
Acinetobacter tandoii ZM06 Assists Glutamicibacter nicotianae ZM05 in Resisting Cadmium Pressure to Preserve Dipropyl Phthalate Biodegradation



{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
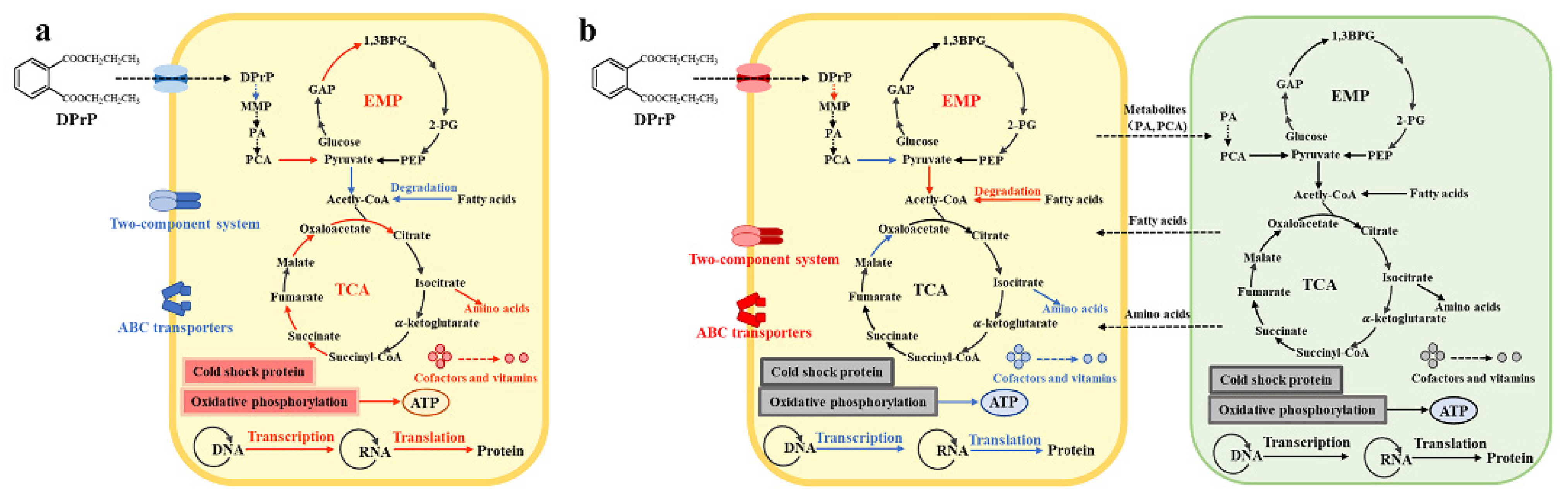
Abstract
:1. Introduction
2. Materials and Methods
2.1. Chemicals and Media
2.2. Isolation and Identification of the DBP-Degrading Bacterium and Cooperative Bacterium
2.3. Monoculture and Coculture Experiments
2.4. Sample Preparation and Analysis
2.5. cDNA Library Construction and RNA-Seq
2.6. RNA-Seq Data Analysis
3. Results
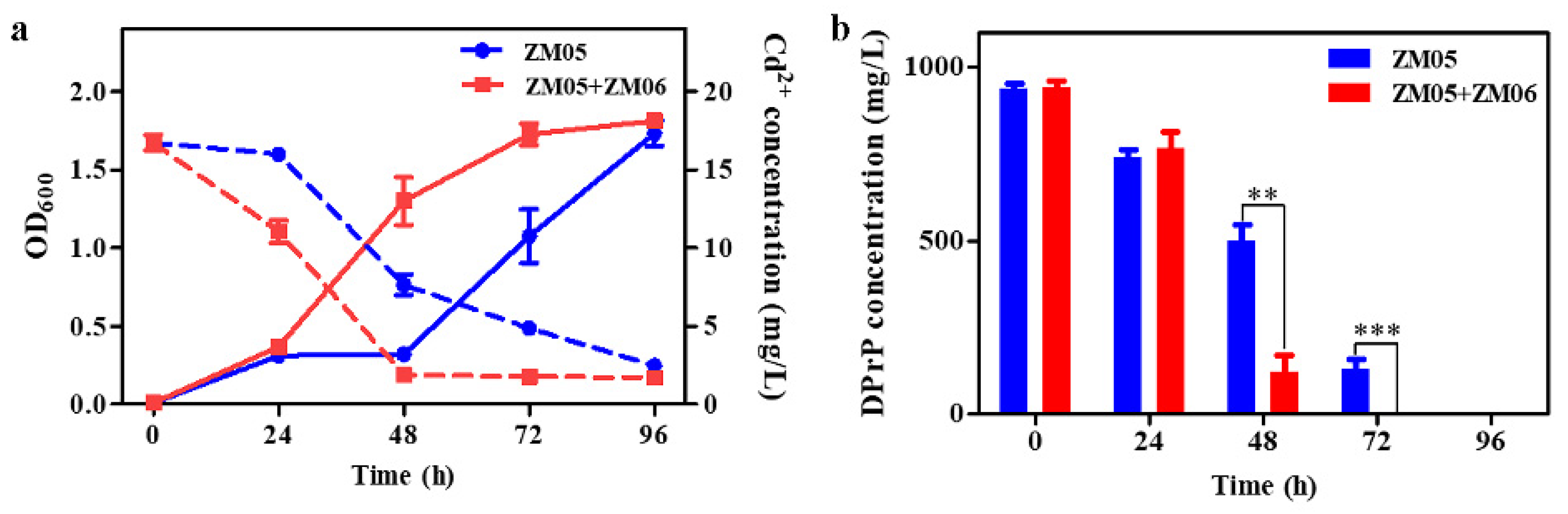
3.1. Construction of a Synergistic Community under Cd Pressure
3.2. Strain ZM06 Relieved Cd (II) Stress in Coculture
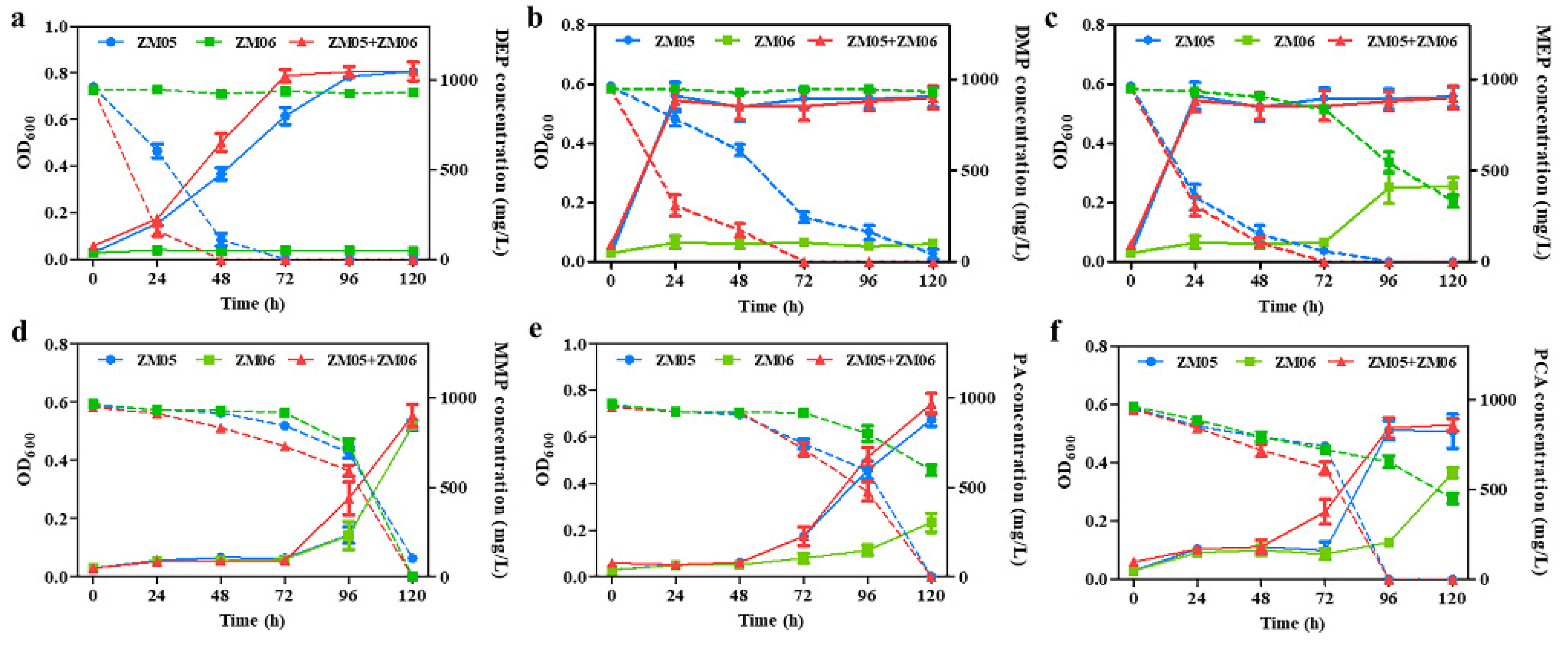
3.3. Strain ZM06 Utilized the Metabolites of DPrP
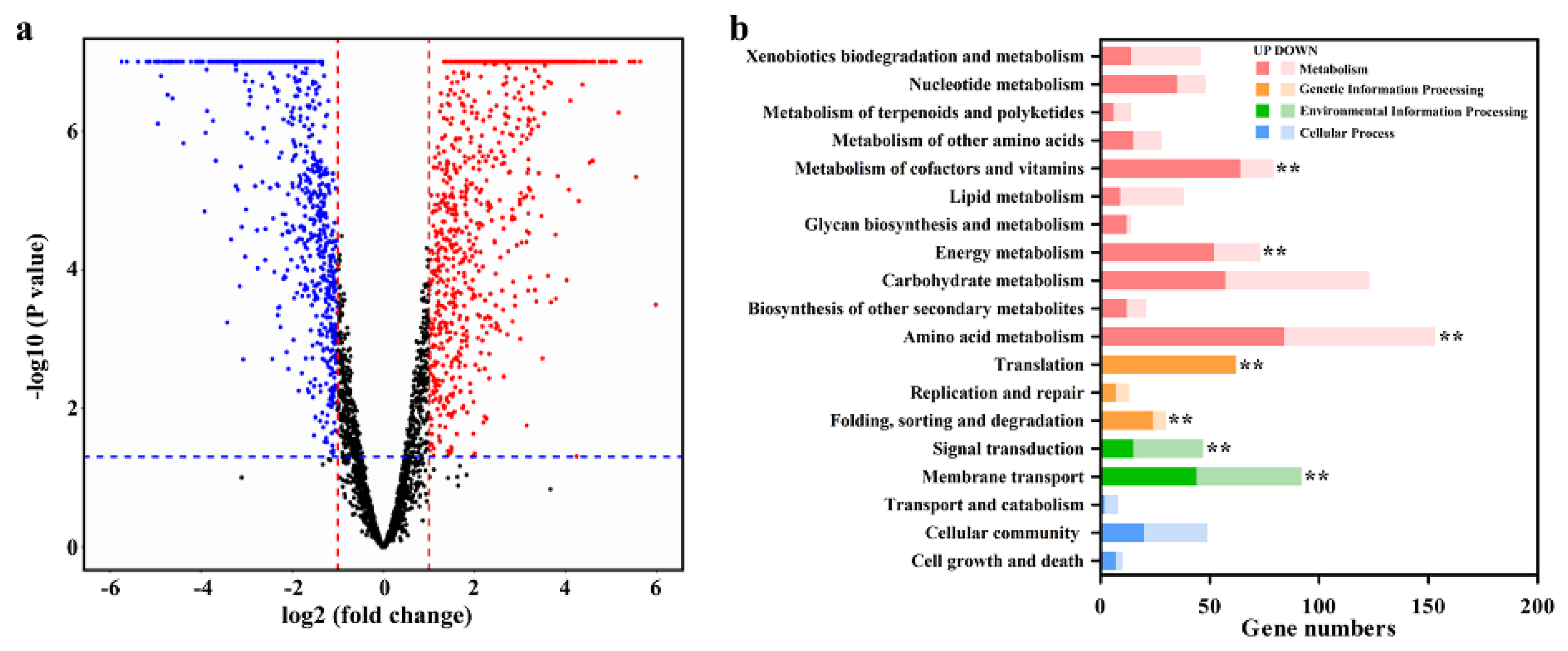
3.4. Transcriptional Response of Strain ZM05 to Cd(II) Stress in Monoculture
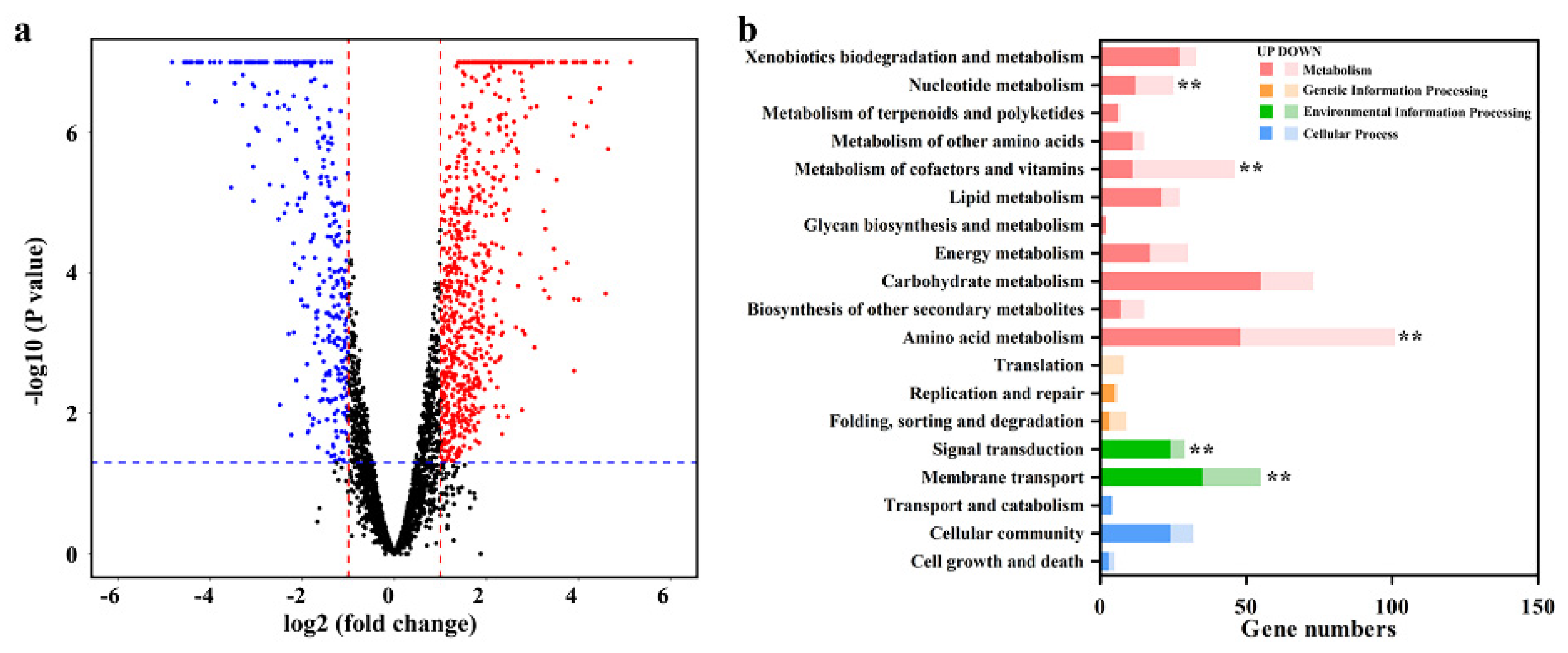
3.5. Gene Expression of Strain ZM05 in Coculture and Monoculture
4. Discussion
4.1. Cd (II) Strongly Stressed Strain ZM05
4.2. Strain ZM06 Accelerated the Degradation of DPrP
4.3. Strain ZM06 Contributed Amino Acids and Fatty Acids to Strain ZM05
4.4. Mechanism of Cooperation between Strain ZM05 and Strain ZM06
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Conflicts of Interest
References
- Liang, D.W.; Zhang, T.; Fang, H.H.P.; He, J. Phthalates biodegradation in the environment. Appl. Microbiol. Biotechnol. 2008, 80, 183–198. [Google Scholar] [CrossRef] [PubMed]
- Kim, Y.H.; Min, J.; Bae, K.D.; Gu, M.B.; Lee, J. Biodegradation of dipropyl phthalate and toxicity of its degradation products: A comparison of Fusarium oxysporum f. sp. pisi cutinase and Candida cylindracea esterase. Arch. Microbiol. 2005, 184, 25–31. [Google Scholar] [CrossRef] [PubMed]
- He, B.S.; Li, J.W. Synthesis of gold nanocubes/PEI-wrinkled CoSe(2)nanomaterials and its application in electrochemical immunosensors for detection of dipropyl phthalate. Rare Met. 2020, 40, 1099–1109. [Google Scholar] [CrossRef]
- Zhang, C.; Tao, Y.; Li, S.; Tian, J.; Ke, T.; Wei, S.; Wang, P.; Chen, L. Simultaneous degradation of trichlorfon and removal of Cd(II) by Aspergillus sydowii strain PA F-2. Environ. Sci. Pollut. Res. 2019, 26, 26844–26854. [Google Scholar] [CrossRef]
- Crain, C.M.; Kroeker, K.; Halpern, B.S. Interactive and cumulative effects of multiple human stressors in marine systems. Ecol. Lett. 2008, 11, 1304–1315. [Google Scholar] [CrossRef] [PubMed]
- Peng, H.; Gao, P.; Chu, G.; Pan, B.; Peng, J.; Xing, B. Enhanced adsorption of Cu(II) and Cd(II) by phosphoric acid-modified biochars. Environ. Pollut. 2017, 229, 846–853. [Google Scholar] [CrossRef] [PubMed]
- Mirlean, N.; Roisenberg, A. The effect of emissions of fertilizer production on the environment contamination by cadmium and arsenic in southern Brazil. Environ. Pollut. 2006, 143, 335–340. [Google Scholar] [CrossRef]
- Rao, Z.X.; Huang, D.Y.; Wu, J.S.; Zhu, Q.H.; Zhu, H.H.; Xu, C.; Xiong, J.; Wang, H.; Duan, M.M. Distribution and availability of cadmium in profile and aggregates of a paddy soil with 30-year fertilization and its impact on Cd accumulation in rice plant. Environ. Pollut. 2018, 239, 198–204. [Google Scholar] [CrossRef]
- Zhang, W.H.; Wu, Y.X.; Simonnot, M.O. Soil Contamination due to E-Waste Disposal and Recycling Activities: A Review with Special Focus on China. Pedosphere 2012, 22, 434–455. [Google Scholar] [CrossRef]
- Adeniyi, A.; Dayomi, M.; Siebe, P.; Okedeyi, O. An assessment of the levels of phthalate esters and metals in the Muledane open dump, Thohoyandou, Limpopo Province, South Africa. Chem. Cent. J. 2008, 2. [Google Scholar] [CrossRef] [Green Version]
- Stoppa, F.; Schiazza, M.; Pellegrini, J.; Ambrosio, F.A.; Rosatelli, G.; D’Orsogna, M.R. Phthalates, heavy metals and PAHs in an overpopulated coastal region: Inferences from Abruzzo, central Italy. Mar. Pollut. Bull. 2017, 125, 501–512. [Google Scholar] [CrossRef]
- Luo, L.; Meng, H.; Gu, J.-D. Microbial extracellular enzymes in biogeochemical cycling of ecosystems. J. Environ. Manag. 2017, 197, 539–549. [Google Scholar] [CrossRef]
- Li, N.; Liu, R.; Chen, J.; Wang, J.; Hou, L.; Zhou, Y. Enhanced phytoremediation of PAHs and cadmium contaminated soils by a Mycobacterium. Sci. Total Environ. 2021, 754. [Google Scholar] [CrossRef] [PubMed]
- Geiger, R.A.; Junghare, M.; Mergelsberg, M.; Ebenau-Jehle, C.; Jesenofsky, V.J.; Jehmlich, N.; von Bergen, M.; Schink, B.; Boll, M. Enzymes involved in phthalate degradation in sulphate-reducing bacteria. Environ. Microbiol. 2019, 21, 3601–3612. [Google Scholar] [CrossRef]
- Liu, Z.B.; He, Z.X.; Huang, H.; Ran, X.B.; Oluwafunmilayo, A.O.; Lu, Z.M. pH Stress-Induced Cooperation between Rhodococcus ruber YYL and Bacillus cereus MLY1 in Biodegradation of Tetrahydrofuran. Front. Microbiol. 2017, 8, 2297. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mikeskova, H.; Novotny, C.; Svobodova, K. Interspecific interactions in mixed microbial cultures in a biodegradation perspective. Appl. Microbiol. Biotechnol. 2012, 95, 861–870. [Google Scholar] [CrossRef]
- Saratale, R.G.; Saratale, G.D.; Kalyani, D.C.; Chang, J.S.; Govindwar, S.P. Enhanced decolorization and biodegradation of textile azo dye Scarlet R by using developed microbial consortium-GR. Bioresour. Technol. 2009, 100, 2493–2500. [Google Scholar] [CrossRef]
- Wu, X.; Liang, R.; Dai, Q.; Jin, D.; Wang, Y.; Chao, W. Complete degradation of di-n-octyl phthalate by biochemical cooperation between Gordonia sp. strain JDC-2 and Arthrobacter sp. strain JDC-32 isolated from activated sludge. J. Hazard. Mater. 2010, 176, 262–268. [Google Scholar] [CrossRef] [PubMed]
- Roane, T.M.; Josephson, K.L.; Pepper, I.L. Dual-bioaugmentation strategy to enhance remediation of cocontaminated soil. Appl. Environ. Microbiol. 2001, 67, 3208–3215. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wang, X.J.; Wu, H.; Wang, X.Y.; Wang, H.X.; Zhao, K.K.; Ma, B.; Lu, Z.M. Network-directed isolation of the cooperator Pseudomonas aeruginosa ZM03 enhanced the dibutyl phthalate degradation capacity of Glutamicibacter nicotianae ZM05 under pH stress. J. Hazard. Mater. 2021, 410. [Google Scholar] [CrossRef]
- Jain, C.; Rodriguez-R, L.M.; Phillippy, A.M.; Konstantinidis, K.T.; Aluru, S. High throughput ANI analysis of 90K prokaryotic genomes reveals clear species boundaries. Nat. Commun. 2018, 9, 5114. [Google Scholar] [CrossRef] [Green Version]
- Lee, I.; Kim, Y.O.; Park, S.C.; Chun, J. OrthoANI: An improved algorithm and software for calculating average nucleotide identity. Int. J. Syst. Evol. Microbiol. 2016, 66, 1100–1103. [Google Scholar] [CrossRef] [PubMed]
- He, Z.X.; Niu, C.Z.; Lu, Z.M. Individual or synchronous biodegradation of di-n-butyl phthalate and phenol by Rhodococcus ruber strain DP-2. J. Hazard. Mater. 2014, 273, 104–109. [Google Scholar] [CrossRef] [PubMed]
- Zhu, F.; Doyle, E.; Zhu, C.; Zhou, D.; Gu, C.; Gao, J. Metagenomic analysis exploring microbial assemblages and functional genes potentially involved in di (2-ethylhexyl) phthalate degradation in soil. Sci. Total Environ. 2020, 715. [Google Scholar] [CrossRef]
- Matsumoto, M.; Kunisawa, A.; Hattori, T.; Kawana, S.; Kitada, Y.; Tamada, H.; Kawano, S.; Hayakawa, Y.; Iida, J.; Fukusaki, E. Free D-amino acids produced by commensal bacteria in the colonic lumen. Sci. Rep. 2018, 8. [Google Scholar] [CrossRef]
- Zhang, S.M.; Wang, H.B.; Zhu, M.J. A sensitive GC/MS detection method for analyzing microbial metabolites short chain fatty acids in fecal and serum samples. Talanta 2019, 196, 249–254. [Google Scholar] [CrossRef]
- Xue, C.; Qi, P.; Li, M.; Liu, Y. Characterization and sorptivity of the Plesiomonas shigelloides strain and its potential use to remove Cd2+ from wastewater. Water 2016, 8, 241. [Google Scholar] [CrossRef]
- Wang, R.; Wei, S.; Jia, P.; Liu, T.; Hou, D.; Xie, R.; Lin, Z.; Ge, J.; Qiao, Y.; Chang, X.; et al. Biochar significantly alters rhizobacterial communities and reduces Cd concentration in rice grains grown on Cd-contaminated soils. Sci. Total Environ. 2019, 676, 627–638. [Google Scholar] [CrossRef] [PubMed]
- Ho, M.T.; Li, M.S.M.; McDowell, T.; MacDonald, J.; Yuan, Z.C. Characterization and genomic analysis of a diesel-degrading bacterium, Acinetobacter calcoaceticus CA16, isolated from Canadian soil. BMC Biotechnol. 2020, 20, 39. [Google Scholar] [CrossRef] [PubMed]
- Whangsuk, W.; Sungkeeree, P.; Nakasiri, M.; Thiengmag, S.; Mongkolsuk, S.; Loprasert, S. Two endocrine disrupting dibutyl phthalate degrading esterases and their compensatory gene expression in Sphingobium sp. SM42. Int. Biodeterior. Biodegrad. 2015, 99, 45–54. [Google Scholar] [CrossRef]
- Bolger, A.M.; Lohse, M.; Usadel, B. Trimmomatic: A flexible trimmer for Illumina sequence data. Bioinformatics 2014, 30, 2114–2120. [Google Scholar] [CrossRef] [Green Version]
- Kim, D.; Langmead, B.; Salzberg, S.L. HISAT: A fast spliced aligner with low memory requirements. Nat. Methods 2015, 12, 357–360. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Anders, S.; Pyl, P.T.; Huber, W. HTSeq—A Python framework to work with high-throughput sequencing data. Bioinformatics 2015, 31, 166–169. [Google Scholar] [CrossRef]
- Pertea, M.; Kim, D.; Pertea, G.M.; Leek, J.T.; Salzberg, S.L. Transcript-level expression analysis of RNA-seq experiments with HISAT, StringTie and Ballgown. Nat. Protoc. 2016, 11, 1650–1667. [Google Scholar] [CrossRef] [PubMed]
- Robinson, M.D.; McCarthy, D.J.; Smyth, G.K. edgeR: A Bioconductor package for differential expression analysis of digital gene expression data. Bioinformatics 2010, 26, 139–140. [Google Scholar] [CrossRef] [Green Version]
- Yu, G.; Wang, L.-G.; Han, Y.; He, Q.-Y. clusterProfiler: An R package for comparing biological themes among gene clusters. OMICS J. Integr. Biol. 2012, 16, 284–287. [Google Scholar] [CrossRef] [PubMed]
- Yang, M.; Zhang, H.; Ni, J.; Chen, W.; Yang, L.; Wei, R. Effect of cadmium on pyrene biodegradation in solution and soil using free and immobilized Escherichia sp. on biochar. Appl. Soil Ecol. 2020, 150. [Google Scholar] [CrossRef]
- Ma, X.K.; Ding, N.; Peterson, E.C.; Daugulis, A.J. Heavy metals species affect fungal-bacterial synergism during the bioremediation of fluoranthene. Appl. Microbiol. Biotechnol. 2016, 100, 7741–7750. [Google Scholar] [CrossRef] [PubMed]
- Ren, C.-G.; Bai, Y.-J.; Kong, C.-C.; Bian, B.; Xie, Z.-H. Synergistic interactions between salt-tolerant rhizobia and arbuscular mycorrhizal fungi on salinity tolerance of Sesbania cannabina plants. J. Plant Growth Regul. 2016, 35, 1098–1107. [Google Scholar] [CrossRef]
- Aimola, P.; Carmignani, M.; Volpe, A.R.; Di Benedetto, A.; Claudio, L.; Waalkes, M.P.; van Bokhoven, A.; Tokar, E.J.; Claudio, P.P. Cadmium induces p53-dependent apoptosis in human prostate epithelial cells. PLoS ONE 2012, 7. [Google Scholar] [CrossRef]
- Anju, R.; Anuj, K.; Ankita, L.; Manu, P. Cellular mechanisms of cadmium-induced toxicity: A review. Int. J. Environ. Health Res. 2014, 24, 378–399. [Google Scholar]
- Cefali, E.; Patane, S.; Arena, A.; Saitta, G.; Guglielmino, S.; Cappello, S.; Nicolo, M.; Allegrini, M. Morphologic variations in bacteria under stress conditions: Near-field optical studies. Scanning 2002, 24, 274–283. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- George, B.; Kumar, J.I.N.; Kumar, R.N.; Sajish, P.R. Biosorption potentiality of living Aspergillus niger Tiegh in removing heavy metal from aqueous solution. Bioremediat. J. 2012, 16, 195–203. [Google Scholar] [CrossRef]
- Chakravarty, R.; Banerjee, P.C. Morphological changes in an acidophilic bacterium induced by heavy metals. Extremophiles 2008, 12, 279–284. [Google Scholar] [CrossRef]
- Holmqvist, E.; Vogel, J. RNA-binding proteins in bacteria. Nat. Rev. Microbiol. 2018, 16, 601–615. [Google Scholar] [CrossRef]
- Chen, J.; Shen, J.; Solem, C.; Jensen, P.R. Oxidative stress at high temperatures in Lactococcus lactis due to an insufficient supply of riboflavin. Appl. Environ. Microbiol. 2013, 79, 6140–6147. [Google Scholar] [CrossRef] [Green Version]
- Ritz, D.; Beckwith, J. Roles of thiol-redox pathways in bacteria. Annu. Rev. Microbiol. 2001, 55, 21–48. [Google Scholar] [CrossRef]
- El-Rab, S.M.F.G.; Shoreit, A.A.F.; Fukumori, Y. Effects of cadmium stress on growth, morphology, and protein expression in Rhodobacter capsulatus B10. Biosci. Biotechnol. Biochem. 2006, 70, 2394–2402. [Google Scholar] [CrossRef]
- Kadam, A.A.; Telke, A.A.; Jagtap, A.S.; Govindwar, S.P. Decolorization of adsorbed textile dyes by developed consortium of Pseudomonas sp. SUK1 and Aspergillus ochraceus NCIM-1146 under solid state fermentation. J. Hazard. Mater. 2011, 189, 486–494. [Google Scholar] [CrossRef]
- Drzyzga, O.; Gottschal, J.C. Tetrachloroethene dehalorespiration and growth of Desulfitobactetium frappieri TCE1 in strict dependence on the activity of Desulfovibrio fructosivorans. Appl. Environ. Microbiol. 2002, 68, 642–649. [Google Scholar] [CrossRef] [Green Version]
- Holmes, V.F.; He, J.; Lee, P.K.H.; Alvarez-Cohen, L. Discrimination of multiple Dehalococcoides strains in a trichloroethene enrichment by quantification of their reductive dehalogenase genes. Appl. Environ. Microbiol. 2006, 72, 5877–5883. [Google Scholar] [CrossRef] [Green Version]
- Lilja, E.E.; Johnson, D.R. Segregating metabolic processes into different microbial cells accelerates the consumption of inhibitory substrates. ISME J. 2016, 10, 1568–1578. [Google Scholar] [CrossRef] [PubMed]
- Nobu, M.K.; Narihiro, T.; Rinke, C.; Kamagata, Y.; Tringe, S.G.; Woyke, T.; Liu, W.T. Microbial dark matter ecogenomics reveals complex synergistic networks in a methanogenic bioreactor. ISME J. 2015, 9, 1710–1722. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zelezniak, A.; Andrejev, S.; Ponomarova, O.; Mende, D.R.; Bork, P.; Patil, K.R. Metabolic dependencies drive species co-occurrence in diverse microbial communities. Proc. Natl. Acad. Sci. USA 2015, 112, 6449–6454. [Google Scholar] [CrossRef] [PubMed] [Green Version]






Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Wang, X.; Shen, S.; Wu, H.; Wang, H.; Wang, L.; Lu, Z. Acinetobacter tandoii ZM06 Assists Glutamicibacter nicotianae ZM05 in Resisting Cadmium Pressure to Preserve Dipropyl Phthalate Biodegradation. Microorganisms 2021, 9, 1417. https://doi.org/10.3390/microorganisms9071417
Wang X, Shen S, Wu H, Wang H, Wang L, Lu Z. Acinetobacter tandoii ZM06 Assists Glutamicibacter nicotianae ZM05 in Resisting Cadmium Pressure to Preserve Dipropyl Phthalate Biodegradation. Microorganisms. 2021; 9(7):1417. https://doi.org/10.3390/microorganisms9071417
Chicago/Turabian StyleWang, Xuejun, Si Shen, Hao Wu, Haixia Wang, Lvjing Wang, and Zhenmei Lu. 2021. "Acinetobacter tandoii ZM06 Assists Glutamicibacter nicotianae ZM05 in Resisting Cadmium Pressure to Preserve Dipropyl Phthalate Biodegradation" Microorganisms 9, no. 7: 1417. https://doi.org/10.3390/microorganisms9071417