
Black Fungi and Hydrocarbons: An Environmental Survey for Alkylbenzene Assimilation

 , and
, and
Abstract
:1. Introduction
2. Materials and Methods
2.1. Isolation and Preservation of Black Fungal Strains
- (a)
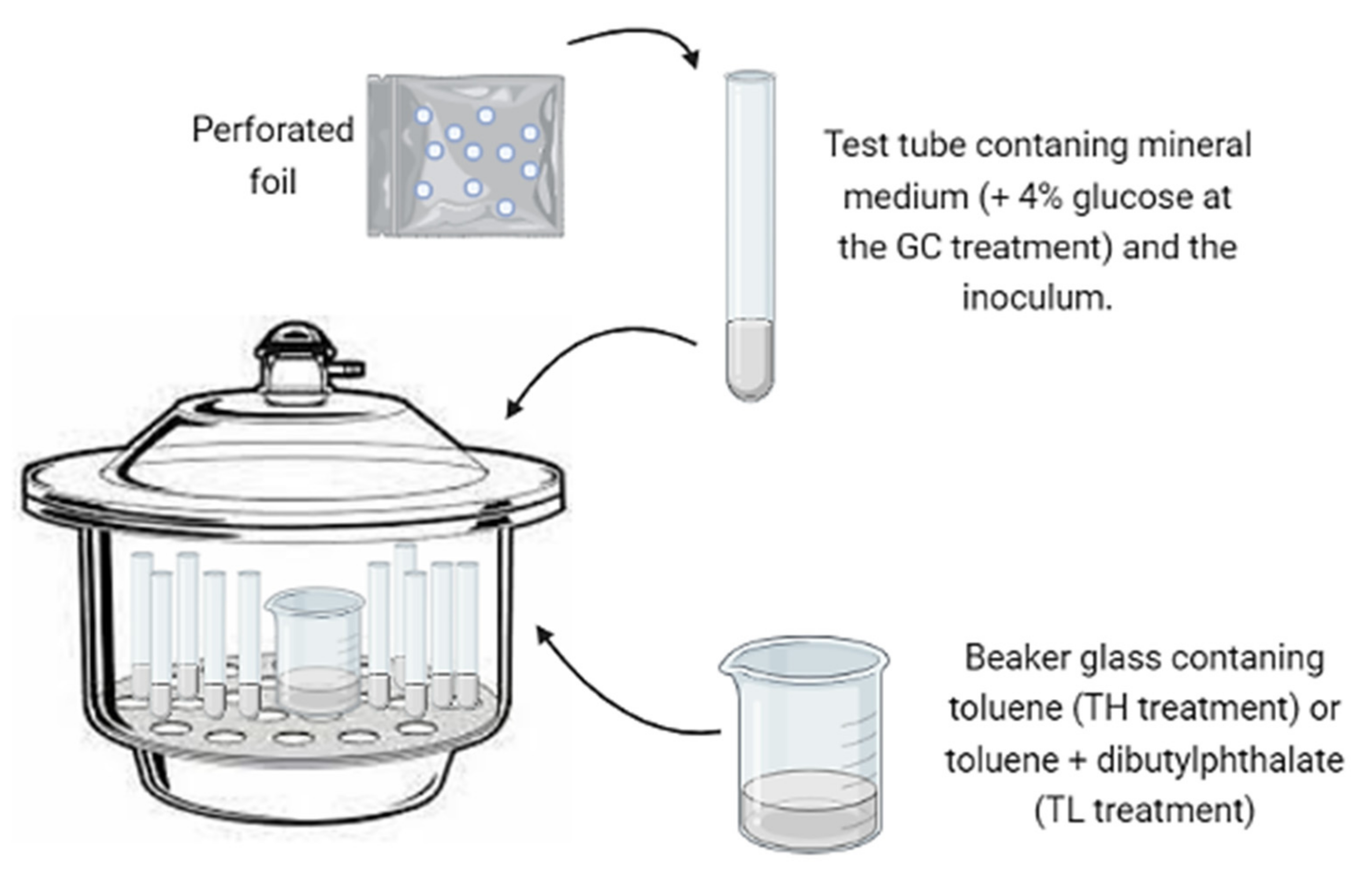
- Oil flotation technique: This isolation method is known to be highly selective for hydrophobic fungi such as black yeasts [32]. Soil samples were taken from a garage shop (city of Rio Claro, São Paulo, Brazil) contaminated with motor oil from vehicles and from land farming soil from an oil refinery (city of Paulínia, São Paulo, Brazil). This method was also applied on bark fragments of Eucalyptus tereticornis and exoskeletons of gynes and drones (winged females and males of leaf-cutting ants) of Atta capiguara and Atta laevigata obtained during the sexual reproduction period known as the “mating flight” (city of Botucatu, São Paulo, Brazil).
- (b)
- Standard serial dilution method: Water samples from a river under the influence of an oil refinery (Atibaia River, São Paulo, Brazil) were processed according to the standard methods [33]. Fungi were isolated through serial dilutions of the samples, followed by plating on potato dextrose and Sabouraud agar (PDA and SA).
- (c)
- Agar walk method: Living individuals of the previously mentioned winged females and males of leaf-cutting ants (Atta capiguara and Atta laevigata) were placed on malt yeast agar plates (MYA; peptic digest of animal tissue, 5.0 g; yeast extract, 3.0 g; malt extract, 3.0 g; dextrose, 10.0 g; agar, 20.0 g per liter of distilled water) added with cycloheximide (500.0 mg), chloramphenicol (200.0 mg), and streptomycin (200.0 mg), and were allowed to walk on the agar for one hour. Then they were removed, and the plates were incubated at 25 °C until the emergence of the black colonies.
2.2. Morphological Analysis
2.3. Molecular Identification
2.4. Biomass Production in Toluene Atmospheres
2.5. Selection of Strains with Toluene Assimilation Potential
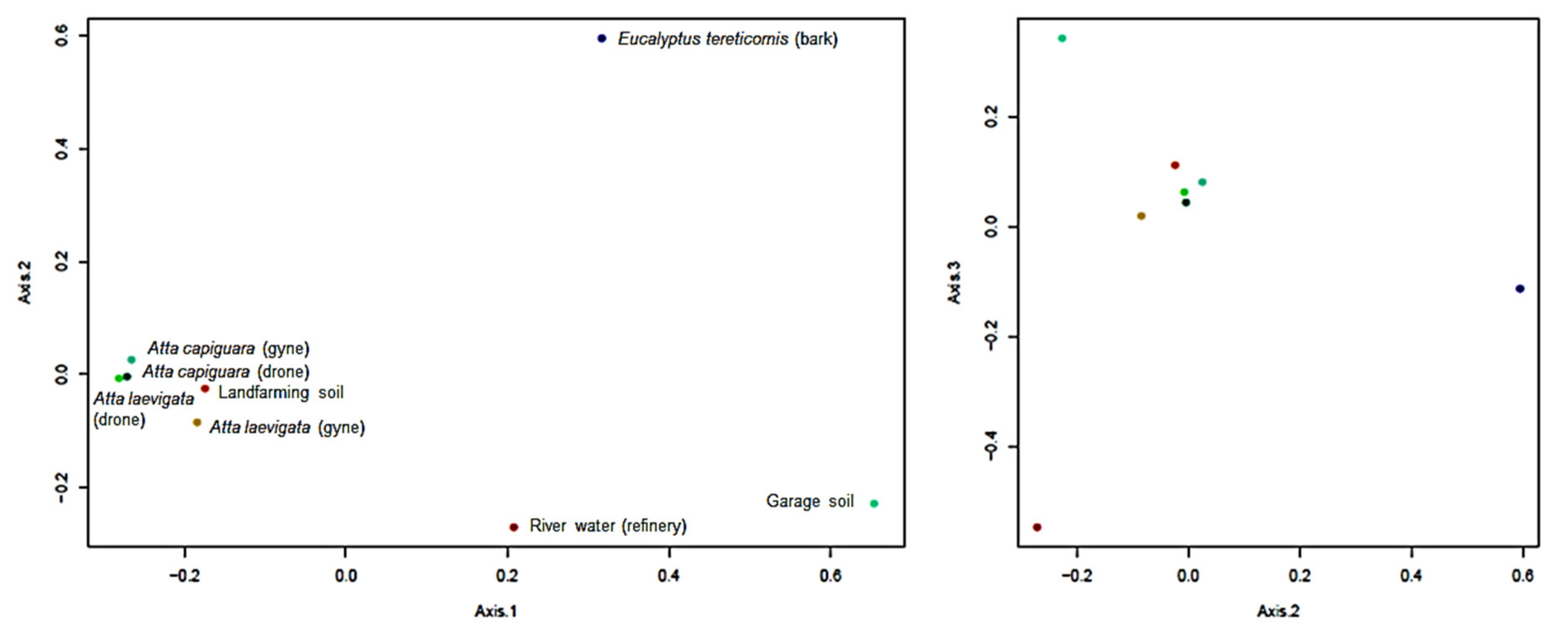
2.6. Multivariate Ecological Analysis
3. Results and Discussion
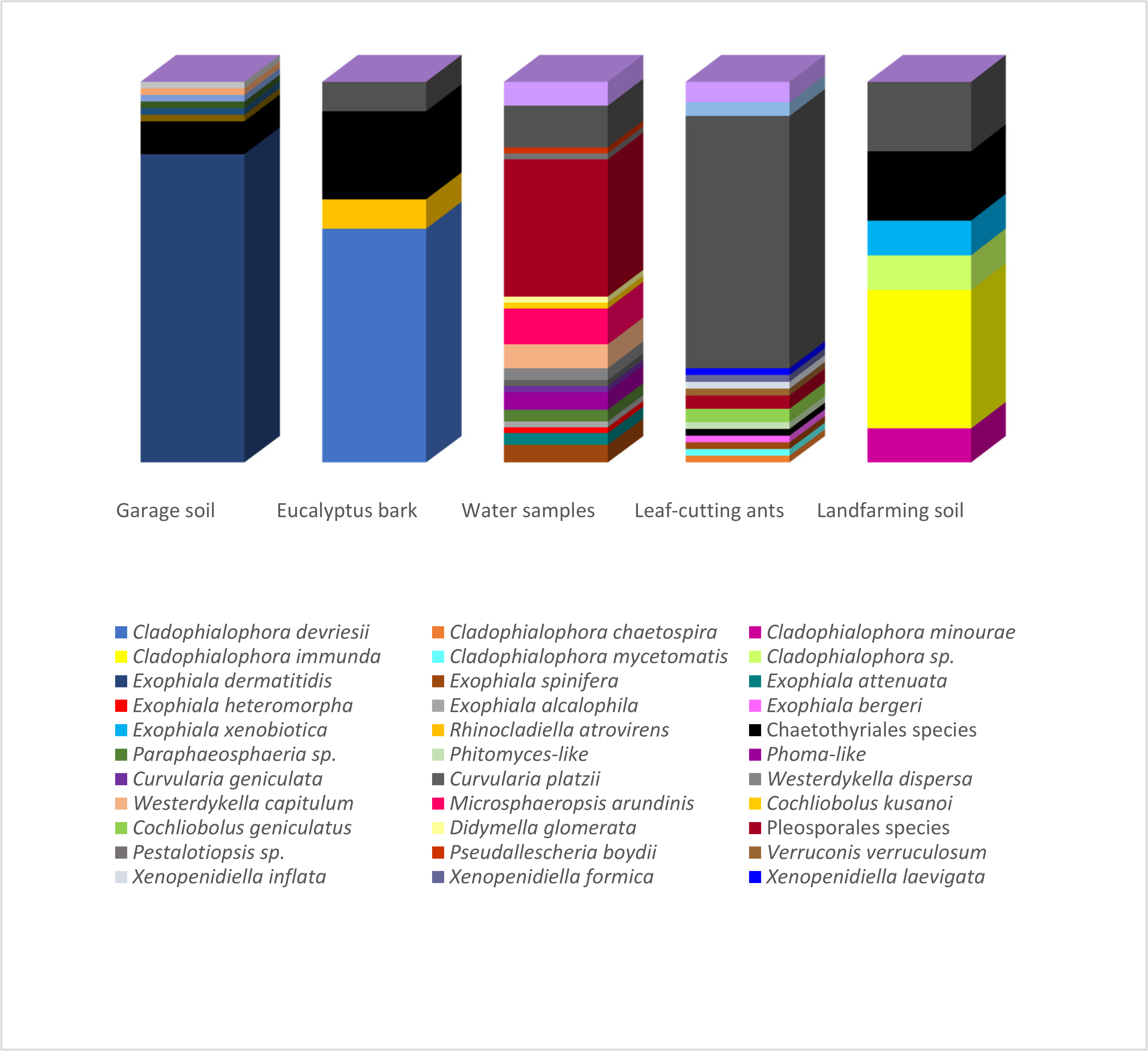
3.1. Isolation and Identification
3.2. Fungal Community Structure
3.3. Growth Specificity Experiments
4. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Mitra, S.; Roy, P. BTEX: A Serious Ground-water Contaminant. Res. J. Environ. Sci. 2011, 5, 394–398. [Google Scholar] [CrossRef] [Green Version]
- Environmental Protection Agency’s (EPA). Toluene. Available online: https://www.epa.gov/sites/production/files/2016-09/documents/toluene.pdf (accessed on 30 October 2020).
- Hobara, T.; Okuda, M.; Gotoh, M.; Oki, K.; Segawa, H.; Kunitsugu, I. Estimation of the Lethal Toluene Concentration from the Accidental Death of Painting Workers. Ind. Health 2000, 38, 228–231. [Google Scholar] [CrossRef] [PubMed]
- Melquiades, R.A.; Lobo, I.; Guedes, C.L.B.; Pinto, J.P. Análise de benzeno, tolueno, etilbenzeno e xilenos em solos por headspace e cromatografia gasosa/detector de ionização de chama. Semin. Ciências Exatas Tecnológicas Londrina 2006, 27, 113–120. [Google Scholar] [CrossRef] [Green Version]
- Wilbur, S.M.A.; Bosh, S.B.S.U.S. Interaction Profile for: Benzene, Toluene, Ethylbenzene, and Xylenes (BTEX); Department of Health and Human Services Public Health Service Agency for Toxic Substances and Disease Registry: Atlanta, GA, USA, 2004. [Google Scholar]
- Occupational Safety and Health Administration (OSHA). Toluene: Occupational Exposure Limits. 2012. Available online: https://www.osha.gov/toluene/occupational-exposure-limits (accessed on 30 October 2020).
- ITOPF. Oil Tanker Spill Statistics. 2018. Available online: http://www.itopf.org/fileadmin/data/Documents/Company_Lit/Oil_Spill_Stats_2018.pdf (accessed on 30 March 2019).
- Brito, N.N.; Zamora, P.P.; Neto, A.L.O.; De Battisti, A.; Paterniani, J.E.S.; Pelegrini, R.T. Utilização de fungos na remediação de efluentes industriais. In IV Fórum de Estudos Contábeis; Faculdades Integradas Claretianas: Rio Claro, SP, Brazil, 2002; pp. 18–22. [Google Scholar]
- Brandão, L.F.P.; Suarez, P.A.Z. Determination of the alternative butanol/gasoline and butanol/diesel fuel blends heats of combustion by a heat-loss compensated semi-microcalorimeter. J. Therm. Anal. Calorim. 2018, 132, 1953–1960. [Google Scholar] [CrossRef]
- Silva, R.L.B.; Barra, C.M.; Monteiro, T.C.N.; Brilhante, O.M. Estudo da contaminação de poços rasos por combustíveis orgânicos e possíveis conseqüências para a saúde pública no município de Itaguaí, Rio de Janeiro, Brasil. Cad. Saúde Públ. 2002, 18, 1599–1607. [Google Scholar] [CrossRef] [Green Version]
- CIMA—Conselho Interministerial do Açúcar e do Álcool. 2013. Available online: http://nxt.anp.gov.br/NXT/gateway.dll/leg/folder_resolucoes/resolucoes_cima/2013/rcima%201%20-%202013.xml?f=templatesfn=document−frame.htmfn=document−frame.htm3.0q=q=x=$nc=9930. (accessed on 8 March 2018).
- Alexander, M. Biodegradation and Bioremediation, 2nd ed.; Academic Press: New York, NY, USA, 1999. [Google Scholar]
- Singh, H. Mycorremediation: Fungal Bioremediation; Wiley-Interscience, John Wiley & Sons, Inc., Publication: Hoboken, NJ, USA, 2006. [Google Scholar]
- Harms, H.; Schlosser, D.; Wick, L.Y. Untapped potential: Exploiting fungi in bioremediation of hazardous chemicals. Nat. Rev. Genet. 2011, 9, 177–192. [Google Scholar] [CrossRef]
- Barrech, D.; Ali, I.; Tareen, M. A Review on Mycoremediation—The fungal bioremediation. Pure Appl. Biol. (PAB) 2018, 7, 343–348. [Google Scholar] [CrossRef]
- Prenafeta-Boldú, F.X.; de Hoog, G.S.; Summerbell, R.C. Fungal Communities in Hydrocarbon Degradation. In Microbial Communities Utilizing Hydrocarbons and Lipids: Members, Metagenomics and Ecophysiology; McGenity, T.J., Ed.; Springer Nature Switzerland AG: Cham, Switzerland, 2019; pp. 1–36. [Google Scholar]
- De Hoog, G.S.; Zeng, J.S.; Harrak, M.J.; Sutton, D.A. Exophiala xenobiotica sp. nov., an opportunistic black yeast inhabiting environments rich in hydrocarbons. Antonie Leeuwenhoek 2006, 90, 257–268. [Google Scholar] [CrossRef]
- Prenafeta-Boldú, F.X.; Kuhn, A.; Luykx, D.M.; Anke, H.; van Groenestijn, J.W.; de Bont, J.A. Isolation and characterisation of fungi growing on volatile aromatic hydrocarbons as their sole carbon and energy source. Mycol. Res. 2001, 105, 477–484. [Google Scholar] [CrossRef]
- Prenafeta-Boldú, F.X.; Summerbell, R.; De Hoog, G.S. Fungi growing on aromatic hydrocarbons: Biotechnology’s unex-pected encounter with biohazard. FEMS Microbiol Rev. 2006, 30, 109–130. [Google Scholar] [CrossRef] [Green Version]
- Zhao, J.; Zeng, J.; De Hoog, G.S.; Attili-Angelis, D.; Prenafeta-Boldú, F.X. Isolation and Identification of Black Yeasts by Enrichment on Atmospheres of Monoaromatic Hydrocarbons. Microb. Ecol. 2010, 60, 149–156. [Google Scholar] [CrossRef] [Green Version]
- Isola, D.; Selbmann, L.; De Hoog, G.S.; Fenice, M.; Onofri, S.; Prenafeta-Boldú, F.X.; Zucconi, L. Isolation and Screening of Black Fungi as Degraders of Volatile Aromatic Hydrocarbons. Mycopathologia 2013, 175, 369–379. [Google Scholar] [CrossRef]
- Gostinčar, C.; Muggia, L.; Grube, M. Polyextremotolerant black fungi: Oligotrophism, adaptive potential, and a link to lichen symbioses. Front. Microbiol. 2012, 3, 390. [Google Scholar] [CrossRef] [Green Version]
- Cordero, R.J.B.; Vij, R.; Casadevall, A. Microbial melanins for radioprotection and bioremediation. Microb. Biotechnol. 2017, 10, 1186–1190. [Google Scholar] [CrossRef] [Green Version]
- Gueidan, C.; Ruibal, C.; De Hoog, S.; Schneider, H. Rock-inhabiting fungi originated during periods of dry climate in the late Devonian and middle Triassic. Fungal Biol. 2011, 115, 987–996. [Google Scholar] [CrossRef]
- Gostinčar, C.; Grube, M.; Gunde-Cimerman, N. Evolution of Fungal Pathogens in Domestic Environments? Fungal Biol. 2011, 115, 1008–1018. [Google Scholar] [CrossRef]
- Gümral, R.; Tümgör, A.; Saraçlı, M.A.; Yıldıran, Ş.T.; Ilkit, M.; de Hoog, G.S. Black yeast diversity on creosoted railway sleepers changes with ambient climatic conditions. Microb. Ecol. 2014, 68, 699–707. [Google Scholar] [CrossRef]
- Nascimento, M.M.; Vicente, V.A.; Bittencourt, J.V.; Gelinski, J.M.L.; Prenafeta-Boldú, F.X.; Romero-Güiza, M.; Fornari, G.; Gomes, R.R.; Santos, G.D.; Ende, A.G.V.D.; et al. Diversity of opportunistic black fungi on babassu coconut shells, a rich source of esters and hydrocarbons. Fungal Biol. 2017, 121, 488–500. [Google Scholar] [CrossRef]
- Badali, H.; Prenafeta-Boldú, F.X.; Guarro, J.; Klaassen, C.H.; Meis, J.F.; De Hoog, G.S. Cladophialophora psammophila, a novel species of Chaetothyriales with a potential use in the bioremediation of volatile aromatic hydrocarbons. Fungal Biol. 2011, 115, 1019–1029. [Google Scholar] [CrossRef]
- Prenafeta-Boldú, F.X.; Illa, J.; Van Groenestijn, J.W.; Flotats, X. Influence of synthetic packing materials on the gas dispersion and biodegradation kinetics in fungal air biofilters. Appl. Microbiol. Biotechnol. 2008, 79, 319–327. [Google Scholar] [CrossRef] [Green Version]
- Teixeira, M.M.; Moreno, L.F.; Stielow, B.; Muszewska, A.; Hainaut, M.; Gonzaga, L.; Abouelleil, A.; Patané, J.S.L.; Priest, M.; Souza, R.; et al. Exploring the genomic diversity of black yeasts and relatives (Chaetothyriales, Ascomycota). Stud. Mycol. 2017, 86, 1–28. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Blasi, B.; Tafer, H.; Kustor, C.; Poyntner, C.; Lopandic, K.; Sterflinger, K. Genomic and transcriptomic analysis of the toluene degrading black yeast Cladophialophora immunda. Sci. Rep. 2017, 7, 11436. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Satow, M.M.; Attili-Angelis, D.; De Hoog, G.S.; Vicente, V.A. Selective factors involved in oil flotation isolation of black yeasts from the environment. Stud. Mycol. 2008, 61, 157–163. [Google Scholar] [CrossRef] [PubMed]
- Standard Methods for the Examination of Water and Wastewater, 19th ed.; American Public Health Association: Washington, DC, USA, 1998.
- Möller, E.M.; Bahnweg, G.; Sandermann, H.; Geiger, H.H. A simple and efficient protocol for isolation of high molecular weight DNA from filamentous fungi, fruit bodies, and infected plant tissues. Nucleic Acids Res. 1992, 20, 6115–6116. [Google Scholar] [CrossRef] [Green Version]
- Gerrits Van Den Ende, A.H.G.; De Hoog, G.S. Variability and molecular diagnostics of the neurotropic species Cladophialophora bantiana. Stud. Mycol. 1999, 43, 151–162. [Google Scholar]
- White, T.J.; Bruns, T.D.; Lee, S.; Taylor, J. Analysis of phylogenetic relationships by amplification and direct sequencing of ribosomal RNA genes. In PCR Protocols: A Guide to Methods and Applications; Innis, M.A., Gelfand, D.H., Sninsky, J.J., White, T.J., Eds.; Academic Press: New York, NY, USA, 1990; pp. 315–322. [Google Scholar]
- Hall, T.A. BioEdit: A User-Friendly Biological Sequence Alignment Editor and Analysis Program for Windows 95/98/NT; Nucleic Acids Symposium Series; Information Retrieval Ltd.: London, UK, 1999; Volume 41, pp. 95–98. [Google Scholar]
- Altschul, S.F.; Gish, W.; Miller, W.; Myers, E.W.; Lipman, D.J. Basic local alignment search tool. J. Mol. Biol. 1990, 215, 403–410. [Google Scholar] [CrossRef]
- Larkin, M.A.; Blackshields, G.; Brown, N.P.; Chenna, R.; McGettigan, P.A.; McWilliam, H.; Valentin, F.; Wallace, I.M.; Lopez, R.; Thompson, J.D.; et al. ClustalW and ClustalX version 2. Bioinformatics 2007, 23, 2947–2948. [Google Scholar] [CrossRef] [Green Version]
- Kumar, S.; Stecher, G.; Li, M.; Knyaz, C.; Tamura, K. MEGA X: Molecular evolutionary genetics analysis across computing platforms. Mol. Biol. Evol. 2018, 35, 1547–1549. [Google Scholar] [CrossRef]
- Kimura, M. A simple method for estimating evolutionary rates of base substitutions through comparative studies of nucleotide sequences. J. Mol. Evol. 1980, 16, 111–120. [Google Scholar] [CrossRef]
- Amoore, J.E.; Hautala, E. Odor as an aid to chemical safety: Odor thresholds compared with threshold limit values and volatilities for 214 industrial chemicals in air and water dilution. J. Appl. Toxicol. 1983, 3, 272–290. [Google Scholar] [CrossRef]
- Huse, S.M.; Welch, D.B.M.; Voorhis, A.; Shipunova, A.; Morrison, H.G.; Eren, A.M.; Sogin, M.L. VAMPS: A website for visualization and analysis of microbial population structures. BMC Bioinform. 2014, 15, 41. [Google Scholar] [CrossRef] [Green Version]
- De Hoog, G.S.; Queiroz-Telles, F.; Haase, G.; Fernandez-Zeppenfeldt, G.; Angelis, D.A.; Van Den Ende, A.; Matos, T.; Peltroche-Llacsahuanga, H.; Pizzirani-Kleiner, A.A.; Rainer, J.; et al. Black fungi: Clinical and pathogenic approaches. Med. Mycol. 2000, 38, 243–250. [Google Scholar] [CrossRef]
- De Hoog, G.S.; Vicente, V.A.; Gorbushina, A.A. The bright future of darkness—The rising power of black fungi: Black yeasts, microcolonial fungi, and their relatives. Mycopathologia 2013, 175, 365–368. [Google Scholar] [CrossRef] [Green Version]
- Moreno, L.F.; Ahmed, A.A.O.; Brankovics, B.; Cuomo, C.A.; Menken, S.B.J.; Taj-Aldeen, S.J.; Faidah, H.; Stielow, J.B.; Teixeira, M.D.M.; Prenafeta-Boldú, F.X.; et al. Genomic Understanding of an Infectious Brain Disease from the Desert. G3 Genes Genomes Genet. 2018, 8, 909–922. [Google Scholar] [CrossRef] [Green Version]
- Moreno, L.F.; Mayer, V.; Voglmayr, H.; Blatrix, R.; Stielow, J.B.; Teixeira, M.M.; Vicente, V.A.; De Hoog, S. Genomic analysis of ant domatia-associated melanized fungi (Chaetothyriales, Ascomycota). Mycol. Prog. 2019, 18, 541–552. [Google Scholar] [CrossRef] [Green Version]
- Badali, H.; Gueidan, C.; Najafzadeh, M.J.; Bonifaz, A.; Gerrits Van Den Ende, A.H.G.; De Hoog, G.S. Biodiversity of Cladophialophora. Stud. Mycol. 2008, 61, 175–191. [Google Scholar] [CrossRef]
- De Hoog, G.S.; Nishikaku, G.; Fernandez-Zeppenfeldt, C.; Padín Gonzáles, E.; Badali, H.; Richar-Yegres, N.; Gerrits Van Den Ende, H.G. Molecular nalysis and pathogenicity of the Cladophialophora carrionii complex, with the description of a novel species. Stud. Mycol. 2007, 58, 219–234. [Google Scholar] [CrossRef]
- Crous, P.W.; Braun, U.; Schubert, K.; Groenewald, J.Z. The genus Cladosporium and similar dematiaceous hypho-mycetes. Preface Stud. Mycol. 2007, 58, 253. [Google Scholar]
- Madrid, H.; Hernandez-Restrepo, M.; Gené, J.; Cano, J.; Guarro, J.; Silva, V. New and interesting chaetothyrialean fungi from Spain. Mycol. Prog. 2016, 15, 1179–1201. [Google Scholar] [CrossRef] [Green Version]
- Das, K.; Lee, S.-Y.; Jung, H.-Y. Cladophialophora lanosa sp. nov., a New Species Isolated from Soil. Mycobiology 2019, 47, 173–179. [Google Scholar] [CrossRef]
- Nunes, T.; Pio, C. Emission of volatile organic compounds from Portuguese eucalyptus forests. Chemosphere Glob. Chang. Sci. 2001, 3, 239–248. [Google Scholar] [CrossRef]
- Winters, A.J.; Adams, M.A.; Bleby, T.M.; Rennenberg, H.; Steigner, D.; Steinbrecher, R.; Kreuzwieser, J. Emissions of isoprene, monoterpene and short-chained carbonyl compounds from Eucalyptus spp. in southern Australia. Atmos. Environ. 2009, 43, 3035–3043. [Google Scholar] [CrossRef]
- Napolitano, R.; Juaréz, N.P. Entomopathogenous fungi degrade epicuticular hydrocarbons of Triatoma infestans. Arch. Biochem. Biophys. 1997, 344, 208–214. [Google Scholar] [CrossRef]
- Attili-Angelis, D.; Duarte AP, M.; Pagnocca, F.C.; Nagamoto, N.S.; De Vries, M.; Stielow, J.B.; de Hoog, G.S. Novel Phialophora species from leaf-cutting ants (tribe Attini). Fungal Divers. 2014, 65, 65–75. [Google Scholar] [CrossRef]
- Sudhadham, M.; Prakitsin, S.; Sivichai, S.; Chaiyarat, R.; Dorrestein, G.; Menken, S.; de Hoog, G. The neurotropic black yeast Exophiala dermatitidis has a possible origin in the tropical rain forest. Stud. Mycol. 2008, 61, 145–155. [Google Scholar] [CrossRef]
- Li, D.M.; Li, R.Y.; De Hoog, G.; Wang, Y.X.; Wang, D.L. Exophiala asiatica, a new species from a fatal case in China. Med. Mycol. 2009, 47, 101–109. [Google Scholar] [CrossRef] [Green Version]
- Del Palacio-Hernanz, A.; Moore, M.K.; Campbell, C.K.; Del Palacio-Perez-Medel, A.; Del Castillo-Cantero, R. Infection of the central nervous system by Rhinocladiella atrovirens in a patient with acquired immunodeficiency syndrome. J. Med. Vet. Mycol. 1989, 27, 127–130. [Google Scholar] [CrossRef]
- Fukasawa, Y. Fungal succession and decomposition of Pinus densiflora snags. Ecol. Res. 2018, 33, 435–444. [Google Scholar] [CrossRef]
- Szewczyk, W.; Kwaśna, H.; Behnke-Borowczyk, J. Fungi inhabiting knotwood of Pinus sylvestris infected by Porodaedalea pini. J. Phytopathol. 2017, 165, 500–507. [Google Scholar] [CrossRef]
- Ruibal, C.; Gonzalo, P.; Bills, G.F. Isolation and characterization of melanized fungi from limestone formations in Mallorca. Mycol. Prog. 2005, 4, 23–38. [Google Scholar] [CrossRef]
- Vicente, V.; Attili-Angelis, D.; Pie, M.; Queiroz-Telles, F.; Cruz, L.; Najafzadeh, M.; de Hoog, G.; Zhao, J.; Pizzirani-Kleiner, A. Environmental isolation of black yeast-like fungi involved in human infection. Stud. Mycol. 2008, 61, 137–144. [Google Scholar] [CrossRef] [PubMed]
- Vicente, V.A.; Angelis, D.A.D.; Queiróz-Telles Filho, F.; Pizzirani-Kleiner, A.A. Isolamento de fungos herpotriquieláceos do ambiente. Braz. J. Microbiol. 2001, 32, 47–51. [Google Scholar]
- Kirk, P.M.; Cannon, P.F.; Minter, D.W.; Staplers, J.A. Dictionary of the Fungi, 10th ed.; CABI Bioscience: Wallingford, UK, 2008. [Google Scholar]
- Zhang, Y.; Crous, P.W.; Schoch, C.L.; Bahkali, A.H.; Guo, L.D.; Hyde, K.D. A molecular, morphological and ecological re-appraisal of Venturiales—A new order of Dothideomycetes. Fungal Divers. 2011, 51, 249–277. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Boerema, H.G.; de Gruyter, J.; Noordeloos, E.M. Contributions towards a monograph of Phoma (Coelomycetes)—IV. Section Heterospora: Taxa with large sized conidial dimorphs, in vivo sometimes as Stagonosporopsis synanamorphs. Pers. Mol. Phylogeny Evol. Fungi 1997, 16, 335–371. [Google Scholar]
- Boerema, G.H.; Gruyter, J.; Noordeloos, M.E.; Hamers, M.E.C. Phoma Identification Manual: Differentiation of Specific and Infra-Specific Taxa in C. Phoma Identification Manual: Differentiation of Specific and Infra-Specific Taxa in Culture; CABI: Wallingford, UK, 2004. [Google Scholar]
- de Gruyter, J.; Woudenberg, J.; Aveskamp, M.; Verkley, G.; Groenewald, J.; Crous, P. Redisposition of phoma-like anamorphs in Pleosporales. Stud. Mycol. 2013, 75, 1–36. [Google Scholar] [CrossRef] [Green Version]
- Ebead, G.A.; Overy, D.P.; Berrué, F.; Kerr, R.G. Westerdykella reniformis sp. nov., producing the antibiotic me-tabolites melinacidin IV and chetracin B. IMA Fungus 2012, 3, 189–201. [Google Scholar] [CrossRef]
- Guedes, F.L.A.; Attili-Angelis, D.; Pagnocca, F.C. Selective isolation of dematiaceous fungi from the workers of Atta laevigata (Formicidae: Attini). Folia Microbiol. 2011, 57, 21–26. [Google Scholar] [CrossRef]
- Dunn, M.; Domsch, K.H.; Gams, W.; Anderson, T.-H. Compendium of Soil Fungi. TAXON 1982, 31, 600. [Google Scholar] [CrossRef]
- Alves, J.L.; Barreto, R.W.; Pereira, O.L.; Soares, D.J. Additions to the mycobiota of the invasive weed Miconia calvescens (Melastomataceae). Mycologia 2010, 102, 69–82. [Google Scholar] [CrossRef]
- Costa, I.P.M.W.; Maia, L.C.; Cavalcanti, M.A. Diversity of leaf endophytic fungi in mangrove plants of Northeast Brazil. Braz. J. Microbiol. 2012, 43, 1165–1173. [Google Scholar] [CrossRef] [Green Version]
- Luo, J.; Liu, X.; Li, E.; Guo, L.; Che, Y. Arundinols A–C and Arundinones A and B from the Plant Endophytic FungusMicrosphaeropsis arundinis. J. Nat. Prod. 2012, 76, 107–112. [Google Scholar] [CrossRef] [PubMed]
- Luedemann, G.M. The Dictyochlamydospore of Peyronellaea glomerata (Corda) Goidanich ex Togliani Contrasted with the Dictyoporospore of Alternaria Tenuis Auct. Mycologia 1959, 51, 772–780. [Google Scholar] [CrossRef]
- Deng, J.X.; Paul, N.C.; Li, M.J.; Seo, E.Y.; Sung, G.H.; Yu, S.H. Molecular Characterization and Morphology of Two Endophytic Peyronellaea Species from Pinus koraiensis in Korea. Mycobiology 2011, 39, 266–271. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Abdollahzadeh, J.; Groenewald, J.Z.; Coetzee MP, A.; Wingfield, M.J.; Crous, P.W. Evolution of lifestyles in Capnodiales. Stud. Mycol. 2020, 95, 381–414. [Google Scholar] [CrossRef]
- Crous, P.; Summerell, B.; Carnegie, A.; Wingfield, M.; Groenewald, J. Novel species of Mycosphaerellaceae and Teratosphaeriaceae. Persoonia Mol. Phylogeny Evol. Fungi 2009, 23, 119–146. [Google Scholar] [CrossRef] [Green Version]
- Bensch, K.; Groenewald, J.; Dijksterhuis, J.; Starink-Willemse, M.; Andersen, B.; Summerell, B.; Shin, H.-D.; Dugan, F.; Schroers, H.-J.; Braun, U.; et al. Species and ecological diversity within the Cladosporium cladosporioides complex (Davidiellaceae, Capnodiales). Stud. Mycol. 2010, 67, 1–94. [Google Scholar] [CrossRef]
- Rodrigues, A.; Cable, R.N.; Mueller, U.G.; Bacci, M., Jr.; Pagnocca, F.C. Antagonistic interactions between garden yeasts and microfungal garden pathogens of leafcutting ants. Antonie Leeuwenhoek 2009, 96, 331–342. [Google Scholar] [CrossRef]
- Rodrigues, A.; Mueller, U.G.; Ishak, H.D.; Bacci, M., Jr.; Pagnocca, F.C.; Bacci, M. Ecology of microfungal communities in gardens of fungus-growing ants (Hymenoptera: Formicidae): A year-long survey of three species of attine ants in Central Texas. FEMS Microbiol. Ecol. 2011, 78, 244–255. [Google Scholar] [CrossRef] [Green Version]
- Pagnocca, F.C.; Rodrigues, A.; Nagamoto, N.S.; Bacci, M. Yeasts and filamentous fungi carried by the gynes of leaf-cutting ants. Antonie Leeuwenhoek 2008, 94, 517–526. [Google Scholar] [CrossRef]
- Mullins, J. Microorganisms in outdoor air. In Microorganisms in Home and Indoor Work Environments: Diversity, Health Impacts, Investigation and Control, 1st ed.; Flannigan, B., Samson, R.A., Miller, J.D., Eds.; Taylor & Francis: London, UK, 2001; pp. 3–16. [Google Scholar]
- Bensch, K.; Braun, U.; Groenewald, J.; Crous, P. The genus Cladosporium. Stud. Mycol. 2012, 72, 1–401. [Google Scholar] [CrossRef] [Green Version]
- Bensch, K.; Groenewald, J.Z.; Braun, U.; Dijksterhuis, J.; de Jesús Yáñez-Morales, M.; Crous, P.W. Common but dif-ferent: The expanding realm of Cladosporium. Stud. Mycol. 2015, 82, 23–74. [Google Scholar] [CrossRef] [Green Version]
- Sandoval-Denis, M.; Gené, J.; Sutton, D.; Wiederhold, N.; Cano-Lira, J.; Guarro, J. New species of Cladosporium associated with human and animal infections. Persoonia Mol. Phylogeny Evol. Fungi 2016, 36, 281–298. [Google Scholar] [CrossRef] [Green Version]
- Bensch, K.; Groenewald, J.Z.; Meijer, M.; Dijksterhuis, J.; Jurjević, Ž.; Andersen, B.; Samson, R.A. Cladosporium species in indoor environments. Stud. Mycol. 2018, 89, 177–301. [Google Scholar] [CrossRef]
- Quaedvlieg, W.; Binder, M.; Groenewald, J.Z.; Summerell, B.A.; Carnegie, A.J.; Burgess, T.I.; Crous, P.W. Intro-ducing the Consolidated Species Concept to resolve species in the Teratosphaeriaceae. Persoonia Mol. Phylogeny Evol. Fungi 2014, 33, 1. [Google Scholar] [CrossRef] [Green Version]
- Duarte, A.; Attili-Angelis, D.; Baron, N.; Groenewald, J.; Crous, P.; Pagnocca, F. Riding with the ants. Persoonia Mol. Phylogeny Evol. Fungi 2017, 38, 81–99. [Google Scholar] [CrossRef] [Green Version]
- Benny, G.L.; Kimbrough, J.W. A synopsis of the orders and families of Plectomycetes with keys to genera. Mycotaxon 1980, 12, 1–91. [Google Scholar]
- Bibashi, E.; De Hoog, G.S.; Kostopoulou, E.; Tsivitanidou, M.; Sevastidou, J.; Geleris, P. Invasive infection caused by Pseudallescheria boydii in an immunocompetent patient. Hippokratia 2009, 13, 184–186. [Google Scholar]
- Martin-Sanchez, P.M.; Gorbushina, A.A.; Toepel, J. Quantification of microbial load in diesel storage tanks using culture- and qPCR-based approaches. Int. Biodet. Biodegr. 2016, 126, 216–223. [Google Scholar] [CrossRef]
- Cazarolli, J.C.; Guzatto, R.; Samios, D.; Peralba, M.D.C.R.; Cavalcanti, E.H.D.S.; Bento, F.M. Susceptibility of linseed, soybean, and olive biodiesel to growth of the deteriogenic fungus Pseudallescheria boydii. Int. Biodeterior. Biodegrad. 2014, 95, 364–372. [Google Scholar] [CrossRef]
- Janda-Ulfig, K.; Ulfig, K.; Cano, J.; Guarro, J. A study of the growth of Pseudallescheria boydii isolates from sewage sludge and clinical sources on tributyrin, rapeseed oil, biodiesel oil and diesel oil. Ann. Agric. Environ. Med. 2008, 15, 45–49. [Google Scholar]
- April, T.M.; Abbott, S.P.; Foght, J.M.; Currah, R.S. Degradation of hydrocarbons in crude oil by the ascomycete Pseudal-lescheria boydii (Microascaceae). Can. J. Microbiol. 1998, 44, 270–278. [Google Scholar] [CrossRef] [PubMed]
- Smith, G.J.; Liew, E.C.; Hyde, K.D. The Xylariales: A Monophyletic Order Containing 7 Families. Fungal Divers. 2003. Available online: http://citeseerx.ist.psu.edu/viewdoc/summary?doi=10.1.1.515.7160 (accessed on 5 May 2021).
- Maharachchikumbura, S.S.N.; Guo, L.-D.; Cai, L.; Chukeatirote, E.; Wu, W.P.; Sun, X.; Crous, P.W.; Bhat, D.J.; McKenzie, E.H.C.; Bahkali, A.H.; et al. A multi-locus backbone tree for Pestalotiopsis, with a polyphasic characterization of 14 new species. Fungal Divers. 2012, 56, 95–129. [Google Scholar] [CrossRef] [Green Version]
- Maharachchikumbura, S.; Hyde, K.; Groenewald, J.; Xu, J.; Crous, P. Pestalotiopsis revisited. Stud. Mycol. 2014, 79, 121–186. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Samerpitak, K.; Van der Linde, E.; Choi, H.-J.; Ende, A.H.G.G.V.D.; Machouart, M.; Gueidan, C.; de Hoog, G.S. Taxonomy of Ochroconis, genus including opportunistic pathogens on humans and animals. Fungal Divers. 2014, 65, 89–126. [Google Scholar] [CrossRef]
- Viana-Bailez, A.M.; Bailez, O.; Malaquias, K.S. Comunicação Química em Formigas-Cortadeiras. In Formigas-Cortadeiras: Da Bioecologia ao Manejo, 1st ed.; Della-Luccia, T.M.C., Ed.; UFV: Abbotsford, BC, Canada, 2011. [Google Scholar]
- Randall, J.D.; Hemmingsen, B.B. A critical evaluation of the fume plate method for the enumeration of bacteria capable of growth on volatile hydrocarbons. In Applied Biotechnology for Site Bioremediation; Hinchee, R.E., Anderson, D.B., Sayles, G.D., Eds.; Lewis Publishers: Boca Raton, FL, USA, 1994; pp. 400–404. [Google Scholar]
- Blasi, B.; Poyntner, C.; Rudavsky, T.; Prenafeta-Boldú, F.X.; De Hoog, S.; Tafer, H.; Sterflinger, K. Pathogenic yet environmentally friendly? Black fungal candidates for bioremediation of pollutants. Geomicrobiol. J. 2016, 33, 308–317. [Google Scholar] [CrossRef] [PubMed]
- Dolatabadi, S.; Rezaei-Matehkolaei, A.; Pawlowska, J.; Hosseini, S.A.; Najafzadeh, M.J.; Madrid, H. Chaetothy-rialean fungi from aromatic hydrocarbon-polluted environments of Iran. Nova Hedwig. 2016, 405–426. [Google Scholar]
- Wainwright, M.; Ali, T.A.; Barakah, F. A review of the role of oligotrophic micro-organisms in biodeterioration. Int. Biodeterior. Biodegrad. 1993, 31, 1–13. [Google Scholar] [CrossRef]
- Parkinson, S.M.; Wainwright, M.; Killham, K. Observations on oligotrophic growth of fungi on silica gel. Mycol. Res. 1989, 93, 529–534. [Google Scholar] [CrossRef]
- Zhdanova, N.N.; Zakharchenko, V.A.; Vember, V.V.; Nakonechnaya, L.T. Fungi from Chernobyl: Mycobiota of the inner regions of the containment structures of the damaged nuclear reactor. Mycol. Res. 2000, 104, 1421–1426. [Google Scholar] [CrossRef]
- Dadachova, E.; Bryan, R.A.; Huang, X.; Moadel, T.; Schweitzer, A.D.; Aisen, P.; Nosanchuk, J.D.; Casadevall, A. Ionizing Radiation Changes the Electronic Properties of Melanin and Enhances the Growth of Melanized Fungi. PLoS ONE 2007, 2, e457. [Google Scholar] [CrossRef] [Green Version]
- Onofri, S.; Selbmann, L.; Zucconi, L.; Pagano, S. Antarctic microfungi as models for exobiology. Planet. Space Sci. 2004, 52, 229–237. [Google Scholar] [CrossRef]




{kind=link}
{kind=link}
{kind=link}
| Substrate of Isolation | Description | Locality (DMS * Coordinates) | Isolation Method |
|---|---|---|---|
| Contaminated soil | Soil from a garage shop | 22°26′13″ S; 47°34′4″ W | Oi flotation |
| Land farming soil from an oil refinery | 22°43′34″ S; 47°8′7″ W | Oil flotation | |
| Plant material | Bark fragments of Eucalyptus tereticornis | 22°24′34.37″ S; 47°32′28.98″ W | Oil flotation |
| Water samples | Water samples from a river related to the activity of an oil refinery | 22°44′23″ S; 47°07′40″ W | Standard serial dilution |
| Insects | Exoskeletons of gynes and drones of Attini ants | 22°50′6″ S; 48°26′1″ W | Oil flotation/Agar walk |
| Environmental Sample | Isolation Method a | No. of Strains | Observed Richness | ACE b | Chao1 | Shannon | Simpson |
|---|---|---|---|---|---|---|---|
| Eucalyptus tereticornis bark | OF | 9 | 3 | 7.07143 | 4 | 0.98643 | 0.37037 |
| Atta capiguara drone | OF, AW | 13 | 5 | 20.7037 | 11 | 1.50588 | 0.49704 |
| Atta laevigata drone | AW | 11 | 4 | 7.22469 | 4.5 | 1.49111 | 0.54545 |
| Garage soil | OF | 47 | 1 | 1 | 1 | 0 | 0 |
| Atta capiguara gyne | OF, AW | 7 | 2 | 3.11111 | 2 | 0.59167 | 0.2449 |
| Atta laevigata gyne | AW | 4 | 4 | error | 10 | 2 | 0.75 |
| Landfarming soil | OF | 7 | 4 | 5.78667 | 4.5 | 1.84237 | 0.69388 |
| Water sample | PP | 43 | 25 | 72.4482 | 47.6667 | 4.255 | 0.92699 |
| Data on the Strains | Biomass (mg) | Ratios | |||||||
|---|---|---|---|---|---|---|---|---|---|
| ID | Species | Substrate of Isolation | GC | NC | TH | TL | GC/NC | TH/NC | TL/NC |
| A55 | Westerdykella dispersa | Water samples | 81.9 | 1.4 | 14.5 | 17.9 | 58.5 | 10.4 | 12.8 |
| A234 | Pestalotiopsis sp. | Water samples | 38.8 | 0.3 | 4.4 | 4.1 | 129.3 | 14.7 | 13.7 |
| N34 | Cladosporium sp. | Atta capiguara drone | 36.9 | 0.4 | 8.1 | 4.2 | 92.3 | 20.3 | 10.5 |
| F21 | Melanized filamentous fungus | Atta capiguara drone | 82.6 | 1.5 | 14.8 | 10.3 | 55.1 | 9.9 | 6.9 |
| D4 | Cladophialophora devriesii | Eucalyptus tereticornis bark | 86.4 | 1.4 | 17.0 | 6.6 | 61.7 | 12.1 | 4.7 |
| D5 | Cladophialophora devriesii | Eucalyptus tereticornis bark | 81.7 | 1.6 | 4.4 | 18.1 | 51.1 | 2.8 | 11.3 |
| D15 | Chaetothyriales sp. | Eucalyptus tereticornis bark | 90.7 | 1.8 | 18.9 | 2.4 | 50.4 | 10.5 | 1.3 |
| D31 | Exophiala dermatitidis | Soil from machine shop | 76.8 | 1.8 | 17.5 | 4.3 | 42.7 | 9.7 | 2.4 |
| D153 | Exophiala dermatitidis | Soil from machine shop | 87.2 | 2.7 | 16.9 | 18.1 | 32.3 | 6.3 | 6.7 |
| D180 | Exophiala dermatitis | Soil from machine shop | 27.3 | 2.0 | 3.0 | 34.5 | 13.7 | 1.5 | 17.3 |
| A126 | Pleosporales sp. | Water samples | 5.9 | 1.2 | 6.1 | 6.6 | 4.9 | 5.1 | 5.5 |
| A304 | Melanized filamentous fungus | Water samples | 67.8 | 1.6 | 4.7 | 15.9 | 42.4 | 2.9 | 9.9 |
| A393 | Pleosporales sp. | Water samples | 22.5 | 1.8 | 17.7 | 6.8 | 12.5 | 9.8 | 3.8 |
| N21 | Cladosporium sp. | Atta capiguara drone | 25.7 | 3.0 | 19.0 | 18.3 | 8.6 | 6.3 | 6.1 |
| N85 | Cladosporium sp. | Atta laevigata drone | 21.4 | 2.4 | 16.5 | 17.3 | 8.9 | 6.9 | 7.2 |
| N92 | Cladosporium sp. | Atta capiguara gyne | 34.6 | 2.7 | 16.6 | 17.5 | 12.8 | 6.1 | 6.5 |
| N101 | Cladosporium sp. | Atta laevigata drone | 49.7 | 2.2 | 17.9 | 17.1 | 22.6 | 8.1 | 7.8 |
| F18 | Cladosporium sp. | Atta capiguara drone | 31.2 | 3.1 | 17.5 | 17.3 | 10.1 | 5.6 | 5.6 |
| NR7 | Pleosporales sp. | Water samples | 81.3 | 1.5 | 16.6 | 4.8 | 54.2 | 11.1 | 3.2 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Baron, N.C.; Pagnocca, F.C.; Otsuka, A.A.; Prenafeta-Boldú, F.X.; Vicente, V.A.; Attili de Angelis, D. Black Fungi and Hydrocarbons: An Environmental Survey for Alkylbenzene Assimilation. Microorganisms 2021, 9, 1008. https://doi.org/10.3390/microorganisms9051008
Baron NC, Pagnocca FC, Otsuka AA, Prenafeta-Boldú FX, Vicente VA, Attili de Angelis D. Black Fungi and Hydrocarbons: An Environmental Survey for Alkylbenzene Assimilation. Microorganisms. 2021; 9(5):1008. https://doi.org/10.3390/microorganisms9051008
Chicago/Turabian StyleBaron, Noemi Carla, Fernando Carlos Pagnocca, Ayumi Aquino Otsuka, Francesc Xavier Prenafeta-Boldú, Vânia Aparecida Vicente, and Derlene Attili de Angelis. 2021. "Black Fungi and Hydrocarbons: An Environmental Survey for Alkylbenzene Assimilation" Microorganisms 9, no. 5: 1008. https://doi.org/10.3390/microorganisms9051008