
Comparison of a Short Linear Antimicrobial Peptide with Its Disulfide-Cyclized and Cyclotide-Grafted Variants against Clinically Relevant Pathogens



Abstract
:1. Introduction
2. Materials and Methods
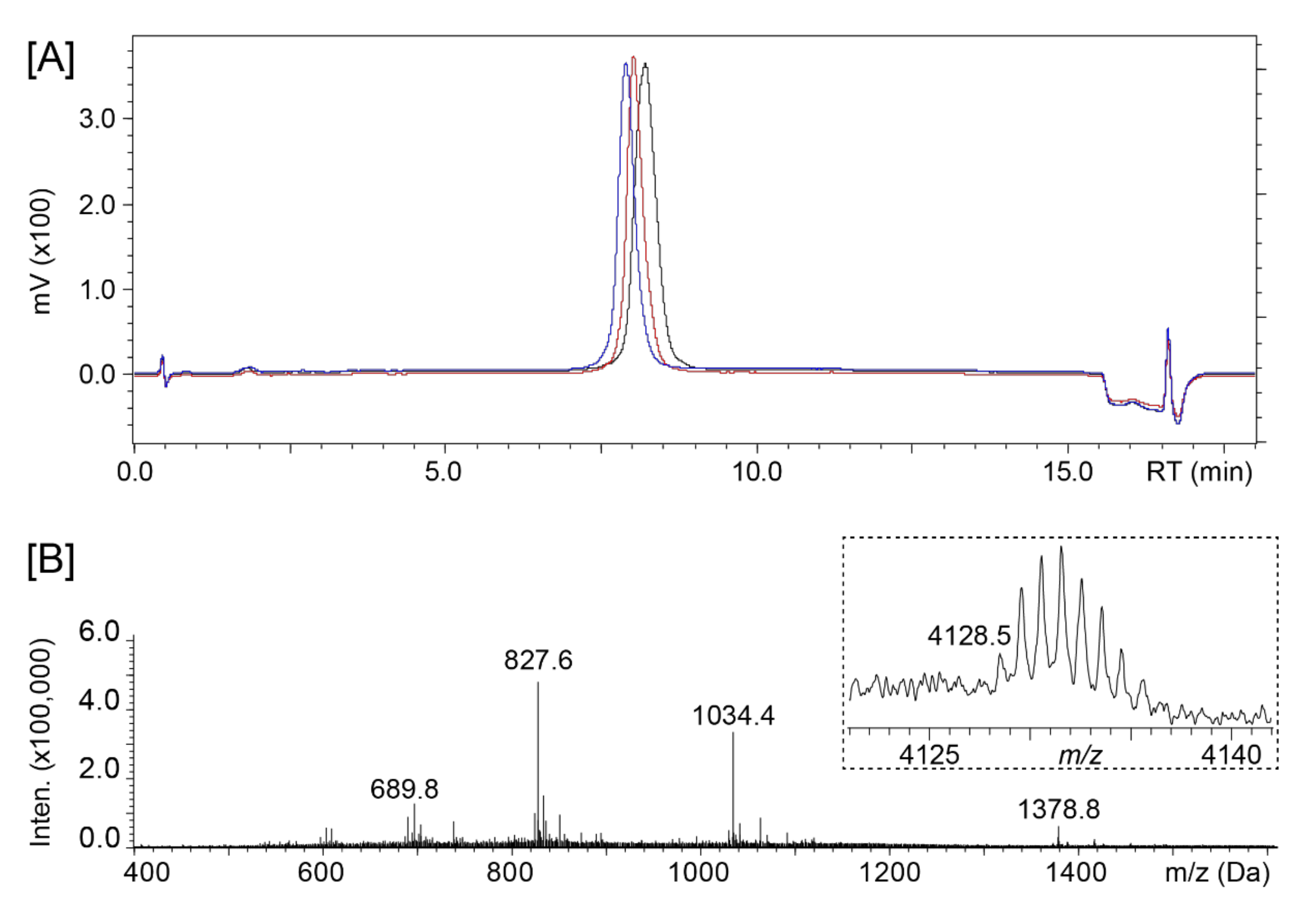
2.1. Peptide Synthesis and Purification
2.2. NMR Sample Analysis
2.3. Bacterial Strains
2.4. Antimicrobial Assays
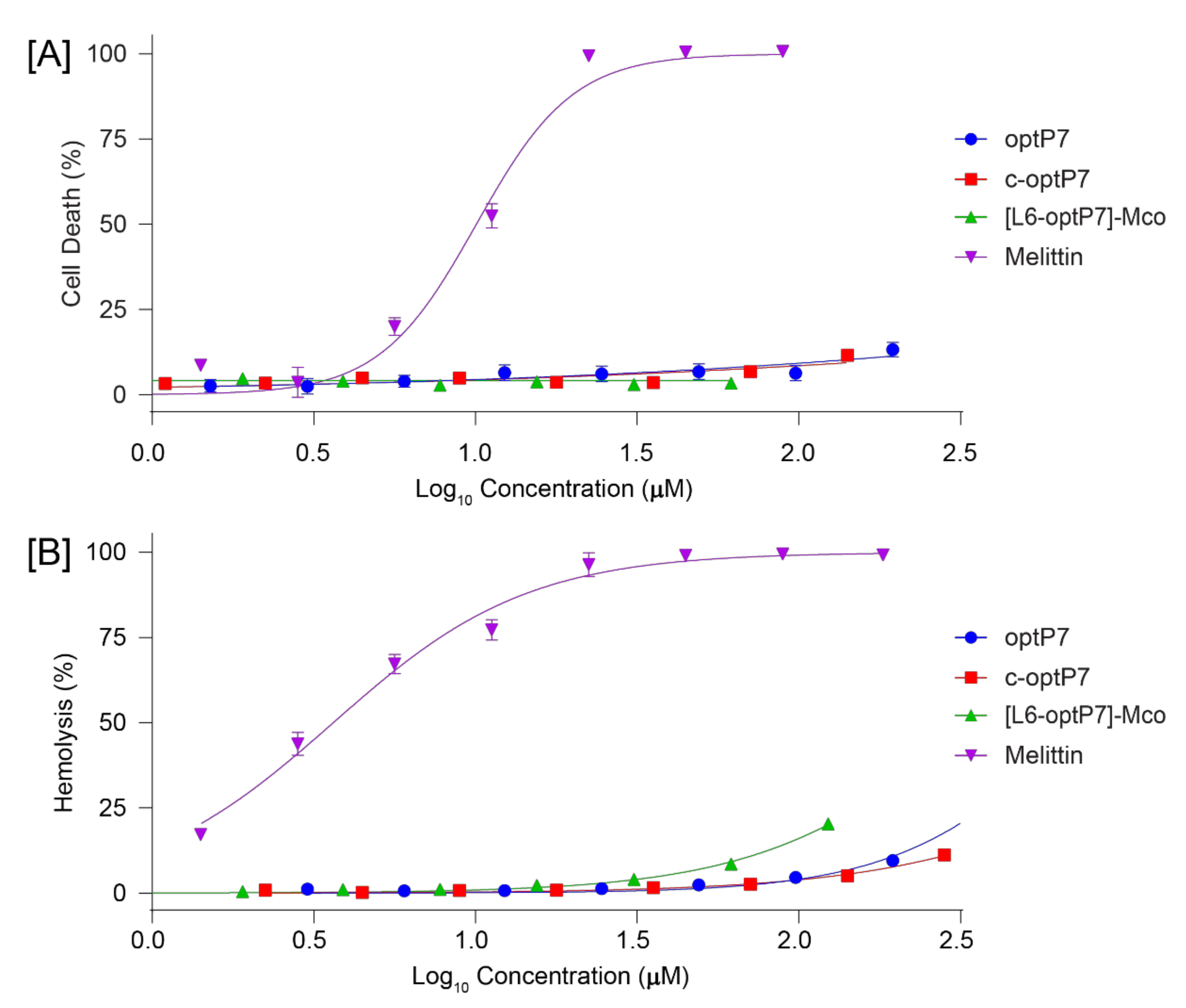
2.5. Cytotoxicity Assays
2.6. Hemolytic Assay
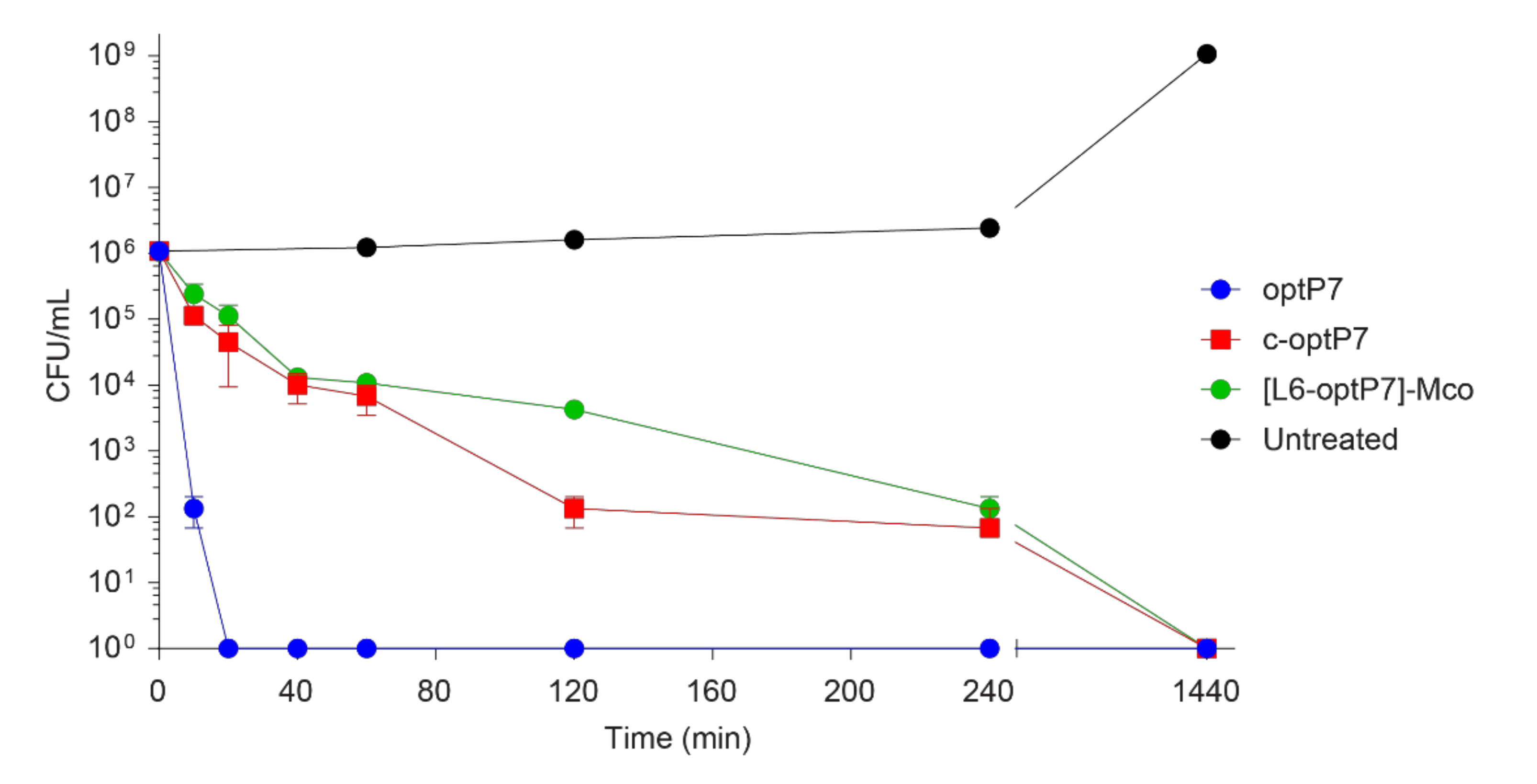
2.7. Time Kill Assay
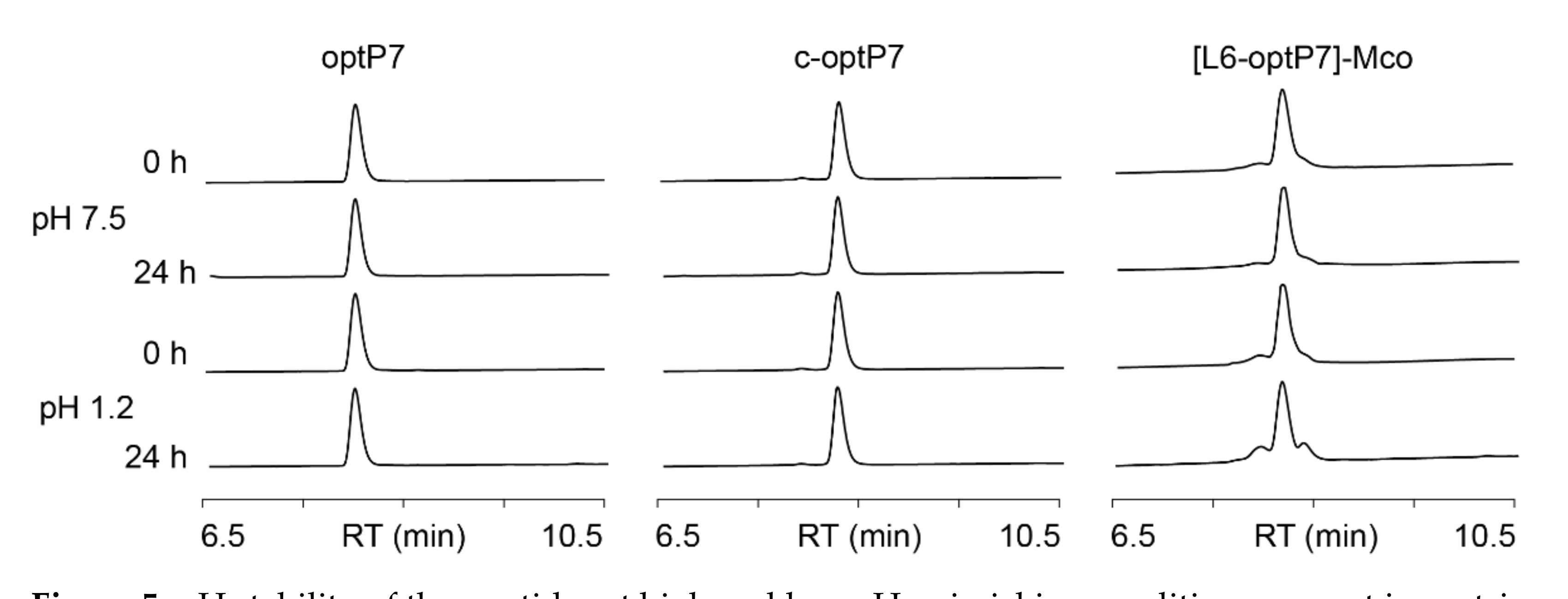
2.8. Stability Assays
3. Results
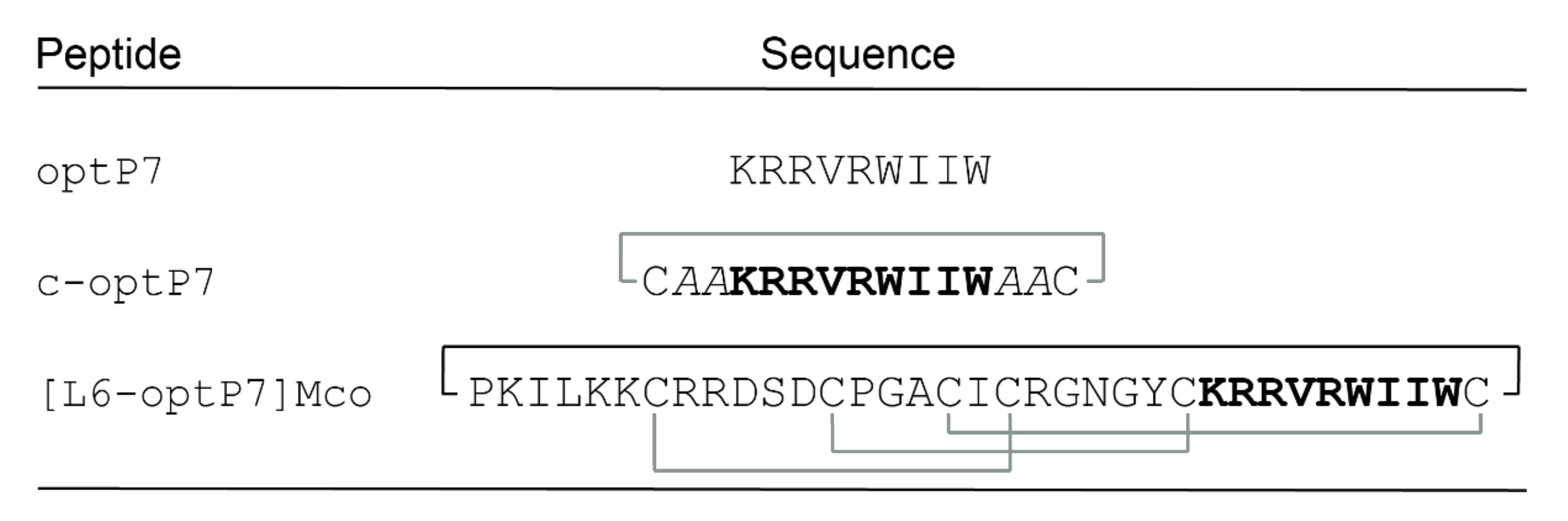
3.1. Design and Synthesis of AMP Grafted MCoTI-II
3.2. Antimicrobial Activity of the Peptides
4. Discussion
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Pendleton, J.N.; Gorman, S.P.; Gilmore, B.F. Clinical relevance of the ESKAPE pathogens. Expert Rev. Anti Infect. Ther. 2013, 11, 297–308. [Google Scholar] [CrossRef]
- Czaplewski, L.; Bax, R.; Clokie, M.; Dawson, M.; Fairhead, H.; Fischetti, V.A.; Foster, S.; Gilmore, B.F.; Hancock, R.E.W.; Harper, D.; et al. Alternatives to antibiotics—A pipeline portfolio review. Lancet Infect. Dis. 2016, 16, 239–251. [Google Scholar] [CrossRef] [Green Version]
- Greber, K.E.; Dawgul, M. Antimicrobial Peptides Under Clinical Trials. Curr. Top. Med. Chem. 2017, 17, 620–628. [Google Scholar] [CrossRef] [PubMed]
- Wang, G.; Li, X.; Wang, Z. APD3: The antimicrobial peptide database as a tool for research and education. Nucleic Acids Res. 2016, 44, D1087–D1093. [Google Scholar] [CrossRef] [Green Version]
- Pirtskhalava, M.; Amstrong, A.A.; Grigolava, M.; Chubinidze, M.; Alimbarashvili, E.; Vishnepolsky, B.; Gabrielian, A.; Rosenthal, A.; Hurt, D.E.; Tartakovsky, M. DBAASP v3: Database of antimicrobial/cytotoxic activity and structure of peptides as a resource for development of new therapeutics. Nucleic Acids Res. 2020, 49, D288–D297. [Google Scholar] [CrossRef]
- Kang, X.; Dong, F.; Shi, C.; Liu, S.; Sun, J.; Chen, J.; Li, H.; Xu, H.; Lao, X.; Zheng, H. DRAMP 2.0, an updated data repository of antimicrobial peptides. Sci. Data 2019, 6, 148. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Koehbach, J.; Craik, D.J. The Vast Structural Diversity of Antimicrobial Peptides. Trends Pharmacol. Sci. 2019, 40, 517–528. [Google Scholar] [CrossRef]
- Falanga, A.; Nigro, E.; De Biasi, M.G.; Daniele, A.; Morelli, G.; Galdiero, S.; Scudiero, O. Cyclic Peptides as Novel Therapeutic Microbicides: Engineering of Human Defensin Mimetics. Molecules 2017, 22, 1217. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mahlapuu, M.; Håkansson, J.; Ringstad, L.; Björn, C. Antimicrobial Peptides: An Emerging Category of Therapeutic Agents. Front. Cell. Infect. Microbiol. 2016, 6, 194. [Google Scholar] [CrossRef] [Green Version]
- Brogden, K.A. Antimicrobial peptides: Pore formers or metabolic inhibitors in bacteria? Nat. Rev. Microbiol. 2005, 3, 238–250. [Google Scholar] [CrossRef] [PubMed]
- Cho, J.H.; Sung, B.H.; Kim, S.C. Buforins: Histone H2A-derived antimicrobial peptides from toad stomach. Biochim. Biophys. Acta 2009, 1788, 1564–1569. [Google Scholar] [CrossRef] [Green Version]
- Kobayashi, S.; Takeshima, K.; Park, C.B.; Kim, S.C.; Matsuzaki, K. Interactions of the novel antimicrobial peptide buforin 2 with lipid bilayers: Proline as a translocation promoting factor. Biochemistry 2000, 39, 8648–8654. [Google Scholar] [CrossRef]
- Xie, Y.; Fleming, E.; Chen, J.L.; Elmore, D.E. Effect of proline position on the antimicrobial mechanism of buforin II. Peptides 2011, 32, 677–682. [Google Scholar] [CrossRef] [Green Version]
- Hale, J.D.; Hancock, R.E. Alternative mechanisms of action of cationic antimicrobial peptides on bacteria. Expert Rev. Anti Infect. Ther. 2007, 5, 951–959. [Google Scholar] [CrossRef] [PubMed]
- Hilpert, K.; McLeod, B.; Yu, J.; Elliott, M.R.; Rautenbach, M.; Ruden, S.; Bürck, J.; Muhle-Goll, C.; Ulrich, A.S.; Keller, S.; et al. Short cationic antimicrobial peptides interact with ATP. Antimicrob. Agents Chemother. 2010, 54, 4480–4483. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Marchand, C.; Krajewski, K.; Lee, H.F.; Antony, S.; Johnson, A.A.; Amin, R.; Roller, P.; Kvaratskhelia, M.; Pommier, Y. Covalent binding of the natural antimicrobial peptide indolicidin to DNA abasic sites. Nucleic Acids Res. 2006, 34, 5157–5165. [Google Scholar] [CrossRef] [PubMed]
- Carlsson, A.; Engström, P.; Palva, E.T.; Bennich, H. Attacin, an antibacterial protein from Hyalophora cecropia, inhibits synthesis of outer membrane proteins in Escherichia coli by interfering with omp gene transcription. Infect. Immun. 1991, 59, 3040–3045. [Google Scholar] [CrossRef] [Green Version]
- Mardirossian, M.; Sola, R.; Beckert, B.; Collis, D.W.P.; Di Stasi, A.; Armas, F.; Hilpert, K.; Wilson, D.N.; Scocchi, M. Proline-Rich Peptides with Improved Antimicrobial Activity against E. coli, K. pneumoniae, and A. baumannii. ChemMedChem 2019, 14, 2025–2033. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mardirossian, M.; Sola, R.; Beckert, B.; Valencic, E.; Collis, D.W.P.; Borišek, J.; Armas, F.; Di Stasi, A.; Buchmann, J.; Syroegin, E.A.; et al. Peptide Inhibitors of Bacterial Protein Synthesis with Broad Spectrum and SbmA-Independent Bactericidal Activity against Clinical Pathogens. J. Med. Chem. 2020, 63, 9590–9602. [Google Scholar] [CrossRef] [PubMed]
- De Leeuw, E.; Li, C.; Zeng, P.; Li, C.; Diepeveen-de Buin, M.; Lu, W.Y.; Breukink, E.; Lu, W. Functional interaction of human neutrophil peptide-1 with the cell wall precursor lipid II. FEBS Lett. 2010, 584, 1543–1548. [Google Scholar] [CrossRef] [Green Version]
- Schmitt, P.; Wilmes, M.; Pugnière, M.; Aumelas, A.; Bachère, E.; Sahl, H.G.; Schneider, T.; Destoumieux-Garzón, D. Insight into invertebrate defensin mechanism of action: Oyster defensins inhibit peptidoglycan biosynthesis by binding to lipid II. J. Biol. Chem. 2010, 285, 29208–29216. [Google Scholar] [CrossRef] [Green Version]
- Knappe, D.; Schmidt, R.; Adermann, K.; Hoffmann, R. Continuous Subcutaneous Delivery of Proline-Rich Antimicrobial Peptide Api137 Provides Superior Efficacy to Intravenous Administration in a Mouse Infection Model. Front. Microbiol. 2019, 10, 2283. [Google Scholar] [CrossRef]
- Yu, K.; Lai, B.F.; Gani, J.; Mikut, R.; Hilpert, K.; Kizhakkedathu, J.N. Interaction of blood components with cathelicidins and their modified versions. Biomaterials 2015, 69, 201–211. [Google Scholar] [CrossRef]
- Craik, D.J.; Daly, N.L.; Bond, T.; Waine, C. Plant cyclotides: A unique family of cyclic and knotted proteins that defines the cyclic cystine knot structural motif. J. Mol. Biol. 1999, 294, 1327–1336. [Google Scholar] [CrossRef]
- Hellinger, R.; Koehbach, J.; Soltis, D.E.; Carpenter, E.J.; Wong, G.K.; Gruber, C.W. Peptidomics of Circular Cysteine-Rich Plant Peptides: Analysis of the Diversity of Cyclotides from Viola tricolor by Transcriptome and Proteome Mining. J. Proteome Res. 2015, 14, 4851–4862. [Google Scholar] [CrossRef]
- Jennings, C.V.; Rosengren, K.J.; Daly, N.L.; Plan, M.; Stevens, J.; Scanlon, M.J.; Waine, C.; Norman, D.G.; Anderson, M.A.; Craik, D.J. Isolation, solution structure, and insecticidal activity of kalata B2, a circular protein with a twist: Do Mobius strips exist in nature? Biochemistry 2005, 44, 851–860. [Google Scholar] [CrossRef] [PubMed]
- Craik, D.J. Host-defense activities of cyclotides. Toxins 2012, 4, 139–156. [Google Scholar] [CrossRef] [Green Version]
- De Veer, S.J.; Kan, M.W.; Craik, D.J. Cyclotides: From Structure to Function. Chem. Rev. 2019, 119, 12375–12421. [Google Scholar] [CrossRef]
- Craik, D.J.; Du, J. Cyclotides as drug design scaffolds. Curr. Opin. Chem. Biol. 2017, 38, 8–16. [Google Scholar] [CrossRef]
- Tam, J.P.; Lu, Y.A.; Yang, J.L.; Chiu, K.W. An unusual structural motif of antimicrobial peptides containing end-to-end macrocycle and cystine-knot disulfides. Proc. Natl. Acad. Sci. USA 1999, 96, 8913–8918. [Google Scholar] [CrossRef] [Green Version]
- Ravipati, A.S.; Henriques, S.T.; Poth, A.G.; Kaas, Q.; Wang, C.K.; Colgrave, M.L.; Craik, D.J. Lysine-rich Cyclotides: A New Subclass of Circular Knotted Proteins from Violaceae. ACS Chem. Biol. 2015, 10, 2491–2500. [Google Scholar] [CrossRef]
- Stromstedt, A.A.; Park, S.; Burman, R.; Goransson, U. Bactericidal activity of cyclotides where phosphatidylethanolamine-lipid selectivity determines antimicrobial spectra. Biochim. Biophys. Acta 2017, 1859, 1986–2000. [Google Scholar] [CrossRef]
- Parsley, N.C.; Sadecki, P.W.; Hartmann, C.J.; Hicks, L.M. Viola “inconspicua” No More: An Analysis of Antibacterial Cyclotides. J. Nat. Prod. 2019, 82, 2537–2543. [Google Scholar] [CrossRef]
- Pränting, M.; Loov, C.; Burman, R.; Göransson, U.; Andersson, D.I. The cyclotide cycloviolacin O2 from Viola odorata has potent bactericidal activity against Gram-negative bacteria. J. Antimicrob. Chemother. 2010, 65, 1964–1971. [Google Scholar] [CrossRef]
- Fensterseifer, I.C.; Silva, O.N.; Malik, U.; Ravipati, A.S.; Novaes, N.R.; Miranda, P.R.; Rodrigues, E.A.; Moreno, S.E.; Craik, D.J.; Franco, O.L. Effects of cyclotides against cutaneous infections caused by Staphylococcus aureus. Peptides 2015, 63, 38–42. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- D’Souza, C.; Henriques, S.T.; Wang, C.K.; Cheneval, O.; Chan, L.Y.; Bokil, N.J.; Sweet, M.J.; Craik, D.J. Using the MCoTI-II Cyclotide Scaffold To Design a Stable Cyclic Peptide Antagonist of SET, a Protein Overexpressed in Human Cancer. Biochemistry 2016, 55, 396–405. [Google Scholar] [CrossRef]
- Wang, C.K.; Gruber, C.W.; Cemazar, M.; Siatskas, C.; Tagore, P.; Payne, N.; Sun, G.; Wang, S.; Bernard, C.C.; Craik, D.J. Molecular grafting onto a stable framework yields novel cyclic peptides for the treatment of multiple sclerosis. ACS Chem. Biol. 2014, 9, 156–163. [Google Scholar] [CrossRef]
- Grimsey, E.; Collis, D.W.P.; Mikut, R.; Hilpert, K. The effect of lipidation and glycosylation on short cationic antimicrobial peptides. Biochim. Biophys. Acta 2020, 1862, 183195. [Google Scholar] [CrossRef]
- Heitz, A.; Hernandez, J.F.; Gagnon, J.; Hong, T.T.; Pham, T.T.; Nguyen, T.M.; Le-Nguyen, D.; Chiche, L. Solution structure of the squash trypsin inhibitor MCoTI-II. A new family for cyclic knottins. Biochemistry 2001, 40, 7973–7983. [Google Scholar] [CrossRef]
- Greenwood, K.P.; Daly, N.L.; Brown, D.L.; Stow, J.L.; Craik, D.J. The cyclic cystine knot miniprotein MCoTI-II is internalized into cells by macropinocytosis. Int. J. Biochem. Cell Biol. 2007, 39, 2252–2264. [Google Scholar] [CrossRef]
- Von Gundlach, A.; Ashby, M.P.; Gani, J.; Lopez-Perez, P.M.; Cookson, A.R.; Ann Huws, S.; Rumancev, C.; Garamus, V.M.; Mikut, R.; Rosenhahn, A.; et al. BioSAXS Measurements Reveal That Two Antimicrobial Peptides Induce Similar Molecular Changes in Gram-Negative and Gram-Positive Bacteria. Front. Pharmacol. 2019, 10, 1127. [Google Scholar] [CrossRef] [Green Version]
- Wiegand, I.; Hilpert, K.; Hancock, R.E. Agar and broth dilution methods to determine the minimal inhibitory concentration (MIC) of antimicrobial substances. Nat. Protoc. 2008, 3, 163–175. [Google Scholar] [CrossRef] [PubMed]
- Gao, Y.; Cui, T.; Lam, Y. Synthesis and disulfide bond connectivity-activity studies of a kalata B1-inspired cyclopeptide against dengue NS2B-NS3 protease. Bioorg. Med. Chem. 2010, 18, 1331–1336. [Google Scholar] [CrossRef] [PubMed]
- Mueller Hinton. Isolation Medium for Antimicrobial Susceptibility Testing; Ref 56137; Bio-Rad: Hercules, CA, USA, 2016. [Google Scholar]
- Mishra, B.; Reiling, S.; Zarena, D.; Wang, G. Host defense antimicrobial peptides as antibiotics: Design and application strategies. Curr. Opin. Chem. Biol. 2017, 38, 87–96. [Google Scholar] [CrossRef]
- Philippe, G.J.B.; Craik, D.J.; Henriques, S.T. Converting peptides into drugs targeting intracellular protein–protein interactions. Drug Discov. Today 2021. [Google Scholar] [CrossRef] [PubMed]
- Clark, R.J.; Fischer, H.; Dempster, L.; Daly, N.L.; Rosengren, K.J.; Nevin, S.T.; Meunier, F.A.; Adams, D.J.; Craik, D.J. Engineering stable peptide toxins by means of backbone cyclization: Stabilization of the alpha-conotoxin MII. Proc. Natl. Acad. Sci. USA 2005, 102, 13767–13772. [Google Scholar] [CrossRef] [Green Version]
- Huang, Y.-H.; Chaousis, S.; Cheneval, O.; Craik, D.J.; Henriques, S.T. Optimization of the cyclotide framework to improve cell penetration properties. Front. Pharmacol. 2015, 6, 17. [Google Scholar] [CrossRef] [Green Version]
- Huang, Y.-H.; Henriques, S.T.; Wang, C.K.; Thorstholm, L.; Daly, N.L.; Kaas, Q.; Craik, D.J. Design of substrate-based BCR-ABL kinase inhibitors using the cyclotide scaffold. Sci. Rep. 2015, 5, 12974. [Google Scholar] [CrossRef] [Green Version]
- Knappe, D.; Ruden, S.; Langanke, S.; Tikkoo, T.; Ritzer, J.; Mikut, R.; Martin, L.L.; Hoffmann, R.; Hilpert, K. Optimization of oncocin for antibacterial activity using a SPOT synthesis approach: Extending the pathogen spectrum to Staphylococcus aureus. Amino Acids 2016, 48, 269–280. [Google Scholar] [CrossRef]
- Knappe, D.; Adermann, K.; Hoffmann, R. Oncocin Onc72 is efficacious against antibiotic-susceptible Klebsiella pneumoniae ATCC 43816 in a murine thigh infection model. Pept. Sci. 2015, 104, 707–711. [Google Scholar] [CrossRef] [PubMed]
- Von Gundlach, A.R.; Garamus, V.M.; Gorniak, T.; Davies, H.A.; Reischl, M.; Mikut, R.; Hilpert, K.; Rosenhahn, A. Small angle X-ray scattering as a high-throughput method to classify antimicrobial modes of action. Biochim. Biophys. Acta 2016, 1858, 918–925. [Google Scholar] [CrossRef]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Bacteria | Growth Media | Fraction 4 | Fraction 5 | Fraction 6 |
|---|---|---|---|---|
| P. aeruginosa | MH 1/5 MH | 31 4–8 | 62 4 | 62 8 |
| E. coli | MH | 62 | 62 | 65 |
| MRSA | MH | 62 | 62 | >62 |
| Bacteria | Growth Media or Ratio | optP7 | c-optP7 | [L6-optP7]Mco | Melittin * |
|---|---|---|---|---|---|
| P. aeruginosa (Gram-negative) | MH | 3 | 142 | 62 | ND |
| MH + 10% serum | 49 | >142 | >62 | ND | |
| Ratio MH + 10% serum/ MH | 16.3 | NA | NA | NA | |
| 1/5 MH | 0.8 | 9 | 8 | ND | |
| 1/5 MH + 10% serum | 6 | 35 | 8 | ND | |
| Ratio 1/5 MH + 10% serum/ 1/5MH | 7.5 | 3.9 | 1 | NA | |
| E. coli (Gram-negative) | MH | 0.8 | 71 | 62 | 16 |
| MH + 10% serum | 6 | >142 | >62 * | ND | |
| Ratio MH + 10% serum/ MH | 7.5 | NA | NA | NA | |
| 1/5 MH | 0.4–0.8 | 4–9 | 2 | 3 | |
| 1/5 MH + 10% serum | 0.8 | 18 | 15 | 11-22 | |
| 1/5 MH + 10% HI serum | 0.8 | 35 | 15 | 11 | |
| Ratio 1/5 MH + 10% serum/ 1/5MH | 2–1 | 4.5–2 | 7.5 | 3.6–7.3 | |
| K. pneumoniae (Gram-negative) | MH | 1.5 | >142 * | 62 * | ND |
| MH + 10% serum | ND | ND | ND | ND | |
| Ratio MH + 10% serum/ MH | NA | NA | NA | NA | |
| 1/5 MH | 1.5 | 71 | 31 | ND | |
| 1/5 MH + 10% serum | 24 | 142 | >62 | ND | |
| Ratio 1/5 MH + 10% serum/ 1/5MH | 16 | 2 | NA | NA | |
| A. baumannii (Gram-negative) | MH | 1.5 | ND | 62 | ND |
| MH + 10% serum | ND | ND | ND | ND | |
| Ratio MH + 10% serum/ MH | NA | NA | NA | NA | |
| 1/5 MH | 0.8 | 4 | 4 | ND | |
| 1/5 MH + 10% serum | 3 | 71 | 31 | ND | |
| Ratio 1/5 MH + 10% serum/ 1/5MH | 3.75 | 17.8 | 7.75 | NA | |
| VRE (Gram-positive) | BHI | 0.8 | >142 * | >62 * | ND |
| BHI + 10% serum | ND | ND | ND | ND | |
| Ratio BHI + 10% serum/ BHI | NA | NA | NA | NA | |
| 1/5 BHI | 0.8 | 18 | 15 | ND | |
| 1/5 BHI + 10% serum | 24 | >142 | 62 | ND | |
| Ratio 1/5 BHI + 10% serum/ 1/5BHI | 30 | >7.8 | 4.1 | NA | |
| MRSA (Gram-positive) | MH | 6 | 142 | >62 | 4 |
| MH + 10% serum | 49 | >142 | >62 | ND | |
| Ratio MH + 10% serum/ MH | 8.1 | NA | NA | NA | |
| 1/5 MH | 1.5 | 18 | 8 | 0.7 | |
| 1/5 MH + 10% serum | 6 | >142 | >62 | 1.4 | |
| 1/5 MH + 10% HI serum | 6 | >142 | >62 | 1.4 | |
| Ratio 1/5 MH + 10% serum/ 1/5MH | 4 | >7.9 | >7.8 | 2 |
| Bacteria | Growth Media | Ratio c-optP7/optP7 | Ratio L6-optP7-Mco/optP7 |
|---|---|---|---|
| P. aeruginosa | MH | 47.3 | 20.6 |
| MH + 10% serum | NA | NA | |
| 1/5 MH | 11.3 | 10.0 | |
| 1/5 MH + 10% serum | 5.8 | 1.3 | |
| E. coli | MH | 88.8 | 77.5 |
| MH + 10% serum | NA | NA | |
| 1/5 MH | 10.0 | 3.3 | |
| 1/5 MH + 10% serum | 22.5 | 18.8 | |
| K. pneumoniae | MH | NA | 41.3 |
| MH + 10% serum | ND | ND | |
| 1/5 MH | 47.3 | 20.7 | |
| 1/5 MH + 10% serum | 5.9 | NA | |
| A. baumannii | MH | ND | 41.3 |
| MH + 10% serum | ND | ND | |
| 1/5 MH | 5 | 5 | |
| 1/5 MH + 10% serum | 23.7 | 10.3 | |
| VRE | BHI | NA | NA |
| BHI + 10% serum | ND | ND | |
| 1/5 BHI | 22.5 | 18.8 | |
| 1/5 BHI + 10% serum | NA | 2.6 | |
| MRSA | MH | 23.6 | ND |
| MH + 10% serum | NA | NA | |
| 1/5 MH | 18 | 8 | |
| 1/5 MH + 10% serum | NA | NA |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Koehbach, J.; Gani, J.; Hilpert, K.; Craik, D.J. Comparison of a Short Linear Antimicrobial Peptide with Its Disulfide-Cyclized and Cyclotide-Grafted Variants against Clinically Relevant Pathogens. Microorganisms 2021, 9, 1249. https://doi.org/10.3390/microorganisms9061249
Koehbach J, Gani J, Hilpert K, Craik DJ. Comparison of a Short Linear Antimicrobial Peptide with Its Disulfide-Cyclized and Cyclotide-Grafted Variants against Clinically Relevant Pathogens. Microorganisms. 2021; 9(6):1249. https://doi.org/10.3390/microorganisms9061249
Chicago/Turabian StyleKoehbach, Johannes, Jurnorain Gani, Kai Hilpert, and David J Craik. 2021. "Comparison of a Short Linear Antimicrobial Peptide with Its Disulfide-Cyclized and Cyclotide-Grafted Variants against Clinically Relevant Pathogens" Microorganisms 9, no. 6: 1249. https://doi.org/10.3390/microorganisms9061249