
Bacillus amyloliquefaciens TL Downregulates the Ileal Expression of Genes Involved in Immune Responses in Broiler Chickens to Improve Growth Performance





Abstract
:1. Introduction
2. Materials and Methods
2.1. Laboratory Animals
2.2. Tissue Sampling
2.3. RNA Preparation and Sequencing
2.4. Quality Control
2.5. Mapping Reads to the Reference Genome
2.6. Quantification of Gene Expression
2.7. Differential Expression Analysis
2.8. Gene Ontology (GO) and Pathway Enrichment
2.9. Quantitative Real-Time Polymerase Chain Reaction Analysis
2.10. Statistical Analyses and Data Records
3. Results
3.1. Growth Performance
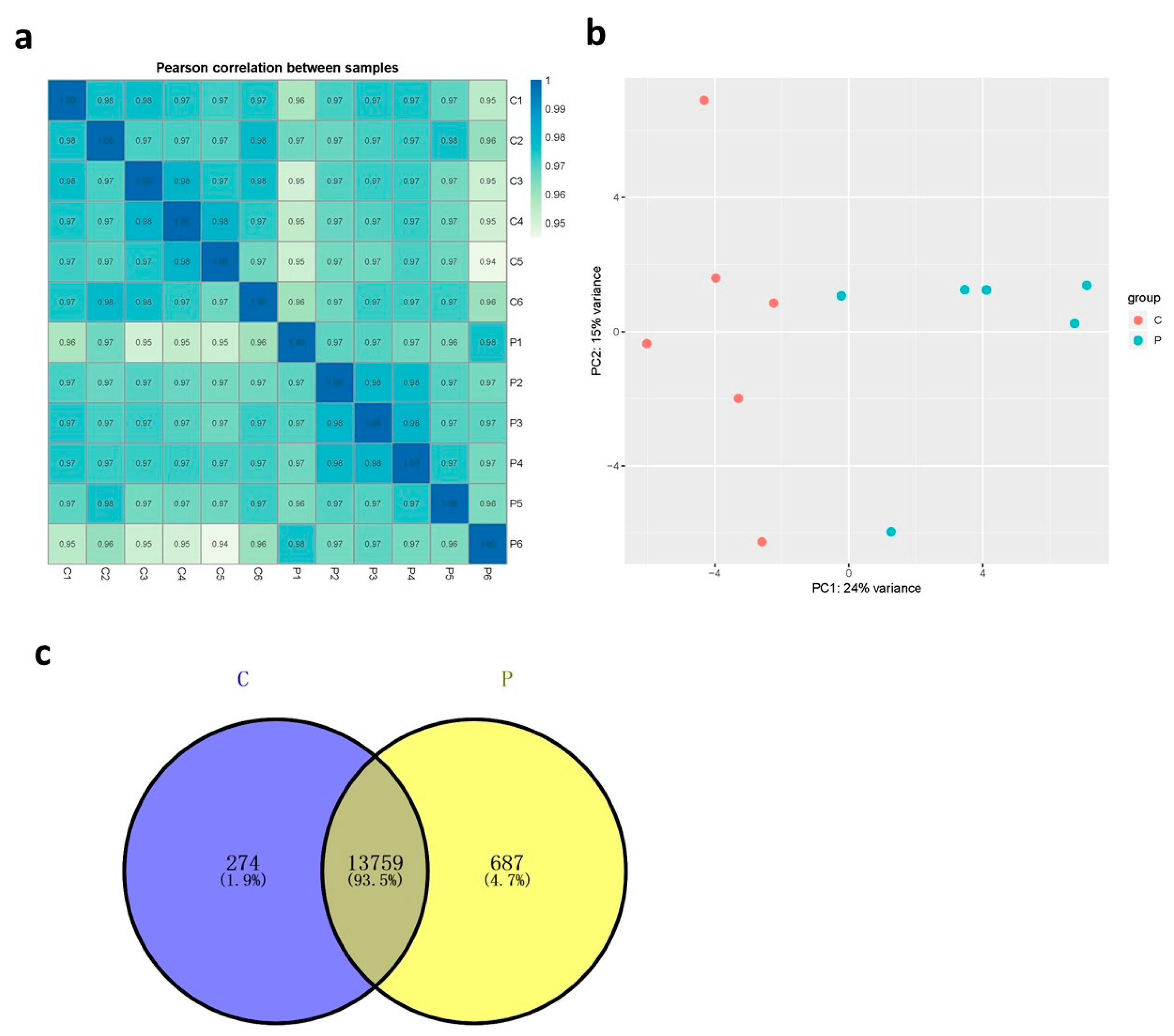
3.2. Transcriptome Analysis
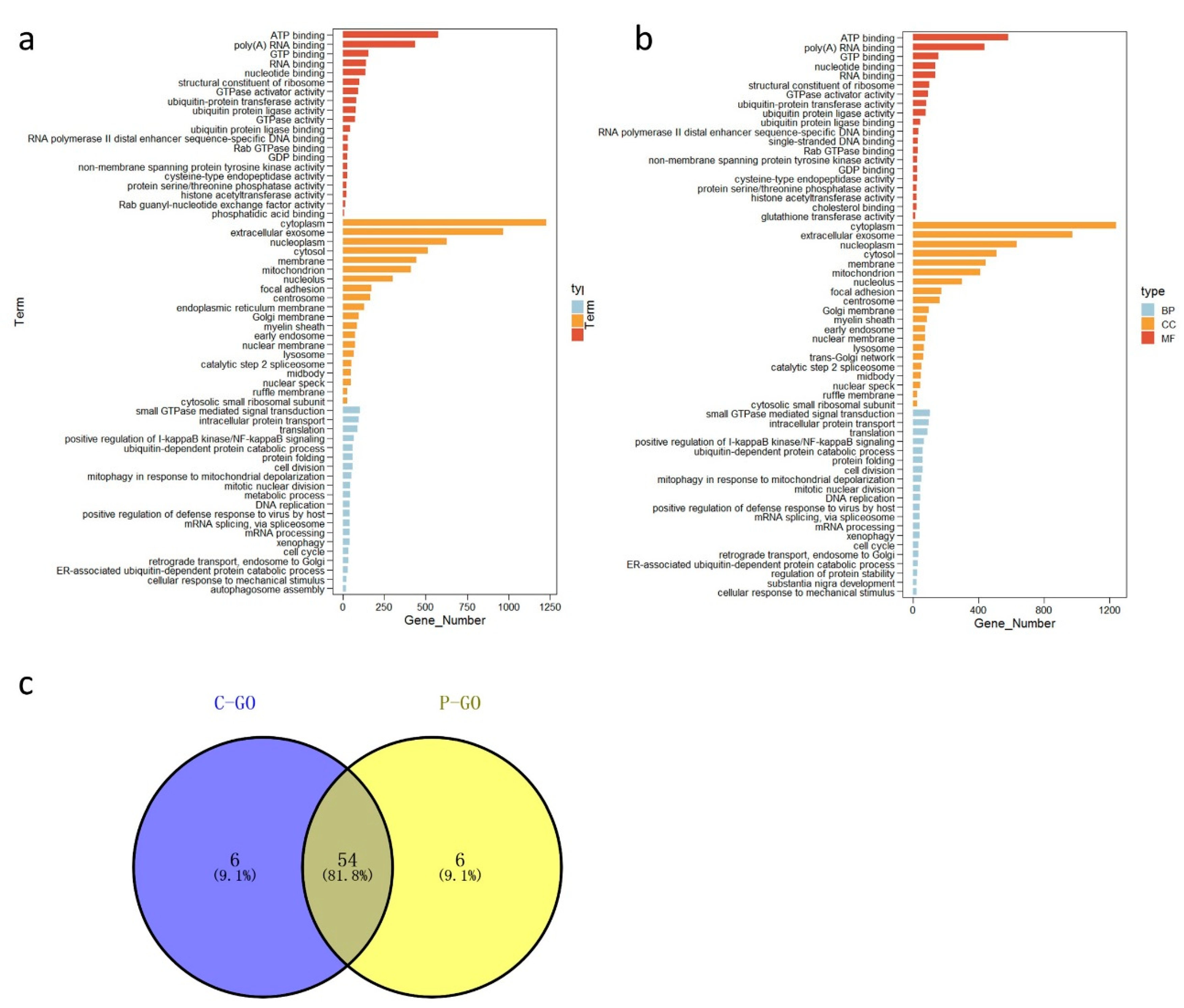
3.3. Functional Analysis of Broiler Chicken Transcriptomes
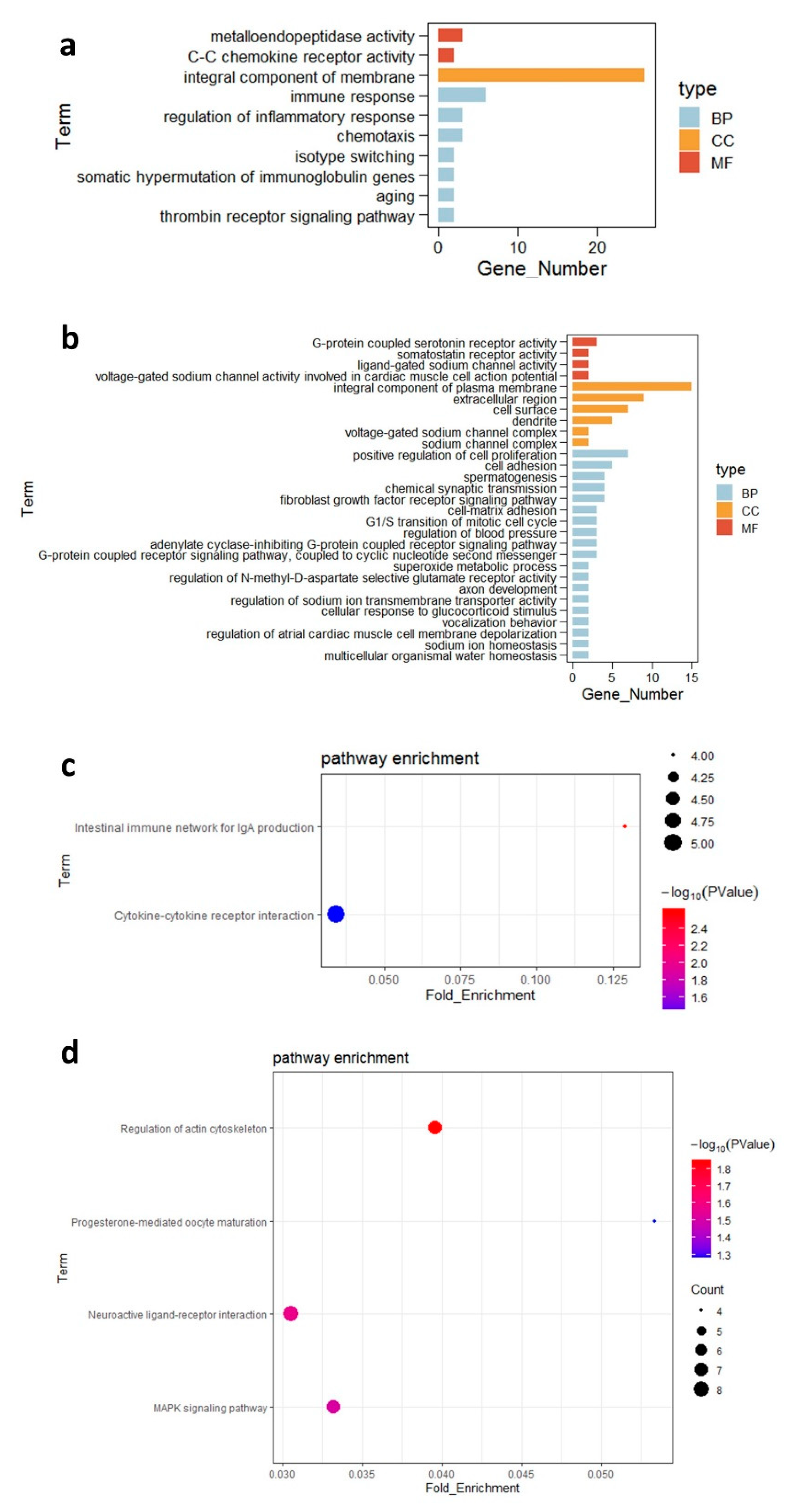
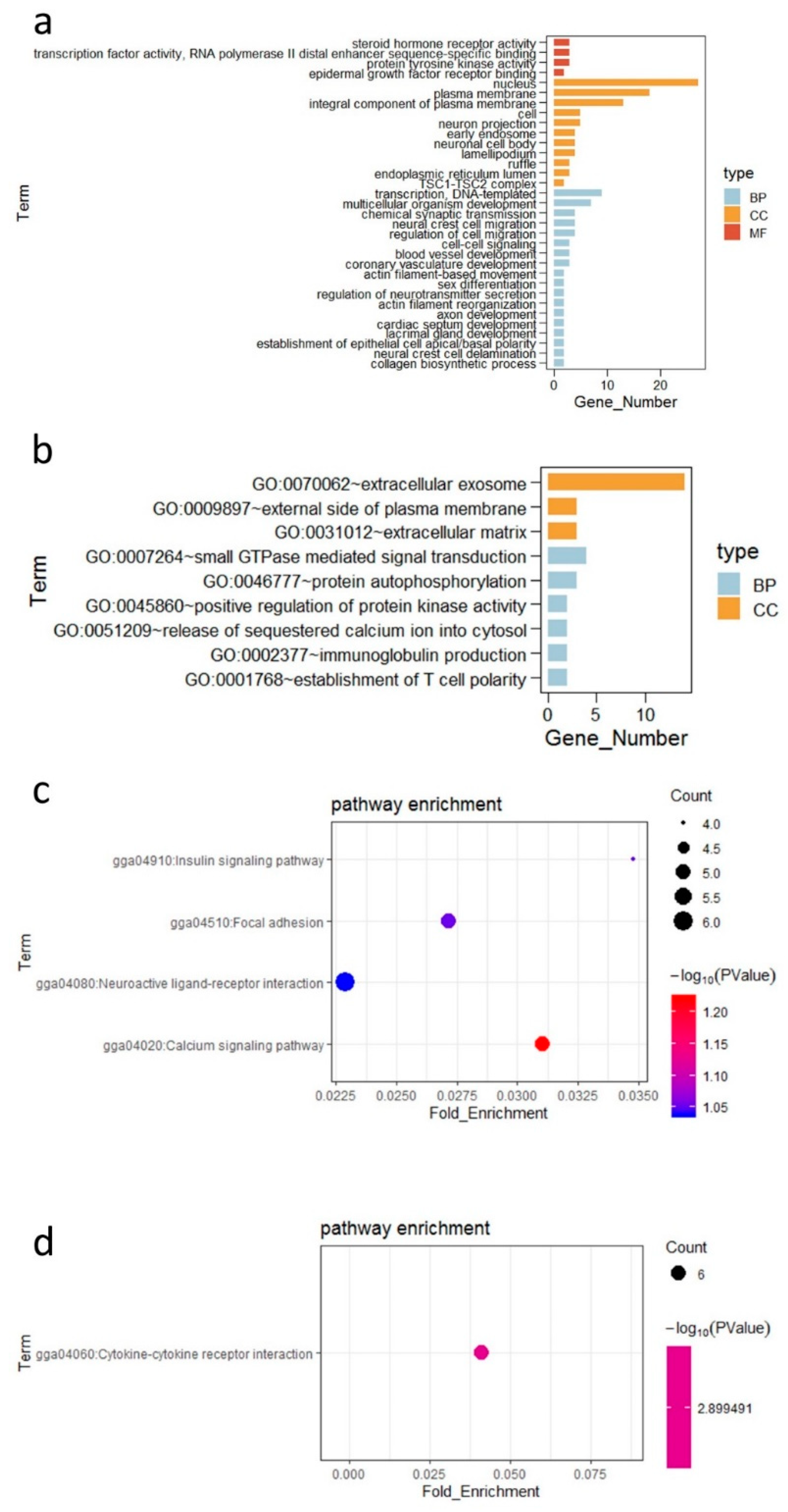
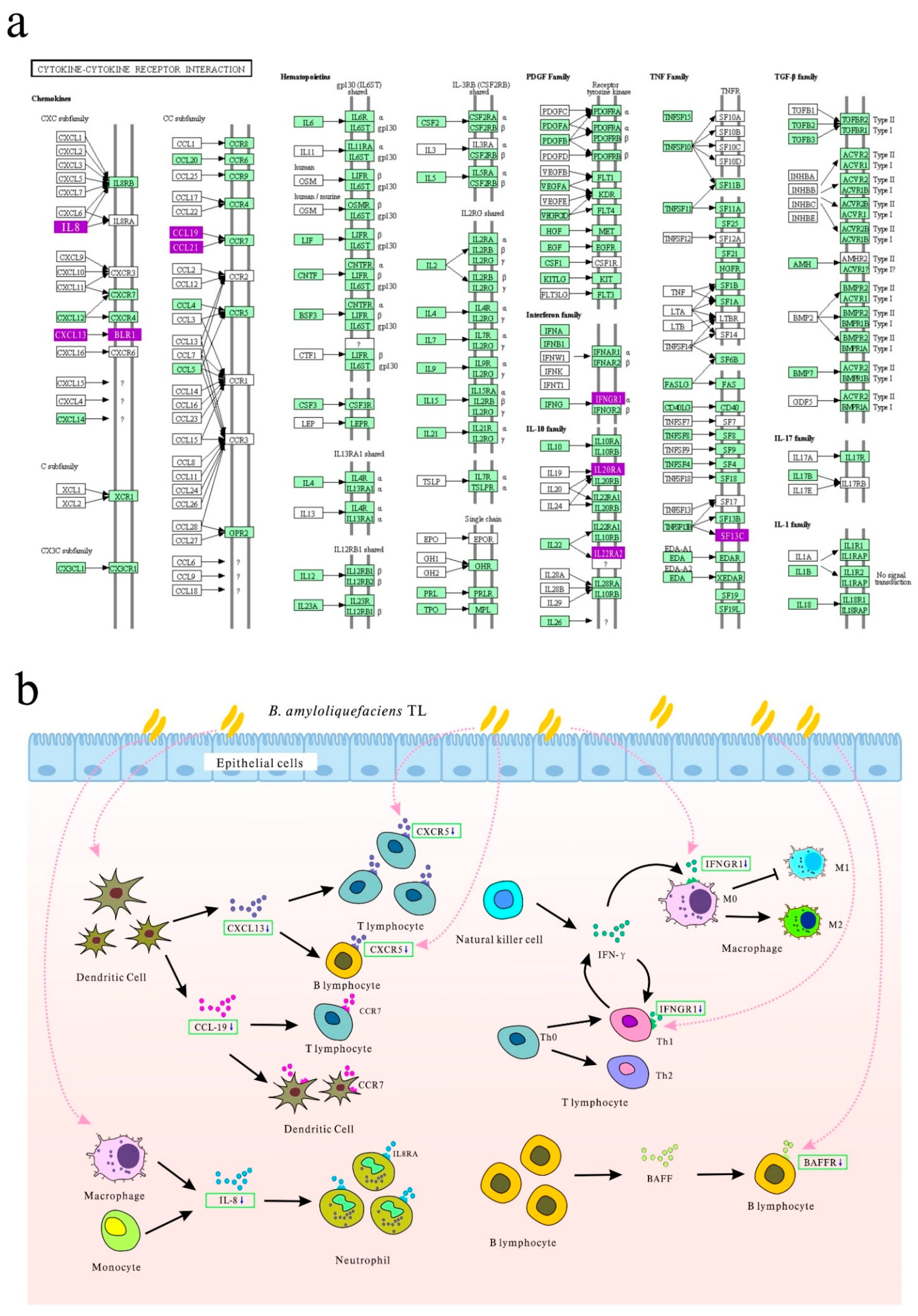
3.4. Differentially Expressed Genes in the Ileal Tissue and Their Functions
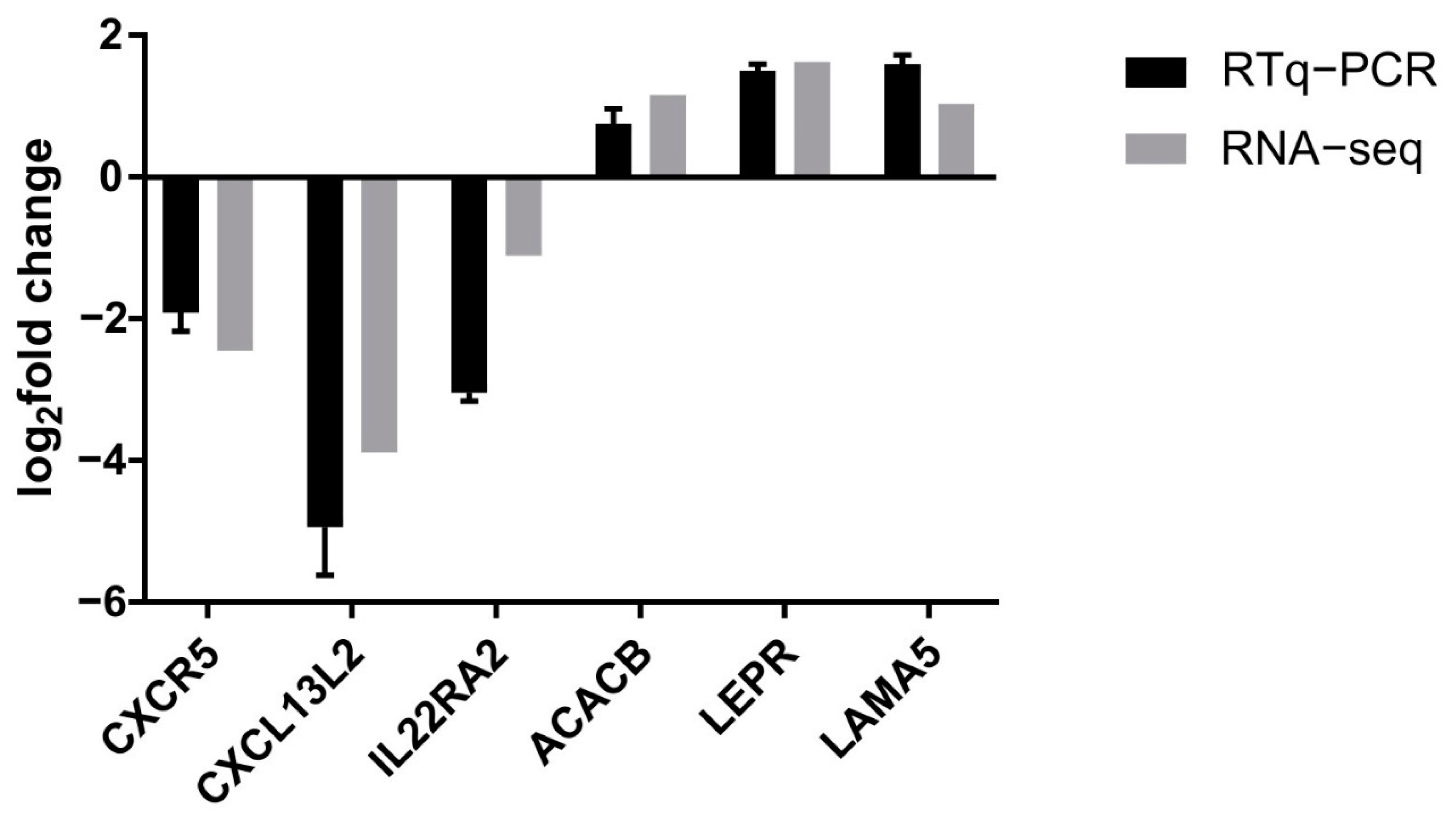
3.5. Confirmation of Gene Expression Data by Quantitative Reverse Transcription Polymerase Chain Reaction
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Jin, L.; Ho, Y.; Abdullah, N.; Jalaludin, S. Probiotics in poultry: Modes of action. World Poult. Sci. J. 1997, 53, 351–368. [Google Scholar] [CrossRef]
- Al-Khalaifa, H.; Al-Nasser, A.; Al-Surayee, T.; Al-Kandari, S.; Al-Enzi, N.; Al-Sharrah, T.; Ragheb, G.; Al-Qalaf, S.; Mohammed, A. Effect of dietary probiotics and prebiotics on the performance of broiler chickens. Poult. Sci. 2019, 98, 4465–4479. [Google Scholar] [CrossRef] [PubMed]
- Wu, Y.; Wang, B.; Zeng, Z.; Liu, R.; Tang, L.; Gong, L.; Li, W. Effects of probiotics Lactobacillus plantarum 16 and Paenibacillus polymyxa 10 on intestinal barrier function, antioxidative capacity, apoptosis, immune response, and biochemical parameters in broilers. Poult. Sci. 2019, 98, 5028–5039. [Google Scholar] [CrossRef] [PubMed]
- Majidi-Mosleh, A.; Sadeghi, A.; Mousavi, S.; Chamani, M.; Zarei, A. Ileal MUC2 gene expression and microbial population, but not growth performance and immune response, are influenced by in ovo injection of probiotics in broiler chickens. Br. Poult. Sci. 2017, 58, 40–45. [Google Scholar] [CrossRef] [PubMed]
- Cheng, Y.H.; Zhang, N.; Han, J.C.; Chang, C.W.; Hsiao, F.S.H.; Yu, Y.H. Optimization of surfactin production from Bacillus subtilis in fermentation and its effects on Clostridium perfringens-induced necrotic enteritis and growth performance in broilers. J. Anim. Physiol. Anim. Nutr. 2018, 102, 1232–1244. [Google Scholar] [CrossRef] [PubMed]
- Li, C.L.; Wang, J.; Zhang, H.J.; Wu, S.G.; Hui, Q.R.; Yang, C.B.; Fang, R.J.; Qi, G.H. Intestinal Morphologic and Microbiota Responses to Dietary Bacillus spp. in a Broiler Chicken Model. Front. Physiol. 2018, 9, 1968. [Google Scholar] [CrossRef]
- Grant, A.; Gay, C.G.; Lillehoj, H.S. Bacillus spp. as direct-fed microbial antibiotic alternatives to enhance growth, immunity, and gut health in poultry. Avian Pathol. 2018, 47, 339–351. [Google Scholar] [CrossRef] [PubMed]
- Whelan, R.A.; Doranalli, K.; Rinttilä, T.; Vienola, K.; Jurgens, G.; Apajalahti, J. The impact of Bacillus subtilis DSM 32315 on the pathology, performance, and intestinal microbiome of broiler chickens in a necrotic enteritis challenge. Poult. Sci. 2019, 98, 3450–3463. [Google Scholar] [CrossRef]
- Ramlucken, U.; Ramchuran, S.O.; Moonsamy, G.; Lalloo, R.; Thantsha, M.S.; Jansen van Rensburg, C. A novel Bacillus based multi-strain probiotic improves growth performance and intestinal properties of Clostridium perfringens challenged broilers. Poult. Sci. 2020, 99, 331–341. [Google Scholar] [CrossRef] [PubMed]
- Opalinski, M.; Maiorka, A.; Dahlke, F.; Cunha, F.; Vargas, F.; Cardozo, E. On the use of a probiotic (Bacillus subtilis-strain DSM 17299) as growth promoter in broiler diets. Braz. J. Poult. Sci. 2007, 9, 99–103. [Google Scholar] [CrossRef] [Green Version]
- Abreu, M.T. Toll-like receptor signalling in the intestinal epithelium: How bacterial recognition shapes intestinal function. Nat. Rev. Immunol. 2010, 10, 131. [Google Scholar] [CrossRef] [PubMed]
- Zampiga, M.; Bertocchi, M.; Bosi, P.; Trevisi, P.; Meluzzi, A.; Sirri, F. Differences in productive performance and intestinal transcriptomic profile in two modern fast-growing chicken hybrids. J. Anim. Physiol. Anim. Nutr. 2019, 103, 125–134. [Google Scholar] [CrossRef] [Green Version]
- Scanes, C.G.; Pierzchala-Koziec, K. Biology of the gastro-intestinal tract in poultry. Avian Biol. Res. 2014, 7, 193–222. [Google Scholar] [CrossRef]
- Sugiharto, S. Role of nutraceuticals in gut health and growth performance of poultry. J. Saudi Soc. Agric. Sci. 2016, 15, 99–111. [Google Scholar] [CrossRef] [Green Version]
- Barr, T.; Sureshchandra, S.; Ruegger, P.; Zhang, J.; Ma, W.; Borneman, J.; Grant, K.; Messaoudi, I. Concurrent gut transcriptome and microbiota profiling following chronic ethanol consumption in nonhuman primates. Gut Microbes 2018, 9, 338–356. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- O’Hara, A.M.; Shanahan, F. The gut flora as a forgotten organ. EMBO Rep. 2006, 7, 688–693. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhang, C.; Liu, Y.; Chen, S.; Qiao, Y.; Zheng, Y.; Xu, M.; Wang, Z.; Hou, J.; Wang, J.; Fan, H. Effects of Intranasal Pseudorabies Virus AH02LA Infection on Microbial Community and Immune Status in the Ileum and Colon of Piglets. Viruses 2019, 11, 518. [Google Scholar] [CrossRef] [Green Version]
- Lin, L.; Zhang, J. Role of intestinal microbiota and metabolites on gut homeostasis and human diseases. BMC Immunol. 2017, 18, 2. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sommer, F.; Bäckhed, F. The gut microbiota—Masters of host development and physiology. Nat. Rev. Microbiol. 2013, 11, 227. [Google Scholar] [CrossRef]
- Mohammed, A.A.; Jiang, S.; Jacobs, J.A.; Cheng, H.W. Effect of a synbiotic supplement on cecal microbial ecology, antioxidant status, and immune response of broiler chickens reared under heat stress. Poult. Sci. 2019, 98, 4408–4415. [Google Scholar] [CrossRef]
- Hu, Y.; Wang, L.; Shao, D.; Wang, Q.; Wu, Y.; Han, Y.; Shi, S. Selectived and Reshaped Early Dominant Microbial Community in the Cecum with Similar Proportions and Better Homogenization and Species Diversity due to Organic Acids as AGP Alternatives Mediate Their Effects on Broilers Growth. Front. Microbiol. 2019, 10, 2948. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yang, X.; Guo, Y.; He, X.; Yuan, J.; Yang, Y.; Wang, Z. Growth performance and immune responses in chickens after challenge with lipopolysaccharide and modulation by dietary different oils. Animal 2008, 2, 216–223. [Google Scholar] [CrossRef] [Green Version]
- Lai, H.T.; Nieuwland, M.G.; Kemp, B.; Aarnink, A.J.; Parmentier, H.K. Effects of repeated intratracheally administered lipopolysaccharide on primary and secondary specific antibody responses and on body weight gain of broilers. Poult. Sci. 2011, 90, 337–351. [Google Scholar] [CrossRef]
- Yang, X.; Li, W.; Feng, Y.; Yao, J. Effects of immune stress on growth performance, immunity, and cecal microflora in chickens. Poult. Sci. 2011, 90, 2740–2746. [Google Scholar] [CrossRef]
- Klasing, K.; Austic, R. Changes in protein degradation in chickens due to an inflammatory challenge. Proc. Soc. Exp. Biol. Med. 1984, 176, 292–296. [Google Scholar] [CrossRef] [PubMed]
- Hong, Y.; Cheng, Y.; Li, Y.; Li, X.; Zhou, Z.; Shi, D.; Li, Z.; Xiao, Y. Preliminary Study on the Effect of Bacillus amyloliquefaciens TL on Cecal Bacterial Community Structure of Broiler Chickens. BioMed. Res. Int. 2019, 2019, 5431354. [Google Scholar] [CrossRef] [Green Version]
- Liu, W.; Qiu, X.; Song, C.; Sun, Y.; Meng, C.; Liao, Y.; Tan, L.; Ding, Z.; Liu, X.; Ding, C. Deep Sequencing-Based Transcriptome Profiling Reveals Avian Interferon-Stimulated Genes and Provides Comprehensive Insight into Newcastle Disease Virus-Induced Host Responses. Viruses 2018, 10, 162. [Google Scholar] [CrossRef] [Green Version]
- Li, X.; Jia, L.; Chen, X.; Dong, Y.; Ren, X.; Dong, Y.; Chen, Y.; Xie, L.; Liu, M.; Shiota, C.; et al. Islet α-cell Inflammation Induced by NF-κB inducing kinase (NIK) Leads to Hypoglycemia, Pancreatitis, Growth Retardation, and Postnatal Death in Mice. Theranostics 2018, 8, 5960–5971. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Y.; Miao, G.; Wu, Q.; Lin, F.; You, C.; Wang, S.; Aweya, J.J.; Ma, H. Transcriptome sequencing and molecular markers discovery in the gonads of Portunus sanguinolentus. Sci. Data 2018, 5, 180131. [Google Scholar] [CrossRef] [PubMed]
- Trapnell, C.; Pachter, L.; Salzberg, S.L. TopHat: Discovering splice junctions with RNA-Seq. Bioinformatics 2009, 25, 1105–1111. [Google Scholar] [CrossRef] [PubMed]
- Trapnell, C.; Williams, B.A.; Pertea, G.; Mortazavi, A.; Kwan, G.; Van Baren, M.J.; Salzberg, S.L.; Wold, B.J.; Pachter, L. Transcript assembly and quantification by RNA-Seq reveals unannotated transcripts and isoform switching during cell differentiation. Nat. Biotechnol. 2010, 28, 511. [Google Scholar] [CrossRef] [Green Version]
- Benjamini, Y.; Hochberg, Y. Controlling the false discovery rate: A practical and powerful approach to multiple testing. J. R. Stat. Soc. B 1995, 57, 289–300. [Google Scholar] [CrossRef]
- Wang, Y.; Gao, M.; Zhu, F.; Li, X.; Yang, Y.; Yan, Q.; Jia, L.; Xie, L.; Chen, Z. METTL3 is essential for postnatal development of brown adipose tissue and energy expenditure in mice. Nat. Commun. 2020, 11, 1648. [Google Scholar] [CrossRef] [Green Version]
- Bai, K.; Huang, Q.; Zhang, J.; He, J.; Wang, T. Supplemental effects of probiotic Bacillus subtilis fmbJ on growth performance, antioxidant capacity, and meat quality of broiler chickens. Poult. Sci. 2016, 96, pew246. [Google Scholar] [CrossRef]
- Willis, W.L.; Isikhuemhen, O.S.; Ibrahim, S.A. Performance assessment of broiler chickens given mushroom extract alone or in combination with probiotics. Poult. Sci. 2007, 86, 1856–1860. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Z.F.; Kim, I.H. Effects of multistrain probiotics on growth performance, apparent ileal nutrient digestibility, blood characteristics, cecal microbial shedding, and excreta odor contents in broilers. Poult. Sci. 2014, 93, 364–370. [Google Scholar] [CrossRef] [PubMed]
- Al Azzaz, J.; Rieu, A.; Aires, V.; Delmas, D.; Chluba, J.; Winckler, P.; Bringer, M.A.; Lamarche, J.; Vervandier-Fasseur, D.; Dalle, F.; et al. Resveratrol-Induced Xenophagy Promotes Intracellular Bacteria Clearance in Intestinal Epithelial Cells and Macrophages. Front. Immunol. 2018, 9, 3149. [Google Scholar] [CrossRef]
- Lee, I.K.; Bae, S.; Gu, M.J.; You, S.J.; Kim, G.; Park, S.-M.; Jeung, W.-H.; Ko, K.H.; Cho, K.J.; Kang, J.S. H9N2-specific IgG and CD4+ CD25+ T cells in broilers fed a diet supplemented with organic acids. Poult. Sci. 2017, 96, 1063–1070. [Google Scholar] [CrossRef] [PubMed]
- Koskela, K.; Arstila, T.P.; Lassila, O. Costimulatory function of CD28 in avian gammadelta T cells is evolutionarily conserved. Scand. J. Immunol. 1998, 48, 635–641. [Google Scholar] [CrossRef] [PubMed]
- Munoz, I.; Berges, M.; Bonsergent, C.; Cormier-Aline, F.; Quéré, P.; Sibille, P. Cloning, expression and functional characterization of chicken CCR6 and its ligand CCL20. Mol. Immunol. 2009, 47, 551–559. [Google Scholar] [CrossRef] [PubMed]
- Wu, Z.; Hu, T.; Kaiser, P. Chicken CCR6 and CCR7 are markers for immature and mature dendritic cells respectively. Dev. Comp. Immunol. 2011, 35, 563–567. [Google Scholar] [CrossRef] [PubMed]
- Rosnet, O.; Blanco-Betancourt, C.; Grivel, K.; Richter, K.; Schiff, C. Binding of free immunoglobulin light chains to VpreB3 inhibits their maturation and secretion in chicken B cells. J. Biol. Chem. 2004, 279, 10228–10236. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Annamalai, T.; Selvaraj, R.K. Chemokine receptor CCR7 and CXCR5 mRNA in chickens following inflammation or vaccination. Poult. Sci. 2011, 90, 1695–1700. [Google Scholar] [CrossRef] [Green Version]
- Wang, G.; Zhang, Y.; Song, X.; Xia, Y.; Lai, P.F.; Ai, L. Lactobacillus casei LC2W can inhibit the colonization of Escherichia coli O157:H7 in vivo and reduce the severity of colitis. Food Funct. 2019, 10, 5843–5852. [Google Scholar] [CrossRef]
- Redweik, G.A.J.; Stromberg, Z.R.; Van Goor, A.; Mellata, M. Protection against avian pathogenic Escherichia coli and Salmonella Kentucky exhibited in chickens given both probiotics and live Salmonella vaccine. Poult. Sci. 2020, 99, 752–762. [Google Scholar] [CrossRef] [PubMed]
- Itoh, N.; Ornitz, D.M. Fibroblast growth factors: From molecular evolution to roles in development, metabolism and disease. J. Biochem. 2011, 149, 121–130. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Matsubara, Y.; Aoki, M.; Endo, T.; Sato, K. Characterization of the expression profiles of adipogenesis-related factors, ZNF423, KLFs and FGF10, during preadipocyte differentiation and abdominal adipose tissue development in chickens. Comp. Biochem. Physiol. B Biochem. Mol. Biol. 2013, 165, 189–195. [Google Scholar] [CrossRef] [PubMed]
- Kliewer, S.A.; Mangelsdorf, D.J. Bile Acids as Hormones: The FXR-FGF15/19 Pathway. Dig. Dis. 2015, 33, 327–331. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rulifson, I.C.; Collins, P.; Miao, L.; Nojima, D.; Lee, K.J.; Hardy, M.; Gupte, J.; Hensley, K.; Samayoa, K.; Cam, C.; et al. In Vitro and In Vivo Analyses Reveal Profound Effects of Fibroblast Growth Factor 16 as a Metabolic Regulator. J. Biol. Chem. 2017, 292, 1951–1969. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Okamoto, M.; Bito, T.; Noji, S.; Ohuchi, H. Subtype-specific expression of Fgf19 during horizontal cell development of the chicken retina. Gene Expr. Patterns 2009, 9, 306–313. [Google Scholar] [CrossRef]
- Olaya-Sánchez, D.; Sánchez-Guardado, L.; Ohta, S.; Chapman, S.C.; Schoenwolf, G.C.; Puelles, L.; Hidalgo-Sánchez, M. Fgf3 and Fgf16 expression patterns define spatial and temporal domains in the developing chick inner ear. Brain Struct. Funct. 2017, 222, 131–149. [Google Scholar] [CrossRef] [Green Version]
- Huang, Y.; Tsai, H.; Wang, S.; Kuo, S.; Fong, Y.; Tang, C. Amphiregulin promotes vascular endothelial growth factor-C expression and lymphangiogenesis through STAT3 activation in human chondrosarcoma cells. Cell Physiol. Biochem. 2019, 52, 1–15. [Google Scholar]
- Busser, B.; Sancey, L.; Brambilla, E.; Coll, J.L.; Hurbin, A. The multiple roles of amphiregulin in human cancer. Biochim. Biophys. Acta 2011, 1816, 119–131. [Google Scholar] [CrossRef] [PubMed]
- Yin, J.; Kim, T.-H.; Park, N.; Shin, D.; Choi, H.I.; Cho, S.; Park, J.B.; Kim, J.H. TRIM71 suppresses tumorigenesis via modulation of Lin28B-let-7-HMGA2 signaling. Oncotarget 2016, 7, 79854. [Google Scholar] [CrossRef]
- Qi, W.; Keenan, H.A.; Li, Q.; Ishikado, A.; Kannt, A.; Sadowski, T.; Yorek, M.A.; Wu, I.-H.; Lockhart, S.; Coppey, L.J. Pyruvate kinase M2 activation may protect against the progression of diabetic glomerular pathology and mitochondrial dysfunction. Nat. Med. 2017, 23, 753. [Google Scholar] [CrossRef] [PubMed]
- Mosoian, A. Intracellular and extracellular cytokine-like functions of prothymosin α: Implications for the development of immunotherapies. Future Med. Chem. 2011, 3, 1199–1208. [Google Scholar] [CrossRef] [PubMed]
- Calderwood, S.K.; Khaleque, M.A.; Sawyer, D.B.; Ciocca, D.R. Heat shock proteins in cancer: Chaperones of tumorigenesis. Trends Biochem. Sci. 2006, 31, 164–172. [Google Scholar] [CrossRef]
- Hassan, F.-U.; Nawaz, A.; Rehman, M.S.; Ali, M.A.; Dilshad, S.M.; Yang, C. Prospects of HSP70 as a genetic marker for thermo-tolerance and immuno-modulation in animals under climate change scenario. Anim. Nutr. 2019, 5, 340–350. [Google Scholar] [CrossRef] [PubMed]
- Xu, J.; Tang, S.; Song, E.; Yin, B.; Bao, E. Inhibition of heat shock protein 70 intensifies heat-stressed damage and apoptosis of chicken primary myocardial cells in vitro. Mol. Med. Rep. 2017, 15, 2881–2889. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Parcells, M.S.; Lin, S.-F.; Dienglewicz, R.L.; Majerciak, V.; Robinson, D.R.; Chen, H.-C.; Wu, Z.; Dubyak, G.R.; Brunovskis, P.; Hunt, H.D. Marek’s disease virus (MDV) encodes an interleukin-8 homolog (vIL-8): Characterization of the vIL-8 protein and a vIL-8 deletion mutant MDV. J. Virol. 2001, 75, 5159–5173. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wu, Y.F.; Shien, J.H.; Yin, H.H.; Chiow, S.H.; Lee, L.H. Structural and functional homology among chicken, duck, goose, turkey and pigeon interleukin-8 proteins. Vet. Immunol. Immunopathol. 2008, 125, 205–215. [Google Scholar] [CrossRef] [PubMed]
- Withanage, G.; Kaiser, P.; Wigley, P.; Powers, C.; Mastroeni, P.; Brooks, H.; Barrow, P.; Smith, A.; Maskell, D.; McConnell, I. Rapid expression of chemokines and proinflammatory cytokines in newly hatched chickens infected with Salmonella enterica serovar typhimurium. Infect. Immun. 2004, 72, 2152–2159. [Google Scholar] [CrossRef] [Green Version]
- Ateya, A.I.; Arafat, N.; Saleh, R.M.; Ghanem, H.M.; Naguib, D.; Radwan, H.A.; Elseady, Y. Intestinal gene expressions in broiler chickens infected with Escherichia coli and dietary supplemented with probiotic, acidifier and synbiotic. Vet. Res. Commun. 2019, 43, 131–142. [Google Scholar] [CrossRef] [PubMed]
- Ohandjo, A.Q.; Liu, Z.; Dammer, E.B.; Dill, C.D.; Griffen, T.L.; Carey, K.M.; Hinton, D.E.; Meller, R.; Lillard, J.W. Transcriptome network Analysis Identifies CXCL13-CXCR5 Signaling Modules in the prostate tumor immune Microenvironment. Sci. Rep. 2019, 9, 1–13. [Google Scholar] [CrossRef] [PubMed]
- Kazanietz, M.G.; Durando, M.; Cooke, M. CXCL13 and Its Receptor CXCR5 in Cancer: Inflammation, Immune Response, and Beyond. Front. Endocrinol. 2019, 10, 471. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Haertle, S.; Alzuheir, I.; Busalt, F.; Waters, V.; Kaiser, P.; Kaufer, B.B. Identification of the receptor and cellular ortholog of the Marek’s Disease Virus (MDV) CXC Chemokine. Front. Microbiol. 2017, 8, 2543. [Google Scholar] [CrossRef] [Green Version]
- Schneider, K.; Kothlow, S.; Schneider, P.; Tardivel, A.; Göbel, T.; Kaspers, B.; Staeheli, P. Chicken BAFF—A highly conserved cytokine that mediates B cell survival. Int. Immunol. 2004, 16, 139–148. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kothlow, S.; Morgenroth, I.; Graef, Y.; Schneider, K.; Riehl, I.; Staeheli, P.; Schneider, P.; Kaspers, B. Unique and conserved functions of B cell-activating factor of the TNF family (BAFF) in the chicken. Int. Immunol. 2007, 19, 203–215. [Google Scholar] [CrossRef] [Green Version]
- Han, X.; Chen, T.; Wang, M. Molecular cloning and characterization of chicken interferon-gamma receptor alpha-chain. J. Interferon Cytokine Res. 2008, 28, 445–454. [Google Scholar] [CrossRef] [PubMed]
- Kaiser, P.; Stäheli, P. Avian cytokines and chemokines. In Avian Immunology; Elsevier: Amsterdam, The Netherlands, 2014; pp. 189–204. [Google Scholar]
- Kidane, F.A.; Mitra, T.; Wernsdorf, P.; Hess, M.; Liebhart, D. Allocation of Interferon Gamma mRNA Positive Cells in Caecum Hallmarks a Protective Trait Against Histomonosis. Front. Immunol. 2018, 9, 1164. [Google Scholar] [CrossRef] [Green Version]
- Bandurska, K.; Krol, I.; Myga-Nowak, M. Interferons: Between structure and function. Postepy Hig. Med. Dosw. (Online) 2014, 68, 428–440. [Google Scholar] [CrossRef] [PubMed]
- Brisbin, J.T.; Gong, J.; Orouji, S.; Esufali, J.; Mallick, A.I.; Parvizi, P.; Shewen, P.E.; Sharif, S. Oral treatment of chickens with lactobacilli influences elicitation of immune responses. Clin. Vaccine Immunol. 2011, 18, 1447–1455. [Google Scholar] [CrossRef]
- Förster, R.; Davalos-Misslitz, A.C.; Rot, A. CCR7 and its ligands: Balancing immunity and tolerance. Nat. Rev. Immunol. 2008, 8, 362–371. [Google Scholar] [CrossRef] [PubMed]
- Ou, C.; Wang, Q.; Zhang, Y.; Kong, W.; Zhang, S.; Yu, Y.; Ma, J.; Liu, X.; Kong, X. Transcription profiles of the responses of chicken bursae of Fabricius to IBDV in different timing phases. Virol. J. 2017, 14, 93. [Google Scholar] [CrossRef] [PubMed]
- Wang, Q.; Ou, C.; Wei, X.; Yu, Y.; Jiang, J.; Zhang, Y.; Ma, J.; Liu, X.; Zhang, G. CC chemokine ligand 19 might act as the main bursal T cell chemoattractant factor during IBDV infection. Poult. Sci. 2019, 98, 688–694. [Google Scholar] [CrossRef] [PubMed]
- Haghighi, H.R.; Abdul-Careem, M.F.; Dara, R.A.; Chambers, J.R.; Sharif, S. Cytokine gene expression in chicken cecal tonsils following treatment with probiotics and Salmonella infection. Vet. Microbiol. 2008, 126, 225–233. [Google Scholar] [CrossRef] [PubMed]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Name. | Sequence (5′–3′) |
|---|---|
| β-actin | F: TATTGCTGCGCTCGTTGTTG |
| R: TGGCCCATACCAACCATCAC | |
| CXCR5 | F: CCTATGACTTGAGCCTGGTGG |
| R: TCCCAGCACGAACATAAGCAG | |
| LAMA5 | F: GCACATCCCATAACGAACGC |
| R: TCAGGACGAGGGGAATTTGC | |
| ACACB | F: TTCGGGACTTCAACCGTGAG |
| R: GGCTGCTTAAAATCCCGCAG | |
| CXCL13 | F: CAGCCATCCTGGAAGCCAAC |
| R: GGATCCACACAGATCCTCTCG | |
| LEPR | F: AGTGCAAACATGCAAGCGAG |
| R: CAGCTTGCCTTCAACCCAAC |
| Group | Control | Probiotics | p-Value | |
|---|---|---|---|---|
| 1 days | 40.84 ± 1.48 | 39.58 ± 1.19 | 0.065 | |
| Average weight | 7 days | 147.48 ± 11.08 | 157.35 ± 8.58 | 0.053 |
| (g) | 14 days | 499.75 ± 20.67 | 532.50 ± 24.09 | 0.056 |
| 21 days | 858.62 ± 30.74 A | 1076.12 ± 98.48 B | 0.000 | |
| - | 35 days | 1901.38 ± 76.36 a | 2156.12 ± 198.61 b | 0.017 |
| 1–7 days | 15.23 ± 1.38 a | 16.82 ± 1.08 b | 0.022 | |
| Average daily | 8–14 days | 50.32 ± 1.51 | 53.59 ± 2.33 | 0.074 |
| gain (g) | 15–21 days | 51.27 ± 1.74 A | 77.66 ± 10.74 B | 0.000 |
| 22–35 days | 80.21 ± 3.72 | 83.08 ± 9.23 | 0.608 | |
| Feed conversion ratio | 2.02 | 1.62 | ||
| KEGG Pathway | p-Value | Gene Number | Molecules |
|---|---|---|---|
| Up group | |||
| Calcium signaling pathway | 0.059866 | 5 | PHKA1, ADCY3, P2RX1, HTR6, ITPR3 |
| Focal adhesion | 0.088308 | 5 | COL27A1, LAMA5, RAPGEF1, KDR, COL5A1 |
| Insulin signaling pathway | 0.089046 | 4 | TSC2, PHKA1, ACACB, RAPGEF1 |
| Neuroactive ligand–receptor interaction | 0.091555 | 6 | SSTR1, GRIK4, GRM2, CHRM4, P2RX1, HTR6 |
| Down group | |||
| Cytokine–cytokine receptor interaction | 0.001260 | 6 | TNFRSF13C, IFNGR1, CXCL13L2, IL20RA, CCL19, CXCR5 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Hong, Y.; Cheng, Y.; Guan, L.; Zhou, Z.; Li, X.; Shi, D.; Xiao, Y. Bacillus amyloliquefaciens TL Downregulates the Ileal Expression of Genes Involved in Immune Responses in Broiler Chickens to Improve Growth Performance. Microorganisms 2021, 9, 382. https://doi.org/10.3390/microorganisms9020382
Hong Y, Cheng Y, Guan L, Zhou Z, Li X, Shi D, Xiao Y. Bacillus amyloliquefaciens TL Downregulates the Ileal Expression of Genes Involved in Immune Responses in Broiler Chickens to Improve Growth Performance. Microorganisms. 2021; 9(2):382. https://doi.org/10.3390/microorganisms9020382
Chicago/Turabian StyleHong, Yuxuan, Yang Cheng, Leluo Guan, Zutao Zhou, Xiaowen Li, Deshi Shi, and Yuncai Xiao. 2021. "Bacillus amyloliquefaciens TL Downregulates the Ileal Expression of Genes Involved in Immune Responses in Broiler Chickens to Improve Growth Performance" Microorganisms 9, no. 2: 382. https://doi.org/10.3390/microorganisms9020382