
Kale Attenuates Inflammation and Modulates Gut Microbial Composition and Function in C57BL/6J Mice with Diet-Induced Obesity


Abstract
:1. Introduction
2. Materials and Methods
2.1. Study Animals and Study Diets
2.2. Randomization and Sample Size Determination
2.3. Determination of Body Weight and Insulin Sensitivity
2.4. Euthanasia, Determination of Fat Accumulation and Tissue Collection
2.5. Determination of Fermentation Variables
2.6. RNA Extraction and cDNA Synthesis
2.7. Analysis of Target Genes in Adipose Tissue by qPCR
2.8. DNA Extraction
2.9. Amplification, Library Preparation, Template Preparation and Sequencing
2.10. Microbial Diversity Analysis and Bioinformatics
2.11. Analysis of Enrichment of Bacteria in Each Study Group
2.12. PICRUSt2
3. Results
3.1. Body Weight, Fat Accumulation, Insulin Sensitivity and Inflammation
3.2. Markers of Fermentation
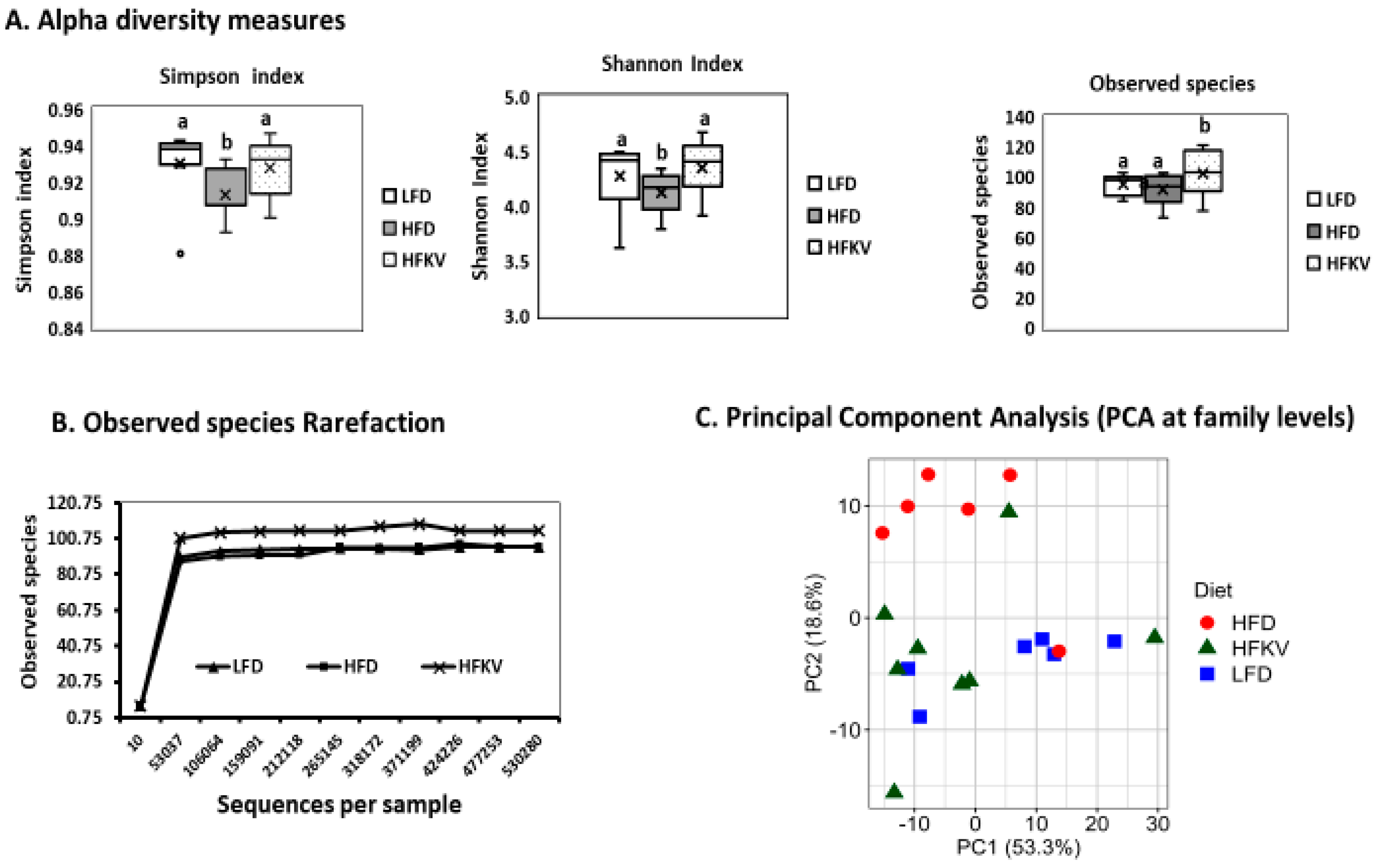
3.3. Diversity Metrics
3.4. Comparative Analysis of the Gut Microbial Composition
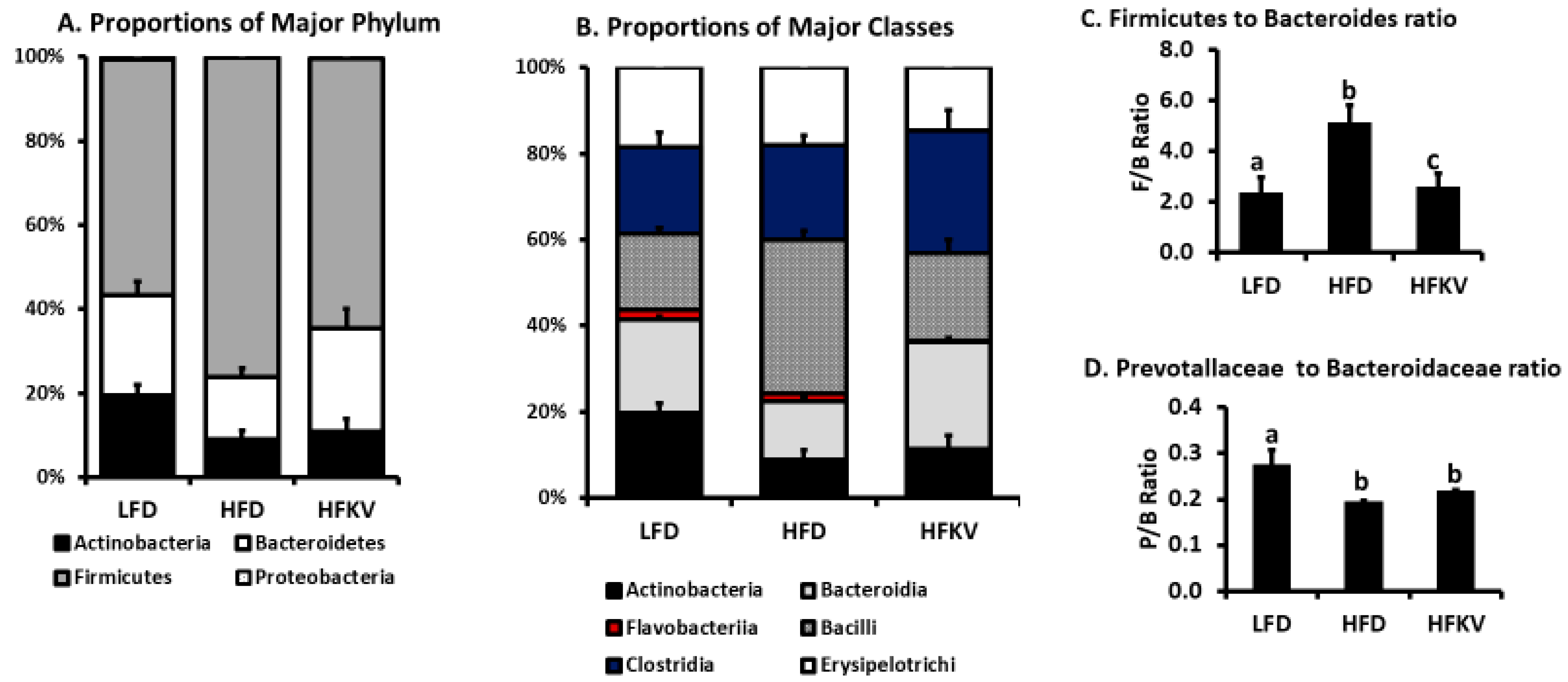
3.5. Comparative Analysis of Phylum Actinobacteria
3.6. Comparative Analysis of Phylum Bacteroidetes
3.7. Comparative Analysis of Phylum Firmicutes
3.8. Comparative Analysis of Phylum Proteobacteria and Verrucomicrobia
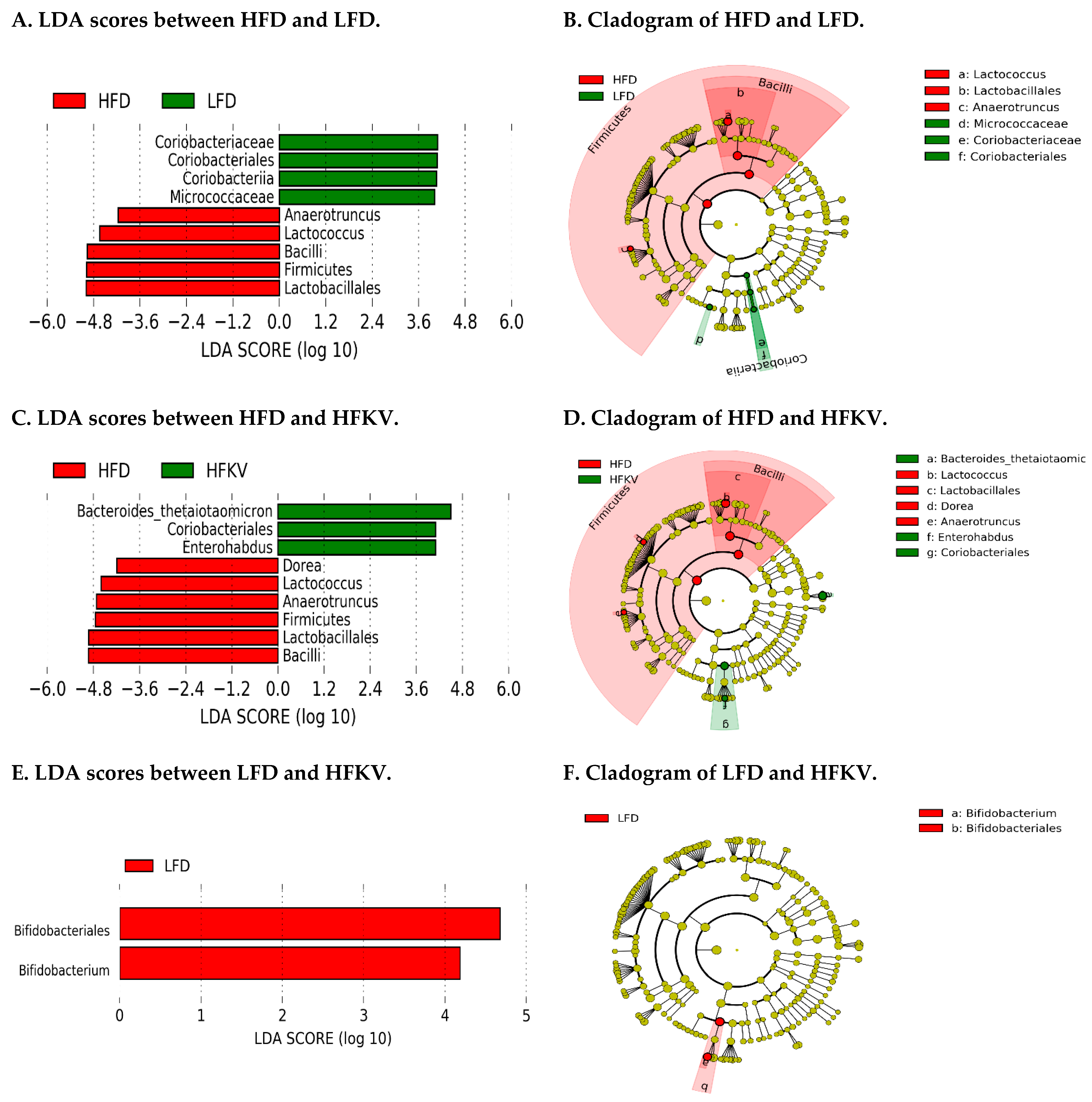
3.9. Linear Discriminant Analysis Effect Size (LEfSe)
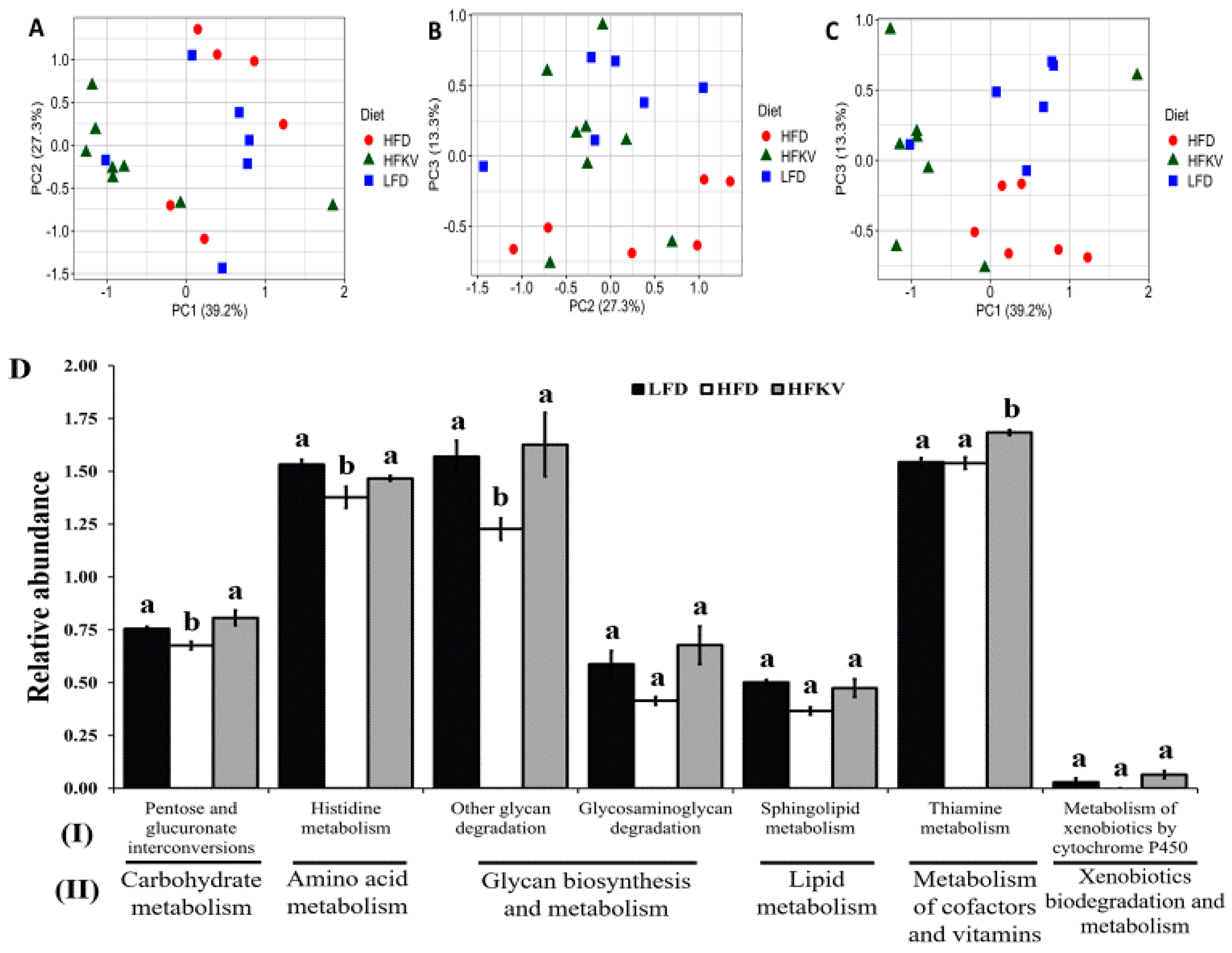
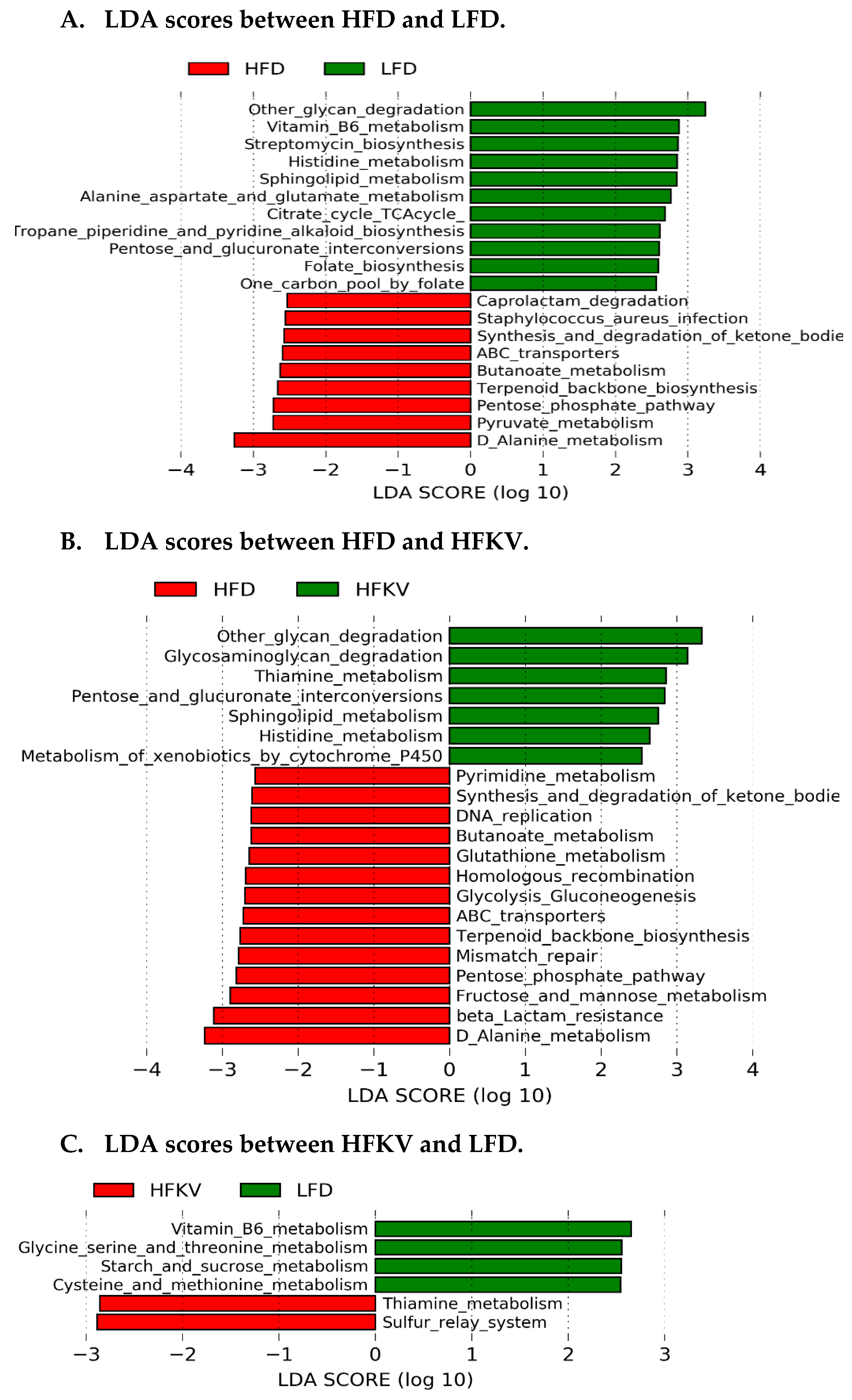
3.10. Predicted Metabolic Functions
4. Discussion
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Migliozzi, M.; Thavarajah, D.; Thavarajah, P.; Smith, P. Lentil and kale: Complementary nutrient-rich whole food sources to combat micronutrient and calorie malnutrition. Nutrients 2015, 7, 9285–9298. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Šamec, D.; Urlić, B.; Sondi, B.S. Kale (Brassica oleracea var. acephala) as a superfood: Review of the scientific evidence behind the statement. Crit. Rev. Food Sci. Nutr. 2018, 59, 2411–2422. [Google Scholar] [CrossRef]
- Olsen, H.; Aaby, K.; Borge, G.I. Characterization and quantification of flavonoids and hydroxycinnamic acids in curly kale (Brassica oleracea L. Convar. acephala var. sabellica) by HPLC-DAD-ESI-MSn. J. Agric. Food. Chem. 2009, 57, 2816–2825. [Google Scholar] [PubMed]
- Dinkova-Kostova, A.T.; Kostov, R.V. Glucosinolates and isothiocyanates in health and disease. Trends Mol. Med. 2012, 18, 337–347. [Google Scholar] [CrossRef] [PubMed]
- Ferreres, F.; Fernandes, F.; Sousa, C.; Valentao, P.; Pereira, J.A.; Andrade, P.B. Metabolic and bioactivity insights into Brassica oleracea var. acephala. J. Agric. Food. Chem. 2009, 57, 8884–8892. [Google Scholar] [CrossRef]
- Kuerban, A.; Yaghmoor, S.S.; Almulaiky, Y.Q.; Mohamed, Y.A.; Razvi, S.S.; Hasan, M.N.; Moselhy, S.S.; Al-Ghafari, A.; AlSufiani, H.M.; Kumosani, T.A.; et al. Therapeutic effects of phytochemicals of brassicaceae for management of obesity. J. Pharm. Res. Int. 2017, 19, 1–11. [Google Scholar] [CrossRef]
- Di Noia, J. Defining powerhouse fruits and vegetables: A nutrient density approach. Prev. Chronic. Dis. 2014, 11, 130390. [Google Scholar] [CrossRef] [Green Version]
- Liu, H.; Ivarsson, E.; Dicksved, J.; Lundh, T.; Lindberg, J.E. Inclusion of chicory (Cichorium intybus L.) in pigs’ diets affects the intestinal microenvironment and the gut microbiota. Appl. Environ. Microbiol. 2012, 78, 4102–4109. [Google Scholar] [CrossRef] [Green Version]
- Ramnani, P.; Gaudier, E.; Bingham, M.; Van Bruggen, P.; Tuohy, K.; Gibson, G. Prebiotic effect of fruit and vegetable shots containing Jerusalem artichoke inulin: A human intervention study. Br. J. Nutr. 2010, 104, 233–240. [Google Scholar] [CrossRef] [Green Version]
- Kaczmarek, J.L.; Liu, X.; Charron, C.S.; Novotny, J.A.; Jeffery, E.H.; Seifried, H.E.; Ross, S.A.; Miller, M.J.; Swanson, K.S.; Holscher, H.D. Broccoli consumption affects the human gastrointestinal microbiota. J. Nutr. Biochem. 2019, 63, 27–34. [Google Scholar] [CrossRef]
- Roza, N.A.V.; Possignolo, L.F.; Palanch, A.C.; Gontijo, J.A.R. Effect of long-term high-fat diet intake on peripheral insulin sensibility, blood pressure, and renal function in female rats. Food Nutr. Res. 2016, 60, 28536. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Xiao, J.; Metzler-Zebeli, B.U.; Zebeli, Q. Gut function-enhancing properties and metabolic effects of dietary indigestible sugars in rodents and rabbits. Nutrients 2015, 7, 8348–8365. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bolyen, E.; Rideout, J.R.; Dillon, M.R.; Bokulich, N.A.; Abnet, C.C.; Al-Ghalith, G.A.; Alexander, H.; Alm, E.J.; Arumugam, M.; Asnicar, F.; et al. Reproducible, interactive, scalable and extensible microbiome data science using QIIME. Nat. Biotechnol. 2019, 37, 852–857. [Google Scholar] [CrossRef] [PubMed]
- Segata, N.; Izard, J.; Waldron, L.; Gevers, L.; Miropolsky, W.S.; Garret, W.S.; Huttenhower, C. Meta-genomic biomarker discovery and explanation. Genome Biol. 2011, 12, R60. [Google Scholar] [CrossRef] [Green Version]
- Douglas, G.M.; Maffei, V.J.; Zaneveld, J.R.; Yurgel, S.N.; Brown, J.R.; Taylor, C.M.; Huttenhower, C.; Langille, M.G.I. PICRUSt2 for prediction of metagenome functions. Nat. Biotechnol. 2020, 38, 685–688. [Google Scholar] [CrossRef]
- Zhao, X.; Zhang, Z.; Hu, B.; Huang, W.; Yuan, C.; Zou, L. Response of gut microbiota to metabolite changes induced by endurance exercise. Front. Microbiol. 2018, 9, 765. [Google Scholar] [CrossRef]
- Liu, H.; Zhang, H.; Wang, X.; Yu, X.; Hu, C.; Zhang, X.-L. The family Coriobacteriaceae is a potential contributor to the beneficial effects of Roux-en-Y gastric bypass on type 2 diabetes. Surg. Obes. Relat. Dis. 2018, 14, 584–593. [Google Scholar] [CrossRef]
- Wexler, H.M.; Daya, S.; Berns, K.I. Bacteroides: The good, the bad, and the nitty-gritty. Clin. Microbiol. Rev. 2007, 20, 593–621. [Google Scholar] [CrossRef] [Green Version]
- Xu, J.; Gordon, J.I. Honor thy symbionts. Proc. Natl. Acad. Sci. USA 2003, 100, 10452–10459. [Google Scholar] [CrossRef] [Green Version]
- Turnbaugh, P.J.; Bäckhed, F.; Fulton, L.A.; Gordon, J.I. Diet-induced obesity is linked to marked but reversible alterations in the mouse distal gut microbiome. Cell Host Microbe 2008, 3, 213–223. [Google Scholar] [CrossRef] [Green Version]
- Petersen, C.; Bell, R.; Klag, K.A.; Lee, S.-H.; Soto, R.; Ghazaryan, A.; Buhrke, K.; Ekiz, H.A.; Ost, K.S.; Boudina, S.; et al. T cell–mediated regulation of the microbiota protects against obesity. Science 2019, 365, eaat9351. [Google Scholar] [CrossRef] [PubMed]
- Atarashi, K.; Tanoue, T.; Oshima, K.; Suda, W.; Nagano, Y.; Nishikawa, H.; Fukuda, S.; Saito, T.; Narushima, S.; Hase, K.; et al. Treg induction by a rationally selected mixture of Clostridia strains from the human microbiota. Nature 2013, 500, 232–236. [Google Scholar] [CrossRef] [PubMed]
- Shang, H.; Sun, J.; Chen, Y.Q. Clostridium butyricum CGMCC0313.1 modulates lipid profile, insulin resistance and colon homeostasis in obese mice. PLoS ONE 2016, 11, e0154373. [Google Scholar] [CrossRef] [PubMed]
- Clavel, T.; Lepage, P.; Charrier, C. The family Coriobacteriaceae. In The Prokaryotes; Rosenberg, E., DeLong, E.F., Lory, S., Stackebrandt, E., Thompson, F., Eds.; Springer: Berlin/Heidelberg, Germany, 2014; pp. 201–238. [Google Scholar]
- Videira, P.A.; Castro-Caldas, M. Linking glycation and glycosylation with inflammation and mitochondrial dysfunction in Parkinson’s disease. Front. Neurosci. 2018, 12, 381. [Google Scholar] [CrossRef]
- Verhelst, X.; Dias, A.M.; Colombel, J.F.; Vermeire, S.; Van Vlierberghe, H.; Callewaert, N.; Pinho, S.S. Protein glycosylation as a diagnostic and prognostic marker of chronic inflammatory gastrointestinal and liver diseases. Gastroenterology 2020, 158, 95–110. [Google Scholar] [CrossRef] [Green Version]
- Tijardović, M.; Marijančević, D.; Bok, D.; Kifer, D.; Lauc, G.; Gornik, O.; Keser, T. Intense physical exercise induces an anti-inflammatory change in IgG N-glycosylation profile. Front. Physiol. 2019, 10, 1522. [Google Scholar] [CrossRef]
- Goulabchand, R.; Vincent, T.; Batteux, F.; Eliaou, J.F.; Guilpain, P. Impact of autoantibody glycosylation in autoimmune diseases. Autoimmun. Rev. 2014, 13, 742–750. [Google Scholar] [CrossRef]
- Page, G.L.J.; Laight, D.; Cummings, M.H. Thiamine deficiency in diabetes mellitus and the impact of thiamine replacement on glucose metabolism and vascular disease. Int. J. Clin. Pract. 2011, 65, 684–690. [Google Scholar] [CrossRef] [Green Version]
- Babaei-Jadidi, R.; Karachalias, N.; Kupich, C.; Ahmed, N.; Thornalley, P.J. High-dose thiamine therapy counters dyslipidaemia in streptozotocin-induced diabetic rats. Diabetologia 2004, 47, 2235–2246. [Google Scholar] [CrossRef] [Green Version]
- Charron, C.S.; Novotny, J.A.; Jeffery, E.H.; Kramer, M.; Ross, S.A.; Seifried, H.E. Consumption of baby kale increased cytochrome P450 1A2 (CYP1A2) activity and influenced bilirubin metabolism in a randomized clinical trial. J. Funct. Foods 2020, 64, 103624. [Google Scholar] [CrossRef]
- Hubbard, T.D.; Murray, I.A.; Nichols, R.G.; Cassel, K.; Podolsky, M.; Kuzu, G.; Tian, Y.; Smith, P.; Kennett, M.J.; Patterson, A.D.; et al. Dietary broccoli impacts microbial community structure and attenuates chemically induced colitis in mice in an Ah receptor dependent manner. J. Funct. Foods 2017, 37, 685–698. [Google Scholar] [CrossRef] [PubMed]
- Denison, M.S.; Nagy, S.R. Activation of the aryl hydrocarbon receptor by structurally diverse exogenous and endogenous chemicals. Annu. Rev. Pharmacol. Toxicol. 2003, 43, 309–334. [Google Scholar] [CrossRef] [PubMed]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Component | Amount (%) | Analytical Methods Used |
|---|---|---|
| Proximate Analysis | ||
| Ash | 10.87 | AOAC: 923.03 |
| Carbohydrates | 73.9 | By calculation |
| Protein (6.25) | 8.71 | AACC 46–30; AOAC 992.15 |
| Total Fat | 1.88 | AOAC: 996.06 |
| Fiber | ||
| Insoluble Dietary Fiber | 37.8 | AOAC: 991.43 |
| Soluble Dietary Fiber | 6.2 | |
| Total Dietary Fiber | 45.1 | |
| Fat | AOAC: 996.06 | |
| Saturated Fat | 0.28 | |
| Monounsaturated Fat | 0.21 | |
| Cis-cis Polyunsaturated Fat | 0.85 | |
| Trans fat | 0.46 | |
| Moisture | 4.65 | AOAC: 945.43, 934.01 |
| Calories per 100 g | ||
| Calories | 347.26 | By calculation |
| Calories, 2020 | 184.74 | |
| Calories from Fat | 17 | |
| Calories from Saturated Fat | 3 | |
| Calories (insoluble fiber subtracted) | 196 |
| Ingredients (g) | LFD | HFD | HFKV |
|---|---|---|---|
| Casein | 200 | 200 | 191.88 |
| L-Cystine | 3 | 3 | 3 |
| Corn starch | 452.2 | 72.8 | 48.28 |
| Maltodextrin 10 | 75 | 100 | 100 |
| Sucrose | 172.8 | 172.8 | 172.8 |
| Cellulose | 50 | 50 | 13.92 |
| Soybean oil | 25 | 25 | 23.46 |
| Lard | 20 | 177.5 | 177.5 |
| Mineral mix | 10 | 10 | 10 |
| Dicalcium phosphate | 13 | 13 | 13 |
| Calcium carbonate | 5.5 | 5.5 | 5.5 |
| Potassium citrate | 16.5 | 16.5 | 16.5 |
| Vitamin mix | 10 | 10 | 10 |
| Choline bitartrate | 2 | 2 | 2 |
| Kale dried powdered leaves | 0 | 0 | 82 |
| Total (g) | 1055.05 | 858.15 | 869.89 |
| Kcal from Specific Nutrients | |||
| Protein | 716 | 716 | 716 |
| Carbohydrate | 2840 | 1422.4 | 1422.4 |
| Fat | 405 | 1822.5 | 1822.5 |
| Total Kcal | 3961 | 3960.9 | 3961 |
| (a) | |||||
| LFD | HFD | HFKV | p-Value | ||
| Obesity Parameters | |||||
| Calorie intake per day (Kcal/d) | 11.2 (0.5) a | 11.5 (0.2) a | 11.4 (0.3) a | ns | |
| Body weight (g) | 26.5 (1.8) a | 36.4 (2.2) b | 33.7 (3.8) b | <0.05 | |
| Total body fat (g) | 0.56 (0.12) a | 1.99 (0.23) b | 1.48 (0.2) b | <0.05 | |
| HOMA-IR | 0.11(0.03) a | 0.88 (0.18) b | 0.78 (0.15) b | <0.05 | |
| Serum Inflammation markers | |||||
| MCP-1 (pg/mL) | 776.1 (140.2) a | 1355.3 (140.2) b | 435.7 (140.2) c | 0.001 | |
| IL-10 (pg/mL) | 195.3 (23.9) a | 284.7 (28.9) b | 332.9 (52.1) c | <0.05 | |
| (b) | |||||
| Marker | LFD | HFD | HFKV | p Value | Primer Sequences |
| TNFα | 1.00 a | 0.81 (0.12) a | 0.21 (0.03) c | <0.05 | 5′ TACTGAACTTCGGGGTGATTGGTCC 3′ 5′ CAGCCTTGTCCCTTGAAGAGAACC 3′ |
| MCP-1 | 1.00 a | 14.9 (1.20) b | 3.23 (0.87) c | <0.05 | 5′ CATCCACGTGTTGGCTCA 3′ 5′ AACTACAGCTTCTTTGGGACA 3′ |
| CD11c | 1.00 a | 2.51(0.30) b | 0.14 (0.02) c | 0.001 | 5′ CTGGATAGCCTTTCTTCTGCTG 3′ 5′ GCACACTGTGTCCGAACTC 3′ |
| F4/80 | 1.00 a | 1.85 (0.03) b | 0.12(0.02) c | 0.001 | 5′ GGAAGGAAATGGAGAGAAAG 3′ 5′ GAAGATCTACCCTGGTGAAT 3′ |
| IL-6 | 1.00 a | 2.84 (0.05) b | 0.27 (0.07) b | 0.001 | 5′ AGCCAGAGTCCTTCAGAGA 3′ 5′ TCCTTAGCCACTCCTTCTGT 3′ |
| IL-10 | 1.00 a | 7.93 (0.04) b | 0.55 (0.04) c | 0.001 | 5′ GTCATCGATTTCTCCCCTGT 3′ 5′ ATGGCCTTGTAGACACCTT 3′ |
| IL-8 | nd | nd | nd | - | 5′ ACAGAAAGGAAGTGATAGCAGT 3′ 5′ GCTTCATTGCCGGTGGAA3′ |
| IL-1β | 1.00 a | 0.12 (0.03) b | 0.16 (0.04) b | <0.05 | 5′ CAACTGTTCCTGAACTCAACT 3′ 5′ ATCTTTTGGGGTCCGTCCAACT 3′ |
| TLR4 | 1.00 a | 0.306 (0.02) b | 1.29 (0.02) c | <0.05 | 5′ GACCGCAACAACGCCATCTA3′ 5′ GGCGTATCAGTGGGGGTCAG3′ |
| NF-κB | 1.00 a | 1.709 (0.02) b | 1.89 (0.03) b | <0.05 | 5′ GCTGAGTCCTGCTCCTTCTAAA 3′ 5′ CCTCTGTGTAGCCCATCTGTTGC 3′ |
| β-Actin | 5′ CAGCTGAGAGGGAAATCGTG 3′ 5′ CGTTGCCAATAGTGATGACC 3′ | ||||
| LFD | HFD | HFKV | p-Value | |
|---|---|---|---|---|
| Weight of full cecum (g) | 0.35 (0.02) | 0.311(0.03) | 0.381 (0.03) | ns |
| Weight of empty cecum (g) | 0.091 (0.03) | 0.086 (0.002) | 0.099 (0.003) | ns |
| pH of colon fecal samples | 8.96 (0.14) | 8.65 (0.02) | 8.11 (0.15) | ns |
| Taxa | LFD | HFD | HFKV | p-Value |
|---|---|---|---|---|
| Firmicutes | ||||
| P-Firmicutes | 57.3 a | 73.5 b | 63.8 c | 0.04 |
| P-Firmicutes; C-Bacillus | 17.9 a | 34.7 b | 19.4 a | <0.005 |
| P-Firmicutes; C-Bacillus; O-Bacillales | 3.6 a | 2.7 b | 2.4 b | ns |
| P-Firmicutes; C-Bacillus; O-Lactobacillales | 14.3 a | 31.9 b | 17.0 a | <0.005 |
| P-Firmicutes; C-Bacillus; O-Lactobacillales; F-Paenibacillaceae | 3.5 | 3.2 | 2.7 | ns |
| P-Firmicutes; C-Bacillus; O-Lactobacillales; F-Lactobacilaceae | 4.49 a | 14.3 b | 9.9 c | <0.001 |
| P-Firmicutes; C-Bacillus; O-Lactobacillales; F-Lactobacilaceae; G-Acetatifactor | 0.31 | 0.23 | 0.70 | ns |
| P-Firmicutes; C-Bacillus; O-Lactobacillales; F-Lactobacilaceae; G-Dorea | 0.06 | 0.06 | 0.02 | ns |
| P-Firmicutes; C-Bacillus; O-Lactobacillales; F-Lactobacilaceae; G-Blautia | 0.11 a | 0.09 a | 0.28 b | <0.05 |
| P-Firmicutes; C-Bacillus; O-Lactobacillales; F-Lactobacilaceae; G-Roseburia | 0.04 a | 0.05 a | 0.48 b | <0.05 |
| P-Firmicutes; C-Bacillus; O-Lactobacillales; F-Lactobacilaceae; G-Tyzerella | 0.03 a | 0.05 | 0.0 a | ns |
| P-Firmicutes; C-Bacillus; O-Lactobacillales; F-Lactobacilaceae; G-Lactobacillus | 0.48 a | 7.6 b | 3.3 c | 0.001 |
| P-Firmicutes; C-Bacillus; O-Lactobacillales; F-Lactobacilaceae; G-Lactobacillus; S-L. johnsonii | 0.24 a | 3.06 b | 1.04 c | <0.05 |
| P-Firmicutes; C-Bacillus; O-Lactobacillales; F-Streptococaceae | 13.8 a | 24.0 b | 14.6 a | 0.04 |
| P-Firmicutes; C-Bacillus; O-Lactobacillales; F-Streptococaceae; G-Lactococcus | 6.72 a | 14.4 b | 7.85 a | 0.04 |
| P-Firmicutes; C-Bacillus; O-Lactobacillales; F-Streptococaceae; G-Lactococcus; S-L. lactis | 6.19 a | 8.41 b | 5.91 a | <0.05 |
| P-Firmicutes; C-Erysipelotrichia | 18.8 a | 18.3 a | 15.1 b | <0.05 |
| P-Firmicutes; C-Erysipelotrichia; O-Erysipelotrichales | 18.7 a | 17.5 a | 14.9 b | ns |
| P-Firmicutes; C-Erysipelotrichia; O-Erysipelotrichales; F-Erysipelotrichaceae | 18.7 a | 10.3 b | 13.1 b | ns |
| P-Firmicutes; C-Erysipelotrichia; O-Erysipelotrichales; F-Erysipelotrichaceae; G-Clostridium | 0.07 | 0.13 | 0.08 | ns |
| P-Firmicutes; C-Erysipelotrichia; O-Erysipelotrichales; F-Erysipelotrichaceae; G-Turicibacter | 1.98 | 2.71 | 2.55 | ns |
| P-Firmicutes; C-Clostridia | 20.7 | 21.4 | 24.1 | ns |
| P-Firmicutes; C-Clostridia; O-Clostridiales | 20.7 | 21.4 | 24.3 | ns |
| P-Firmicutes; C-Clostridia; O-Clostridiales; F-Eubacteriaceae | 0.20 | 0.22 | 0.26 | ns |
| P-Firmicutes; C-Clostridia; O-Clostridiales; F-Eubacteriaceae; G-Eubactrium | 0.01 a | 0.02 a | 0.39 b | 0.04 |
| P-Firmicutes; C-Clostridia; O-Clostridiales; F-Lachnospiraceae | 9.5 a | 11.3 b | 13.2 b | 0.04 |
| P-Firmicutes; C-Clostridia; O-Clostridiales; F-Peptostreptococcaceae | 0.56 a | 1.67 b | 1.75 b | 0.04 |
| P-Firmicutes; C-Clostridia; O-Clostridiales; F-Oscillospiraceae | 0.35 | 0.22 | 0.10 | ns |
| P-Firmicutes; C-Clostridia; O-Clostridiales; F-Ruminococaceae | 3.74 | 4.70 | 5.64 | ns |
| P-Firmicutes; C-Clostridia; O-Clostridiales; F-Ruminococaceae; G-Ruminococcus | 3.43 | 4.33 | 5.06 | ns |
| P-Firmicutes; C-Clostridia; O-Clostridiales; F-Ruminococaceae; G-Ruminococcus; S-R. gnavus | 2.60 | 3.29 | 3.55 | ns |
| P-Firmicutes; C-Clostridia; O-Clostridiales; F-Ruminococaceae; G-Faecalibacterium | 0.31 | 0.37 | 0.64 | ns |
| P-Firmicutes; C-Clostridia; O-Clostridiales; F-Ruminococaceae; G-Faecalibacterium; S-F. prausnitzii | 0.07 | 0.09 | 0.03 | ns |
| P-Firmicutes; C-Clostridia; O-Clostridiales; F-Clostridiaceae | 7.66 | 6.17 | 8.44 | ns |
| P-Firmicutes; C-Clostridia; O-Clostridiales; F-Clostridiaceae; G-Clostridium | 7.51 | 5.94 | 7.81 | ns |
| P-Firmicutes; C-Clostridia; O-Clostridiales; F-Clostridiaceae; G-Clostridium; S-C. vincentii | 3.01 | 2.08 | 2.38 | ns |
| P-Firmicutes; C-Clostridia; O-Clostridiales; F-Clostridiaceae; G-Clostridium; S-C. disporicum | 0.52 | 0.52 | 0.35 | ns |
| P-Firmicutes; C-Clostridia; O-Clostridiales; F-Clostridiaceae; G-Clostridium; S-C. scindens | 0.17 | 0.15 | 0.08 | ns |
| P-Firmicutes; C-Clostridia; O-Clostridiales; F-Clostridiaceae; G-Clostridium; S-C. hiranonis | 0.10 | 0.34 | 0.42 | ns |
| Bacteroidetes | ||||
| P-Bacteroidetes | 24.2 a | 14.4 b | 24.5 a | 0.01 |
| P-Bacteroidetes; C-Cytophagia | 1.02 a | 0.31 b | 0.35 b | <0.05 |
| P-Bacteroidetes; C-Sphingobacteria | 0.03 | 0.03 | 0.04 | ns |
| P-Bacteroidetes; C-Flavobacteria | 1.62 a | 2.93 b | 0.93 c | <0.05 |
| P-Bacteroidetes; C-Flavobacteria; O-Flavobacteriales | 1.16 | 0.83 | 0.87 | ns |
| P-Bacteroidetes; C-Flavobacteria; O-Flavobacteriales; F-Flavobacteraceae | 1.6 a | 2.32 b | 0.52 c | <0.05 |
| P-Bacteroidetes; C-Bacteroidia | 20.7 a | 18.7 b | 23.1 a | 0.01 |
| P-Bacteroidetes; C-Bacteroidia; O-Bacteroidales | 20.7 a | 17.6 b | 23.1 a | <0.05 |
| P-Bacteroidetes; C-Bacteroidia; O-Bacteroidales; F-Bacteroidaceae | 13.9 a | 12.91 a | 15.9 b | <0.05 |
| P-Bacteroidetes; C-Bacteroidia; O-Bacteroidales; F-Bacteroidaceae; G-Bacteroides | 7.53 a | 6.88 b | 9.76 b | 0.01 |
| P-Bacteroidetes; C-Bacteroidia; O-Bacteroidales; F-Bacteroidaceae; G-Bacteroides; S-B. thetamicron | 6.48 a | 5.69 b | 8.86 c | <0.01 |
| P-Bacteroidetes; C-Bacteroidia; O-Bacteroidales; F-Porphyromonadaceae | 3.84 | 4.16 | 3.32 | ns |
| P-Bacteroidetes; C-Bacteroidia; O-Bacteroidales; F-Prevotallaceae | 2.39a | 1.57b | 2.91a | <0.05 |
| P-Bacteroidetes; C-Bacteroidia; O-Bacteroidales; F-Rikenellaceae | 0.64 | 1.24 | 0.97 | ns |
| P-Bacteroidetes; C-Bacteroidia; O-Bacteroidales; F-Rikenellaceae; G-Alistipes | 0.44 a | 0.51 a | 0.83 b | 0.01 |
| Actinobacteria | ||||
| P-Actinobacteria | 21.2 a | 8.9 b | 11.2 b | 0.001 |
| P-Actinobacteria; C-Actinobacteria | 20.0 a | 8.6 b | 10.8 b | 0.001 |
| P-Actinobacteria; C-Actinobacteria; O-Actinomycetales | 0.07 | 0.03 | 0.05 | ns |
| P-Actinobacteria; C-Actinobacteria; O-Bifidobacteriales | 18.5 a | 7.72 b | 7.43 b | 0.001 |
| P-Actinobacteria; C-Actinobacteria; O-Bifidobacteriales; F-Bifidobacteroidaceae | 18.6 a | 7.72 b | 7.43 b | 0.001 |
| P-Actinobacteria; C-Actinobacteria; O-Bifidobacteriales; F-Bifidobacteroidaceae; G-Bifidobacterium | 17.8 a | 7.34 b | 7.09 b | 0.001 |
| P-Actinobacteria; C-Actinobacteria; O-Bifidobacteriales; F-Bifidobacteroidaceae; G-Bifidobacterium; S-B. pseudolongum | 12.6 a | 5.49 b | 5.73 b | 0.001 |
| P-Actinobacteria; C-Actinobacteria; O-Bifidobacteriales; F-Bifidobacteroidaceae; G-Bifidobacterium; S-B. choerinum | 0.41 a | 0.15 b | 0.10 b | 0.001 |
| P-Actinobacteria; C-Coriobacteriia | 1.34 a | 0.89 a | 3.40 c | <0.001 |
| P-Actinobacteria; C-Coriobacteriia; O-Coriobacteriales | 1.39 a | 0.87 a | 3.38 c | <0.001 |
| P-Actinobacteria; C-Coriobacteriia; O-Coriobacteriales; F-Coriobacteriaceae | 1.39 a | 0.87 a | 3.38 c | <0.001 |
| P-Actinobacteria; C-Coriobacteriia; O-Coriobacteriales; F-Coriobacteriaceae; G-Atopobium | 0.67 a | 0.26 a | 1.26 c | <0.05 |
| P-Actinobacteria; C-Coriobacteriia; O-Coriobacteriales; F-Coriobacteriaceae; G-Enterorhabdus | 0.04 a | 0.03 a | 0.25 b | 0.05 |
| P-Actinobacteria; C-Coriobacteriia; O-Coriobacteriales; F-Coriobacteriaceae; G-Enterorhabdus; S-E. caecimuris | 0.04 a | 0.02 a | 0.19 b | 0.05 |
| Proteobacteria | ||||
| P-Proteobacteria | 0.68 | 0.22 | 0.31 | ns |
| P-Proteobacteria; C-Betaproteobacteria | 0.58 | 0.16 | 0.17 | ns |
| P-Proteobacteria; C-Betaproteobacteria; O-Bukholderiales | 0.56 | 0.16 | 0.13 | ns |
| P-Proteobacteria; C-Betaproteobacteria; O-Bukholderiales; F-Sutterellaceae | 0.56 | 0.13 | 0.16 | ns |
| P-Proteobacteria; C-Betaproteobacteria; O-Bukholderiales; F-Sutterellaceae; G-Parasutterella | 0.55 | 0.12 | 0.14 | ns |
| P-Proteobacteria; C-Betaproteobacteria; O-Bukholderiales; F-Sutterellaceae; G-Parasutterella; S-P. excrementihominis | 0.45 | 0.11 | 0.12 | ns |
| P-Proteobacteria; C-Betaproteobacteria; O-Neisseriales | 0.05 | 0.1 | 0.1 | ns |
| P-Proteobacteria; C-Betaproteobacteria; O-Neisseriales; F-Neisseriaceae | 0.05 | 0.1 | 0.1 | ns |
| P-Proteobactetria; C-Gammaproteobacteria | 0.02 a | 1.63 b | 0.73 c | 0.04 |
| P-Proteobactetria; C-Gammaproteobacteria; O-Enterobacterales | 0.02 a | 1.63 b | 0.69 b | 0.04 |
| P-Proteobactetria; C-Gammaproteobacteria; O-Enterobacterales; F-Enterobacteriaceae | 0.02 a | 1.63 b | 0.69 b | 0.04 |
| Verrucomicrobia | ||||
| P-Verrucomicrobia | 0.06 a | 0.24 b | 0.22 b | <0.05 |
| P-Verrucomicrobia; G-Akkermansia | 0.04 a | 0.22 b | 0.21 b | <0.05 |
| P-Verrucomicrobia; G-Akkermansia; S-A. municiphila | 0.04 a | 0.22 b | 0.21 b | <0.05 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Shahinozzaman, M.; Raychaudhuri, S.; Fan, S.; Obanda, D.N. Kale Attenuates Inflammation and Modulates Gut Microbial Composition and Function in C57BL/6J Mice with Diet-Induced Obesity. Microorganisms 2021, 9, 238. https://doi.org/10.3390/microorganisms9020238
Shahinozzaman M, Raychaudhuri S, Fan S, Obanda DN. Kale Attenuates Inflammation and Modulates Gut Microbial Composition and Function in C57BL/6J Mice with Diet-Induced Obesity. Microorganisms. 2021; 9(2):238. https://doi.org/10.3390/microorganisms9020238
Chicago/Turabian StyleShahinozzaman, Md, Samnhita Raychaudhuri, Si Fan, and Diana N. Obanda. 2021. "Kale Attenuates Inflammation and Modulates Gut Microbial Composition and Function in C57BL/6J Mice with Diet-Induced Obesity" Microorganisms 9, no. 2: 238. https://doi.org/10.3390/microorganisms9020238