
Culture Degeneration Reduces Sex-Related Gene Expression, Alters Metabolite Production and Reduces Insect Pathogenic Response in Cordyceps militaris

 ,
,
Abstract
:1. Introduction
2. Material and Methods
2.1. Fungal Culture Media
2.2. Reverse Transcription and Quantitative PCR (RT-qPCR)
2.3. Liquid Chromatography-Mass Spectrometry (LC-MS)
2.3.1. Fungal Sample Extraction
2.3.2. Liquid Chromatography
2.3.3. Mass Spectrometry
2.4. Galleria mellonella Caterpillar Treatments
2.5. Statistical Analysis
3. Results
3.1. Comparing Parental Control and Degenerated CM2 Strains
3.1.1. Growth Rates of the Parental Control and Degenerated C. militaris Strains
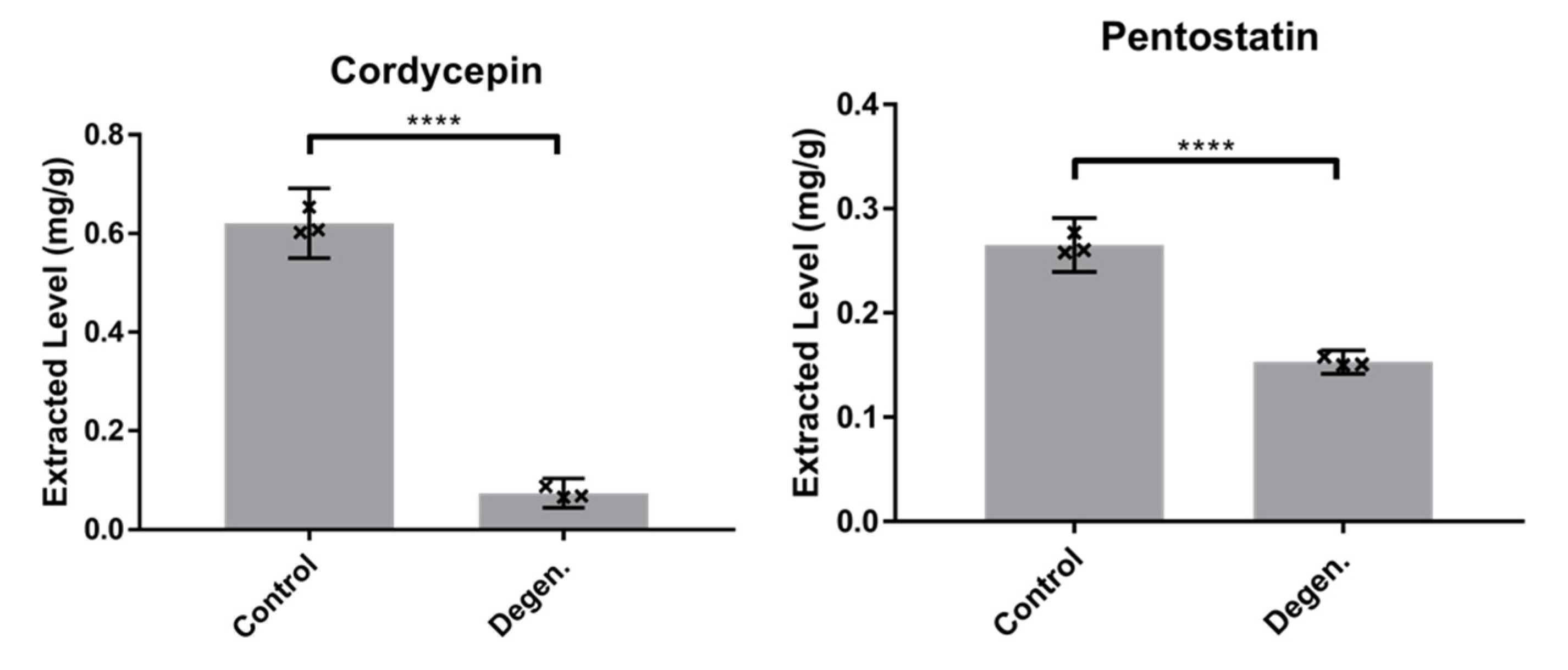
3.1.2. Comparison of Cordycepin and Pentostatin Levels in Parental Control and Degenerated C. militaris Strains
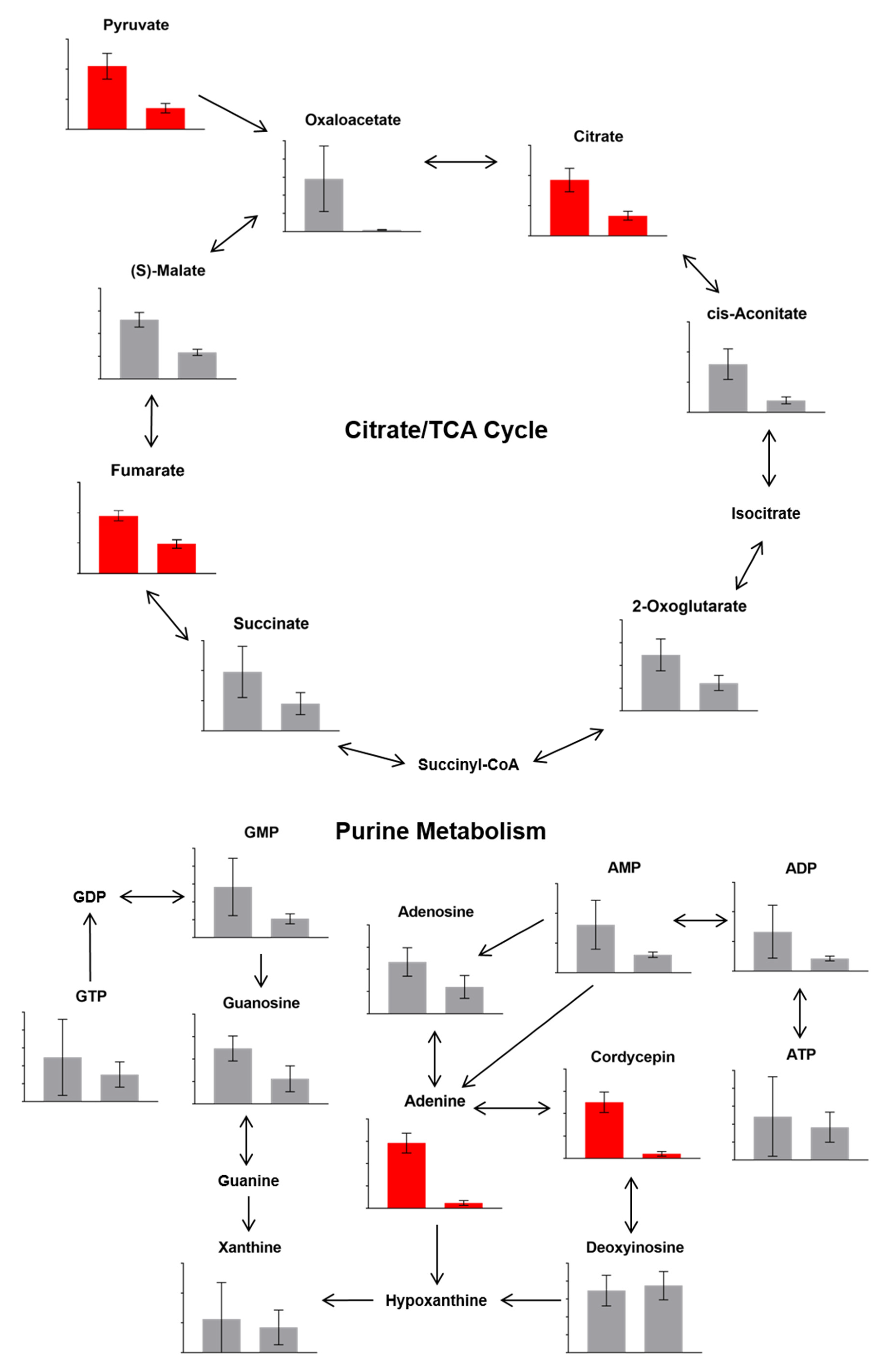
3.1.3. Differences in Metabolites Detected
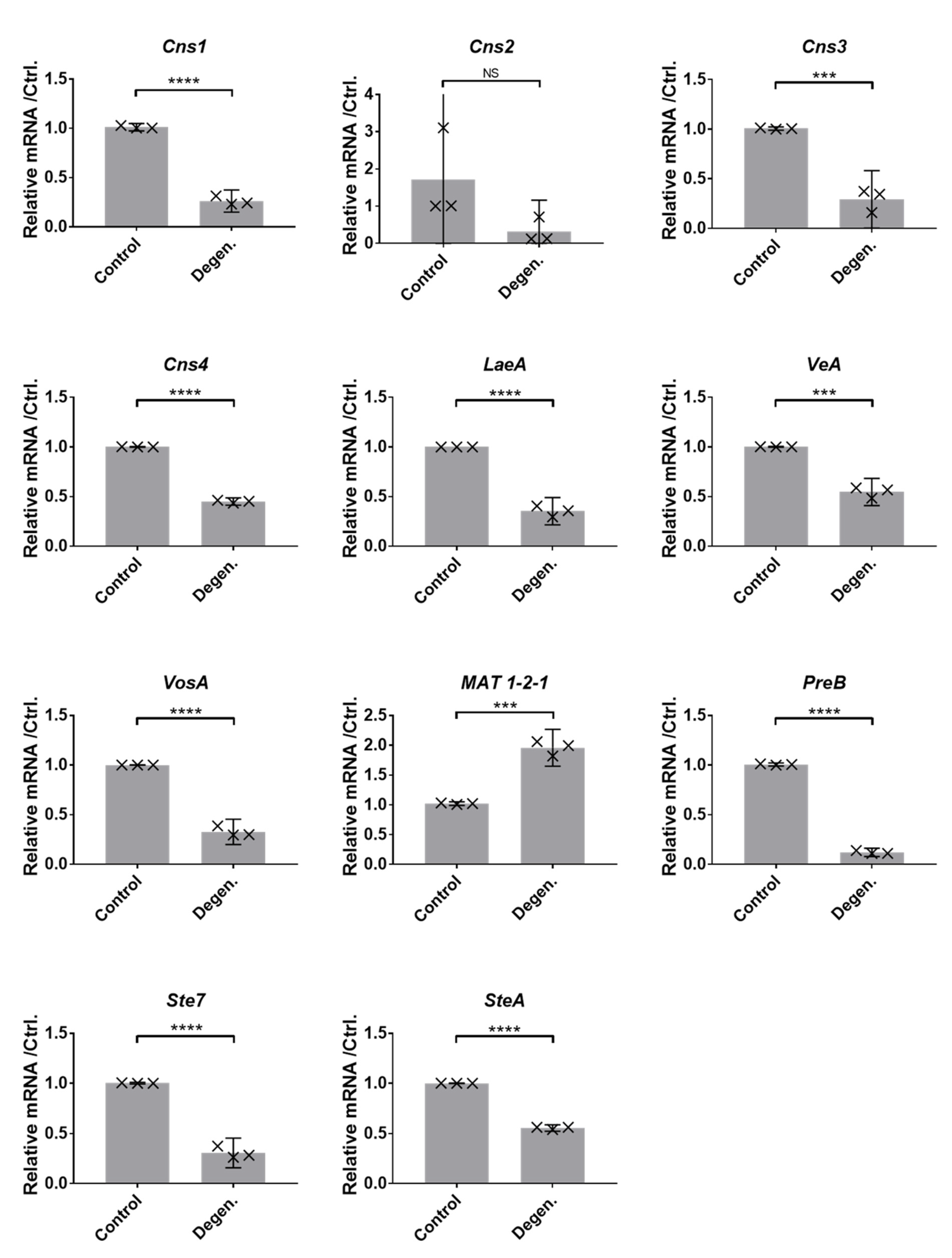
3.1.4. Gene Expression Differences
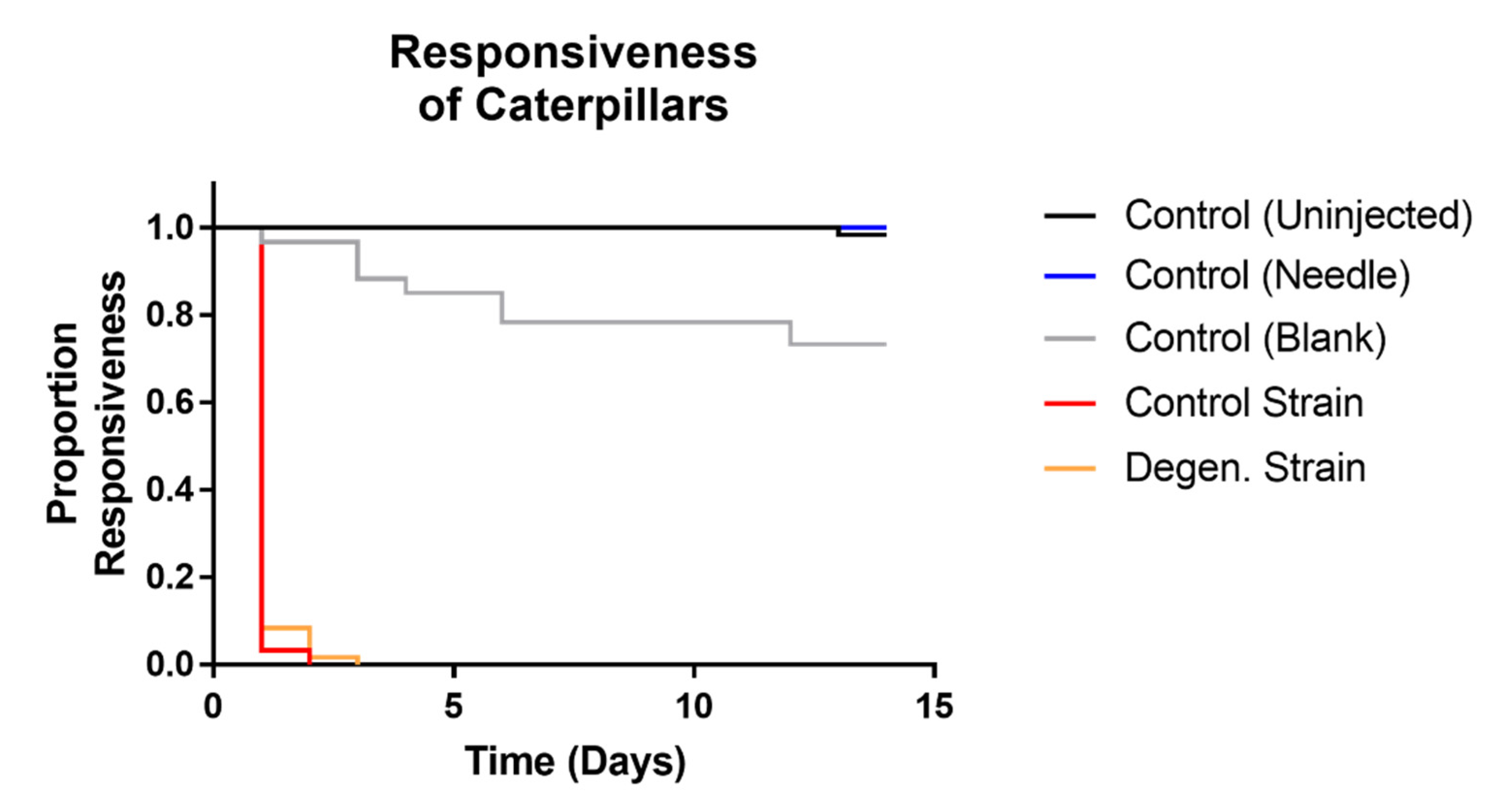
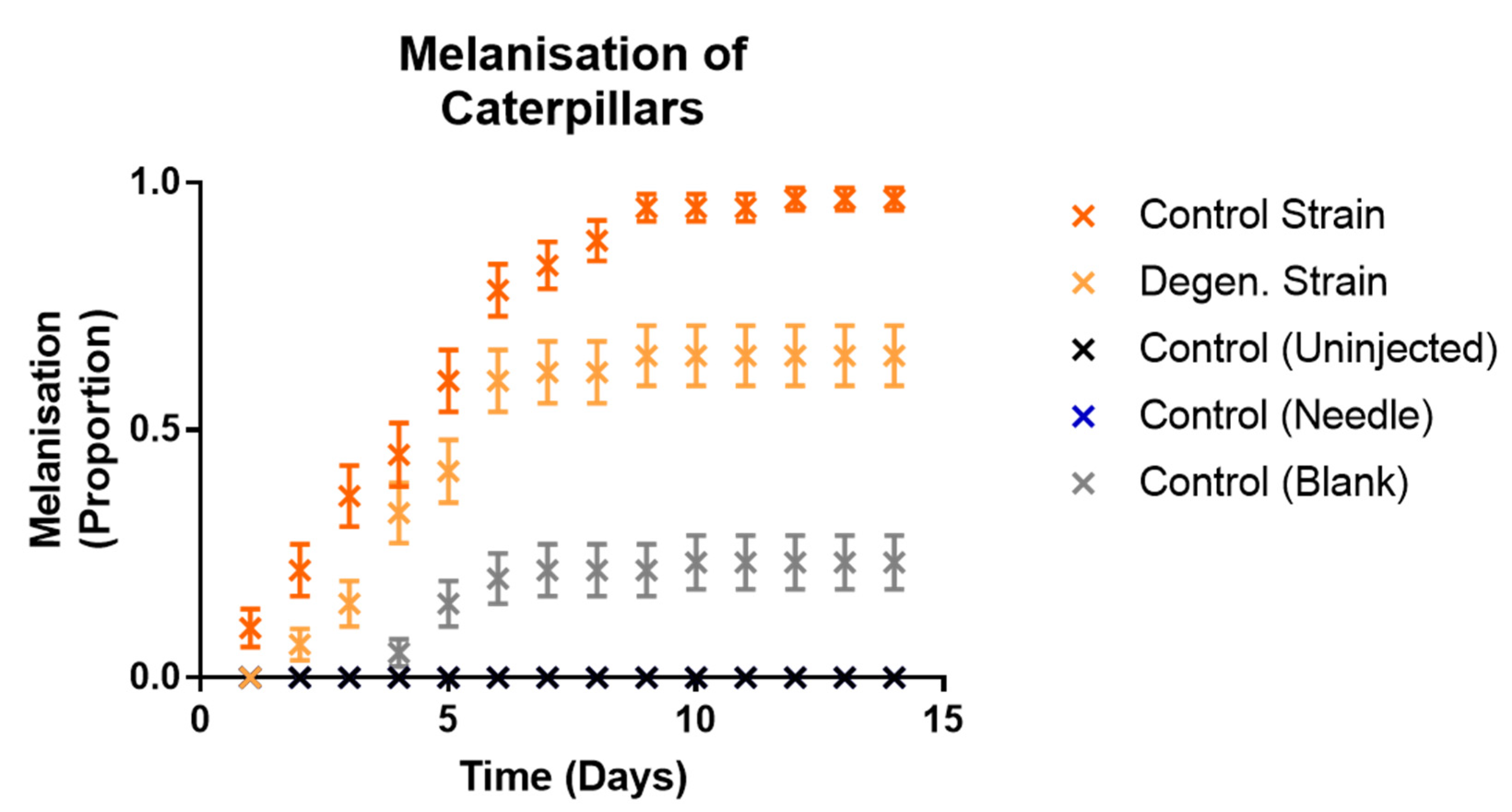
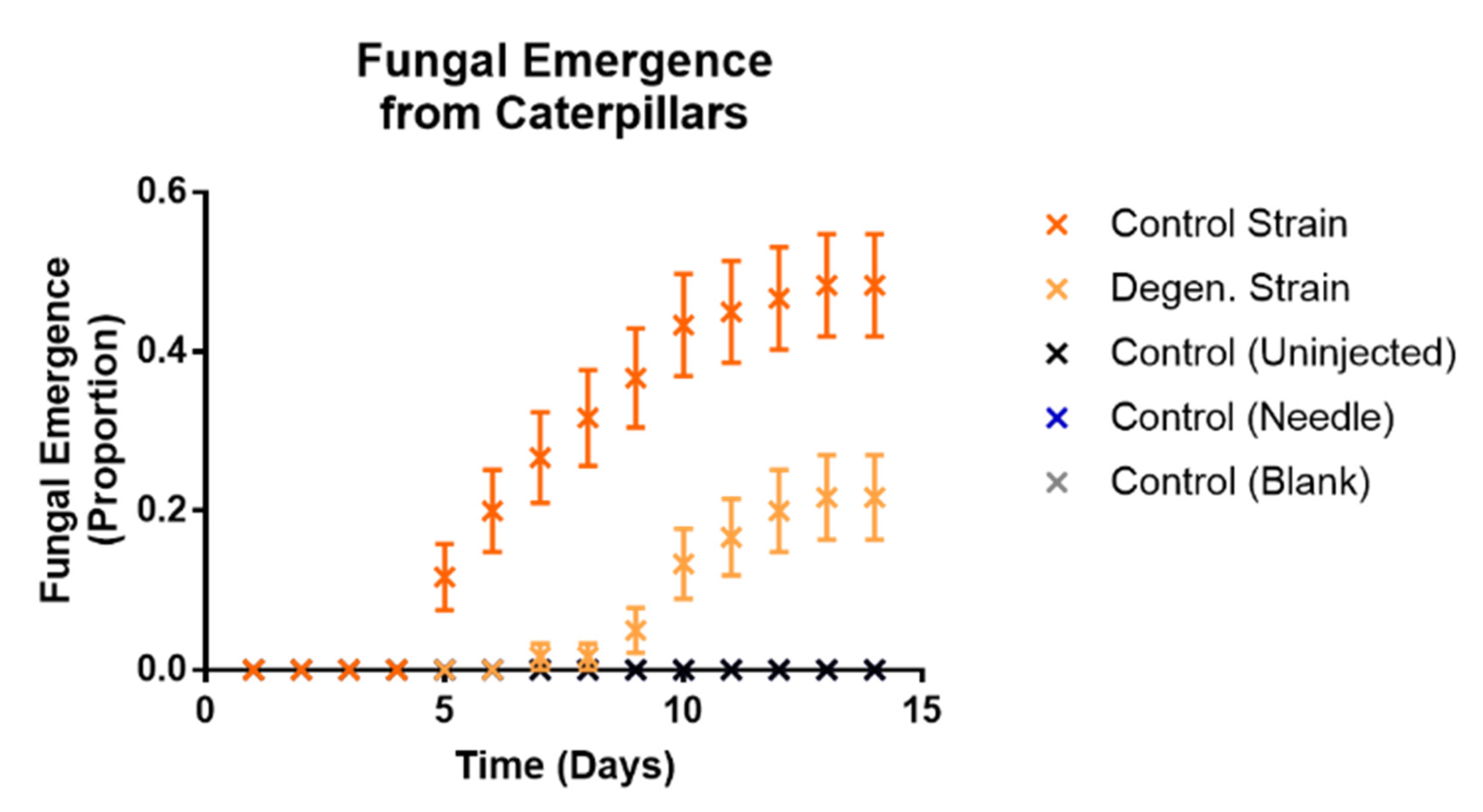
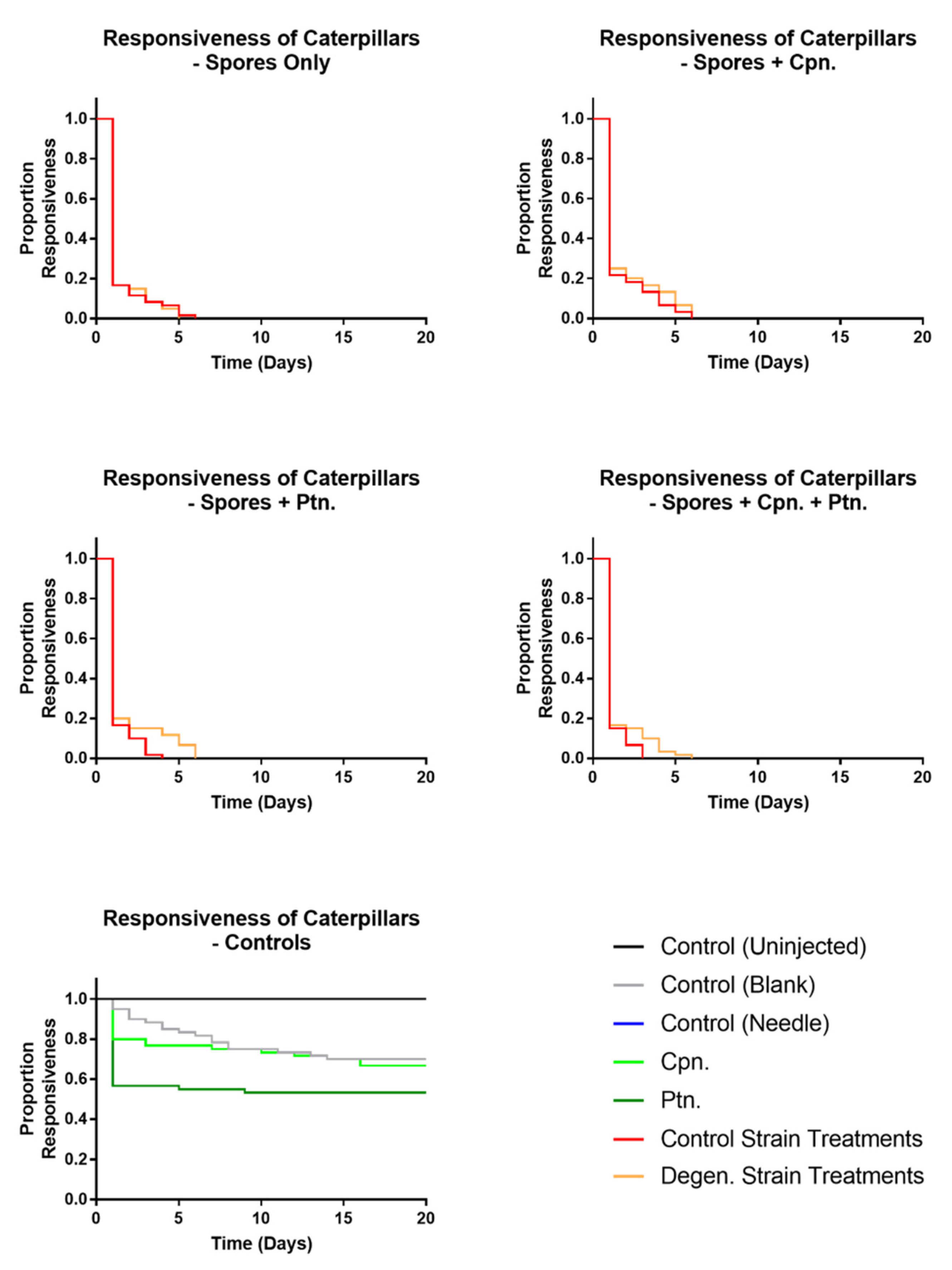
3.2. Effects on Galleria mellonella Caterpillars
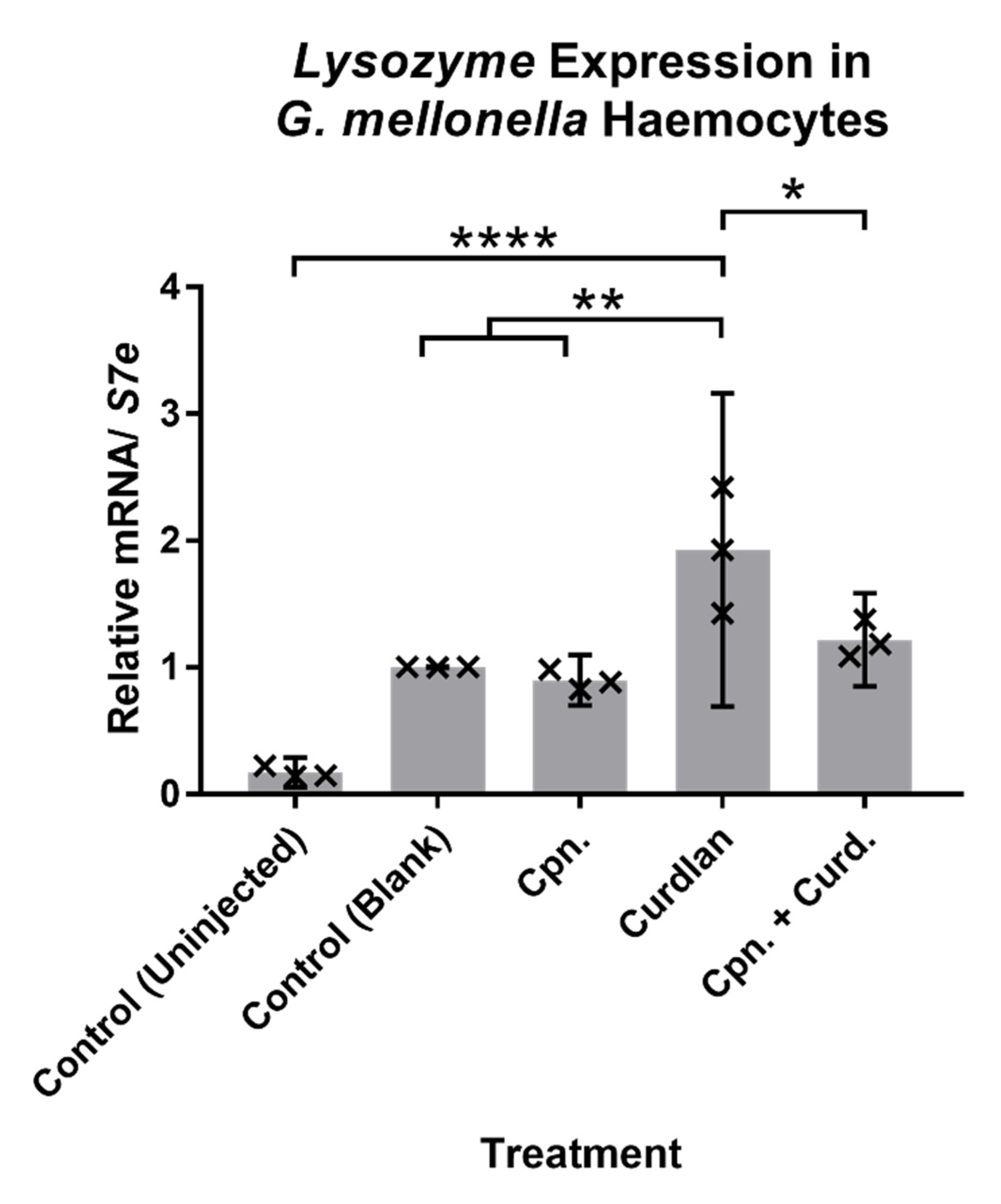
3.2.1. Immune-Related Gene Expression in Response to Cordycepin in Caterpillars
3.2.2. Effects of Fungal Injections on Caterpillars
4. Discussion
Supplementary Materials
Author Contributions
Funding
Data Availability Statement
Conflicts of Interest
References
- Shrestha, B.; Zhang, W.; Zhang, Y.; Liu, X. The medicinal fungus Cordyceps militaris: Research and development. Mycol. Prog. 2012, 11, 559–614. [Google Scholar] [CrossRef]
- Cunningham, K.G.; Manson, W.; Spring, F.S. Cordycepin, a metabolic product isolated from cultures of Cordyceps militaris (Linn.) Link. Nature 1950, 166, 949. [Google Scholar] [CrossRef] [PubMed]
- Kondrashov, A.; Meijer, H.A.; Barthet-Barateig, A.; Parker, H.N.; Khurshid, A.; Tessier, S.; Sicard, M.; Knox, A.J.; Pang, L.; de Moor, C.H. Inhibition of polyadenylation reduces inflammatory gene induction. RNA 2012, 18, 2236–2250. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ashraf, S.; Radhi, M.; Gowler, P.; Burston, J.J.; Gandhi, R.D.; Thorn, G.J.; Piccinini, A.M.; Walsh, D.A.; Chapman, V.; de Moor, C.H. The polyadenylation inhibitor cordycepin reduces pain, inflammation and joint pathology in rodent models of osteoarthritis. Sci. Rep. 2019, 9, 4696. [Google Scholar] [CrossRef]
- Wong, Y.Y.; Moon, A.; Duffin, R.; Barthet-Barateig, A.; Meijer, H.A.; Clemens, M.J.; de Moor, C.H. Cordycepin Inhibits Protein Synthesis and Cell Adhesion through Effects on Signal Transduction. J. Biol. Chem. 2010, 285, 2610–2621. [Google Scholar] [CrossRef] [Green Version]
- Xia, Y.; Luo, F.; Shang, Y.; Chen, P.; Lu, Y.; Wang, C. Fungal cordycepin biosynthesis is coupled with the production of the safeguard molecule pentostatin. Cell Chem. Biol. 2017, 24, 1479–1489. [Google Scholar] [CrossRef] [Green Version]
- Boomsma, J.J.; Jensen, A.B.; Meyling, N.V.; Eilenberg, J. Evolutionary interaction networks of insect pathogenic fungi. Annu. Rev. Entomol. 2014, 59, 467–485. [Google Scholar] [CrossRef]
- Kim, J.R.; Yeon, S.H.; Kimn, H.S.; Ahn, Y.J. Larvicidal activity against Plutella xylostella of cordycepin from the fruiting body of Cordyceps militaris. Pest Manag. Sci. 2002, 58, 713–717. [Google Scholar] [CrossRef]
- Schmid-Hempel, P. Immune defence, parasite evasion strategies and their relevance for “macroscopic phenomea” such as virulence. Philos. Trans. R. Soc. B 2009, 364, 85–98. [Google Scholar] [CrossRef] [Green Version]
- Wellham, P.A.D.; Kim, D.H.; Brock, M.; de Moor, C.H. Coupled biosynthesis of cordycepin and pentostatin in Cordyceps militaris: Implications for fungal biology and medicinal natural products. Annal. Transl. Med. 2019, 7 (Suppl. 3), S85. [Google Scholar] [CrossRef]
- Woolley, V.C.; Teakle, G.R.; Prince, G.; de Moor, C.H.; Chandler, D. Cordycepin, a metabolite of Cordyceps militaris, reduces immune-related gene expression in insects. J. Invertebr. Pathol. 2020, 177, 107480. [Google Scholar] [CrossRef]
- Kato, T.; Nishimura, K.; Suparmin, A.; Ikeo, K.; Park, E.Y. Effects of cordycepin in Cordyceps militaris during its injection to silkworm larvae. Microorganisms 2021, 9, 681. [Google Scholar] [CrossRef]
- Shrestha, B.; Han, S.K.; Yoon, H.S.; Sung, J.M. Morphological characteristics of conidiogenesis in Cordyceps militaris. J. Mycobiol. 2005, 33, 69–76. [Google Scholar] [CrossRef] [Green Version]
- Gago, S.; Garcia-Rodas, R.; Cuesta, I.; Mellado, E.; Alastruey-Izquierdo, A. Candida parapsilosis, Candida orthopsilosis, and Candida metapsilosis virulence in the non-conventional host Galleria mellonella. Virulence 2014, 5, 278–285. [Google Scholar] [CrossRef] [Green Version]
- Scorzoni, L.; De Paula e Silva, A.C.A.; Singulani, J.D.L.; Leite, F.S.; De Oliveira, H.C.; Moraes da Silva, R.A.; Fusco-Almeida, A.M.; Mendes-Giannini, M.J.S. Comparison of virulence between Paracoccidioides brasiliensis and Paracoccidioides lutzii using Galleria mellonella as a host model. Virulence 2015, 6, 766–776. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Beekman, C.N.; Meckler, L.; Kim, E.; Bennett, R.J. Galleria mellonella as an insect model for P. destructans, the cause of white-nose syndrome in bats. PLoS ONE 2018, 13, e0201915. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Maurer, E.; Hörtnagl, C.; Lackner, M.; Grässle, D.; Naschberger, V.; Moser, P.; Segal, E.; Semis, M.; Lass-Flörl, C.; Binder, U. Galleria mellonella as a model system to study virulence potential of mucormycetes and evaluation of antifungal treatment. Med. Mycol. 2019, 57, 351–362. [Google Scholar] [CrossRef] [Green Version]
- Ramarao, N.; Nielsen-Leroux, C.; Lereclus, D. The insect Galleria mellonella as a powerful infection model to investigate bacterial pathogenesis. J. Vis. Exp. 2012, 70, e4392. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jorjão, A.L.; Oliveira, L.D.; Scorzoni, L.; Figueiredo-Godoi, L.M.A.; Cristina APrata, M.; Jorge, A.O.C.; Junqueira, J.C. From moths to caterpillars: Ideal conditions for Galleria mellonella rearing for in vivo microbiological studies. Virulence 2018, 9, 383–389. [Google Scholar] [CrossRef] [Green Version]
- Lange, A.; Beier, S.; Huson, D.H.; Parusel, R.; Iglauer, F.; Frick, J.S. Genome sequence of Galleria mellonella (greater wax moth). Genome Announc. 2018, 6, e01220-17. [Google Scholar] [CrossRef] [Green Version]
- Altincicek, B.; Vilcinskas, A. Metamorphosis and collagen-IV-fragments stimulate innate immune response in the greater wax moth, Galleria mellonella. Dev. Comp. Immun. 2006, 30, 1108–1118. [Google Scholar] [CrossRef] [PubMed]
- Wojda, I.; Jakubowicz, T. Humoral immune response upon mild heat-shock conditions in Galleria mellonella. J. Insect Phys. 2007, 53, 1134–1144. [Google Scholar] [CrossRef]
- Wojda, I.; Kowalski, P.; Jakubosicz, T. Hummoral immune response of Galleria mellonella larvae after infection by Beauveria bassiana under optimal and heat-schock conditions. J. Insect Phys. 2009, 6, 525–531. [Google Scholar] [CrossRef] [PubMed]
- Sun, S.J.; Deng, C.H.; Zhang, L.Y.; Hu, K.H. Molecular analysis and biochemical characteristics of degenerated strains of Cordyceps militaris. Arch. Microbiol. 2017, 199, 939–944. [Google Scholar] [CrossRef] [PubMed]
- Yin, J.; Xin, X.; Weng, Y.; Gui, Z. Transcriptome-wide analysis reveals the progress of Cordyceps militaris subculture degeneration. PLoS ONE 2017, 12, E0182679. [Google Scholar] [CrossRef] [Green Version]
- Safavi, S.A. Attenuation of the entomopathogenic fungus Beauveria bassiana following serial in vitro transfers. Biologia 2012, 67, 1062–1068. [Google Scholar] [CrossRef]
- Ansari, M.A.; Butt, T.M. Effects of successive subculturing on stability, virulence, conidial yield, germination and shelf-life of entomopathogenic fungi. J. Appl. Microbiol. 2011, 110, 1460–1469. [Google Scholar] [CrossRef]
- White, T.J.; Bruns, T.; Lee, S.J.W.T.; Taylor, J. Amplification and direct sequencing of fungal ribosomal RNA genes for phylogenetics. In PCR Protocols: A Guide to Methods and Applications; Academic Press: New York, NY, USA, 1990; pp. 315–322. [Google Scholar]
- Schatschneider, S.; Abdelrazig, S.; Safo, L.; Henstra, A.M.; Millat, T.; Kim, D.H.; Winzer, K.; Minton, N.P.; Barrett, D.A. Quantitative isotope-dilution high-resolution-mass-spectrometry analysis of multiple intracellular metabolites in Clostridium autoethanogenum with uniformly 13C-labeled standards derived from Spirulina. Anal. Chem. 2018, 90, 4470–4477. [Google Scholar] [CrossRef]
- Want, E.J. Global metabolic profiling procedures for urine using UPLC-MS. Nat. Protoc. 2010, 5, 1005–1018. [Google Scholar] [CrossRef] [PubMed]
- Tautenhahn, R.; Bottcher, C.; Neumann, S. Highly sensitive feature detection for high resolution LC/MS. BMC Bioinform. 2008, 9, 504. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Scheltema, R.A.; Jankevics, A.; Jansen, R.C.; Swertz, M.A.; Breitling, R. PeakML/mzMatch: A File Format, Java Library, R Library, and Tool-Chain for Mass Spectrometry Data Analysis. Anal. Chem. 2011, 83, 2786–2793. [Google Scholar] [CrossRef] [Green Version]
- Creek, D.J.; Jankevics, A.; Burgess KE, V.; Breitling, R.; Barrett, M.P. IDEOM: An Excel interface for analysis of LC-MS-based metabolomics data. Bioinformatics 2012, 28, 1048–1049. [Google Scholar] [CrossRef]
- Sumner, L.W.; Amberg, A.; Barrett, D.; Beale, M.H.; Beger, R.; Daykin, C.A.; Fan, T.W.M.; Fiehn, O.; Goodacre, R.; Griffin, J.L.; et al. Proposed minimum reporting standards for chemical analysis. Metabolomics 2007, 3, 211–221. [Google Scholar] [CrossRef] [Green Version]
- Sumner, L.W.; Lei, Z.; Nikolau, B.J.; Saito, K.; Roessner, U.; Trengove, R. Proposed quantitative and alphanumeric metabolite identification metrics. Metabolomics 2014, 10, 1047–1049. [Google Scholar] [CrossRef]
- Chambers, M.C.; Lightfield, K.L.; Schneider, D.S. How the fly balances its ability to combat different pathogens. PLoS Pathog. 2012, 8, e1002970. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cereneus, L.; Lee, B.L.; Soderhall, K. The proPO-system: Pros and cons for its role in invertebrate immunity. Trends in Immunol. 2008, 29, 263–271. [Google Scholar] [CrossRef] [PubMed]
- Lu, A.; Zhang, Q.; Zhang, J.; Yang, B.; Wu, K.; Xie, W.; Luan, Y.X.; Ling, E. Insect prophenoloxidase: The view beyond immunity. Front. Physiol. 2014, 5, 252. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chandler, D. Basic and applied research on entomopathogenic fungi. In Microbial Agents of Insect and Mite Pests: From Theory to Practice; Lacey, L., Ed.; Academic Press: New York, NY, USA, 2016; pp. 42–60. [Google Scholar]
- Dubovskiy, I.M.; Whitten, M.A.; Kryukov, V.Y.; Yaroslavtseva, O.N.; Grizanova, E.V.; Greig, C.; Mukherjee, K.; Vilcinskas, A.; Mitkovets, P.V.; Glupov, V.V.; et al. More than a colour change: Insect melanism, disease resistance and fecundity. P. Roy. Soc. Lond. B. Bio. 2013, 280, 20130584. [Google Scholar] [CrossRef] [PubMed]
- Jin, L.H.; Shim, J.; Yoon, J.S.; Kim, B.; Kim, J.; Kim-Ha, J.; Kim, Y.J. Identification and functional analysis of antifungal immune response genes in Drosophila. PLoS Pathog. 2008, 4, e1000168. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bayram, O.; Braus, G.H. Coordination of secondary metabolism and development in fungi: The velvet family of regulatory proteins. FEMS Microbiol. Rev. 2012, 36, 1–24. [Google Scholar] [CrossRef] [Green Version]
- Sarikaya Bayram, Ö.; Bayram, Ö.; Valerius, O.; Park, H.S.; Irniger, S.; Gerke, J.; Ni, M.; Han, K.H.; Yu, J.H.; Braus, G.H. Laea control of velvet family regulatory proteins for light-dependent development and fungal cell-type specificity. PLoS Genet. 2010, 6, e1001226. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kryukov, V.Y.; Kosman, E.; Tomilova, O.; Polenogova, O.; Rotskaya, U.; Tyurin, M.; Alikina, T.; Yaroslavtseva, O.; Kabilov, M.; Glupov, V. Interplay between fungal infection and bacterial associates in the wax moth Galleria mellonella under different temperature conditions. J. Fungi. 2020, 6, 170. [Google Scholar] [CrossRef] [PubMed]














{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
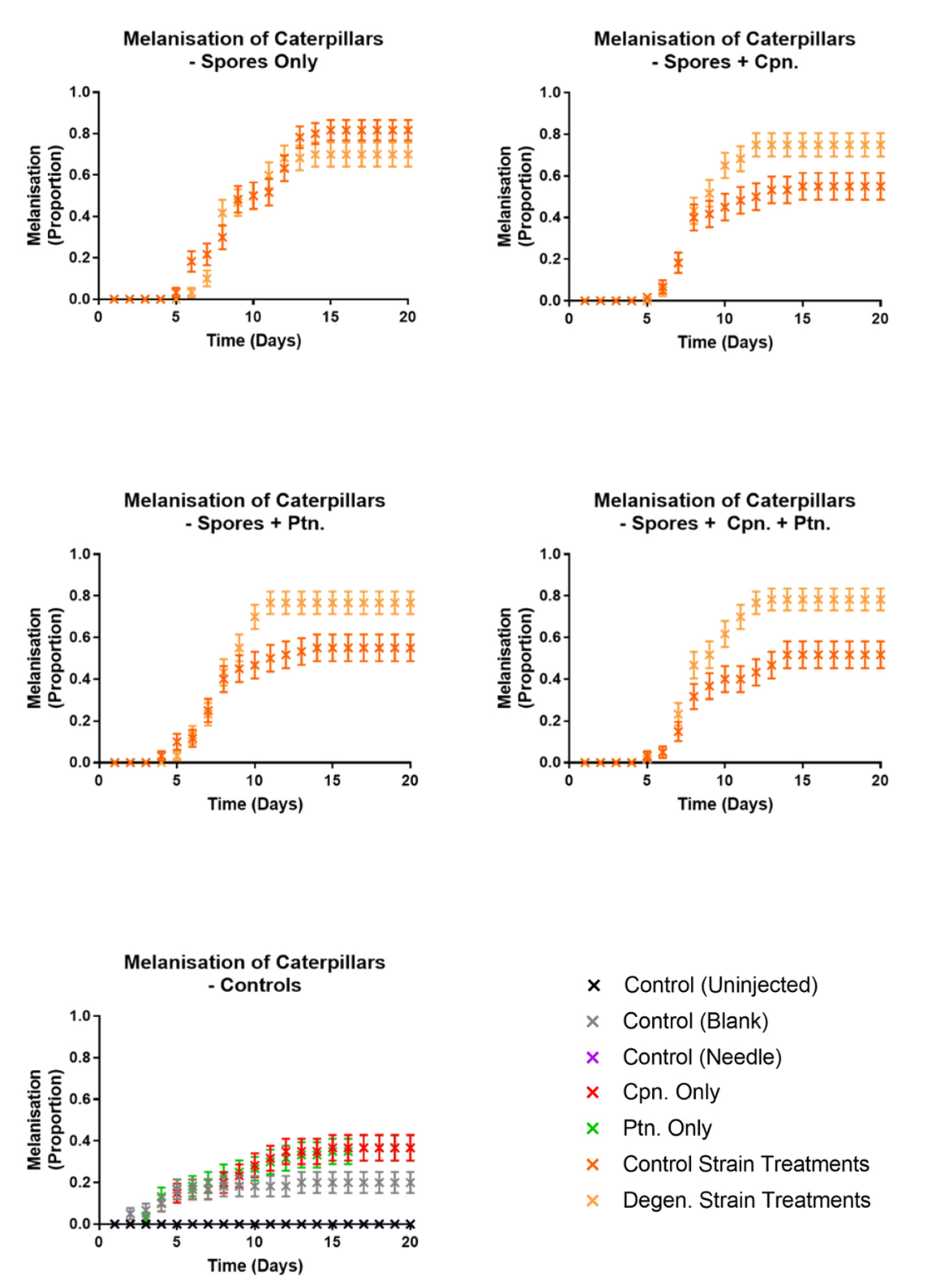{kind=link}
{kind=link}
| Gene | Sequences (5′-3′) | Species | Primer Reference |
|---|---|---|---|
| Actin | CGAAGACGTTGCCGCTCTGG CGGAGGCTGAGAATGCC | C. militaris | This study |
| Calmodulin | GCTTGCGCCCTCTCGCTGC CTCTGACTCGGAAGGGTTCTGGCC | C. militaris | This study |
| Glyceraldehyde-3-Phosphate Dehydrogenase (GAPDH) | CGTCAAGGTTGGCATCAACGG GCCGTTGACGACGAGATCG | C. militaris | This study |
| Ribosomal Protein S3 (rps3) | CCTTTCGCCCAAGAATAATTTAG GTTGAATGTAAAGATGTTTTTTGTAATCC | C. militaris | This study |
| Cordycepin Synthesis 1 (Cns1) | GCTTATCCGACTACATTTCCATCC CCAGCCGCTCCAGGTGC | C. militaris | This study |
| Cordycepin Synthesis 2 (Cns2) | CGCCGGTGTCCTCCAGAC CGAGGCGTGTGACACGC | C. militaris | This study |
| Cordycepin Synthesis 3 (Cns3) | CGAGTCAACCGCCTACACC GTAGGACTGGGGCAGCGG | C. militaris | This study |
| Cordycepin Synthesis 4 (Cns4) | GCCGGACAAAGAGAAACGAC CCAAGAGCATCTCTCCCGG | C. militaris | This study |
| Loss of aflR-Expression A (LaeA) | GGCTGTCGATCTGAACAAAATCC CGAGGGTAGCCAATGAATTTCGACC | C. militaris | This study |
| Velvet A (VeA) | CCAGTGCCCAGTGCCAGTTG GCAATGGGCGAGGGCGAG | C. militaris | This study |
| Variability of Spores A (VosA) | CTCTTCATACACATCACCAAAGGC CTGTTGGCGCACCTCAAGG | C. militaris | This study |
| Mating type alpha pheromone receptor (PreB) | CGTCGTTTGACCGCTTCGC CGACGCAGAGCGCGAG | C. militaris | This study |
| MAT 1-2-1 | CTCAGTATCGACGGTCTCATCTACC CCAGTGCCGGACATCAAATGTCG | C. militaris | This study |
| MAP kinase kinase Ste7 (Ste7) | GATGGTCAACTCGAGATCGGG GTTGACAATGTAATCCGAGTGACAG | C. militaris | This study |
| Transcription Factor SteA (SteA) | GCTTTCTGCTCCCTACCGG GAAAGTCGAGAAACTGGCTCTTG | C. militaris | This study |
| Ribosomal Gene S7e (S7e) | TCCCAACTCTTGACCGACGA AGTGGTTGCGCCATCCATAC | G. mellonella | [22] |
| Lysozyme (Lyso) | ATGTGCCAATGCCCAAGTTG GTGGCTAGGCTTGGGAAGAAT | G. mellonella | [21] |
| Gallerimycin (Gale) | TATCATTGGCCTTCTTGGCTG GCACTCGTAAAATACACATCCGG | G. mellonella | [22] |
| Galiomicin (Gali) | TCGTATCGTCACCGCAAAATG GCCGCAATGACCACCTTTATA | G. mellonella | [23] |
| Insect metalloproteinase inhibitor (IMPI) | AGATGGCTATGCAAGGGATG AGGACCTGTGCAGCATTTCT | G. mellonella | [21] |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Wellham, P.A.D.; Hafeez, A.; Gregori, A.; Brock, M.; Kim, D.-H.; Chandler, D.; de Moor, C.H. Culture Degeneration Reduces Sex-Related Gene Expression, Alters Metabolite Production and Reduces Insect Pathogenic Response in Cordyceps militaris. Microorganisms 2021, 9, 1559. https://doi.org/10.3390/microorganisms9081559
Wellham PAD, Hafeez A, Gregori A, Brock M, Kim D-H, Chandler D, de Moor CH. Culture Degeneration Reduces Sex-Related Gene Expression, Alters Metabolite Production and Reduces Insect Pathogenic Response in Cordyceps militaris. Microorganisms. 2021; 9(8):1559. https://doi.org/10.3390/microorganisms9081559
Chicago/Turabian StyleWellham, Peter A. D., Abdul Hafeez, Andrej Gregori, Matthias Brock, Dong-Hyun Kim, David Chandler, and Cornelia H. de Moor. 2021. "Culture Degeneration Reduces Sex-Related Gene Expression, Alters Metabolite Production and Reduces Insect Pathogenic Response in Cordyceps militaris" Microorganisms 9, no. 8: 1559. https://doi.org/10.3390/microorganisms9081559