
The Fungal Effector Mlp37347 Alters Plasmodesmata Fluxes and Enhances Susceptibility to Pathogen

 , ,
, ,  , ,
, , {kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Materials and Methods
2.1. Cloning Procedures and Plasmid Constructs
2.2. Plant Material, Growth Conditions, and Transgenic Production
2.3. Expression in Nicotiana Benthamiana
2.4. Pathogen Infection Assays
2.5. Confocal Microscopy Assay
2.6. mCherry Diffusion Assay
2.7. DANS Assay and Callose Quantification
2.8. Y2H Reporter Assays
2.9. Western Blot Analysis
2.10. RNA Extraction and Transcriptome Analysis
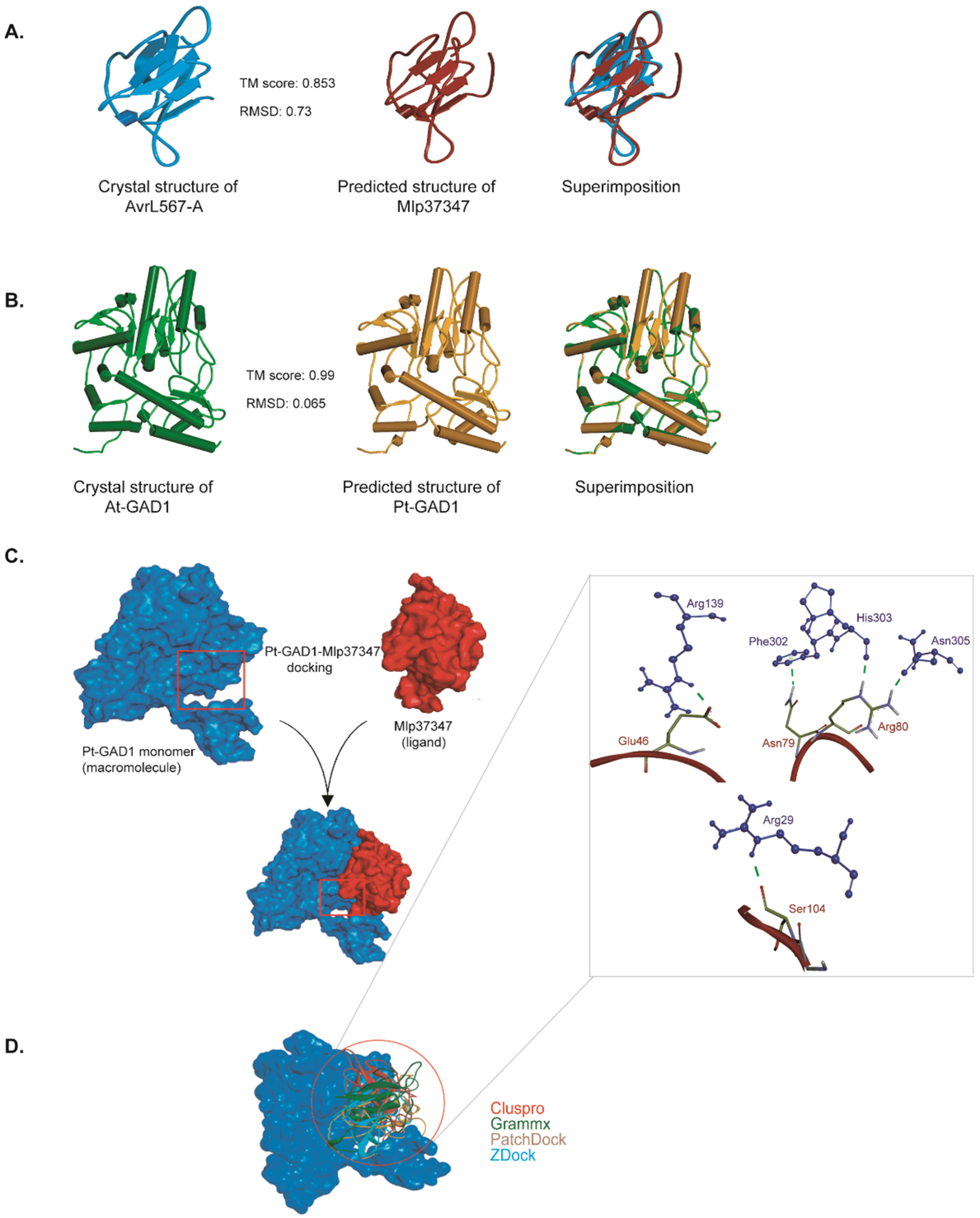
2.11. In Silico Protein–Protein Binding
3. Results
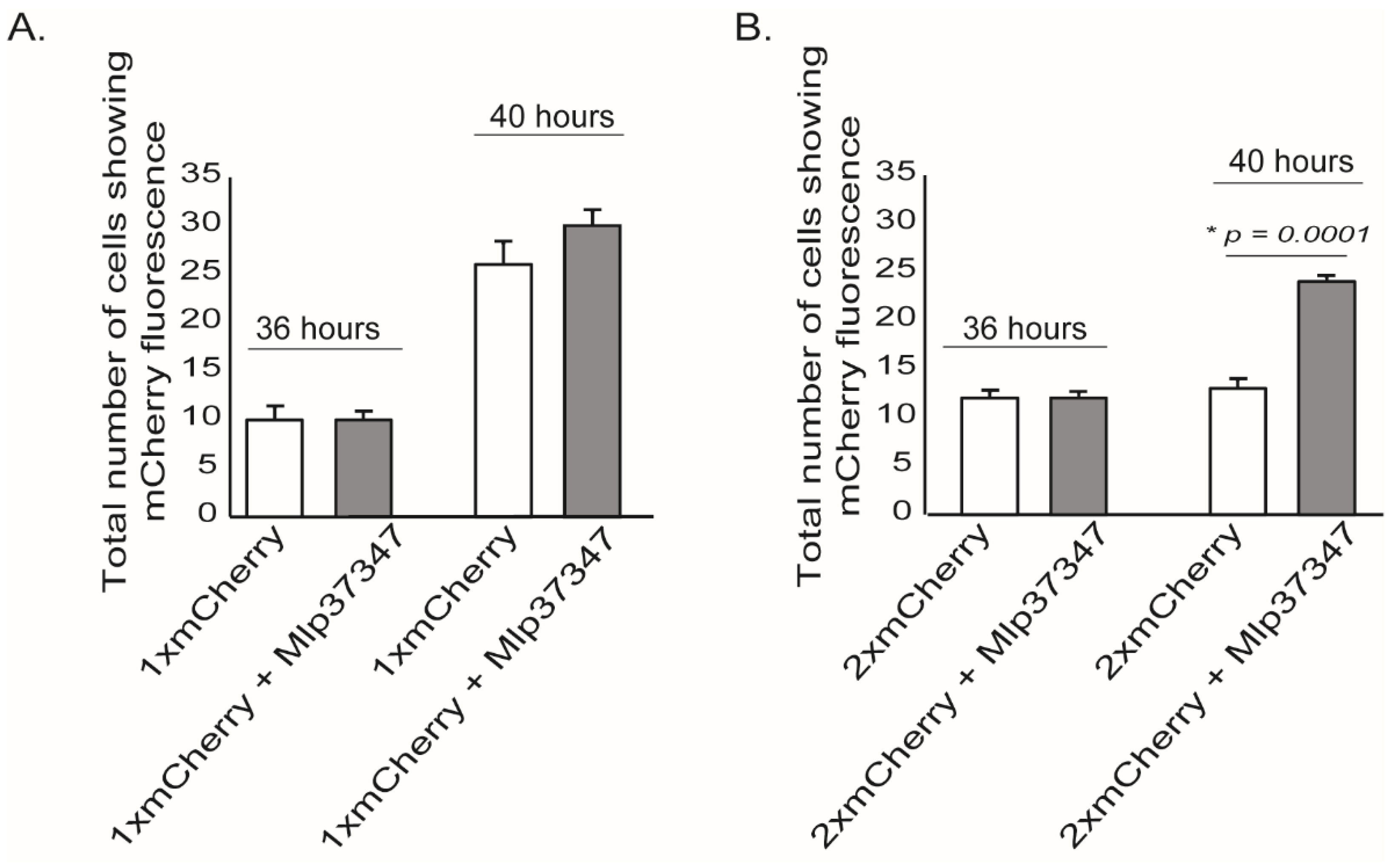
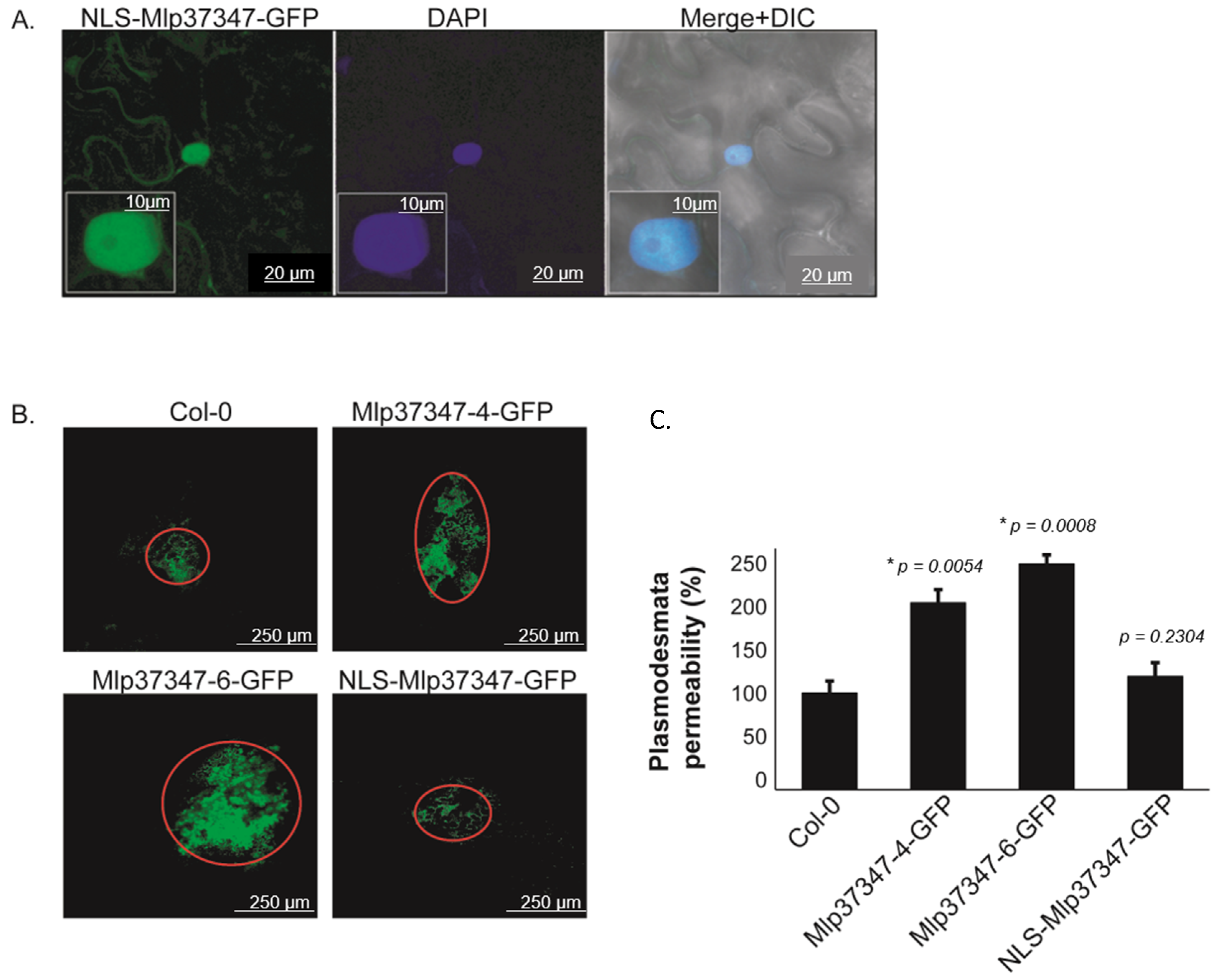
3.1. Mlp37347 Enhances Plasmodesmata Flux
3.2. Plasmodesmata Localization of Mlp37347 Is Required for Enhanced Plasmodesmatal Flux
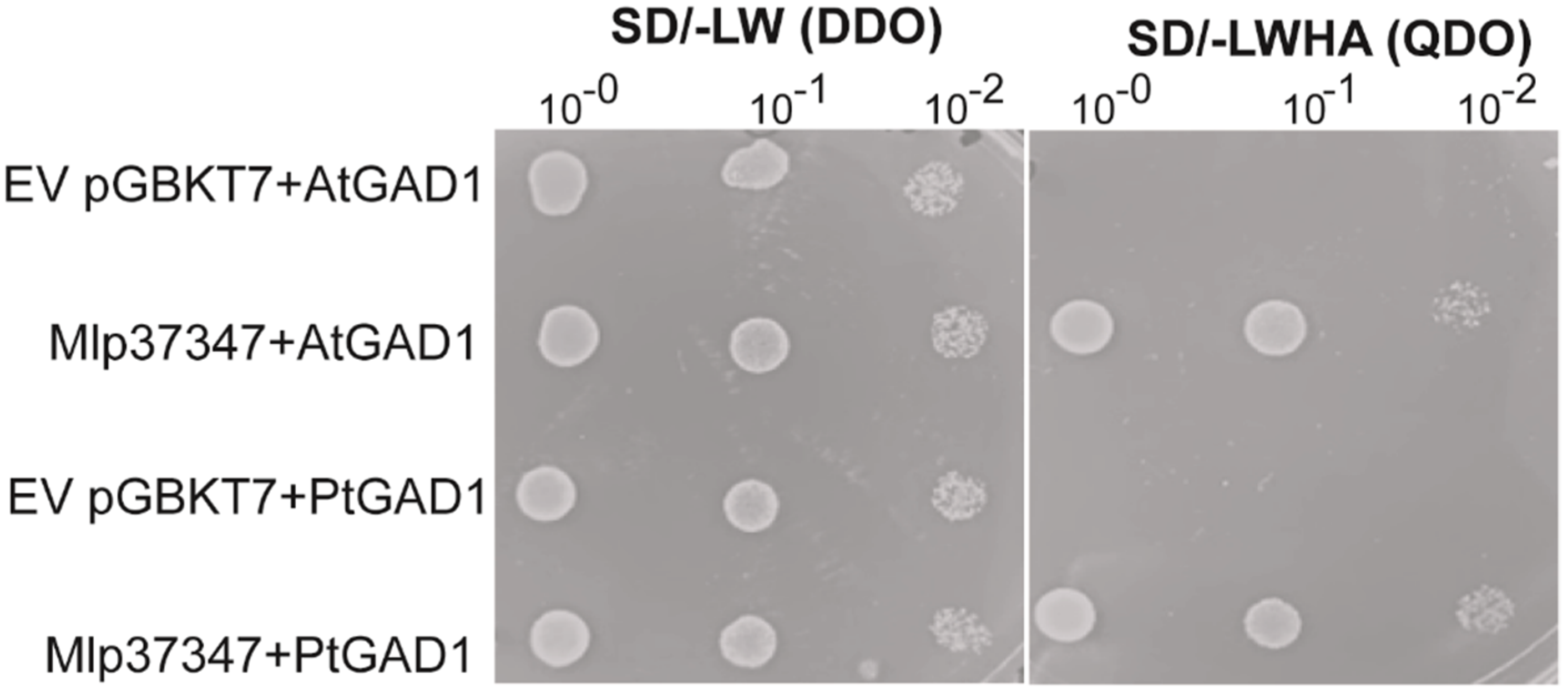
3.3. Mlp37347–GAD1 Interaction
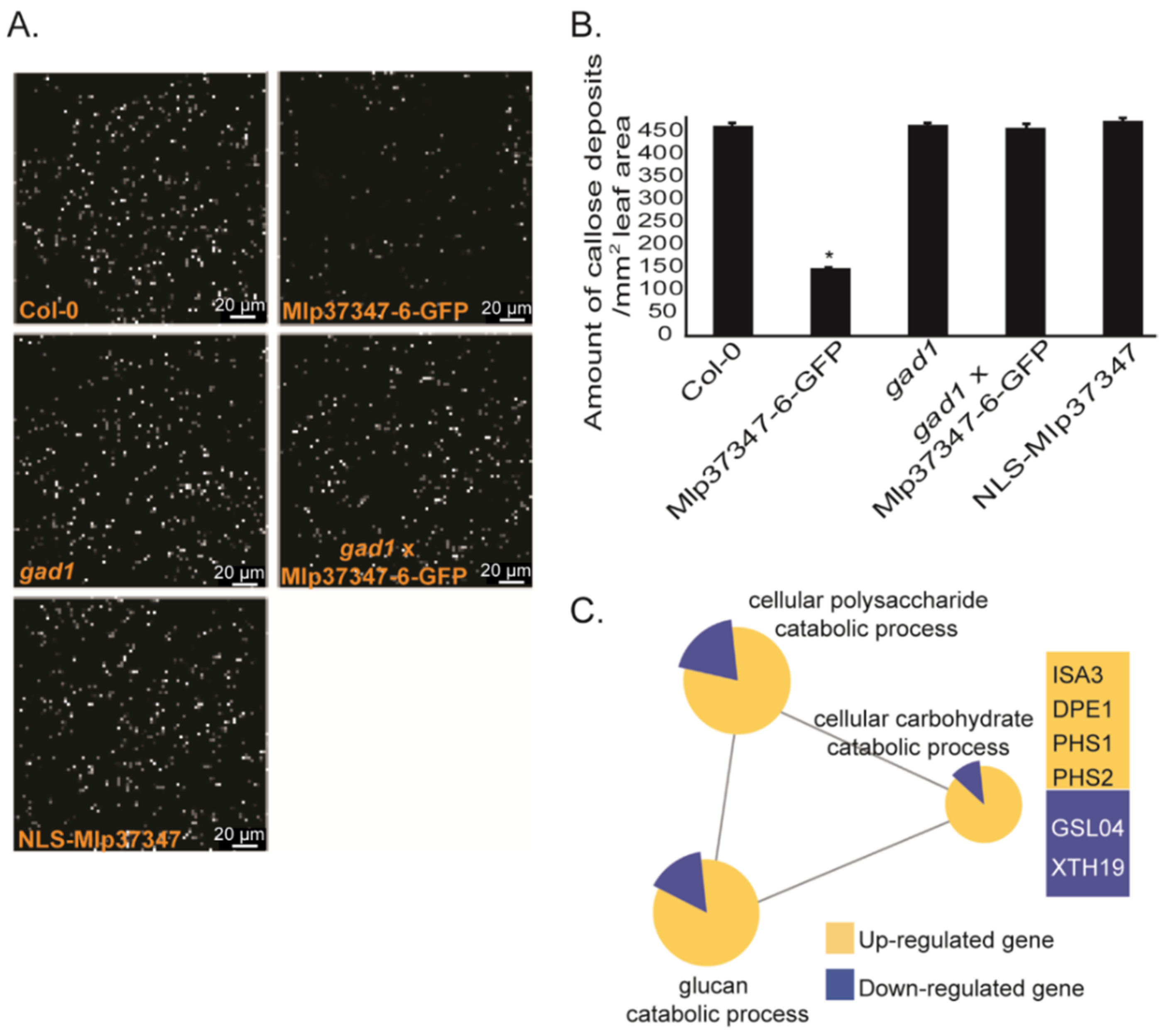
3.4. Mlp37347 Decreases Plasmodesmatal Callose Deposition and Affects Callose Metabolism Gene Expression
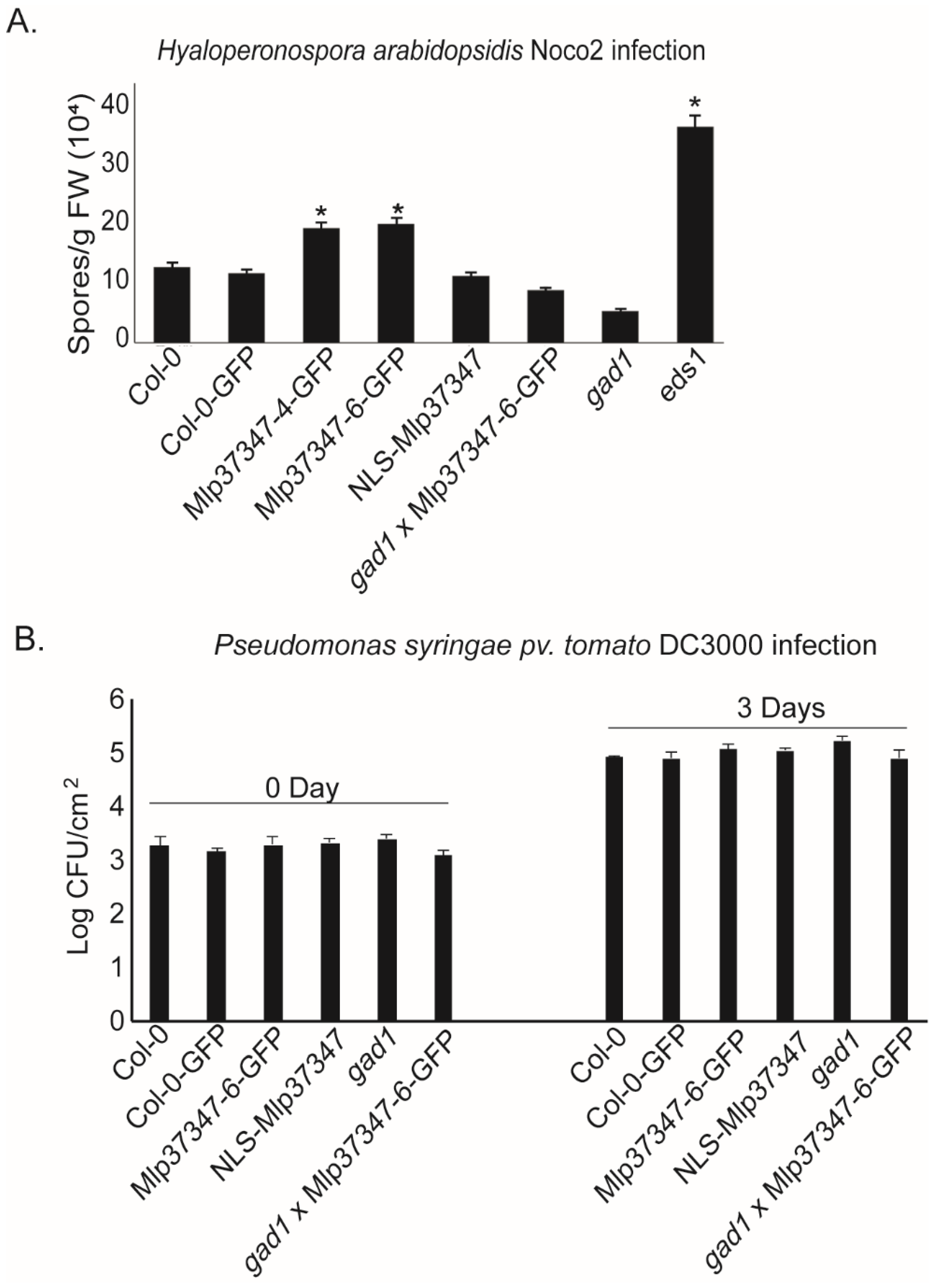
3.5. Mlp37347 Increases the Susceptibility of A. thaliana to H. arabidopsidis
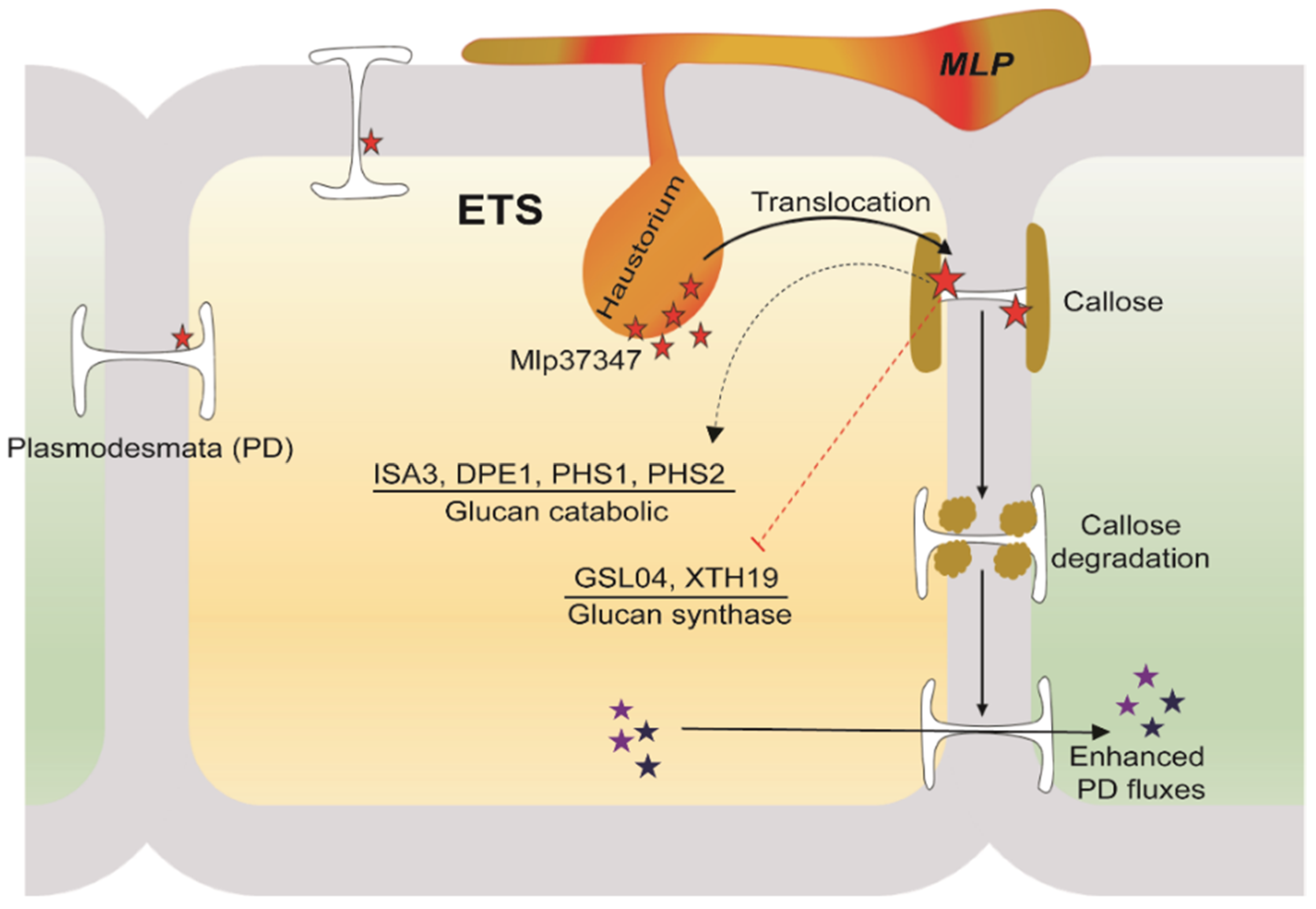
4. Discussion
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Chiang, Y.-H.; Coaker, G. Effector Triggered Immunity: NLR Immune Perception and Downstream Defense Responses. Arab. Book 2015, 13, e0183. [Google Scholar] [CrossRef] [Green Version]
- Zipfel, C. Plant pattern-recognition receptors. Trends Immunol. 2014, 35, 345–351. [Google Scholar] [CrossRef]
- Hogenhout, S.A.; Van Der Hoorn, R.A.L.; Terauchi, R.; Kamoun, S. Emerging Concepts in Effector Biology of Plant-Associated Organisms. Mol. Plant-Microbe Interact. 2009, 22, 115–122. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Petre, B.; Kamoun, S. How Do Filamentous Pathogens Deliver Effector Proteins into Plant Cells? PLoS Biol. 2014, 12, e1001801. [Google Scholar] [CrossRef] [Green Version]
- Rafiqi, M.; Ellis, J.G.; Ludowici, V.A.; Hardham, A.R.; Dodds, P.N. Challenges and progress towards understanding the role of effectors in plant–fungal interactions. Curr. Opin. Plant Biol. 2012, 15, 477–482. [Google Scholar] [CrossRef]
- Duplessis, S.; Cuomo, C.A.; Lin, Y.-C.; Aerts, A.; Tisserant, E.; Veneault-Fourrey, C.; Joly, D.L.; Hacquard, S.; Amselem, J.; Cantarel, B.L.; et al. Obligate biotrophy features unraveled by the genomic analysis of rust fungi. Proc. Natl. Acad. Sci. USA 2011, 108, 9166–9171. [Google Scholar] [CrossRef] [Green Version]
- Hacquard, S.; Delaruelle, C.; Legué, V.; Tisserant, E.; Kohler, A.; Frey, P.; Martin, F.; Duplessis, S. Laser Capture Microdissection of Uredinia Formed by Melampsora larici-populina Revealed a Transcriptional Switch Between Biotrophy and Sporulation. Mol. Plant-Microbe Interact. 2010, 23, 1275–1286. [Google Scholar] [CrossRef] [PubMed]
- Hacquard, S.; Joly, D.L.; Lin, Y.-C.; Tisserant, E.; Feau, N.; Delaruelle, C.; Legué, V.; Kohler, A.; Tanguay, P.; Petre, B.; et al. A Comprehensive Analysis of Genes Encoding Small Secreted Proteins Identifies Candidate Effectors in Melampsora larici-populina (Poplar Leaf Rust). Mol. Plant-Microbe Interact. 2012, 25, 279–293. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Joly, D.L.; Feau, N.; Tanguay, P.; Hamelin, R.C. Comparative analysis of secreted protein evolution using expressed sequence tags from four poplar leaf rusts (Melampsora spp.). BMC Genom. 2010, 11, 422. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Germain, H.; Joly, D.L.; Mireault, C.; Plourde, M.B.; Letanneur, C.; Stewart, D.; Morency, M.-J.; Petre, B.; Duplessis, S.; Séguin, A. Infection assays in Arabidopsis reveal candidate effectors from the poplar rust fungus that promote susceptibility to bacteria and oomycete pathogens. Mol. Plant Pathol. 2018, 19, 191–200. [Google Scholar] [CrossRef] [Green Version]
- Petre, B.; Saunders, D.G.O.; Sklenar, J.; Lorrain, C.; Win, J.; Duplessis, S.; Kamoun, S. Candidate Effector Proteins of the Rust Pathogen Melampsora larici-populina Target Diverse Plant Cell Compartments. Mol. Plant-Microbe Interact. 2015, 28, 689–700. [Google Scholar] [CrossRef] [Green Version]
- Lucas, W.J.; Lee, J.-Y. Plasmodesmata as a supracellular control network in plants. Nat. Rev. Mol. Cell Biol. 2004, 5, 712–726. [Google Scholar] [CrossRef]
- De Storme, N.; Geelen, D. Callose homeostasis at plasmodesmata: Molecular regulators and developmental relevance. Front. Plant Sci. 2014, 5, 138. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Caillaud, M.-C.; Piquerez, S.J.M.; Fabro, G.; Steinbrenner, J.; Ishaque, N.; Beynon, J.; Jones, J.D.G. Subcellular localization of the Hpa RxLR effector repertoire identifies a tonoplast-associated protein HaRxL17 that confers enhanced plant susceptibility. Plant J. 2011, 69, 252–265. [Google Scholar] [CrossRef] [PubMed]
- Caillaud, M.-C.; Wirthmueller, L.; Fabro, G.; Piquerez, S.J.M.; Asai, S.; Ishaque, N.; Jones, J.D.G. Mechanisms of Nuclear Suppression of Host Immunity by Effectors from the Arabidopsis Downy Mildew Pathogen Hyaloperonospora arabidopsidis (Hpa). Cold Spring Harb. Symp. Quant. Biol. 2012, 77, 285–293. [Google Scholar] [CrossRef] [PubMed]
- Du, Y.; Berg, J.; Govers, F.; Bouwmeester, K. Immune activation mediated by the late blight resistance protein R1 requires nuclear localization of R1 and the effector AVR 1. New Phytol. 2015, 207, 735–747. [Google Scholar] [CrossRef] [PubMed]
- Gaouar, O.; Morency, M.-J.; Letanneur, C.; Séguin, A.; Germain, H. The 124202 candidate effector of Melampsora larici-populina interacts with membranes in Nicotiana and Arabidopsis. Can. J. Plant Pathol. 2016, 38, 197–208. [Google Scholar] [CrossRef]
- Kunjeti, S.G.; Iyer, G.; Johnson, E.; Li, E.; Broglie, K.E.; Rauscher, G.; Rairdan, G.J. Identification of Phakopsora pachyrhizi Candidate Effectors with Virulence Activity in a Distantly Related Pathosystem. Front. Plant Sci. 2016, 7, 269. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Petre, B.; Lorrain, C.; Saunders, D.G.O.; Win, J.; Sklenar, J.; Duplessis, S.; Kamoun, S. Rust fungal effectors mimic host transit peptides to translocate into chloroplasts. Cell. Microbiol. 2015, 18, 453–465. [Google Scholar] [CrossRef] [Green Version]
- Karimi, M.; Inzé, D.; Depicker, A. GATEWAY™ vectors for Agrobacterium-mediated plant transformation. Trends Plant Sci. 2002, 7, 193–195. [Google Scholar] [CrossRef]
- Mireault, C.; Paris, L.-E.; Germain, H. Enhancement of the Arabidopsis floral dip method with XIAMETER OFX-0309 as alternative to Silwet L-77 surfactant. Botany 2014, 92, 523–525. [Google Scholar] [CrossRef] [Green Version]
- Miyashita, Y.; Good, A.G. Contribution of the GABA shunt to hypoxia-induced alanine accumulation in roots of Arabidopsis thaliana. Plant Cell Physiol. 2008, 49, 92–102. [Google Scholar] [CrossRef] [PubMed]
- Krenek, P.; Samajova, O.; Luptovciak, I.; Doskocilova, A.; Komis, G.; Samaj, J. Transient plant transformation mediated by Agrobacterium tumefaciens: Principles, methods and applications. Biotechnol. Adv. 2015, 33, 1024–1042. [Google Scholar] [CrossRef]
- Ahmed, M.B.; Santos, K.C.G.D.; Sanchez, I.B.; Petre, B.; Lorrain, C.; Plourde, M.B.; Duplessis, S.; Desgagné-Penix, I.; Germain, H. A rust fungal effector binds plant DNA and modulates transcription. Sci. Rep. 2018, 8, 1–14. [Google Scholar] [CrossRef]
- Cao, L.; Blekemolen, M.C.; Tintor, N.; Cornelissen, B.J.; Takken, F.L. The Fusarium oxysporum Avr2-Six5 Effector Pair Alters Plasmodesmatal Exclusion Selectivity to Facilitate Cell-to-Cell Movement of Avr2. Mol. Plant 2018, 11, 691–705. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cui, W.; Wang, X.; Lee, J.-Y. Drop-ANd-See: A simple, real-time, and noninvasive technique for assaying plasmodesmal permeability. In Plasmodesmata; Springer: Berlin/Heidelberg, Germany, 2015; pp. 149–156. [Google Scholar]
- Zavaliev, R.; Epel, B.L. Imaging callose at plasmodesmata using aniline blue: Quantitative confocal microscopy. In Plasmodesmata; Springer: Berlin/Heidelberg, Germany, 2015; pp. 105–119. [Google Scholar]
- Germain, H.; Gray-Mitsumune, M.; LaFleur, É.; Matton, D.P. ScORK17, a transmembrane receptor-like kinase predominantly expressed in ovules is involved in seed development. Planta 2008, 228, 851–862. [Google Scholar] [CrossRef]
- Bindea, G.; Galon, J.; Mlecnik, B. CluePedia Cytoscape plugin: Pathway insights using integrated experimental and in silico data. Bioinformatics 2013, 29, 661–663. [Google Scholar] [CrossRef]
- Love, M.I.; Huber, W.; Anders, S. Moderated estimation of fold change and dispersion for RNA-seq data with DESeq2. Genome Biol. 2014, 15, 550. [Google Scholar] [CrossRef] [Green Version]
- Yang, J.; Yan, R.; Roy, A.; Xu, D.; Poisson, J.; Zhang, Y. The I-TASSER Suite: Protein structure and function prediction. Nat. Methods 2015, 12, 7–8. [Google Scholar] [CrossRef] [Green Version]
- Kozakov, D.; Hall, D.R.; Xia, B.; Porter, K.A.; Padhorny, D.; Yueh, C.; Beglov, D.; Vajda, S. The ClusPro web server for protein–protein docking. Nat. Protoc. 2017, 12, 255–278. [Google Scholar] [CrossRef]
- Pierce, B.; Hourai, Y.; Weng, Z. Accelerating Protein Docking in ZDOCK Using an Advanced 3D Convolution Library. PLoS ONE 2011, 6, e24657. [Google Scholar] [CrossRef]
- Schneidman-Duhovny, D.; Inbar, Y.; Nussinov, R.; Wolfson, H.J. PatchDock and SymmDock: Servers for rigid and symmetric docking. Nucleic Acids Res. 2005, 33 (Suppl. 2), W363–W367. [Google Scholar] [CrossRef] [Green Version]
- Tovchigrechko, A.; Vakser, I.A. GRAMM-X public web server for protein-protein docking. Nucleic Acids Res. 2006, 34 (Suppl. 2), W310–W314. [Google Scholar] [CrossRef] [PubMed]
- Wang, C.-I.A.; Gunčar, G.; Forwood, J.K.; Teh, T.; Catanzariti, A.-M.; Lawrence, G.J.; Loughlin, F.E.; Mackay, J.; Schirra, H.J.; Anderson, P.A.; et al. Crystal Structures of Flax Rust Avirulence Proteins AvrL567-A and -D Reveal Details of the Structural Basis for Flax Disease Resistance Specificity. Plant Cell 2007, 19, 2898–2912. [Google Scholar] [CrossRef] [Green Version]
- Faulkner, C. Receptor-mediated signaling at plasmodesmata. Front. Plant Sci. 2013, 4, 521. [Google Scholar] [CrossRef] [Green Version]
- Zambryski, P.; Crawford, K. Plasmodesmata: Gatekeepers for Cell-to-Cell Transport of Developmental Signals in Plants. Annu. Rev. Cell Dev. Biol. 2000, 16, 393–421. [Google Scholar] [CrossRef] [Green Version]
- Sun, Y.; Huang, D.; Chen, X. Dynamic regulation of plasmodesmatal permeability and its application to horticultural research. Hortic. Res. 2019, 6, 1–11. [Google Scholar] [CrossRef] [Green Version]
- Han, X.; Huang, L.-J.; Feng, D.; Jiang, W.; Miu, W.; Li, N. Plasmodesmata-Related Structural and Functional Proteins: The Long Sought-After Secrets of a Cytoplasmic Channel in Plant Cell Walls. Int. J. Mol. Sci. 2019, 20, 2946. [Google Scholar] [CrossRef] [Green Version]
- Benitez-Alfonso, Y.; Faulkner, C.; Ritzenthaler, C.; Maule, A.J. Plasmodesmata: Gateways to Local and Systemic Virus Infection. Mol. Plant-Microbe Interact. 2010, 23, 1403–1412. [Google Scholar] [CrossRef] [PubMed]
- Epel, B.L. Plant viruses spread by diffusion on ER-associated movement-protein-rafts through plasmodesmata gated by viral induced host β-1,3-glucanases. Semin. Cell Dev. Biol. 2009, 20, 1074–1081. [Google Scholar] [CrossRef]
- Kankanala, P.; Czymmek, K.; Valent, B. Roles for Rice Membrane Dynamics and Plasmodesmata during Biotrophic Invasion by the Blast Fungus. Plant Cell 2007, 19, 706–724. [Google Scholar] [CrossRef] [Green Version]
- Yamaoka, N.; Ohta, T.; Danno, N.; Taniguchi, S.; Matsumoto, I.; Nishiguchi, M. The role of primary germ tubes in the life cycle of Blumeria graminis: The primary germ tube is responsible for the suppression of resistance induction of a host plant cell. Physiol. Mol. Plant Pathol. 2007, 71, 184–191. [Google Scholar] [CrossRef]
- Zavaliev, R.; Ueki, S.; Epel, B.L.; Citovsky, V. Biology of callose (β-1,3-glucan) turnover at plasmodesmata. Protoplasma 2010, 248, 117–130. [Google Scholar] [CrossRef]
- Sevilem, I.; Miyashima, S.; Helariutta, Y. Cell-to-cell communication via plasmodesmata in vascular plants. Cell Adhes. Migr. 2013, 7, 27–32. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Vatén, A.; Dettmer, J.; Wu, S.; Stierhof, Y.-D.; Miyashima, S.; Yadav, S.R.; Roberts, C.J.; Campilho, A.; Bulone, V.; Lichtenberger, R.; et al. Callose Biosynthesis Regulates Symplastic Trafficking during Root Development. Dev. Cell 2011, 21, 1144–1155. [Google Scholar] [CrossRef] [Green Version]
- Conti, G.; Rodriguez, M.C.; Manacorda, C.A.; Asurmendi, S. Transgenic Expression of Tobacco mosaic virus Capsid and Movement Proteins Modulate Plant Basal Defense and Biotic Stress Responses in Nicotiana tabacum. Mol. Plant-Microbe Interact. 2012, 25, 1370–1384. [Google Scholar] [CrossRef] [Green Version]
- Wu, S.; Gallagher, K.L. Mobile protein signals in plant development. Curr. Opin. Plant Biol. 2011, 14, 563–570. [Google Scholar] [CrossRef]
- De Storme, N.; De Schrijver, J.; Van Criekinge, W.; Wewer, V.; Dörmann, P.; Geelen, D. GLUCAN SYNTHASE-LIKE8 and STEROL METHYLTRANSFERASE2 Are Required for Ploidy Consistency of the Sexual Reproduction System in Arabidopsis. Plant Cell 2013, 25, 387–403. [Google Scholar] [CrossRef] [Green Version]
- Grison, M.S.; Kirk, P.; Brault, M.L.; Na Wu, X.; Schulze, W.X.; Benitez-Alfonso, Y.; Immel, F.; Bayer, E.M. Plasma Membrane-Associated Receptor-like Kinases Relocalize to Plasmodesmata in Response to Osmotic Stress. Plant Physiol. 2019, 181, 142–160. [Google Scholar] [CrossRef] [Green Version]
- Su, S.; Liu, Z.; Chen, C.; Zhang, Y.; Wang, X.; Zhu, L.; Miao, L.; Wang, X.-C.; Yuan, M. Cucumber Mosaic Virus Movement Protein Severs Actin Filaments to Increase the Plasmodesmal Size Exclusion Limit in Tobacco. Plant Cell 2010, 22, 1373–1387. [Google Scholar] [CrossRef] [Green Version]
- Xian, L.; Yu, G.; Wei, Y.; Rufian, J.S.; Li, Y.; Zhuang, H.; Xue, H.; Morcillo, R.J.; Macho, A.P. A Bacterial Effector Protein Hijacks Plant Metabolism to Support Pathogen Nutrition. Cell Host Microbe 2020, 28, 548–e7. [Google Scholar] [CrossRef] [PubMed]







Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Rahman, M.S.; Madina, M.H.; Plourde, M.B.; dos Santos, K.C.G.; Huang, X.; Zhang, Y.; Laliberté, J.-F.; Germain, H. The Fungal Effector Mlp37347 Alters Plasmodesmata Fluxes and Enhances Susceptibility to Pathogen. Microorganisms 2021, 9, 1232. https://doi.org/10.3390/microorganisms9061232
Rahman MS, Madina MH, Plourde MB, dos Santos KCG, Huang X, Zhang Y, Laliberté J-F, Germain H. The Fungal Effector Mlp37347 Alters Plasmodesmata Fluxes and Enhances Susceptibility to Pathogen. Microorganisms. 2021; 9(6):1232. https://doi.org/10.3390/microorganisms9061232
Chicago/Turabian StyleRahman, Md. Saifur, Mst Hur Madina, Mélodie B. Plourde, Karen Cristine Gonçalves dos Santos, Xiaoqiang Huang, Yang Zhang, Jean-François Laliberté, and Hugo Germain. 2021. "The Fungal Effector Mlp37347 Alters Plasmodesmata Fluxes and Enhances Susceptibility to Pathogen" Microorganisms 9, no. 6: 1232. https://doi.org/10.3390/microorganisms9061232