celussi, M.; Cataletto, B. Annual dynamics of bacterioplankton assemblages in the Gulf of Trieste (Northern Adriatic Sea). Gene 2007, 406, 113–123. [Google Scholar] [CrossRef]
- Xu, Z.; Te, S.H.; He, Y.; Gin, K.Y.-H. The characteristics and dynamics of cyanobacteria–heterotrophic bacteria between two estuarine reservoirs–tropical versus sub-tropical regions. Front. Microbiol. 2018, 9, 2531. [Google Scholar] [CrossRef] [Green Version]
- Li, D.; Wu, N.; Tang, S.; Su, G.; Li, X.; Zhang, Y.; Wang, G.; Zhang, J.; Liu, H.; Hecker, M. Factors associated with blooms of cyanobacteria in a large shallow lake, China. Environ. Sci. Eur. 2018, 30, 27. [Google Scholar] [CrossRef]
- Xue, Y.; Chen, H.; Yang, J.R.; Liu, M.; Huang, B.; Yang, J. Distinct patterns and processes of abundant and rare eukaryotic plankton communities following a reservoir cyanobacterial bloom. ISME J. 2018, 12, 2263–2277. [Google Scholar] [CrossRef] [Green Version]
- Paerl, H.W.; Fulton, R.S.; Moisander, P.H.; Dyble, J.; Boynton, A. Harmful freshwater algal blooms, with an emphasis on cyanobacteria. Sci. World J. 2001, 1, 76–113. [Google Scholar] [CrossRef] [PubMed]
- Filstrup, C.T.; Hillebrand, H.; Heathcote, A.J.; Harpole, W.S.; Downing, J.A. Cyanobacteria dominance influences resource use efficiency and community turnover in phytoplankton and zooplankton communities. Ecol. Lett. 2014, 17, 464–474. [Google Scholar] [CrossRef]
- McClure, R.S.; Overall, C.C.; Hill, E.A.; Song, H.-S.; Charania, M.; Bernstein, H.C.; McDermott, J.E.; Beliaev, A.S. Species-specific transcriptomic network inference of interspecies interactions. ISME J. 2018, 12, 2011–2023. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Singh, D.P.; Tyagi, M.B.; Kumar, A.; Thakur, J.K.; Kumar, A. Antialgal activity of a hepatotoxin-producing cyanobacterium, Microcystis aeruginosa. World J. Microbiol. Biotechnol. 2001, 17, 15–22. [Google Scholar] [CrossRef]
- Li, Y.; Li, D. Competition between toxic Microcystis aeruginosa and nontoxic Microcystis wesenbergii with Anabaena PCC7120. J. Appl. Phycol. 2012, 24, 69–78. [Google Scholar] [CrossRef] [Green Version]
- Berry, M.A.; Davis, T.W.; Cory, R.M.; Duhaime, M.B.; Johengen, T.H.; Kling, G.W.; Marino, J.A.; Den Uyl, P.A.; Gossiaux, D.; Dick, G.J. Cyanobacterial harmful algal blooms are a biological disturbance to Western Lake Erie bacterial communities. Environ. Microbiol. 2017, 19, 1149–1162. [Google Scholar] [CrossRef] [PubMed]
- Hu, Z.q.; Liu, Y.d.; Li, D.h. Physiological and biochemical analyses of microcystin-RR toxicity to the cyanobacterium Synechococcus elongatus. Environ. Toxicol. 2004, 19, 571–577. [Google Scholar] [CrossRef] [PubMed]
- Do Carmo Bittencourt-Oliveira, M.; Camargo-Santos, D.; do Nascimento Moura, A.; Francisco, I.B.; dos Santos Dias, C.T.; Molica, R.J.R.; Cordeiro-Araújo, M.K. Effects of toxic and non-toxic crude extracts on different Microcystis species (Cyanobacteria). Afr. J. Microbiol. Res. 2013, 7, 2596–2600. [Google Scholar] [CrossRef]
- Toporowska, M.; Mazur-Marzec, H.; Pawlik-Skowrońska, B. The effects of cyanobacterial bloom extracts on the biomass, Chl-a, MC and other oligopeptides contents in a natural Planktothrix agardhii population. Int. J. Environ. Res. Public Health 2020, 17, 2881. [Google Scholar] [CrossRef] [PubMed]
- Omidi, A.; Esterhuizen-Londt, M.; Pflugmacher, S. Desmodesmus subspicatus co-cultured with microcystin producing (PCC 7806) and the non-producing (PCC 7005) strains of Microcystis aeruginosa. Ecotoxicology 2019, 28, 834–842. [Google Scholar] [CrossRef] [Green Version]
- Ma, Z.; Fang, T.; Thring, R.W.; Li, Y.; Yu, H.; Zhou, Q.; Zhao, M. Toxic and non-toxic strains of Microcystis aeruginosa induce temperature dependent allelopathy toward growth and photosynthesis of Chlorella vulgaris. Harmful Algae 2015, 48, 21–29. [Google Scholar] [CrossRef]
- Daniel, E.; Weiss, G.; Murik, O.; Sukenik, A.; Lieman-Hurwitz, J.; Kaplan, A. The response of Microcystis aeruginosa strain MGK to a single or two consecutive H2O2 applications. Environ. Microbiol. Rep. 2019, 11, 621–629. [Google Scholar] [CrossRef]
- Omidi, A.; Esterhuizen-Londt, M.; Pflugmacher, S. Interspecies interactions between Microcystis aeruginosa PCC 7806 and Desmodesmus subspicatus SAG 86.81 in a co-cultivation system at various growth phases. Environ. Int. 2019, 131, 105052. [Google Scholar] [CrossRef]
- Dong, J.; Li, C.; Chang, M.; Dai, D.; Liu, S.; Quan, B.; Zhang, Y.; Gao, Y. Effects of toxic cyanobacterium Microcystis aeruginosa on the morphology of green alga Chlorella vulgaris. Ann. Limnol. Int. J. Limnol. 2019, 55, 7. [Google Scholar] [CrossRef]
- Vardi, A.; Schatz, D.; Beeri, K.; Motro, U.; Sukenik, A.; Levine, A.; Kaplan, A. Dinoflagellate-cyanobacterium communication may determine the composition of phytoplankton assemblage in a mesotrophic lake. Curr. Biol. 2002, 12, 1767–1772. [Google Scholar] [CrossRef] [Green Version]
- Kearns, K.D.; Hunter, M.D. Green algal extracellular products regulate antialgal toxin production in a cyanobacterium. Environ. Microbiol. 2000, 2, 291–297. [Google Scholar] [CrossRef] [PubMed]
- Chen, J.; Guo, R. Inhibition effect of green alga on cyanobacteria by the interspecies interactions. Int. J. Environ. Sci. Technol. 2014, 11, 839–842. [Google Scholar] [CrossRef] [Green Version]
- Harel, M.; Weiss, G.; Lieman-Hurwitz, J.; Gun, J.; Lev, O.; Lebendiker, M.; Temper, V.; Block, C.; Sukenik, A.; Zohary, T.; et al. Interactions between Scenedesmus and Microcystis may be used to clarify the role of secondary metabolites. Environ. Microbiol. Rep. 2013, 5, 97–104. [Google Scholar] [CrossRef]
- Gulati, R.; Demott, W. The role of food quality for zooplankton: Remarks on the state-of-the-art, perspectives and priorities. Freshw. Biol. 1997, 38, 753–768. [Google Scholar] [CrossRef]
- Müller-Navarra, D.C.; Brett, M.T.; Liston, A.M.; Goldman, C.R. A highly unsaturated fatty acid predicts carbon transfer between primary producers and consumers. Nature 2000, 403, 74–77. [Google Scholar] [CrossRef] [PubMed]
- Gliwicz, Z.M.; Lampert, W. Food thresholds in Daphnia species in the absence and presence of blue-green filaments. Ecology 1990, 71, 691–702. [Google Scholar] [CrossRef]
- DeMott, W.R.; Gulati, R.D.; Van Donk, E. Daphnia food limitation in three hypereutrophic Dutch lakes: Evidence for exclusion of large-bodied species by interfering filaments of cyanobacteria. Limnol. Oceanogr. 2001, 46, 2054–2060. [Google Scholar] [CrossRef]
- Wilson, A.E.; Sarnelle, O.; Tillmanns, A.R. Effects of cyanobacterial toxicity and morphology on the population growth of freshwater zooplankton: Meta-analyses of laboratory experiments. Limnol. Oceanogr. 2006, 51, 1915–1924. [Google Scholar] [CrossRef] [Green Version]
- Hulot, F.D.; Carmignac, D.; Legendre, S.; Yepremian, C.; Bernard, C. Effects of microcystin-producing and microcystin-free strains of Planktothrix agardhii on long-term population dynamics of Daphnia magna. Ann. Limnol. Int. J. Limnol. 2012, 48, 337–347. [Google Scholar] [CrossRef] [Green Version]
- Kurmayer, R.; Schober, E.; Tonk, L.; Visser, P.M.; Christiansen, G. Spatial divergence in the proportions of genes encoding toxic peptide synthesis among populations of the cyanobacterium Planktothrix in European lakes. FEMS Microbiol. Lett. 2011, 317, 127–137. [Google Scholar] [CrossRef] [Green Version]
- Schwarzenberger, A.; Kurmayer, R.; Martin-Creuzburg, D. Toward disentangling the multiple nutritional constraints imposed by Planktothrix: The significance of harmful secondary metabolites and sterol limitation. Front. Microbiol. 2020, 11. [Google Scholar] [CrossRef]
- Tang, H.; Hou, X.; Xue, X.; Chen, R.; Zhu, X.; Huang, Y.; Chen, Y. Microcystis aeruginosa strengthens the advantage of Daphnia similoides in competition with Moina micrura. Sci. Rep. 2017, 7, 1–9. [Google Scholar] [CrossRef] [Green Version]
- Pflugmacher, S.; Wiegand, C.; Oberemm, A.; Beattie, K.A.; Krause, E.; Codd, G.A.; Steinberg, C.E. Identification of an enzymatically formed glutathione conjugate of the cyanobacterial hepatotoxin microcystin-LR: The first step of detoxication. Biochim. Biophys. Acta Gen. Subj. 1998, 1425, 527–533. [Google Scholar] [CrossRef]
- Gustafsson, S.; Rengefors, K.; Hansson, L.-A. Increased consumer fitness following transfer of toxin tolerance to offspring via maternal effects. Ecology 2005, 86, 2561–2567. [Google Scholar] [CrossRef] [Green Version]
- Sarnelle, O.; Wilson, A.E. Local adaptation of Daphnia pulicaria to toxic cyanobacteria. Limnol. Oceanogr. 2005, 50, 1565–1570. [Google Scholar] [CrossRef] [Green Version]
- Herrera, N.A.; Echeverri, L.F.; Ferrao-Filho, A.S. Effects of phytoplankton extracts containing the toxin microcystin-LR on the survival and reproduction of cladocerans. Toxicon 2015, 95, 38–45. [Google Scholar] [CrossRef] [PubMed]
- Herrera, N.; Palacio, J.; Echeverri, F.; Ferrão-Filho, A. Effects of a cyanobacterial bloom sample containing microcystin-LR on the ecophysiology of Daphnia similis. Toxicol. Rep. 2014, 1, 909–914. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Freitas, E.C.; Pinheiro, C.; Rocha, O.; Loureiro, S. Can mixtures of cyanotoxins represent a risk to the zooplankton? The case study of Daphnia magna Straus exposed to hepatotoxic and neurotoxic cyanobacterial extracts. Harmful Algae 2014, 31, 143–152. [Google Scholar] [CrossRef]
- Smutná, M.; Babica, P.; Jarque, S.; Hilscherová, K.; Maršálek, B.; Haeba, M.; Bláha, L. Acute, chronic and reproductive toxicity of complex cyanobacterial blooms in Daphnia magna and the role of microcystins. Toxicon 2014, 79, 11–18. [Google Scholar] [CrossRef]
- Chislock, M.F.; Sarnelle, O.; Jernigan, L.M.; Wilson, A.E. Do high concentrations of microcystin prevent Daphnia control of phytoplankton? Water Res. 2013, 47, 1961–1970. [Google Scholar] [CrossRef]
- Sadler, T.; von Elert, E. Physiological interaction of Daphnia and Microcystis with regard to cyanobacterial secondary metabolites. Aquat. Toxicol. 2014, 156, 96–105. [Google Scholar] [CrossRef]
- Paes, T.A.S.V.; Costa, I.A.S.d.; Silva, A.P.C.; Eskinazi-Sant’ anna, E. Can microcystins affect zooplankton structure community in tropical eutrophic reservoirs? Braz. J. Biol. 2016, 76, 450–460. [Google Scholar] [CrossRef]
- Zhao, S.; Wang, Y.; Li, D. Effects of toxic and non-toxic Microcystis aeruginosa in different mixtures with Scenedesmus obliquus on growth of Brachionus calyciflorus. J. Freshw. Ecol. 2014, 29, 377–386. [Google Scholar] [CrossRef] [Green Version]
- Liang, Y.; Ouyang, K.; Chen, X.; Su, Y.; Yang, J. Life strategy and grazing intensity responses of Brachionus calyciflorus fed on different concentrations of microcystin-producing and microcystin-free Microcystis aeruginosa. Sci. Rep. 2017, 7, 43127. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Nandini, S.; Sánchez-Zamora, C.; Sarma, S. Toxicity of cyanobacterial blooms from the reservoir Valle de Bravo (Mexico): A case study on the rotifer Brachionus calyciflorus. Sci. Total Environ. 2019, 688, 1348–1358. [Google Scholar] [CrossRef] [PubMed]
- Liang, Y.; Gao, T.; Shao, L.; Min, Y.; Yang, J. Effects of microcystin-LR and nitrite on the lifespan, reproduction, and heat shock responses of rotifer Brachionus calyciflorus at different temperatures. Aquat. Sci. 2020, 82, 1–14. [Google Scholar] [CrossRef]
- Koski, M.; Schmidt, K.; Engström-Öst, J.; Viitasalo, M.; Jónasdóttir, S.; Repka, S.; Sivonen, K. Calanoid copepods feed and produce eggs in the presence of toxic cyanobacteria Nodularia spumigena. Limnol. Oceanogr. 2002, 47, 878–885. [Google Scholar] [CrossRef]
- Wilson, A.E.; Hay, M.E. A direct test of cyanobacterial chemical defense: Variable effects of microcystin-treated food on two Daphnia pulicaria clones. Limnol. Oceanogr. 2007, 52, 1467–1479. [Google Scholar] [CrossRef] [Green Version]
- Ferrão-Filho, A.d.S.; Da Costa, S.M.; Ribeiro, M.G.L.; Azevedo, S.M. Effects of a saxitoxin-producer strain of Cylindrospermopsis raciborskii (cyanobacteria) on the swimming movements of cladocerans. Environ. Toxicol. 2008, 23, 161–168. [Google Scholar] [CrossRef] [PubMed]
- Ger, K.A.; Teh, S.J.; Goldman, C.R. Microcystin-LR toxicity on dominant copepods Eurytemora affinis and Pseudodiaptomus forbesi of the upper San Francisco Estuary. Sci. Total Environ. 2009, 407, 4852–4857. [Google Scholar] [CrossRef] [PubMed]
- Ger, K.A.; Hansson, L.A.; Lürling, M. Understanding cyanobacteria-zooplankton interactions in a more eutrophic world. Freshw. Biol. 2014, 59, 1783–1798. [Google Scholar] [CrossRef] [Green Version]
- Ullah, H.; Nagelkerken, I.; Goldenberg, S.U.; Fordham, D.A. Climate change could drive marine food web collapse through altered trophic flows and cyanobacterial proliferation. PLoS Biol. 2018, 16, e2003446. [Google Scholar] [CrossRef]
- Amorim, C.A.; Ulisses, C.; Moura, A.N. Biometric and physiological responses of Egeria densa Planch. cultivated with toxic and non-toxic strains of Microcystis. Aquat. Toxicol. 2017, 191, 201–208. [Google Scholar] [CrossRef]
- Senavirathna, M.J.; Muhetaer, G.; Zhaozhi, L.; Fujino, T. Allelopathic influence of low concentration Microcystis aeruginosa on Egeria densa under different light intensities. Chem. Ecol. 2020. [Google Scholar] [CrossRef]
- Li, Q.; Gu, P.; Zhang, H.; Luo, X.; Zhang, J.; Zheng, Z. Response of submerged macrophytes and leaf biofilms to the decline phase of Microcystis aeruginosa: Antioxidant response, ultrastructure, microbial properties, and potential mechanism. Sci. Total Environ. 2020, 699, 134325. [Google Scholar] [CrossRef]
- Pei, Y.; Liu, L.; Hilt, S.; Xu, R.; Wang, B.; Li, C.; Chang, X. Root exudated algicide of Eichhornia crassipes enhances allelopathic effects of cyanobacteria Microcystis aeruginosa on green algae. Hydrobiologia 2018, 823, 67–77. [Google Scholar] [CrossRef]
- Pflugmacher, S. Promotion of oxidative stress in the aquatic macrophyte Ceratophyllum demersum during biotransformation of the cyanobacterial toxin microcystin-LR. Aquat. Toxicol. 2004, 70, 169–178. [Google Scholar] [CrossRef] [PubMed]
- Chen, G.; Zheng, Z.; Bai, M.; Li, Q. Chronic effects of microcystin-LR at environmental relevant concentrations on photosynthesis of Typha angustifolia Linn. Ecotoxicology 2020, 29, 514–523. [Google Scholar] [CrossRef]
- Wang, N.; Wang, C. Effects of microcystin-LR on the tissue growth and physiological responses of the aquatic plant Iris pseudacorus L. Aquat. Toxicol. 2018, 200, 197–205. [Google Scholar] [CrossRef] [PubMed]
- Christensen, V.G.; Khan, E. Freshwater neurotoxins and concerns for human, animal, and ecosystem health: A review of anatoxin-a and saxitoxin. Sci. Total Environ. 2020, 736, 139515. [Google Scholar] [CrossRef]
- Bouma-Gregson, K.; Olm, M.R.; Probst, A.J.; Anantharaman, K.; Power, M.E.; Banfield, J.F. Microbial diversity and metabolic potential in cyanotoxin producing cyanobacterial mats throughout a river network. bioRxiv 2018, 294421. [Google Scholar] [CrossRef] [Green Version]
- Thomson-Laing, G.; Puddick, J.; Laroche, O.; Fulton, S.; Steiner, K.; Heath, M.W.; Wood, S.A. Broad and fine scale variability in bacterial diversity and cyanotoxin quotas in benthic cyanobacterial mats. Front. Microbiol. 2020, 11, 129. [Google Scholar] [CrossRef]
- Cerasino, L.; Salmaso, N. Co-occurrence of anatoxin-a and microcystins in Lake Garda and other deep subalpine lakes. Adv. Oceanogr. Limnol. 2020, 11. [Google Scholar] [CrossRef]
- Bouma-Gregson, K.; Kudela, R.M.; Power, M.E. Widespread anatoxin-a detection in benthic cyanobacterial mats throughout a river network. PLoS ONE 2018, 13, e0197669. [Google Scholar] [CrossRef] [Green Version]
- Li, Q.; Gu, P.; Zhang, C.; Luo, X.; Zhang, H.; Zhang, J.; Zheng, Z. Combined toxic effects of anatoxin-a and microcystin-LR on submerged macrophytes and biofilms. J. Hazard. Mater. 2020, 389, 122053. [Google Scholar] [CrossRef] [PubMed]
- Papenfort, K.; Bassler, B.L. Quorum sensing signal–response systems in Gram-negative bacteria. Nat. Rev. Microbiol. 2016, 14, 576. [Google Scholar] [CrossRef] [PubMed]
- Chia, M.A.; Cordeiro-Araujo, M.K.; do Carmo Bittencourt-Oliveira, M. Growth and antioxidant response of Microcystis aeruginosa (Cyanobacteria) exposed to anatoxin-a. Harmful Algae 2015, 49, 135–146. [Google Scholar] [CrossRef]
- Chia, M.A.; Cordeiro-Araújo, M.K.; Lorenzi, A.S.; do Carmo Bittencourt-Oliveira, M. Does anatoxin-a influence the physiology of Microcystis aeruginosa and Acutodesmus acuminatus under different light and nitrogen conditions? Environ. Sci. Pollut. Res. 2016, 23, 23092–23102. [Google Scholar] [CrossRef] [PubMed]
- Chia, M.A.; Kramer, B.J.; Jankowiak, J.G.; Bittencourt-Oliveira, M.d.C.; Gobler, C.J. The individual and combined effects of the cyanotoxins, anatoxin-a and microcystin-LR, on the growth, toxin production, and nitrogen fixation of prokaryotic and eukaryotic algae. Toxins 2019, 11, 43. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kearns, K.; Hunter, M. Toxin-producing Anabaena flos-aquae induces settling of Chlamydomonas reinhardtii, a competing motile alga. Microb. Ecol. 2001, 42, 80–86. [Google Scholar] [CrossRef]
- De Abreu, F.Q.; da S. Ferrão-Filho, A. Effects of an Anatoxin-a (s)-producing strain of Anabaena spiroides (Cyanobacteria) on the survivorship and somatic growth of two Daphnia similis clones. J. Environ. Prot. 2013, 12–18. [Google Scholar] [CrossRef] [Green Version]
- Bownik, A.; Pawlik-Skowrońska, B. Early indicators of behavioral and physiological disturbances in Daphnia magna (Cladocera) induced by cyanobacterial neurotoxin anatoxin-a. Sci. Total Environ. 2019, 695, 133913. [Google Scholar] [CrossRef]
- Schwarzenberger, A.; Martin-Creuzburg, D. Daphnia’s adaptive molecular responses to the cyanobacterial neurotoxin anatoxin-α are maternally transferred. Toxins 2021, 13, 326. [Google Scholar] [CrossRef]
- Ha, M.-H.; Pflugmacher, S. Time-dependent alterations in growth, photosynthetic pigments and enzymatic defense systems of submerged Ceratophyllum demersum during exposure to the cyanobacterial neurotoxin anatoxin-a. Aquat. Toxicol. 2013, 138, 26–34. [Google Scholar] [CrossRef] [PubMed]
- Ha, M.-H.; Contardo-Jara, V.; Pflugmacher, S. Uptake of the cyanobacterial neurotoxin, anatoxin-a, and alterations in oxidative stress in the submerged aquatic plant Ceratophyllum demersum. Ecotoxicol. Environ. Saf. 2014, 101, 205–212. [Google Scholar] [CrossRef]
- Pratheepa, V.; Vasconcelos, V. Binding and pharmacokinetics of the sodium channel blocking toxins (Saxitoxin and the Tetrodotoxins). Mini Rev. Med. Chem. 2017, 17, 320–327. [Google Scholar] [CrossRef]
- Walker, J.R.; Novick, P.A.; Parsons, W.H.; McGregor, M.; Zablocki, J.; Pande, V.S.; Du Bois, J. Marked difference in saxitoxin and tetrodoxin affinity for the human nociceptive voltage-gated sodium channel (Naᵥ1. 7). Proc. Natl. Acad. Sci. USA 2012, 109, 18102–18107. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Trainer, V.L.; Baden, D.G. High affinity binding of red tide neurotoxins to marine mammal brain. Aquat. Toxicol. 1999, 46, 139–148. [Google Scholar] [CrossRef]
- Pomati, F.; Rossetti, C.; Calamari, D.; Neilan, B.A. Effects of saxitoxin (STX) and veratridine on bacterial Na+-K+ fluxes: A prokaryote-based STX bioassay. Appl. Environ. Microbiol. 2003, 69, 7371–7376. [Google Scholar] [CrossRef] [Green Version]
- Cusick, K.D.; Sayler, G.S. An overview on the marine neurotoxin, saxitoxin: Genetics, molecular targets, methods of detection and ecological functions. Mar. Drugs 2013, 11, 991–1018. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhang, X. Biodiversity of the symbiotic bacteria associated with toxic marine dinoflagellate Alexandrium tamarense. JBM 2015, 3, 23. [Google Scholar] [CrossRef] [Green Version]
- Laureano-Rosario, A.E.; McFarland, M.; Bradshaw II, D.J.; Metz, J.; Brewton, R.A.; Pitts, T.; Perricone, C.; Schreiber, S.; Stockley, N.; Wang, G. Dynamics of microcystins and saxitoxin in the Indian River Lagoon, Florida. Harmful Algae 2021, 103, 102012. [Google Scholar] [CrossRef]
- Raudonis, R.A. Bacteria Associated with Paralytic Shellfish Toxin-Producing Strains of Anabaena circinalis. Master’s Thesis, University of Alaska Fairbanks, College, AK, USA, December 2007. [Google Scholar]
- Do Carmo Bittencourt-Oliveira, M.; Chia, M.A.; Camargo-Santos, D.; Dias, C.T. The effect of saxitoxin and non-saxitoxin extracts of Cylindrospermopsis raciborskii (Cyanobacteria) on cyanobacteria and green microalgae. J. Appl. Phycol. 2016, 28, 241–250. [Google Scholar] [CrossRef] [Green Version]
- Perreault, F.; Matias, M.S.; Melegari, S.P.; de Carvalho Pinto, C.R.S.; Creppy, E.E.; Popovic, R.; Matias, W.G. Investigation of animal and algal bioassays for reliable saxitoxin ecotoxicity and cytotoxicity risk evaluation. Ecotoxicol. Environ. Saf. 2011, 74, 1021–1026. [Google Scholar] [CrossRef] [PubMed]
- Melegari, S.P.; Perreault, F.; Moukha, S.; Popovic, R.; Creppy, E.E.; Matias, W.G. Induction to oxidative stress by saxitoxin investigated through lipid peroxidation in Neuro 2A cells and Chlamydomonas reinhardtii alga. Chemosphere 2012, 89, 38–43. [Google Scholar] [CrossRef] [Green Version]
- Haney, J.F.; Sasner, J.J.; Ikawa, M. Effects of products released by Aphanizomenon flos-aquae and purified saxitoxin on the movements of Daphnia carinata feeding appendages. Limnol. Oceanogr. 1995, 40, 263–272. [Google Scholar] [CrossRef]
- Ferrão-Filho, A.d.S.; da Silva, D.A.C. Saxitoxin-producing Raphidiopsis raciborskii (cyanobacteria) inhibits swimming and physiological parameters in Daphnia similis. Sci. Total Environ. 2020, 706, 135751. [Google Scholar] [CrossRef]
- Ferrão-Filho, A.d.S.; Dias, T.M.; Pereira, U.J.; dos Santos, J.A.A.; Kozlowsky-Suzuki, B. Nutritional and toxicity constraints of phytoplankton from a Brazilian reservoir to the fitness of cladoceran species. Environ. Sci. Pollut. Res. 2019, 26, 12881–12893. [Google Scholar] [CrossRef] [PubMed]
- Ferrão-Filho, A.S.; Pereira, U.J.; Vilar, M.C.; de Magalhães, L.; Marinho, M.M. Can small-bodied Daphnia control Raphidiopsis raciborskii in eutrophic tropical lakes? A mesocosm experiment. Environ. Sci. Pollut. Res. 2020, 27, 35459–35473. [Google Scholar] [CrossRef]
- Rangel, L.M.; Ger, K.A.; Silva, L.H.; Soares, M.C.S.; Faassen, E.J.; Lürling, M. Toxicity overrides morphology on Cylindrospermopsis raciborskii grazing resistance to the calanoid copepod Eudiaptomus gracilis. Microb. Ecol. 2016, 71, 835–844. [Google Scholar] [CrossRef] [Green Version]
- Santos, G.D.; Vilar, M.C.P.; de Oliveira Azevedo, S.M.F. Acute toxicity of neurotoxin-producing Raphidiopsis (Cylindrospermopsis) raciborskii ITEP-A1 (Cyanobacteria) on the neotropical cladoceran Macrothrix spinosa. Ecotoxicol. Environ. Contam. 2020, 15, 1–8. [Google Scholar] [CrossRef]
- Ferrão-Filho, A.d.S.; de Abreu, S.D.; de Oliveira, T.A.; de Magalhães, V.F.; Pflugmacher, S.; da Silva, E.M. Single and combined effects of microcystin and saxitoxin-producing cyanobacteria on the fitness and antioxidant defenses of cladocerans. Environ. Toxicol. Chem. 2017, 36, 2689–2697. [Google Scholar] [CrossRef]
- Moreira, C.; Azevedo, J.; Antunes, A.; Vasconcelos, V. Cylindrospermopsin: Occurrence, methods of detection and toxicology. J. Appl. Microbiol. 2013, 114, 605–620. [Google Scholar] [CrossRef]
- Antunes, J.T.; Leão, P.N.; Vasconcelos, V.M. Cylindrospermopsis raciborskii: Review of the distribution, phylogeography, and ecophysiology of a global invasive species. Front. Microbiol. 2015, 6, 473. [Google Scholar] [CrossRef] [Green Version]
- Rzymski, P.; Poniedziałek, B. In search of environmental role of cylindrospermopsin: A review on global distribution and ecology of its producers. Water Res. 2014, 66, 320–337. [Google Scholar] [CrossRef]
- Rasmussen, J.P.; Cursaro, M.; Froscio, S.M.; Saint, C.P. An examination of the antibiotic effects of cylindrospermopsin on common gram-positive and gram-negative bacteria and the protozoan Naegleria lovaniensis. Environ. Toxicol. 2008, 23, 36–43. [Google Scholar] [CrossRef]
- Wormer, L.; Cirés, S.; Carrasco, D.; Quesada, A. Cylindrospermopsin is not degraded by co-occurring natural bacterial communities during a 40-day study. Harmful Algae 2008, 7, 206–213. [Google Scholar] [CrossRef]
- Dziga, D.; Kokocinski, M.; Maksylewicz, A.; Czaja-Prokop, U.; Barylski, J. Cylindrospermopsin biodegradation abilities of Aeromonas sp. isolated from Rusałka Lake. Toxins 2016, 8, 55. [Google Scholar] [CrossRef] [Green Version]
- Martínez-Ruiz, E.B.; Cooper, M.; Fastner, J.; Szewzyk, U. Manganese-Oxidizing bacteria isolated from natural and technical systems remove cylindrospermopsin. Chemosphere 2020, 238, 124625. [Google Scholar] [CrossRef] [PubMed]
- B-Béres, V.; Vasas, G.; Dobronoki, D.; Gonda, S.; Nagy, S.A.; Bácsi, I. Effects of cylindrospermopsin producing cyanobacterium and its crude extracts on a benthic green alga—competition or allelopathy? Mar. Drugs 2015, 13, 6703–6722. [Google Scholar] [CrossRef] [Green Version]
- Antunes, J.T.; Leão, P.N.; Vasconcelos, V.M. Influence of biotic and abiotic factors on the allelopathic activity of the cyanobacterium Cylindrospermopsis raciborskii strain LEGE 99043. Microb. Ecol. 2012, 64, 584–592. [Google Scholar] [CrossRef] [PubMed]
- Pinheiro, C.; Azevedo, J.; Campos, A.; Loureiro, S.; Vasconcelos, V. Absence of negative allelopathic effects of cylindrospermopsin and microcystin-LR on selected marine and freshwater phytoplankton species. Hydrobiologia 2013, 705, 27–42. [Google Scholar] [CrossRef] [Green Version]
- Kovács, A.W.; Tóth, V.R.; Pálffy, K. The effects of interspecific interactions between bloom forming cyanobacteria and Scenedesmus quadricauda (chlorophyta) on their photophysiology. Acta Biol. Hung. 2018, 69, 210–223. [Google Scholar] [CrossRef] [Green Version]
- Mello, M.M.E.; Soares, M.C.S.; Roland, F.; Lürling, M. Growth inhibition and colony formation in the cyanobacterium Microcystis aeruginosa induced by the cyanobacterium Cylindrospermopsis raciborskii. J. Plankton Res. 2012, 34, 987–994. [Google Scholar] [CrossRef] [Green Version]
- Chia, M.A.; Cordeiro-Araújo, M.K.; Lorenzi, A.S.; do Carmo Bittencourt-Oliveira, M. Cylindrospermopsin induced changes in growth, toxin production and antioxidant response of Acutodesmus acuminatus and Microcystis aeruginosa under differing light and nitrogen conditions. Ecotoxicol. Environ. Saf. 2017, 142, 189–199. [Google Scholar] [CrossRef]
- Bar-Yosef, Y.; Sukenik, A.; Hadas, O.; Viner-Mozzini, Y.; Kaplan, A. Enslavement in the water body by toxic Aphanizomenon ovalisporum, inducing alkaline phosphatase in phytoplanktons. Curr. Biol. 2010, 20, 1557–1561. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Dobronoki, D.; Viktória, B.; Vasas, G.; Gonda, S.; Nagy, S.A.; Bácsi, I. Potential role of the cellular matrix of Aphanizomenon strains in the effects of cylindrospermopsin—an experimental study. J. Appl. Phycol. 2019, 31, 1805–1817. [Google Scholar] [CrossRef] [Green Version]
- Leonard, J.A.; Paerl, H.W. Zooplankton community structure, micro-zooplankton grazing impact, and seston energy content in the St. Johns river system, Florida as influenced by the toxic cyanobacterium Cylindrospermopsis raciborskii. Hydrobiologia 2005, 537, 89–97. [Google Scholar] [CrossRef]
- Nogueira, I.C.; Saker, M.L.; Pflugmacher, S.; Wiegand, C.; Vasconcelos, V.M. Toxicity of the cyanobacterium Cylindrospermopsis raciborskii to Daphnia magna. Environ. Toxicol. 2004, 19, 453–459. [Google Scholar] [CrossRef]
- Nogueira, I.C.; Lobo-da-Cunha, A.; Vasconcelos, V.M. Effects of Cylindrospermopsis raciborskii and Aphanizomenon ovalisporum (cyanobacteria) ingestion on Daphnia magna midgut and associated diverticula epithelium. Aquat. Toxicol. 2006, 80, 194–203. [Google Scholar] [CrossRef]
- Dao, T.S.; Ortiz-Rodríguez, R.; Do-Hong, L.C.; Wiegand, C. Non-microcystin and non-cylindrospermopsin producing cyanobacteria affect the biochemical responses and behavior of Daphnia magna. Int. Rev. Hydrobiol. 2013, 98, 235–244. [Google Scholar] [CrossRef]
- Flores-Rojas, N.C.; Esterhuizen-Londt, M.; Pflugmacher, S. Uptake, growth, and pigment changes in Lemna minor L. exposed to environmental concentrations of cylindrospermopsin. Toxins 2019, 11, 650. [Google Scholar] [CrossRef] [Green Version]
- Flores-Rojas, N.C.; Esterhuizen-Londt, M.; Pflugmacher, S. Antioxidative stress responses in the floating macrophyte Lemna minor L. with cylindrospermopsin exposure. Aquat. Toxicol. 2015, 169, 188–195. [Google Scholar] [CrossRef]
- Flores-Rojas, N.C.; Esterhuizen, M. Uptake and effects of cylindrospermopsin: Biochemical, physiological and biometric responses in the submerged macrophyte Egeria densa Planch. Water 2020, 12, 2997. [Google Scholar] [CrossRef]
- Chiswell, R.K.; Shaw, G.R.; Eaglesham, G.; Smith, M.J.; Norris, R.L.; Seawright, A.A.; Moore, M.R. Stability of cylindrospermopsin, the toxin from the cyanobacterium, Cylindrospermopsis raciborskii: Effect of pH, temperature, and sunlight on decomposition. Environ. Toxicol. 1999, 14, 155–161. [Google Scholar] [CrossRef]
- Santos, C.; Azevedo, J.; Campos, A.; Vasconcelos, V.; Pereira, A.L. Biochemical and growth performance of the aquatic macrophyte Azolla filiculoides to sub-chronic exposure to cylindrospermopsin. Ecotoxicology 2015, 24, 1848–1857. [Google Scholar] [CrossRef]
- Jámbrik, K.; Máthé, C.; Vasas, G.; Bácsi, I.; Surányi, G.; Gonda, S.; Borbély, G.; M.-Hamvas, M. Cylindrospermopsin inhibits growth and modulates protease activity in the aquatic plants Lemna minor L. and Wolffia arrhiza (L.) Horkel. Acta Biol. Hung. 2010, 61, 77–94. [Google Scholar] [CrossRef] [PubMed]
- Downing, S.; Banack, S.; Metcalf, J.; Cox, P.; Downing, T. Nitrogen starvation of cyanobacteria results in the production of β-N-methylamino-L-alanine. Toxicon 2011, 58, 187–194. [Google Scholar] [CrossRef]
- Cox, P.A.; Banack, S.A.; Murch, S.J.; Rasmussen, U.; Tien, G.; Bidigare, R.R.; Metcalf, J.S.; Morrison, L.F.; Codd, G.A.; Bergman, B. Diverse taxa of cyanobacteria produce β-N-methylamino-L-alanine, a neurotoxic amino acid. Proc. Natl. Acad. Sci. USA 2005, 102, 5074–5078. [Google Scholar] [CrossRef] [Green Version]
- Esterhuizen-Londt, M.; Downing, T. β-N-methylamino-L-alanine (BMAA) in novel South African cyanobacterial isolates. Ecotoxicol. Environ. Saf. 2008, 71, 309–313. [Google Scholar] [CrossRef]
- Réveillon, D.; Séchet, V.; Hess, P.; Amzil, Z. Production of BMAA and DAB by diatoms (Phaeodactylum tricornutum, Chaetoceros sp., Chaetoceros calcitrans and, Thalassiosira pseudonana) and bacteria isolated from a diatom culture. Harmful Algae 2016, 58, 45–50. [Google Scholar] [CrossRef]
- Jiang, L.; Eriksson, J.; Lage, S.; Jonasson, S.; Shams, S.; Mehine, M.; Ilag, L.L.; Rasmussen, U. Diatoms: A novel source for the neurotoxin BMAA in aquatic environments. PLoS ONE 2014, 9, e84578. [Google Scholar] [CrossRef]
- Lage, S.; Costa, P.R.; Moita, T.; Eriksson, J.; Rasmussen, U.; Rydberg, S.J. BMAA in shellfish from two Portuguese transitional water bodies suggests the marine dinoflagellate Gymnodinium catenatum as a potential BMAA source. Aquat. Toxicol. 2014, 152, 131–138. [Google Scholar] [CrossRef] [PubMed]
- Visser, C. Evaluation of Model Systems for the Study of Protein Association/Incorporation of Beta-Methylamino-L-Alanine (BMAA). Master’s Thesis, Nelson Mandela University, Port Elizabeth, South Africa, January 2011. [Google Scholar]
- Van Onselen, R.; Cook, N.A.; Phelan, R.R.; Downing, T.G. Bacteria do not incorporate β-N-methylamino-l-alanine into their proteins. Toxicon 2015, 102, 55–61. [Google Scholar] [CrossRef] [PubMed]
- Berntzon, L.; Erasmie, S.; Celepli, N.; Eriksson, J.; Rasmussen, U.; Bergman, B. BMAA inhibits nitrogen fixation in the cyanobacterium Nostoc sp. PCC 7120. Mar. Drugs 2013, 11, 3091–3108. [Google Scholar] [CrossRef] [Green Version]
- Downing, S.; Downing, T.G. The metabolism of the non-proteinogenic amino acid β-N-methylamino-L-alanine (BMAA) in the cyanobacterium Synechocystis PCC6803. Toxicon 2016, 115, 41–48. [Google Scholar] [CrossRef]
- Popova, A.A.; Rasmussen, U.; Semashko, T.A.; Govorun, V.M.; Koksharova, O.A. Stress effects of cyanotoxin β-methylamino-L-alanine (BMAA) on cyanobacterial heterocyst formation and functionality. Environ. Microbiol. Rep. 2018, 10, 369–377. [Google Scholar] [CrossRef] [PubMed]
- Lürling, M.; Faassen, E.J.; Van Eenennaam, J.S. Effects of the cyanobacterial neurotoxin β-N-methylamino-L-alanine (BMAA) on the survival, mobility and reproduction of Daphnia magna. J. Plankton Res. 2011, 33, 333–342. [Google Scholar] [CrossRef] [Green Version]
- Faassen, E.J.; García-Altares, M.; e Mello, M.M.; Lürling, M. Trans generational effects of the neurotoxin BMAA on the aquatic grazer Daphnia magna. Aquat. Toxicol. 2015, 168, 98–107. [Google Scholar] [CrossRef] [PubMed]
- Esterhuizen-Londt, M.; Wiegand, C.; Downing, T.G. β-N-methylamino-L-alanine (BMAA) uptake by the animal model, Daphnia magna and subsequent oxidative stress. Toxicon 2015, 100, 20–26. [Google Scholar] [CrossRef] [PubMed]
- Al-Sammak, M.A.; Hoagland, K.D.; Cassada, D.; Snow, D.D. Co-occurrence of the cyanotoxins BMAA, DABA and anatoxin-a in Nebraska reservoirs, fish, and aquatic plants. Toxins 2014, 6, 488–508. [Google Scholar] [CrossRef] [Green Version]
- Esterhuizen-Londt, M.; Pflugmacher, S.; Downing, T. β-N-Methylamino-L-alanine (BMAA) uptake by the aquatic macrophyte Ceratophyllum demersum. Ecotoxicol. Environ. Saf. 2011, 74, 74–77. [Google Scholar] [CrossRef]
- Esterhuizen-Londt, M.; Pflugmacher, S.; Downing, T. The effect of β-N-methylamino-L-alanine (BMAA) on oxidative stress response enzymes of the macrophyte Ceratophyllum demersum. Toxicon 2011, 57, 803–810. [Google Scholar] [CrossRef]
- Downing, S.; Esterhuizen-Londt, M.; Downing, T.G. β-N-methylamino-L-alanine (BMAA) metabolism in the aquatic macrophyte Ceratophyllum demersum. Ecotoxicol. Environ. Saf. 2015, 120, 88–92. [Google Scholar] [CrossRef]
- Contardo-Jara, V.; Kuehn, S.; Pflugmacher, S. Single and combined exposure to MC-LR and BMAA confirm suitability of Aegagropila linnaei for use in green liver systems®–A case study with cyanobacterial toxins. Aquat. Toxicol. 2015, 165, 101–108. [Google Scholar] [CrossRef] [PubMed]
- Lemaire, V.; Brusciotti, S.; van Gremberghe, I.; Vyverman, W.; Vanoverbeke, J.; De Meester, L. Genotype× genotype interactions between the toxic cyanobacterium Microcystis and its grazer, the waterflea Daphnia. Evol. Appl. 2012, 5, 168–182. [Google Scholar] [CrossRef] [PubMed]
- Dziallas, C.; Grossart, H.P. Temperature and biotic factors influence bacterial communities associated with the cyanobacterium Microcystis sp. Environ. Microbiol. 2011, 13, 1632–1641. [Google Scholar] [CrossRef] [PubMed]
- Wiltsie, D.; Schnetzer, A.; Green, J.; Vander Borgh, M.; Fensin, E. Algal blooms and cyanotoxins in Jordan Lake, North Carolina. Toxins 2018, 10, 92. [Google Scholar] [CrossRef] [Green Version]
- Graham, J.L.; Loftin, K.A.; Meyer, M.T.; Ziegler, A.C. Cyanotoxin mixtures and taste-and-odor compounds in cyanobacterial blooms from the Midwestern United States. Environ. Sci. Technol. 2010, 44, 7361–7368. [Google Scholar] [CrossRef] [PubMed]
- Christensen, V.G.; Maki, R.P.; Stelzer, E.A.; Norland, J.E.; Khan, E. Phytoplankton community and algal toxicity at a recurring bloom in Sullivan Bay, Kabetogama Lake, Minnesota, USA. Sci. Rep. 2019, 9, 1–11. [Google Scholar] [CrossRef]
- Esterhuizen-Londt, M.; Von Schnehen, M.; Kühn, S.; Pflugmacher, S. Oxidative stress responses in the animal model, Daphnia pulex exposed to a natural bloom extract versus artificial cyanotoxin mixtures. Aquat. Toxicol. 2016, 179, 151–157. [Google Scholar] [CrossRef] [PubMed]
- Briand, E.; Reubrecht, S.; Mondeguer, F.; Sibat, M.; Hess, P.; Amzil, Z.; Bormans, M. Chemically mediated interactions between Microcystis and Planktothrix: Impact on their growth, morphology and metabolic profiles. Environ. Microbiol. 2019, 21, 1552–1566. [Google Scholar] [CrossRef] [PubMed] [Green Version]


Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Omidi, A.; Pflugmacher, S.; Kaplan, A.; Kim, Y.J.; Esterhuizen, M. Reviewing Interspecies Interactions as a Driving Force Affecting the Community Structure in Lakes via Cyanotoxins. Microorganisms 2021, 9, 1583. https://doi.org/10.3390/microorganisms9081583
Omidi A, Pflugmacher S, Kaplan A, Kim YJ, Esterhuizen M. Reviewing Interspecies Interactions as a Driving Force Affecting the Community Structure in Lakes via Cyanotoxins. Microorganisms. 2021; 9(8):1583. https://doi.org/10.3390/microorganisms9081583
Chicago/Turabian StyleOmidi, Azam, Stephan Pflugmacher, Aaron Kaplan, Young Jun Kim, and Maranda Esterhuizen. 2021. "Reviewing Interspecies Interactions as a Driving Force Affecting the Community Structure in Lakes via Cyanotoxins" Microorganisms 9, no. 8: 1583. https://doi.org/10.3390/microorganisms9081583